ra hmeae httertaamedm - universiti putra malaysia papers/jtas vol. 40 (2) may... · ra hmeae...
TRANSCRIPT

Pertanika J. Trop. Agric. Sci. 40 (2): 305 - 318 (2017)
ISSN: 1511-3701 © Universiti Putra Malaysia Press
TROPICAL AGRICULTURAL SCIENCEJournal homepage: http://www.pertanika.upm.edu.my/
Article history:Received: 27 September 2016Accepted: 10 November 2016
ARTICLE INFO
E-mail addresses: [email protected] (Kamarul Rahim Kamarudin),[email protected] (Maryam Mohamed Rehan),[email protected] (Hanina Mohd Noor),[email protected] (Nur Zazarina Ramly),[email protected] (Aisyah Mohamed Rehan)* Corresponding author
Comparisan of Ossicle Shape and 12S rRNA Gene Sequencing Techniques for Species Identification of Gamat-based beche-de-mer from Langkawi Island, Kedah
Kamarul Rahim Kamarudin1*, Maryam Mohamed Rehan1, Hanina Mohd Noor1, Nur Zazarina Ramly1 and Aisyah Mohamed Rehan2 1Food Biotechnology, Faculty of Science and Technology, Universiti Sains Islam Malaysia, Bandar Baru Nilai, 71800 USIM, Nilai, Negeri Sembilan, Malaysia2Department of Biotechnology, Kulliyyah of Science, International Islamic University Malaysia, 25200 IIUM, Kuantan, Pahang, Malaysia
ABSTRACT
Due to the issues of species substitution and product mislabelling of beche-de-mer worldwide, this study aimed to identify the species of seven gamat-based beche-de-mer specimens from Kuah, Langkawi Island, Kedah, Malaysia based on ossicle shapes and non-protein-coding 12S mitochondrial rRNA gene sequences. In general, ossicles were well extracted from the specimens. At least eight ossicle shapes were observed i.e. large I-shaped rod, perforated plate, table, C-shaped rod, button, rosette, I-shaped rod and X-shaped rod. Except for button, I-shaped rod and X-shaped rod, the other five ossicle shapes are common in Stichopus horrens. However, the species status could not be resolved at this level due to the presence of uncommon ossicle shapes and the physical features of the specimens could not be used as supporting data as they were different from the live or unprocessed sea cucumber. In this study, 12S mitochondrial rRNA gene sequences were analysed using the Basic Local Alignment Search Tool programme for Nucleotides (blastn), resulting in the species identification of the beche-de-mer specimens as S. horrens,
known locally as gamat emas (golden sea cucumber) with 96-99% similarity (an average of 98%). The phylogenetic trees based on the Neighbour-Joining method, Maximum Parsimony method and Maximum Likelihood method indicated that all 12S mitochondrial rRNA gene sequences of the beche-de-mer specimens clustered with the reference samples of S.

Kamarul Rahim Kamarudin, Maryam Mohamed Rehan, Hanina Mohd Noor, Nur Zazarina Ramly and Aisyah Mohamed Rehan
306 Pertanika J. Trop. Agric. Sci. 40 (2) 306 - 318 (2017)
horrens from Pangkor Laut, Pangkor Island, Perak, Malaysia, supporting the BLASTN results and confirming their species status as S. horrens. Furthermore, 10 partial 12S mitochondrial rRNA gene sequences of the reference samples and the beche-de-mer specimens of S. horrens were registered with the GenBank (Accession No.: KX879628-KX879637). Overall, the findings suggested that the species identification of the beche-de-mer specimens using 12S mitochondrial rRNA gene sequence gave better inference than ossicle-shape identification. The outcomes of this study benefit enforcement agencies in their work of monitoring and overcoming the issues of species substitution and product mislabelling of beche-de-mer or commercial dried sea cucumber in Malaysian markets as well as in global markets.
Keywords: 12S mitochondrial rRNA gene, gamat-
based beche-de-mer, ossicle shape, phylogenetic trees,
Stichopus horrens
INTRODUCTION
Sea cucumber (Phylum Echinodermata: Class Holothuroidea) is a marine heritage of Malaysia due to its high value in traditional medicine and the trepang or beche-de-mer industry (Hashim, 2011). The marine-dwelling organism was categorised as gamat and timun laut (Kamarudin et al., 2009, 2015). The term gamat refers to all species from the family Stichopodidae e.g. Stichopus herrmanni (the curryfish) and Thelenota anax (the amberfish), while the term timun laut refers to all non-gamat
species e.g. Bohadschia vitiensis (the brown sandfish) and Holothuria (Microthele) fuscopunctata (the elephant trunkfish). In terms of species richness, Kamarudin et al. (2015) reported the presence of 10 gamat species in Malaysia including eight Stichopus species and two Thelenota species. Furthermore, Choo (2008) reported that at least six gamat species are of commercial importance in Malaysia. According to Conand et al. (2014), Thelenota ananas (the prickly redfish) and S. herrmanni – both gamat species were recorded in Malaysia – were included in the International Union for Conservation of Nature (IUCN) Red List for aspidochirotid holothuroids as endangered or at risk of extinction and vulnerable or at risk of extinction, respectively.
In Sabah, Malaysia, timun laut and gamat species have been exploited as food in the beche-de-mer industries. The term beche-de-mer refers to sea cucumber that is cooked and dried for commercial purposes (Purcell, 2014). An estimated 139 tonnes of sea cucumber was landed in Sabah from the year 2000 until 2005 (Annual Fisheries Statistics, Sabah, 2000-2005). In contrast to timun laut, the gamat species is popular as the main ingredient in gamat-based traditional products in Peninsular Malaysia e.g. body fluid extracts (air gamat) and lipid extracts (minyak gamat). Gamat is usually cooked and dried into beche-de-mer, locally known as ibu gamat, before being used as the main ingredient in traditional products. Ibu gamat is sold in Malaysian markets. Langkawi Island, Kedah and Pangkor Island, Perak are the two main production

Species Identification of beche-de-mer from Langkawi Island
307Pertanika J. Trop. Agric. Sci. 40 (2): 307 - 318 (2017)
incorrectly labelled. In Malaysian markets, Kamarudin et al. (2015a, 2015b) reported the presence of unlabelled beche-de-mer products that could be related to the issue of intentional species substitution. In fact, the global issues have been affecting the trading of processed sea cucumber. In order to address the issues, rapid, reproducible and reliable techniques for identifying the animal origin of biological specimens have been discovered and developed since 1992 (Bartlett & Davidson, 1992; Wen et al., 2011).
Therefore, the aim of this study was to identify the species of gamat-based beche-de-mer specimens from Kuah, Langkawi Island, Kedah, Malaysia based on ossicle shapes and non-protein-coding 12S mitochondrial rRNA gene sequences. Toral-Granda (2005) has suggested that ossicle, the small part of calcified material from the sea cucumber’s body, is informative and useful in identifying the species of sea cucumber in any form e.g. fresh, salted and dried forms. In fact, the ossicle continued to be an important characteristic for morphological identification of sea cucumber species (Kamarudin & Mohamed Rehan, 2015). However, the absence of common ossicle shapes and the presence of uncommon ossicle shapes will cause uncertainty in species status. For that reason, besides the ossicle shape identification technique, 12S mitochondrial rRNA gene sequencing technique was included in this study in order to obtain a more accurate conclusion and comparison. In summary, the outcomes of this study suggest that
sites of gamat-based traditional products in Peninsular Malaysia (Kamarudin et al., 2015). However, between the two islands, Langkawi Island, Kedah is more popular with the gamat-based traditional products. Gamat Asli Enterprise is one of the most established producers and distributors of gamat-based products in Langkawi Island, Kedah and it has been operating since 1960s (Wariman, 2002). Nowadays, in Malaysia, gamat-based products are also manufactured using modern technologies.
A number of 19 sea cucumber species are commercialised in Malaysia (Choo, 2008). Preceding beche-de-mer marketing, sea cucumber is gutted, boiled, roasted and finally preserved through drying, smoking or freezing. Pickling and canning are other options that may be used in the extensive processing. Processing usually leads to deformation of the body of the sea cucumber and subsequently, causes difficulties in species identification and confirmation of the processed sea cucumber or beche-de-mer in markets through their morphology. There have been cases where sea cucumber-based products were not labelled with their species names and other details such as the name, address and contact information of the manufacturer as well as quantity information. Rasmussen and Morrissey (2008) mentioned that the presence of intentional species substitution and mislabelling of sea cucumber products have been reported worldwide. Wen et al. (2011) reported that seven samples of commercial sea cucumber products from Guangzhou, China (63.6%) were

Kamarul Rahim Kamarudin, Maryam Mohamed Rehan, Hanina Mohd Noor, Nur Zazarina Ramly and Aisyah Mohamed Rehan
308 Pertanika J. Trop. Agric. Sci. 40 (2) 308 - 318 (2017)
the species identification of the beche-de-mer specimens using 12S mitochondrial rRNA gene sequence gave better inference as compared to identification based on ossicle shape. Despite that, both techniques are recommended to be used together for accurate species identification and confirmation of processed sea cucumber, in this case the commercial gamat-based beche-de-mer specimens from Kuah, Langkawi Island, Kedah, Malaysia. National enforcement agencies around the world are recommended to use both techniques for monitoring and overcoming the issues of species substitution and product mislabelling of beche-de-mer in global markets.
MATERIALS AND METHODS
Study site and sampling
Seven specimens of gamat-based beche-de-mer (i.e. in dried form) or ibu gamat distributed by Gamat Asli Enterprise were bought in Kuah, Langkawi Island, Kedah, Malaysia [Global Positioning System (GPS) position: 4° 21’ 32.53”N, 100° 46’ 24.91”E)]. The specimens were labelled LKIG 1 to LKIG 7 (Figure 1) and stored in a -20°C chest freezer for long-term storage with proper cataloguing at the Faculty of Science and Technology (FST), Universiti Sains Islam Malaysia (USIM), Nilai, Negeri Sembilan.
Ossicle extraction and shape observation
The methods outlined by Kamarudin and Mohamed Rehan (2015) were used with
little modification. A small piece of tissue (i.e. 20 mg) from each beche-de-mer specimen was cut with a sterile blade and then placed on a glass microscope slide. Several drops of liquid household bleach were applied onto the tissue portion to dissolve away the soft tissue. The mixture was left at room temperature for 30 min until a white pellet of ossicles was formed in the liquid solution. The ossicle shapes were observed at 400x magnification using Olympus culture microscope model CKX41. During the observation, the images of the ossicle shapes were captured and recorded for morphological identification. The main focus in this study was to identify the shapes of ossicles; therefore, the definite microscopic size of each ossicle type was not entirely counted (Figure 2).
Total genomic DNA extraction
A 200 mg tissue from each beche-de-mer specimen was disrupted and homogenised using the QIAGEN TissueRuptor prior to
Figure 1. Specimens of gamat-based beche-de-mer from Kuah, Langkawi, Kedah, Malaysia supplied by Gamat Asli Enterprise
15
Figure 1. Specimens of gamat-based beche-de-mer from Kuah, Langkawi, Kedah, Malaysia supplied by Gamat Asli Enterprise.

Species Identification of beche-de-mer from Langkawi Island
309Pertanika J. Trop. Agric. Sci. 40 (2): 309 - 318 (2017)
the total genomic DNA extraction using the DNeasy mericon Food Kit by QIAGEN. The protocol was modified in order to obtain a better yield of total genomic DNA. The approximate yield of the total genomic DNA was determined using 1% agarose gel with FloroSafe DNA Stain through horizontal gel electrophoresis. The total genomic DNA extracts were stored in a -20°C chest freezer for long-term storage.
Polymerase chain reaction (PCR)
Non-protein-coding 12S mitochondrial rRNA gene was amplified through standard PCR procedures using the 2x TopTaq Master Mix Kit by QIAGEN [~360 bp of fragment length based on Palumbi et al. (1991)].
AB12SA-Lf (forward) (25 bases) 5’- AAA CTG GGA TTA GAT
ACC CCA CTA T -3’
AB12SB-Hr (reverse) (20 bases) 5’- GAG GGT GAC GGG CGG
TGT GT -3’
The PCR run was programmed for 35 cycles. Cycle parameters for the PCR run were set for 2 min at 95°C for initial denaturation, 30 s at 95°C for denaturation, 30 s at 50.3°C for annealing, 45 s at 72°C for extension, repetition of step 2-4 for another 34 cycles and 5 min at 72°C for final extension.
PCR product purification and DNA sequencing
The PCR products were purified using the QIAquick PCR Purification Kit by QIAGEN (for direct purification of single PCR fragment) and QIAquick Gel Extraction Kit by QIAGEN (for purification of desired PCR fragment from agarose gel). The purified PCR products were sent for DNA sequencing at the First BASE Laboratories Sdn Bhd, Seri Kembangan, Selangor, Malaysia.
Phylogenetic analyses and GenBank submission
Sequenced 12S mitochondrial rRNA gene PCR products were displayed using Chromas programme version 2.5.1 (Copyright© 1998-2016 Technelysium Pty Ltd). Online Basic Local Alignment Search Tool programme for Nucleotides (blastn) was then accessed to assign each DNA sequence obtained from this study to a particular sea cucumber species or genus. For the multiple sequence alignment of forward reaction sequences, ClustalX programme version 2.1 (Thompson et al., 1997) was used prior to the phylogenetic tree reconstruction.
Figure 2. Ossicle shapes in the gamat-based beche-de-mer specimens from Kuah, Langkawi, Kedah, Malaysia. (a) Large I-shaped rods, (b) tables, (c) C-shaped rods, (d) perforated plates, (e) rosettes, (f) small rods, (g) buttons, and (h) X-shaped rod
16
Figure 2. Ossicle shapes in the gamat-based beche-de-mer specimens from Kuah, Langkawi, Kedah, Malaysia. (a) Large I-shaped rods, (b) tables, (c) C-shaped rods, (d) perforated plates, (e) rosettes, (f) small rods, (g) buttons, and (h) X-shaped rod.

Kamarul Rahim Kamarudin, Maryam Mohamed Rehan, Hanina Mohd Noor, Nur Zazarina Ramly and Aisyah Mohamed Rehan
310 Pertanika J. Trop. Agric. Sci. 40 (2) 310 - 318 (2017)
The reconstruction of a Neighbour-Joining (NJ) tree, Maximum Parsimony (MP) tree and Maximum Likelihood (ML) tree were computed using Molecular Evolutionary Genetics Analysis 6 programme (MEGA6) (Tamura et al., 2013). In addition, a 12S mitochondrial rRNA gene sequence of Holothuria (Mertensiothuria) leucospilota from Teluk Nipah, Pangkor Island, Perak (HL1, GenBank Accession No.: KX768273) was included as the outgroup while three 12S mitochondrial rRNA gene sequence of morphospecies Stichopus horrens from Pangkor Laut, Pangkor Island, Perak (SHP1-SHP3) were incorporated as the commercial species standard or reference samples. In contrast to the specimens of beche-de-mer from Kuah, Langkawi, Kedah, the total genomic DNA of the reference samples was extracted from their fresh tissues i.e. unprocessed samples. Regarding the GenBank submission, Sequin version 15.10 programme was used to prepare sequence data in the submission entry in order to obtain the accession numbers from the GenBank, National Center for
Biotechnology Information (NCBI), U. S. National Library of Medicine.
RESULTS AND DISCUSSION
The ossicles from the commercial beche-de-mer specimens from Kuah, Langkawi Island, Kedah, Malaysia were successfully extracted even though the specimens underwent extensive processing including gutting, boiling, roasting and subsequent preservation procedures for storage prior to marketing and consumption. Despite the fact that the processes caused body deformation and this led to difficulties for clear species identification and confirmation on the basis of morphology, the ossicles were still observable. At least eight ossicle shapes were microscopically observed without the size measurement i.e. large I-shaped rod, perforated plate, table, C-shaped rod, button, rosette, I-shaped rod and X-shaped rod (Figure 2, Table 1). According to Selenka (1867), the large I-shaped rod, perforated plate, table, C-shaped rod and rosette (Figure 2(a)-2(e)) were common ossicle shapes in S. horrens. All the common ossicles were
Table 1 List of ossicle shapes in the gamat-based beche-de-mer specimens from Kuah, Langkawi, Kedah
Specimen Large I-shaped rod
Table C-shaped rod
Perforated plate
Rosette I-shaped rod
Button X-shaped rod
LKIG 1 √ √ √ √ x x √ xLKIG 2 √ √ √ x √ √ x xLKIG 3 √ √ √ √ √ √ √ xLKIG 4 √ √ x √ x √ x xLKIG 5 √ √ √ √ √ x √ xLKIG 6 √ √ √ √ √ √ x √LKIG 7 √ √ √ x √ √ √ √

Species Identification of beche-de-mer from Langkawi Island
311Pertanika J. Trop. Agric. Sci. 40 (2): 311 - 318 (2017)
present in LKIG 3, LKIG 5 and LKIG 6 while the other four specimens showed the absence of one or two common ossicle shape(s) (Table 1). The absence could have been due to the effects from the beche-de-mer processing methods. Besides that, some ossicle shapes could be overlooked during the microscopic observation.
Interestingly, three ossicle shapes i.e. button, I-shaped rod and X-shaped rod were also recorded in this study (Figure 2(f)-2(h)). The button shape was observed in LKIG 1, LKIG 3, LKIG 5 and LKIG 7; the I-shaped rod in LKIG 2, LKIG 3, LKIG 4, LKIG 6 and LKIG 7; and the X-shaped rod in LKIG 6 and LKIG 7. Kamarudin and Mohamed Rehan (2015) also recorded the presence of the I-shaped rod in the tentacles and respiratory trees of S. horrens specimens from Pangkor Island, Perak, Malaysia as well as the X-shaped rod in the tentacles and gastrointestines of the species. However, the button shape was not listed. As a result, the species status of the commercial beche-de-mer specimens was hard to be confirmed at this level due to the absence of the common ossicle shapes and the presence of the uncommon ossicle shapes. It was also difficult to identify the specimens due to their deformed physical appearance that was different from the live or unprocessed sea cucumbers.
The blastn results for the non-protein-coding 12S mitochondrial rRNA gene sequences of the beche-de-mer specimens demonstrated that the specimens were identified as S. horrens with 96-99%
similarity i.e. an average of 98% similarity. All the partial 12S mitochondrial rRNA gene sequences of the beche-de-mer specimens of S. horrens were registered with the GenBank (Accession No.: KX879630 - KX879637). In Peninsular Malaysia, S. horrens or the dragonfish is well known locally as gamat emas or the golden sea cucumber (Kamarudin et al., 2009; 2015). In terms of the multiple sequence alignment of the forward reaction sequences against the outgroup using the ClustalX programme version 2.1 (Figure 3), the results showed that 21 out of 356 base positions were found to contain variable bases within the sequences of the beche-de-mer specimens. Eighteen base positions presented transversion, two base positions showed transition and one base position demonstrated an insertion. Interestingly, there were 13 variable bases in the sequence of LKIG7 specimen consisting of one transition (i.e. adenine to thymine), one insertion of adenine and 11 transversions. Despite that, the blastn result identified it as S. horrens with 96% similarity.
Regarding the phylogenetic analyses, the tree reconstruction based on the distanced-based NJ method with clustering algorithm, along with the character-based MP method and the character-based ML method both with optimality criterion, involved 11 nucleotide sequences including 10 in-groups and an out-group to root each tree. All positions containing gaps and missing data were eliminated. As a result, there were a total of 346 positions in the final dataset. The optimal NJ tree with

Kamarul Rahim Kamarudin, Maryam Mohamed Rehan, Hanina Mohd Noor, Nur Zazarina Ramly and Aisyah Mohamed Rehan
312 Pertanika J. Trop. Agric. Sci. 40 (2) 312 - 318 (2017)
the sum of branch length=0.38155689 is shown in Figure 4. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches (Felsenstein, 1985). The tree was drawn to scale, with branch lengths in the same units as the trees’ evolutionary distance used to infer the phylogenetic tree. Evolutionary distance was computed using the Maximum
Figure 3. Multiple sequence alignment of forward reaction sequences of gamat-based beche-de-mer specimens from Kuah, Langkawi, Kedah, Malaysia using ClustalX programme version 2.1 (Thompson et al., 1997). Twenty-one out of 356 base positions, highlighted in red numbers, were found to contain variable bases
17
Figure 3. Multiple sequence alignment of forward reaction sequences of gamat-based beche-de-mer specimens from Kuah, Langkawi, Kedah, Malaysia using ClustalX programme version 2.1 (Thompson et al., 1997). Twenty-one out of 356 base positions, highlighted in red numbers, were found to contain variable bases.
Figure 4. The phylogenetic inference of gamat-based beche-de-mer specimens from Kuah, Langkawi, Kedah, Malaysia using the Neighbour-Joining method (Saitou & Nei, 1987)
18
Figure 4. The phylogenetic inference of gamat-based beche-de-mer specimens from Kuah, Langkawi, Kedah, Malaysia using the Neighbour-Joining method (Saitou & Nei, 1987).
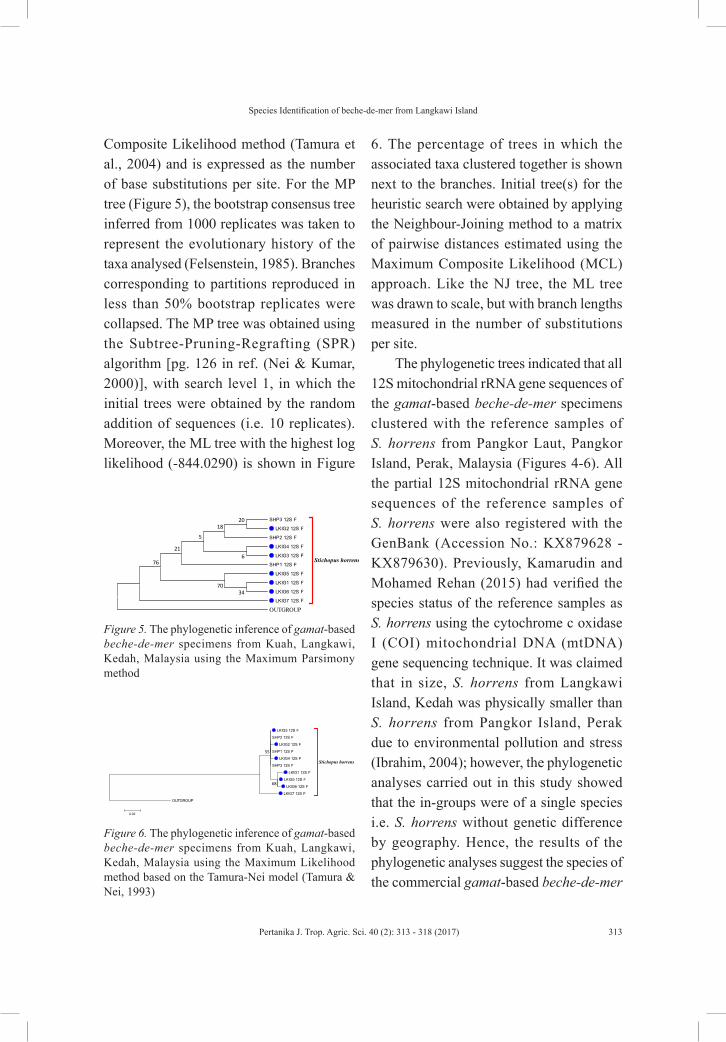
Figure 5. The phylogenetic inference of gamat-based beche-de-mer specimens from Kuah, Langkawi, Kedah, Malaysia using the Maximum Parsimony method.
LKIG4 12S F
LKIG2 12S F
SHP3 12S F
SHP1 12S F
LKIG3 12S F
SHP2 12S F
LKIG1 12S F
LKIG5 12S F
LKIG6 12S F
LKIG7 12S F
Stichopus horrens
OUTGROUP
4653
61
7315912
0.05
SHP3 12S F
LKIG2 12S F
SHP2 12S F
LKIG4 12S F
LKIG3 12S F
SHP1 12S F
LKIG5 12S F
LKIG1 12S F
LKIG6 12S F
LKIG7 12S F
Stichopus horrens
OUTGROUP
3470
76
2018
621
5
Species Identification of beche-de-mer from Langkawi Island
313Pertanika J. Trop. Agric. Sci. 40 (2): 313 - 318 (2017)
Composite Likelihood method (Tamura et al., 2004) and is expressed as the number of base substitutions per site. For the MP tree (Figure 5), the bootstrap consensus tree inferred from 1000 replicates was taken to represent the evolutionary history of the taxa analysed (Felsenstein, 1985). Branches corresponding to partitions reproduced in less than 50% bootstrap replicates were collapsed. The MP tree was obtained using the Subtree-Pruning-Regrafting (SPR) algorithm [pg. 126 in ref. (Nei & Kumar, 2000)], with search level 1, in which the initial trees were obtained by the random addition of sequences (i.e. 10 replicates). Moreover, the ML tree with the highest log likelihood (-844.0290) is shown in Figure
6. The percentage of trees in which the associated taxa clustered together is shown next to the branches. Initial tree(s) for the heuristic search were obtained by applying the Neighbour-Joining method to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach. Like the NJ tree, the ML tree was drawn to scale, but with branch lengths measured in the number of substitutions per site.
The phylogenetic trees indicated that all 12S mitochondrial rRNA gene sequences of the gamat-based beche-de-mer specimens clustered with the reference samples of S. horrens from Pangkor Laut, Pangkor Island, Perak, Malaysia (Figures 4-6). All the partial 12S mitochondrial rRNA gene sequences of the reference samples of S. horrens were also registered with the GenBank (Accession No.: KX879628 - KX879630). Previously, Kamarudin and Mohamed Rehan (2015) had verified the species status of the reference samples as S. horrens using the cytochrome c oxidase I (COI) mitochondrial DNA (mtDNA) gene sequencing technique. It was claimed that in size, S. horrens from Langkawi Island, Kedah was physically smaller than S. horrens from Pangkor Island, Perak due to environmental pollution and stress (Ibrahim, 2004); however, the phylogenetic analyses carried out in this study showed that the in-groups were of a single species i.e. S. horrens without genetic difference by geography. Hence, the results of the phylogenetic analyses suggest the species of the commercial gamat-based beche-de-mer
Figure 5. The phylogenetic inference of gamat-based beche-de-mer specimens from Kuah, Langkawi, Kedah, Malaysia using the Maximum Parsimony method
18
Figure 4. The phylogenetic inference of gamat-based beche-de-mer specimens from Kuah, Langkawi, Kedah, Malaysia using the Neighbour-Joining method (Saitou & Nei, 1987).
Figure 5. The phylogenetic inference of gamat-based beche-de-mer specimens from Kuah, Langkawi, Kedah, Malaysia using the Maximum Parsimony method.
LKIG4 12S F
LKIG2 12S F
SHP3 12S F
SHP1 12S F
LKIG3 12S F
SHP2 12S F
LKIG1 12S F
LKIG5 12S F
LKIG6 12S F
LKIG7 12S F
Stichopus horrens
OUTGROUP
4653
61
7315912
0.05
SHP3 12S F
LKIG2 12S F
SHP2 12S F
LKIG4 12S F
LKIG3 12S F
SHP1 12S F
LKIG5 12S F
LKIG1 12S F
LKIG6 12S F
LKIG7 12S F
Stichopus horrens
OUTGROUP
3470
76
2018
621
5
Figure 6. The phylogenetic inference of gamat-based beche-de-mer specimens from Kuah, Langkawi, Kedah, Malaysia using the Maximum Likelihood method based on the Tamura-Nei model (Tamura & Nei, 1993)
19
Figure 6. The phylogenetic inference of gamat-based beche-de-mer specimens from Kuah, Langkawi, Kedah, Malaysia using the Maximum Likelihood method based on the Tamura-Nei model (Tamura & Nei, 1993). Table 1 List of Ossicle Shapes in the Gamat-Based Beche-De-Mer Specimens from Kuah, Langkawi, Kedah
Specimen Large I-shaped
rod Table C-shaped
rod Perforated
plate Rosette I-shaped
rod Button X-shaped
rod LKIG 1 √ √ √ √ x x √ x LKIG 2 √ √ √ x √ √ x x LKIG 3 √ √ √ √ √ √ √ x LKIG 4 √ √ x √ x √ x x LKIG 5 √ √ √ √ √ x √ x LKIG 6 √ √ √ √ √ √ x √ LKIG 7 √ √ √ x √ √ √ √
LKIG3 12S F
SHP2 12S F
LKIG2 12S F
SHP1 12S F
LKIG4 12S F
SHP3 12S F
LKIG1 12S F
LKIG5 12S F
LKIG6 12S F
LKIG7 12S F
Stichopus horrens
OUTGROUP
68
55
0.02

Kamarul Rahim Kamarudin, Maryam Mohamed Rehan, Hanina Mohd Noor, Nur Zazarina Ramly and Aisyah Mohamed Rehan
314 Pertanika J. Trop. Agric. Sci. 40 (2) 314 - 318 (2017)
specimens from Kuah, Langkawi Island, Kedah, Malaysia to be S. horrens.
Since the gamat-based beche-de-mer specimens in this study underwent body deformation, their physical features could not be used to identify their sea cucumber species as they were not completely similar to the original morphological features. Therefore, the ossicle shape was used for species identification. However, species identification of the specimens based on ossicle shape itself could not be fully resolved. Even though there were common ossicle shapes of S. horrens in the microscopic observation, an additional approach was still required to support and then confirm species identity due to the presence of additional ossicle shapes. This study also suggested that species identification of the processed sea cucumbers using 12S mitochondrial rRNA gene sequences gave a more accurate conclusion compared to identification by ossicle shape, suggesting that 12S mitochondrial rRNA gene sequencing can be a suitable additional approach for identifying sea cucumber species. This study also discovered the potential of the 12S mitochondrial rRNA gene sequencing technique for use by enforcement agencies to resolve the issues of species substitution and product mislabelling of processed sea cucumber in Malaysia as well as throughout the world. Effective maternal inheritance, apparent haploid genome, considerable non-recombination, continuous replication and greater rate of substitution compared to the ‘single-copy’ nuclear has made
mitochondrial DNA, including the 12S mitochondrial rRNA gene, the main interest in genetic studies (Nabholz et al., 2008). By taking into account the observation of five common ossicle shapes of S. horrens in the specimens, both the non-protein-coding 12S mitochondrial rRNA gene sequencing along with the ossicle shape observation can be used to provide better resolution of species status of beche-de-mer specimens.
In general, the specimens of gamat-based beche-de-mer or ibu gamat distributed by the Gamat Asli Enterprise are S. horrens, the Langkawi gamat or gamat emas, which contain high therapeutic value, including the capabilities to hasten wound healing and rejuvenate tissues (Hashim, 2011). The company is one of the most established producers and distributors of gamat-based products in Langkawi Island (Wariman, 2002). However, sea cucumber ranching being carried out by the Langkawi Development Authority (LADA) in Teluk Yu, Temoyong, Langkawi and Tuba Island, Langkawi uses Holothuria (Metriatyla) scabra instead of S. horrens (Sharif & Osman, 2016). H. scabra is regarded as one of the timun laut species (Kamarudin et al., 2015) but some people in Malaysia also regard it as a gamat species. According to Choo (2008), S. horrens and H. scabra are among Malaysia’s commercial species of sea cucumber. However, the sandfish is regarded as “endangered, or at a high risk of extinction” based on the IUCN Red List for aspidochirotid holothuroids (Conand et al., 2014). The use of H. scabra in sea ranching

Species Identification of beche-de-mer from Langkawi Island
315Pertanika J. Trop. Agric. Sci. 40 (2): 315 - 318 (2017)
could be due to its well-studied and well-developed hatchery and culture techniques that are used worldwide.
CONCLUSION
Ossicles from the commercial beche-de-mer specimens from Kuah, Langkawi Island, Kedah, Malaysia were extracted with at least eight ossicle shapes i.e. large I-shaped rod, perforated plate, table, C-shaped rod, button, rosette, I-shaped rod and X-shaped rod. The first five shapes are common in S. horrens. Nonetheless, the absence of one or two common ossicle shape(s) and the presence of additional uncommon shapes required the use of an additional species identification approach. In terms of species status and genetic relationship based on the non-protein-coding 12S mitochondrial rRNA gene sequence, the blastn results suggested that the beche-de-mer specimens were S. horrens (i.e. gamat emas) with an average of 98% similarity. In total, 10 partial 12S mitochondrial rRNA gene sequences of S. horrens resulting from this study were registered with the GenBank, NCBI, U. S. National Library of Medicine (Accession No.: KX879628-KX879637). The phylogenetic trees of NJ, MP and ML demonstrated that all 12S mitochondrial rRNA gene sequences of the beche-de-mer specimens clustered with the reference samples of S. horrens from Pangkor Laut, Pangkor Island, Perak, Malaysia, confirming their species status as S. horrens. Besides this, the findings suggest that species identification of the beche-de-mer specimens using 12S mitochondrial
rRNA gene sequence gave better inference compared to the ossicle shape. Despite that, species identification by means of the non-protein-coding 12S mitochondrial rRNA gene sequencing along with the ossicle shape observation gave better resolution in species identification and status confirmation. For monitoring and overcoming the issues of species substitution and product mislabelling of processed sea cucumber in Malaysian markets and also in global markets, this study showed the potential of 12S mitochondrial rRNA gene sequencing along with ossicle shape observation for use by enforcement agencies for the purpose of monitoring and overcoming the issues of species substitution and product mislabelling of beche-de-mer or commercial sea cucumber in Malaysian markets as well as in global markets.
ACKNOWLEDGEMENTS
Thank you to all reviewers of this paper, all members of the Food Biotechnology programme, Faculty of Science and Technology (FST), Universiti Sains Islam Malaysia (USIM), Nilai, Negeri Sembilan Darul Khusus, especially Ms. Sarina Irma Binti Saidon, Mr. Muhammad Izzat B. Redzuan and Mrs. Siti Nabilah Bt Mohd Rusly (the laboratory assistants) and the Malaysian Ministry of Higher Education (MOHE) for valuable input and great assistance. This research was funded by the Research Acculturation Grant Scheme (RAGS) Phase 1/2014 from the Ministry of Education (MOE) – Ref: USIM/RAGS/FST/36/50414. For further details, visit

Kamarul Rahim Kamarudin, Maryam Mohamed Rehan, Hanina Mohd Noor, Nur Zazarina Ramly and Aisyah Mohamed Rehan
316 Pertanika J. Trop. Agric. Sci. 40 (2) 316 - 318 (2017)
the Sea Cucumber (Echinodermata: Holothuroidea) Database at http://sites.google.com/site/malaysianseacucumber/.
REFERENCESAFS. (2000-2005). Annual Fisheries Statistics, Sabah.
Department of Fisheries, Kota Kinabalu, Sabah.
Bartlett, S. E., & Davidson, W. S. (1992). FINS (Forens ica l ly Informat ive Nucleot ide Sequencing): A procedure for identifying the animal origin of biological specimens. Bio Tech, 12(3), 408–11.
Choo, P. S. (2008). Population status, fisheries and trade of sea cucumbers in Asia. In V. Toral-Granda, A. Lovatelli, & M. Vasconcellos (Eds.), Sea cucumbers. A global review of fisheries and trade (pp. 81-118). Rome: Food and Agriculture Organization of the United Nations.
Conand, C., Polidoro, B., Mercier, A., Gamboa, R., Hamel, J. F., & Purcell, S. (2014). The IUCN Red List assessment of aspidochirotid sea cucumbers and its implications. SPC Beche-de-mer Information Bulletin, 34(2014 May), 3–7.
Felsenstein, J. (1985). Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 39(4), 783–791.
Hashim, R. (2011). Timun laut warisan Malaysia. Kuala Lumpur, Malaysia: Research Management Centre, International Islamic University Malaysia.
Ibrahim, J. (2004, November 22). Populasi gamat di Langkawi tinggal 30 peratus. Berita Harian. Malaysia.
Kamarudin, K. R., & Rehan, M. M. (2015). Morphological and molecular identification of Holothuria (Merthensiothuria) leucospilota and Stichopus horrens from Pangkor Island, Malaysia. Tropical Life Sciences Research, 26(1), 87–99.
Kamarudin, K. R., Rehan, M. M., Noor, H. M., Ramly, N. Z., & Rehan, A. M. (2015a). 16S rDNA barcoding technique for molecular identification of processed sea cucumbers from selected Malaysian markets. In Proceedings of the 3rd International Postgraduate Conference on Science and Mathematics 2015 (IPCSM’15) – Science and Mathematics Empower Innovative Generation (pp. 22–26). Sultan Idris Education University (UPSI), Tanjong Malim, Perak Darul Ridzuan, Malaysia.
Kamarudin, K. R., Rehan, A. M., Noor, H. M., Ramly, N. Z., & Rehan, M. M. (2015b). Molecular species identification of processed sea cucumbers from selected Malaysian markets using COI MtDNA gene. In Proceedings of International Conference on Biodiversity and Conservation 2015 (ICBC 2015) – “Sustaining Biodiversity for Sustainable Future” (pp. 5–28). Sultan Idris Education University (UPSI), Tanjong Malim, Perak Darul Ridzuan, Malaysia.
Kamarudin, K. R., Rehan, A. M., Lukman, A. L., Ahmad, H. F., Anua, M. H., Nordin, N. F. H. … Usup, G. (2009). Coral reef sea cucumbers in Malaysia. Malaysian Journal of Science, 28(2), 171–186.
Kamarudin, K. R., Usup, G., Hashim, R., & Rehan, M. M. (2015). Sea cucumber (Echinodermata: Holothuroidea) species richness at selected localities in Malaysia. Pertanika Journal of Tropical Agricultural Science, 38(1), 7–32.
Nabholz, B., Mauffrey, J. F., Bazin, E., Galtier, N., & Glemin, S. (2008). Determination of mitochondrial genetic diversity in mammals. Genetics, 178(1), 351–361.
Palumbi, S., Martin, A., Romano, S., McMillan, W. O., Stice, L., & Grabowski, G. (1991). The simple fool’s guide to PCR. Honolulu: Department of Zoology and Kewalo Marine Laboratory, Universiti of Hawaii.

Species Identification of beche-de-mer from Langkawi Island
317Pertanika J. Trop. Agric. Sci. 40 (2): 317 - 318 (2017)
Purcell, S. W. (2014). Processing sea cucumbers into beche-de-mer: A manual for Pacific Island fishers (p. 9). New South Wales: Southern Cross University and Noumea Cedex: The Secretariat of the Pacific Community.
Rasmussen, R. S., & Morrissey, M. T. (2008). DNA-based methods for the identification of commercial fish and seafood species. Comprehensive Reviews in Food Science and Food Safety, 7(3), 280–295.
Saitou, N., & Nei, M. (1987). The neighbor-joining method: A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, 4(4), 406–425.
Selenka, E. (1867). Beiträge zur Anatomis un Systematik der Holothurien. Zeitschrift für wissenschaftliche Zoologie, 17, 291–374.
Sharif, A., & Osman, H. (2016, February 16). Prospek cerah pulihara gamat. Harian Metro. Ruangan Setempat. Langkawi, Malaysia.
Tamura, K., & Nei, M. (1993). Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Molecular Biology and Evolution, 10(3), 512–526.
Tamura, K., Nei, M., & Kumar, S. (2004). Prospects for inferring very large phylogenies by using the neighbor-joining method. Proceedings of the National Academy of Sciences (USA), 101(30), 11030–11035.
Tamura, K., Stecher, G., Peterson, D., Filipski, A., & Kumar, S. (2013). MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Molecular Biology and Evolution, 30(12), 2725–2729.
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F., & Higgins, D. G. (1997). The ClustalX windows interface: Flexible strategies for multiple sequence alignment aided by the quality analysis tools. Nucleic Acids Research, 25(24), 4876–4882.
Toral-Granda, M. V. (2005). The use of calcareous spicules for the identification of the Galápagos sea cucumber Isostichopus fuscus on the international market. SPC Beche-de-mer Information Bulletin, 22(2005, July), 3–5.
Wariman, N. H. (2002, November 18). Langkawi gamat-based health products maker eyes South Korean market. Pertubuhan Berita Nasional Malaysia (BERNAMA). Langkawi, Kedah, Malaysia.
Wen, J., Hu, C., Zhang, L., & Fan, S. (2011). Genetic identification of global commercial sea cucumber species on the basis of mitochondrial DNA sequences. Food Control, 22(2011), 72–77.
