putative receptor growth auxinidentified characterizedauxin-binding...
TRANSCRIPT

Proc. Nati. Acad. Sci. USAVol. 88, pp. 5479-5483, July 1991Biochemistry
Putative receptor for the plant growth hormone auxin identifiedand characterized by anti-idiotypic antibodies
(auxin-binding proteins/soybean/2,4-dichlorophenoxyacetic acid/indole-3-acetic acid/plant growth regulators)
PARUCHURI V. PRASAD*t AND ALAN M. JONESt*Department of Molecular Biology, Scripps Clinic and Research Foundation, La Jolla, CA 92037; and tDepartment of Biology, CB# 3280, Coker Hall, TheUniversity of North Carolina at Chapel Hill, Chapel Hill, NC 27599
Communicated by Charles S. Levings III, March 7, 1991 (received for review December 17, 1990)
ABSTRACT The molecular mechanism of action of aux-ins, which are an important class of plant growth regulators,is not known. While they are thought to influence cellularprocesses such as gene expression and membrane hyperpolar-ization by binding to specific receptors, no receptor mediatingthese processes has been unambiguously identified. We de-scribe the identification of a putative auxin receptor by usinganti-idiotypic antibodies.' We have demonstrated that a 65-kilodalton protein found in a wide range of plants specificallybinds auxins. Furthermore, since it is localized to the nucleus,it is a candidate for an auxin receptor that mediates somenuclear process.
Auxins are a group ofplant hormones that influence a numberof physiological and developmental processes (1). While themolecular mechanism of action of auxins is unknown, theyare expected to interact with cellular receptors, leading to theactivation of a number of biochemical processes. Auxin-binding proteins (ABPs) have been reported in both themembrane and soluble fractions of some plants (2-6). A22-kilodalton (kDa) subunit ABP in maize membranes hasbeen purified and characterized (2), and a 40- and 42-kDaauxin-binding doublet in tomato and zucchini has been iden-tified (4, 5). While there is some indirect evidence suggestingthat the maize ABP may be involved in elongation growth ofthe coleoptile (7), the function oftomato and zucchini plasmamembrane ABPs is unclear. Since auxins accumulate in thecell (8), an intracellular site for an auxin receptor is bothplausible and consistent with its rapid effect on gene tran-scription. Auxins, exhibiting multiple effects in target cells,may act through several receptors located in different com-partments in various tissues. Therefore, it is important toassign functions to all ABPs to gain a full understanding ofauxin action. Cytosolic auxin-binding sites have been re-ported, but a characterization of the biochemical identity orthe function of these sites'has been hampered by a lack ofreliable ligand-binding assays in crude plant extracts and thelow concentrations of binding proteins (9, 10). These diffi-culties have necessitated a fresh approach to identifying andcharacterizing soluble auxin receptors. Several novel ap-proaches such as photoaffinity labeling, affinity chromatog-raphy, and anti-idiotypic antibodies have been developed forthe isolation of receptors for physiologically important lig-ands (3-5, 11-16). The anti-idiotypic antibody approach hasbeen utilized to identify receptors for several biologicaleffectors in animal systems (11, 13-15, 17, 18), although it hasnot been used to identify a receptor for a plant growthregulator. In this communication,' we report the application ofthis strategy to the identification of a ubiquitous ABP inplants.
MATERIALS AND METHODSChemicals and Plant Materials. 2,4-Dichlorophenoxy[2-
14C]acetic acid ([14C]2,4-D; specific activity 56 mCi/mmol; 1Ci =' 37 GBq), indole-3-[2-14C]acetic acid ([14C]IAA; specificactivity 56 mCi/mmol), and [5-3H]indole-3-acetic acid([3H]IAA; specific activity 27.6 Ci/mmol) were obtainedfrom Amersham. All chemicals were of the highest puritycommercially available. Soybean (Glycine max var. Wayne)seeds were obtained from Wilkens Seeds, Pontiac, IL andwere germinated in the dark in moist vermiculite. All otherseeds were obtained from the local market.Production of Antisera. Antibodies to IAA were raised by
injecting female New Zealand White (NZW) rabbits with IAAconjugated to bovine serum albumin (19, 20). The firstinjection consisted of 1 mg of conjugate emulsified in Fre-und's complete adjuvant (FCA) at multiple subcutaneoussites. The animals were boosted at 2-week intervals with 0.5mg of conjugate in incomplete Freund's adjuvant. The ani-mals were bled from the ear vein after three boosts and theantibody titer was measured by a radioimmunoassay (19, 20).The specificity of this antiserum for IAA was demonstratedby analog competition of0.5 puM [14CJIAA. It was shown that100 AM IAA inhibited the -binding of [14C]IAA by 51%,whereas 5-nitroindole, indole-3-aldehyde, 3-indoleglyoxyla-mide, indole-3-carboxylic acid, indole-2-carboxylic acid,L-tryptophan, indole-3-carbinol, each at 100 AM, did notinhibit the binding of [14C]IAA. Indole-3-acetamide, ethylin-dole-3-acetate, and indole-3-acetone inhibited a maximum of19%, indicating that the antiserum is specific for IAA and thatthe indoleacetyl group is important for antibody recognition.For the production of anti-idiotypic antibodies (17, 18),
affinity purified anti-IAA antibodies (0.3 mg) emulsified inFCA were injected into NZW rabbits as described above.Subsequent boosts were given in incomplete Freund's adju-vant with 0.2 mg of antibodies per animal. Anti-idiotypicresponse was monitored by an enzyme-linked immunosor-bent assay (ELISA) and was detected after three boosterinjections.
Purification and Characterization of Antisera. Anti-IAAantibodies for the preparation of anti-idiotypic antisera werepurified by affinity chromatography on a column of Affigel-102 (Bio-Rad) containing immobilized IAA coupled throughthe ring nitrogen via a six-atom spacer arm to the matrix (19,20). The antisera were dialyzed against phosphate-bufferedsaline (PBS, 10 mM potassium phosphate/0.15 M NaCl, pH7.4) and passed through a column containing the affinitymatrix at a flow rate of 10 ml/hr. The column was washed
Abbreviations: IAA, indole-3-acetic acid; 2,4-D, 2,4-dichlorophe-noxyacetic acid; NAA, naphthalene-1-acetic acid; 2,4,5-T, 2,4,5-trichlorophenoxyacetic acid; ABP, auxin-binding protein; FCA,Freund's complete adjuvant.tTo whom reprint requests should be addressed at present address:Department of Biology, CB# 3280, Coker Hall, The University ofNorth Carolina at Chapel Hill, Chapel Hill, NC 27599.
5479
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

5480 Biochemistry: Prasad and Jones
thoroughly in PBS and the bound antibodies were eluted by0.5 mM IAA. The eluted antibodies were dialyzed to removeIAA and injected into rabbits.For some studies, the IgG fraction purified on a staphylo-
coccal protein A column (Bio-Rad) was used. The IgGfraction was digested with immobilized papain (Pierce) andpassed over a column of protein A to isolate Fab fragments.The IgG fractions of anti-idiotypic and preimmune sera
were immobilized on Affi-Gel 10 (Bio-Rad) according to themanufacturer's instructions. The amount of IgG coupled wasestimated to be 10 mg/ml of gel. An ELISA was used toanalyze the interaction of anti-idiotypic antisera with anti-IAA Fab fragments immobilized on polystyrene plates (0.25Aug per well). The amount of anti-idiotypic antibody bound tothe immobilized Fab fragments was estimated colorimetri-cally by using protein G-alkaline phosphatase conjugate(Calbiochem).Immunoblot Analysis of Plant Extracts. Etiolated seedlings
(10 g) were homogenized with 50 mM Tris-HCl/1 mMEDTA/10 mM 2-mercaptoethanol, pH 7.6 (buffer 1) in aWaring blender. The homogenate was filtered throughcheesecloth and centrifuged at 105,000 x g for 60 min.Samples with a known protein concentration (21) were pre-pared for SDS/polyacrylamide gel electrophoresis (SDS/PAGE) (22) as follows. The supernatants after centrifugation(0.5 mg of protein per ml) were mixed with an equal volumeofphenol/chloroform (1:1) on a Vortex mixer and centrifuged(12,000 x g) for 4 min. The lower organic phase and theinterphase were extracted three more times with an equalvolume of buffer and the aqueous layer was discarded aftereach centrifugation. Finally, the proteins in the interphasewere precipitated with 5 vol of cold (-20°C) acetone andrecovered by centrifugation. This step in sample preparation,which will be described fjrther in a following report, wasfound to be necessary for efficient antibody recognition. Thepellets were washed twice in cold acetone, dried, and dis-solved in SDS-sample buffer [62.5 mM Tris HCl, pH 6.8/2%SDS/8 M urea/5% (vol/vol) 2-mercaptoethanol] by heatingat 60°C for 10 min. The proteins were resolved by SDS/12%PAGE and transferred to nitrocellulose (23). The blots wereprobed with a 1:500 dilution of anti-idiotypic antiserumfollowed by an anti-rabbit IgG-alkaline phosphatase conju-gate (Bio-Rad, 1:2000 dilution). The blots were scanned in anImagequant gel scanner (Molecular Dynamics, MountainView, CA).Sample Preparation for Auxin-Binding Assays. All centrif-
ugations were for 20 min at 25,000 x g and all samplemanipulations were conducted at 4°C. Seven-day old soy-bean seedlings were homogenized in buffer 1 and the homog-enate was filtered. After centrifugation, 10 mM MnSO4 wasadded to the supernatant and stirred for 15 min. The precip-itate was removed by centrifugation, and solid ammoniumsulfate was added to the supernatant to give 60% saturation.After stirring and centrifugation, the pellet was discarded andthe concentration of ammonium sulfate in the supernatantwas raised to 80%6. The pellet obtained after centrifugationwas dissolved in buffer and desalted on a Bio-Gel P-6 column(Bio-Rad).Auxin-Binding Assays. Auxin binding in partially purified
extracts was measured by the following two methods. (i)Equilibrium gel filtration (24). A 1-ml column of SephadexG-25 (Pharmacia) was equilibra~ted with 1 ,uM [14C]2,4-D in 50mM sodium citrate/5 mM MgCl2, pH 5.5. Partially purifiedsoybean extract (2 mg ofprotein) was incubated under similarconditions in a total volume of 100 ,ul and loaded on thepreviously equilibrated column. The column was eluted withthe same buffer containing [14C]2,4-D and fractions werecollected and their radioactivities were measured in a liquidscintillation counter. (it) Ammonium sulfate pelleting assay(25). An alternative method that utilizes ammonium sulfate to
pellet auxin-protein complexes was also used. After theprotein had been incubated with ['4C]2,4-D (1 ttM) at 40C for15 min in a total volume of 0.1 ml, saturated ammoniumsulfate (0.9 ml) was added and the precipitated proteins werepelleted at 47,000 x g for 20 min. The supernatant wascompletely removed and the pellets were washed once with1 ml of 90% saturated ammonium sulfate. The radioactivityassociated with the pellets was measured.
Isolation of Nuclei. Nuclei from soybean plumules wereprepared by the method of Hagen and Guilfoyle (26). Thepurity was estimated by light microscopy using differentialinterference contrast (DIC) optics and fluorescence micros-copy in the presence of Hoechst 33258 dye (MolecularProbes) in a Nikon Microphot FXA microscope. Proteinsfrom the nuclear and cytosolic fractions were prepared forSDS/PAGE and immunoblot analysis as described previ-ously.
RESULTSProduction of Anti-idiotypic Antibodies. Purified IgG and
Fab fractions of anti-IAA antibodies had binding constantsfor IAA of 4.1 x 107 M1 and 1.1 x 107 M-1, respectively,which are in good agreement with reported values (19). Theanti-IAA antibodies purified on a column ofimmobilized IAAshowed [3H]IAA binding activity after the removal of freeIAA, whereas the nonretained fraction lacked such an activ-ity (data not shown), indicating that IAA chromatographywas effective. When anti-idiotypic antibodies were raisedagainst purified anti-IAA antibodies in rabbits, the antiserumfrom one of the animals recognized the Fab fragments ofanti-IAA antisera, whereas preimmune serum from the sameanimal had no such activity (data not shown). When 1 AMIAA was included in the incubation mixture, the binding ofanti-idiotypic antibodies to the Fab fragments was partiallyinhibited (38-47%), indicating the presence of antibodiesdirected against the variable regions of anti-IAA antibodies.
Reactivity of Anti-idibtypic Antibodies with Plant Extracts.The antibodies were used to probe immunoblots of total cellextracts from several plants representing both monocots anddicots. A protein of 65 kDa was recognized by the anti-idiotypic antiserum in all the plants tested (Fig. 1A) whereaspreimmune serum did not react with any proteins in ourextracts (Fig. 1B). Soybean and mung bean extracts con-tained an additional band at 14 kDa that may be a proteolyticproduct of the 65-kDa band, since in freshly prepared ex-tracts from these plants the smaller protein was not seen. InArabidopsis, a protein of 35 kDa is also recognized by theantiserum in addition to the 65-kDa band. While we do not
A BkDa a b c d e f g h a b c d e f g h
9765
29-. _
,-....:.
FIG. 1. Reactivity of anti-idiotypic antiserum with plant extracts.Extracts were prepared from whole seedlings and the proteins weresubjected to SDS/12% PAGE and immunoblot analysis. Duplicateblots were probed with either the anti-idiotypic antiserum (A) or thepreimmune serum (B). Approximately 100 1Lg of protein was loadedin each lane. Corresponding lanes from panels A and B containproteins from Vigna radiata (a), Glycine max (b), Arabidopsisthaliana (c), Zea mays (d), Triticum aestivum (e), Lycopersiconesculentum (t), Nicotiana tabacum (g), and Petunia hybrida (h).
Proc. NatL Acad. Sci. USA 88 (1991)
Proc. Natl. Acad. Sci. USA 88 (1991) 5481
know the relationship between these two proteins in Arabi-dopsis, either they may represent two different ABPs sharinghomology at the ligand-binding site or the 35-kDa protein maybe derived from the 65-kDa protein by proteolysis. Theanti-idiotypic antibodies did not recognize the purified 22-kDa maize ABP or any other membrane-associated ABPs onblots. However, preliminary results show that the 22-kDaABP is retained on the anti-idiotypic immunoaffinity column(data not shown).
In theory, antibodies interacting at or near the auxin-binding site should be displaced by auxin (13). We tested thisby incubating the soybean protein on blots with anti-idiotypicantiserum in the presence of several concentrations of theauxins IAA and 2,4-D and salicylic acid, a structurally similarcompound that is not an auxin. As seen in Fig. 2, both thenatural and the synthetic auxin modified the immunoreactiv-ity of the anti-idiotypic antibodies with the 65-kDa protein,whereas salicylic acid, an inactive compound, had no effect.These compounds also modulated the immunoreactivity ofthe 14-kDa protein seen in soybean (data not shown).A range of ligand concentration was used to determine the
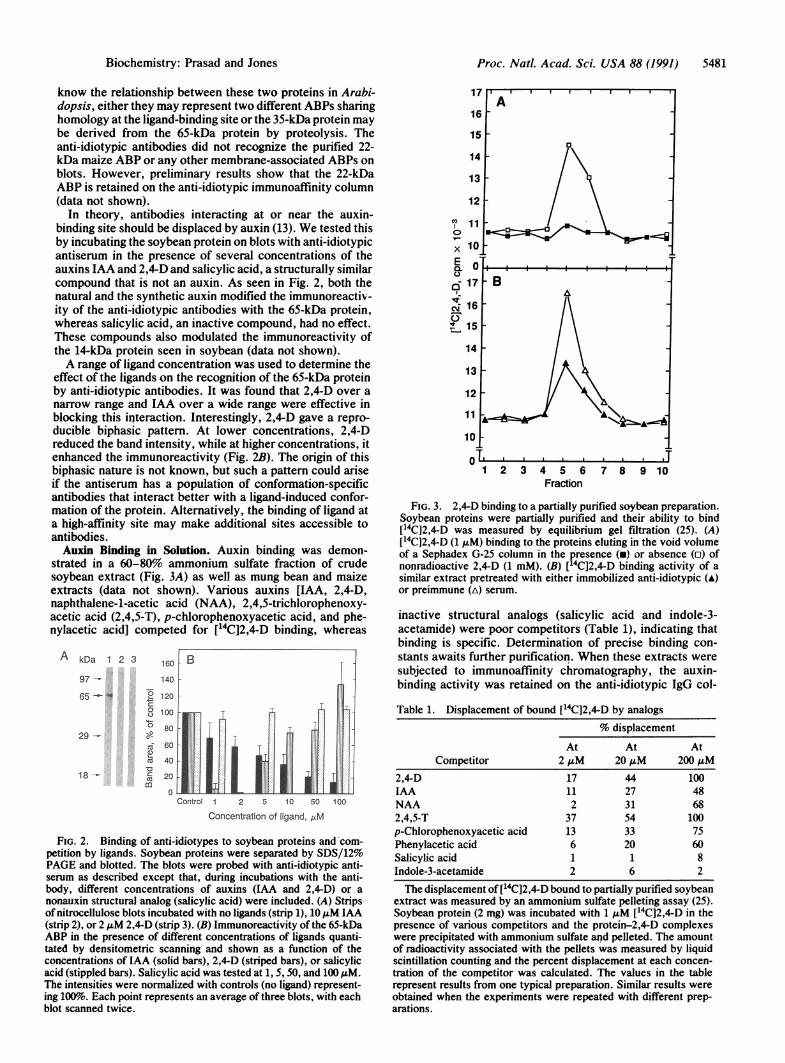
effect of the ligands on the recognition of the 65-kDa proteinby anti-idiotypic antibodies. It was found that 2,4-D over anarrow range and IAA over a wide range were effective inblocking this interaction. Interestingly, 2,4-D gave a repro-ducible biphasic pattern. At lower concentrations, 2,4-Dreduced the band intensity, while at higher concentrations, itenhanced the immunoreactivity (Fig. 2B). The origin of thisbiphasic nature is not known, but such a pattern could ariseif the antiserum has a population of conformation-specificantibodies that interact better with a ligand-induced confor-mation of the protein. Alternatively, the binding of ligand ata high-affinity site may make additional sites accessible toantibodies.Auxin Binding in Solution. Auxin binding was demon-
strated in a 60-80% ammonium sulfate fraction of crudesoybean extract (Fig. 3A) as well as mung bean and maizeextracts (data not shown). Various auxins [IAA, 2,4-D,naphthalene-1-acetic acid (NAA), 2,4,5-trichlorophenoxy-acetic acid (2,4,5-T), p-chlorophenoxyacetic acid, and phe-nylacetic acid] competed for [14C]2,4-D binding, whereas
A kDa 1 2 3 BT
97 140
65 - 120
0100
080T
29 _60Ta~~~~~~~~40
18- 20
0
Control 1 2 5 10 50 100
Concentration of ligand, AM
FIG. 2. Binding of anti-idiotypes to soybean proteins and com-petition by ligands. Soybean proteins were separated by SDS/12%PAGE and blotted. The blots were probed with anti-idiotypic anti-serum as described except that, during incubations with the anti-body, different concentrations of auxins (IAA and 2,4-D) or anonauxin structural analog (salicylic acid) were included. (A) Stripsof nitrocellulose blots incubated with no ligands (strip 1), 10 ,uM IAA(strip 2), or 2 ,uM 2,4-D (strip 3). (B) Immunoreactivity of the 65-kDaABP in the presence of different concentrations of ligands quanti-tated by densitometric scanning and shown as a function of theconcentrations of IAA (solid bars), 2,4-D (striped bars), or salicylicacid (stippled bars). Salicylic acid was tested at 1, 5, 50, and 100 /AM.The intensities were normalized with controls (no ligand) represent-ing 100%o. Each point represents an average of three blots, with eachblot scanned twice.
co 11 ,. _
X 10
EL 0 i, ,Xc17 B
C\F 16
15
14-
13-
12-
1102101
1 2 3 4 5 6 7 8 9 10Fraction
FIG. 3. 2,4-D binding to a partially purified soybean preparation.Soybean proteins were partially purified and their ability to bind[14C]2,4-D was measured by equilibrium gel filtration (25). (A)[14C]2,4-D (1 ,uM) binding to the proteins eluting in the void volumeof a Sephadex G-25 column in the presence (m) or absence (o) ofnonradioactive 2,4-D (1 mM). (B) [14C]2,4-D binding activity of asimilar extract pretreated with either immobilized anti-idiotypic (A)
or preimmune (A) serum.
inactive structural analogs (salicylic acid and indole-3-acetamide) were poor competitors (Table 1), indicating thatbinding is specific. Determination of precise binding con-stants awaits further purification. When these extracts weresubjected to immunoaffinity chromatography, the auxin-binding activity was retained on the anti-idiotypic IgG col-
Table 1. Displacement of bound [14C]2,4-D by analogs% displacement
At At AtCompetitor 2 AiM 20 ,uM 200 AzM
2,4-D 17 44 100IAA 11 27 48NAA 2 31 682,4,5-T 37 54 100p-Chlorophenoxyacetic acid 13 33 75Phenylacetic acid 6 20 60Salicylic acid 1 1 8Indole-3-acetamide 2 6 2
The displacement of [14C]2,4-D bound to partially purified soybeanextract was measured by an ammonium sulfate pelleting assay (25).Soybean protein (2 mg) was incubated with 1 AuM [14C]2,4-D in thepresence of various competitors and the protein-2,4-D complexeswere precipitated with ammonium sulfate and pelleted. The amountof radioactivity associated with the pellets was measured by liquidscintillation counting and the percent displacement at each concen-tration of the competitor was calculated. The values in the tablerepresent results from one typical preparation. Similar results wereobtained when the experiments were repeated with different prep-arations.
Biochemistry: Prasad and Jones
5482 Biochemistry: Prasad and Jones
umn but not on the preimmune IgG column. Retention on thecolumn was demonstrated by measuring auxin-binding ac-tivity in the flow-through of immune and preimmune col-umns, using the equilibrium gel filtration method (Fig. 3B).These results are consistent with the results obtained withligand competition on immunoblots (Fig. 2) and show that theantiserum recognizes an ABP in solution. ['4C]2,4-D waschosen as the radioligand in these experiments to minimizethe possibility ofdecomposition, which readily occurs to IAAin crude plant extracts.
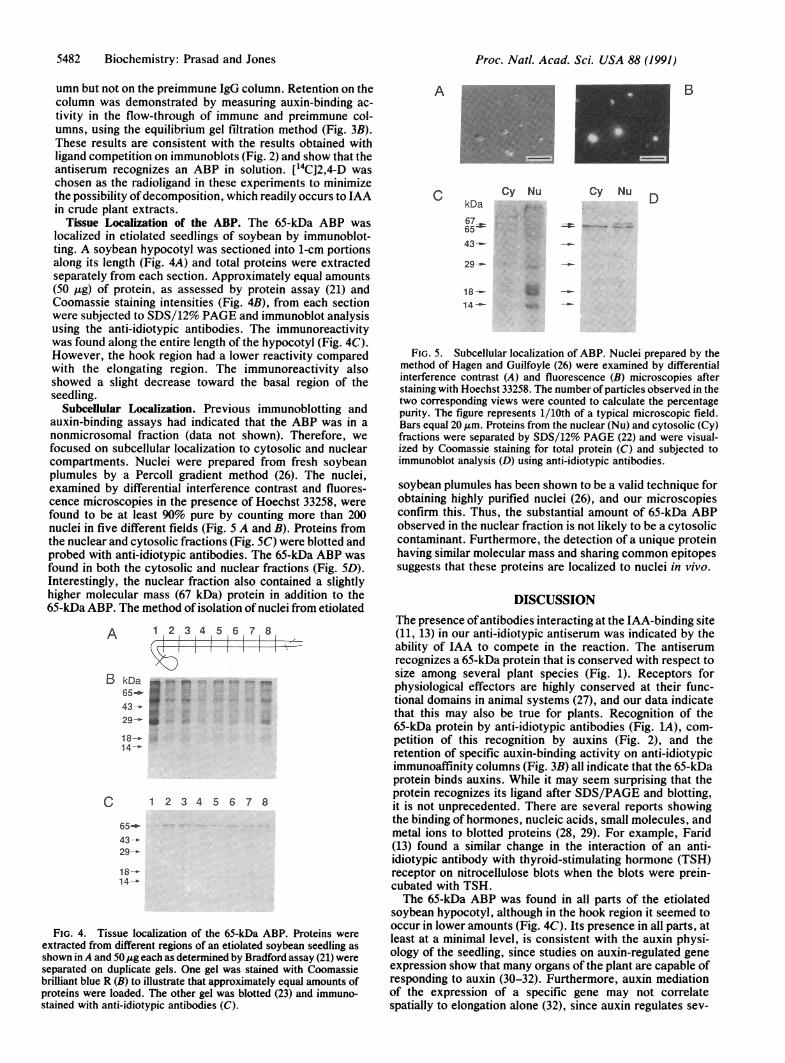
Tissue Localization of the ABP. The 65-kDa ABP waslocalized in etiolated seedlings of soybean by immunoblot-ting. A soybean hypocotyl was sectioned into 1-cm portionsalong its length (Fig. 4A) and total proteins were extractedseparately from each section. Approximately equal amounts(50 /Lg) of protein, as assessed by protein assay (21) andCoomassie staining intensities (Fig. 4B), from each sectionwere subjected to SDS/12% PAGE and immunoblot analysisusing the anti-idiotypic antibodies. The immunoreactivitywas found along the entire length of the hypocotyl (Fig. 4C).However, the hook region had a lower reactivity comparedwith the elongating region. The immunoreactivity alsoshowed a slight decrease toward the basal region of theseedling.
Subcellular Localization. Previous immunoblotting andauxin-binding assays had indicated that the ABP was in anonmicrosomal fraction (data not shown). Therefore, wefocused on subcellular localization to cytosolic and nuclearcompartments. Nuclei were prepared from fresh soybeanplumules by a Percoll gradient method (26). The nuclei,examined by differential interference contrast and fluores-cence microscopies in the presence of Hoechst 33258, werefound to be at least 90% pure by counting more than 200nuclei in five different fields (Fig. 5 A and B). Proteins fromthe nuclear and cytosolic fractions (Fig. SC) were blotted andprobed with anti-idiotypic antibodies. The 65-kDa ABP wasfound in both the cytosolic and nuclear fractions (Fig. SD).Interestingly, the nuclear fraction also contained a slightlyhigher molecular mass (67 kDa) protein in addition to the65-kDa ABP. The method of isolation of nuclei from etiolated
A 1 2 3 4 5 6 7 8
B kDa65
4291
18-14-
C 1 2 3 4 5 6 7 8
65-43-29-
18 --14-
FIG. 4. Tissue localization of the 65-kDa ABP. Proteins wereextracted from different regions of an etiolated soybean seedling asshown inA and 50kg each as determined by Bradford assay (21) wereseparated on duplicate gels. One gel was stained with Coomassiebrilliant blue R (B) to illustrate that approximately equal amounts ofproteins were loaded. The other gel was blotted (23) and immuno-stained with anti-idiotypic antibodies (C).
A
C
B
Cy Nu Cy NukDa .
6765_43-_
D
18--14 -_
FIG. 5. Subcellular localization of ABP. Nuclei prepared by themethod of Hagen and Guilfoyle (26) were examined by differentialinterference contrast (A) and fluorescence (B) microscopies afterstaining with Hoechst 33258. The number of particles observed in thetwo corresponding views were counted to calculate the percentagepurity. The figure represents 1/10th of a typical microscopic field.Bars equal 20 ,um. Proteins from the nuclear (Nu) and cytosolic (Cy)fractions were separated by SDS/12% PAGE (22) and were visual-ized by Coomassie staining for total protein (C) and subjected toimmunoblot analysis (D) using anti-idiotypic antibodies.
soybean plumules has been shown to be a valid technique forobtaining highly purified nuclei (26), and our microscopiesconfirm this. Thus, the substantial amount of 65-kDa ABPobserved in the nuclear fraction is not likely to be a cytosoliccontaminant. Furthermore, the detection of a unique proteinhaving similar molecular mass and sharing common epitopessuggests that these proteins are localized to nuclei in vivo.
DISCUSSIONThe presence ofantibodies interacting at the IAA-binding site(11, 13) in our anti-idiotypic antiserum was indicated by theability of IAA to compete in the reaction. The antiserumrecognizes a 65-kDa protein that is conserved with respect tosize among several plant species (Fig. 1). Receptors forphysiological effectors are highly conserved at their func-tional domains in animal systems (27), and our data indicatethat this may also be true for plants. Recognition of the65-kDa protein by anti-idiotypic antibodies (Fig. 1A), com-petition of this recognition by auxins (Fig. 2), and theretention of specific auxin-binding activity on anti-idiotypicimmunoaffinity columns (Fig. 3B) all indicate that the 65-kDaprotein binds auxins. While it may seem surprising that theprotein recognizes its ligand after SDS/PAGE and blotting,it is not unprecedented. There are several reports showingthe binding of hormones, nucleic acids, small molecules, andmetal ions to blotted proteins (28, 29). For example, Farid(13) found a similar change in the interaction of an anti-idiotypic antibody with thyroid-stimulating hormone (TSH)receptor on nitrocellulose blots when the blots were prein-cubated with TSH.The 65-kDa ABP was found in all parts of the etiolated
soybean hypocotyl, although in the hook region it seemed tooccur in lower amounts (Fig. 4C). Its presence in all parts, atleast at a minimal level, is consistent with the auxin physi-ology of the seedling, since studies on auxin-regulated geneexpression show that many organs of the plant are capable ofresponding to auxin (30-32). Furthermore, auxin mediationof the expression of a specific gene may not correlatespatially to elongation alone (32), since auxin regulates sev-
Proc. NatL Acad. Sci. USA 88 (1991)

Proc. Natl. Acad. Sci. USA 88 (1991) 5483
eral physiological and developmental processes besides cellelongation. Thus, it is probable that all parts of the plantcontain receptors capable of perceiving the auxin signal andtransducing it uniquely.Auxins cause rapid changes in specific gene transcription
in isolated organs, in some cases increasing the messagelevels of specific transcripts within minutes (26, 30, 31). Themolecular mechanism by which these rapid effects arebrought about is unknown, although endogenous and appliedauxins were localized to the nucleus in several plant species(33-35), suggesting that auxins bind some nuclear compo-nent. Analogously to the steroid receptor family in animals,it is possible that a receptor-auxin complex interacts directlywith regulatory sequences on genomic DNA to affect tran-scription. Therefore, the localization of the 65-kDa ABP tonuclei raises the possibility that the 65-kDa ABP and/or theunique 67-kDa protein mediates auxin-regulated gene expres-sion. The relationship between the two nuclear ABPs isunclear; however, one possibility is that the 67-kDa proteinis a covalently modified form ofthe 65-kDaABP similar to thephosphorylated nuclear estrogen receptor (36). Due to thepossibility of nuclear rupture and the leakage of nuclearcontent into the cytoplasm during the isolation of nuclei, wecannot say unequivocally that the 65-kDa ABP is also locatedin the cytoplasm in vivo.The 65-kDa ABP described here differs from other ABPs
previously described. The maize ABP (ABP1) is locatedprimarily in the endoplasmic reticulum (37), the zucchini42-kDa ABP is located on the plasma membrane (5), and the65-kDa ABP is nuclear and possibly cytosolic in location. Theligand-binding characteristics of these proteins are quitedifferent (cf. refs. 2, 3, and 5 and Table 1). Zucchini 42-kDaABP and the 65-kDa ABP appear to be more widely distrib-uted among plant species than ABP1, which has been re-ported to occur mainly in cereals. Thus, the conservation ofthe 65-kDa ABP in several plants and its unique localizationto the nucleus make it a strong candidate for an auxinreceptor that mediates important auxin-regulated biologicalprocesses.The anti-idiotypic antibody strategy that we have used may
be widely applicable to the study of receptor systems forother plant growth regulators (e.g., see ref. 38). While theidentification of a binding protein by this approach does notconstitute a positive proof that it is a physiological receptorunless such a claim is validated by a functional biochemicalassay (39), it can nevertheless be used as a first screen toisolate putative receptors whose function can then be bio-chemically evaluated.
We thank Dr. James B. Hicks (Department of Molecular Biology,Scripps Clinic and Research Foundation, La Jolla, CA) for providingthe support and encouragement to initiate this work. Financialsupport was provided by PPG Industries, Inc. (Pittsburgh, PA) (toP.V.P.) and the National Science Foundation (to A.M.J.).
1. Davies, P. J. (1987) in Plant Hormones and Their Role in PlantGrowth and Development, ed. Davies, P. J. (Nijhoff, Dor-drecht, The Netherlands), pp. 1-11.
2. Lobler, M. & Klambt, D. (1985) J. Biol. Chem. 260, 9848-9853.
3. Jones, A. M. & Venis, M. A. (1989) Proc. Natl. Acad. Sci.USA 86, 6153-6156.
4. Hicks, G. R., Rayle, D. L. & Lomax, T. L. (1989) Science 245,52-54.
5. Hicks, G. R., Rayle, D. L., Jones, A. M. & Lomax, T. L.(1989) Proc. Natl. Acad. Sci. USA 86, 4948-4952.
6. Jones, A. M. (1990) Physiol. Plant. 80, 154-158.7. Barbier-Brygoo, H., Ephritikhine, G., Klambt, D., Ghislain,
M. & Guern, J. (1989) Proc. Natl. Acad. Sci. USA 86, 891-895.8. Rubery, P. H. (1987) in Plant Hormones and Their Role in
Plant Growth and Development, ed. Davies, P. J. (Nijhoff,Dordrecht, The Netherlands), pp. 341-362.
9. Libbenga, K. R. & Mennes, A. M. (1987) in Plant Hormonesand Their Role in Plant Growth and Development, ed. Davies,P. J. (Nijhoff, Dordrecht, The Netherlands), pp. 194-221.
10. Venis, M. A. (1985) in Hormone Binding Sites in Plants,(Longman, New York), pp. 106-114.
11. Farid, N. & Lo, T. C. Y. (1985) Endocrine Rev. 6, 1-23.12. Pain, D. K., Kanwar, Y. S. & Blobel, G. (1988) Nature (Lon-
don) 331, 232-237.13. Farid, N. (1989) Methods Enzymol. 178, 191-212.14. Vaux, D., Tooze, J. & Fuller, S. (1990) Nature (London) 345,
495-502.15. Pain, D., Murakami, H. & Blobel, G. (1990) Nature (London)
347, 444-449.16. Wilchek, M., Miron, T. & Kohn, J. (1984) Methods Enzymol.
104, 3-55.17. Wasserman, N. H., Penn, A. S., Freimuth, P. I., Trepton, N.,
Wentzel, S., Cleveland, W. L. & Erlanger, B. F. (1982) Proc.Natl. Acad. Sci. USA 79, 4810-4814.
18. Schreiber, M., Fogelfeld, L., Souroujon, M. C., Kohen, F. &Fuchs, S. (1983) Life Sci. 33, 1519-1526.
19. Pengelly, W. & Meins, F., Jr. (1977) Planta 136, 173-180.20. Pence, V. C. & Caruso, J. L. (1987) in Plant Hormones and
Their Role in Plant Growth and Development, ed. Davies, P. J.(Nijhoff, Dordrecht, The Netherlands), pp. 240-256.
21. Bradford, M. (1976) Anal. Biochem. 72, 248-254.22. Laemmli, U. K. (1970) Nature (London) 227, 680-685.23. Towbin, H., Staehelin, T. & Gordon, J. (1979) Proc. Natl.
Acad. Sci. USA 76, 4350-4354.24. Hummel, J. P. & Dreyer, W. J. (1962) Biochim. Biophys. Acta
63, 530-532.25. Polya, G. M. & Davies, A. W. (1978) Planta 139, 139-147.26. Hagen, G. & Guilfoyle, T. (1985) Mol. Cell. Biol. 5, 1197-1203.27. Gustafson, J. A., Carlstedt-Duke, J., Poellinger, L., Okret, S.,
Wikstrom, A. C., Bronnegard, M., Gillner, M., Dong, Y.,Fuxe, K., Cintra, A., Harfstrand, A. & Agnati, L. (1987)Endocrine Rev. 8, 185-234.
28. Bers, G. & Garfin, D. (1985) Biotechniques 3, 276-288.29. Gershoni, J. M. (1989) Methods Biochem. Anal. 33, 1-59.30. Guilfoyle, T. (1986) CRC Crit. Rev. Plant Sci. 4, 247-276.31. Theologis, A., Huynh, T. V. & Davis, R. W. (1985) J. Mol.
Biol. 183, 53-68.32. Wright, R. M., Hagen, G. & Guilfoyle, T. (1987) Plant Mol.
Biol. 9, 625-634.33. Liao, S.-H. & Hamilton, R. H. (1966) Science 151, 822-824.34. Perbal, G. & Driss-Ecole, D. (1988) Biol. Cell 1988 62, 275-280.35. Ohmiya, A., Hayashi, T. & Kakiuchi, N. (1990) Plant Cell
Physiol. 31, 711-715.36. Washburn, T., Hocutt, A., Brautigan, D. L. & Korach, K. S.
(1991) Mol. Endocrinol. 5, 235-242.37. Jones, A. M., Lamerson, P. & Venis, M. A. (1989) Planta 179,
409-413.38. Hooley, R., Beale, M. H., Smith, S. J. & MacMillan, J. (1990)
in Plant Growth Substances 1988, eds. Pharis, R. P. & Rood,S. B. (Springer, Berlin), pp. 145-153.
39. Meyer, D. I. (1990) Nature (London) 347, 424-425.
Biochemistry: Prasad and Jones