purification of cartilage-derived growth factor by heparin affinity ... · purification of...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1985 by The American Society of Biological Chemists, h e .
Vol. 260, No. 4, Issue of February 25, pp. 2399-2403,1985 Printed in U S. A.
Purification of Cartilage-derived Growth Factor by Heparin Affinity Chromatography*
(Received for publication, August 13, 1984)
Robert Sullivan and Michael KlagsbrunS From the DeDartments of Biolopical Chemistry and Surgery, The Children’s Hospital and Harvard Medical School, ~~
Boston, MasBachusetts 021 15 -
Cartilage-derived growth factor (CDGF) was found to bind tightly to columns of immobilized heparin and could be eluted with concentrations of salt in the order of 1.6-1.8 M NaC1. The molecular weight of CDGF was estimated to be 18,000-20,000 by high performance liquid-size exclusion chromatography. The affinity of CDGF for heparin greatly facilitated its purification. Highly purified CDGF active at about 1-2 ng/ml was obtained when crude cartilage extract was applied to heparin-Sepharose and the growth factor activity was recycled over heparin-Sepharose two more times. Analysis by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and silver stain visualization of highly purified CDGF showed one major polypeptide band with a molecular weight of about 19,000 containing over 95% of the protein and one minor polypeptide band containing the rest of the protein. Only the M, 19,000 polypeptide was active after elution from the polyacrylamide gel. Although CDGF bound tightly to immobilized heparin, it did not bind to immobilized chondroitin sulfate or hyaluronic acid. In addition, CDGF bound to heparin much more tightly than did platelet-derived growth factor even though these two growth factors had similar isoelectric points of about 10. These results suggest that the binding of CDGF to heparin was due to a specific affinity of the 2 molecules for each other.
In order to have a better understanding of the regulation of cartilage and bone metabolism, a great deal of investigation has been focused on the role of growth and maturation factors found in these tissues. Growth and maturation factors have been isolated from cartilage (1-5) and from bone (5-9). Sev- eral years ago our laboratory purified a cartilage-derived growth factor to apparent homogeneity (2). The purified CDGF’ was active in stimulating DNA synthesis in 3T3 cells a t 1-5 pg/ml. However, other purified 3T3 cell growth factors such as fibroblast growth factor and platelet-derived growth factor were active at 10 ng/ml. I t was thought that the relatively low specific activity might have been in part due to the harsh denaturation techniques used in the purification of
*This work was supported by Grant R01 CA 37392 from the National Cancer Institute. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ To whom correspondence should be addressed at: The Children’s Hospital, 300 Longwood Ave., Boston, MA 02115.
The abbreviations used are: CDGF, cartilage-derived growth fac- tor; HPLC, high performance liquid chromatography; SDS-PAGE,
platelet-derived growth factor. sodium dodecyl sulfate-polyacrylamide gel electrophoresis; PDGF,
CDGF. Recently, a chondrosarcoma-derived growth factor with a molecular weight of about 18,000 was purified to apparent homogeneity in our laboratory by taking advantage of its strong affinity for heparin (10). Two steps were required for the purification: cation exchange and heparin affinity chromatography. In this report we demonstrate that a growth factor with a strong affinity for heparin can be isolated from normal cartilage. CDGF can be purified to a very high degree by using several cycles of heparin affinity chromatography as the only biochemical purification step. CDGF purified by this method has a molecular weight of 19,000 and is active at about 1-2 ng/ml.
EXPERIMENTAL PROCEDURES
Preparation of Cartiluge-derived Growth Factor Extracts-Extracts of CDGF were prepared from bovine scapular cartilage, bovine artic- ular cartilage, human costal cartilage, and chick sternum cartilage. Although this report describes bovine scapula CDGF, CDGF from the other sources gave the same results. Two methods were used for the preparation of cartilage extracts. 1) Cartilage was extracted with 1 M guanidine-HC1, pH 6.5, for 24 h at room temperature as described previously (2). 2) Cartilage was digested with 2 mg/ml of clostridial collagenase to produce an extracellular matrix fraction as described previously (11).
Heparin-Sepharose Chromatography-Heparin-Sepharose was purchased from Pharmacia. Heparin-Sepharose chromatography of CDGF was carried out either by hatch elution or by gradient elution. 1) Batch elution was used as a first step for large scale purification. Extracts of CDGF (5-20 g/300 ml) were dissolved in a column equilibration buffer of 0.1 M NaCl and 0.01 M Tris-HC1, pH 7.5. The extracts were applied at 4 “C to columns of heparin-Sepharose (160- ml bed volume, 5 X 8 cm) at a flow rate of 60 ml/h. Alternatively, cartilage extract (15 g/300 ml) and heparin-Sepharose (160 ml) were mixed by shaking overnight a t 4 “C. The supernatant was discarded and the heparin-Sepharose was washed with equilibration buffer and was poured into a column. The heparin-Sepharose columns were subsequently washed with consecutive rinses of 0.01 M Tris-HCI, pH 7.5, supplemented with 0.1 M NaCl (400 ml), with 0.5 M NaCl (300 ml), and with 3.0 M NaCl (300 ml). Virtually all of the CDGF was found in the 0.5-3.0 M NaCl fraction. Starting with about 10 g of guanidine extract, about 10 mg of partially purified CDGF containing about 100,000 units of growth factor activity were recovered in the 0.5-3.0 M NaCl batch eluted fraction. 2) Gradient elution was used for samples of 1 g or less. Columns of heparin-Sepharose (10-40-ml bed volume) were equilibrated with 0.1 M NaCl and 0.01 M Tris-HC1, pH 7.5. Samples (40 mg/ml or less) were applied to the column. After a rinse with 4-10 column volumes of equilibration buffer, CDGF was eluted with either a 0.1-3.0 M NaCl gradient or a 0.6-3.0 M NaCl gradient a t a flow rate of about 30-40 ml/h. The volume of the gradient ranged from 200-600 ml depending on the bed volume and the purity of sample (see figure legends). About 320-600 ml were used for 6-40-ml bed volumes when crude or partially purified CDGF was applied and 160-200 ml were used for 4-10-ml bed volumes when highly purified CDGF was applied. Fractions of 8 ml were collected and each fraction was monitored directly for absorbance, conductiv- ity, and growth factor activity. Fractions containing growth factor activity obtained by either hatch or gradient elution were dialyzed (Spectraphor dialysis tubing, 6,000-8,000 molecular weight cutoff)
2399

2400 Cartilage-derived Growth Factor
'
/
/ /
/ /'
/
- 3.0
' 2.5
z 0
z 1.5 - 2.0 Q
- n
n I I I
1.0 I Y
0.5
0
l4.3
D a3
3.0 aJ UJ
z 2.5 D 0 m
2.0
Iv 1.5 03 0 3
1.0 3 n
0.5 I v
0 IO 20 30 40 50 60 70
FRACTION NUMBER FIG. 1. Heparin-Sepharose chromatography of CDGF. Bovine scapula cartilage was extracted with 1 M
guanidine HCI, pH 6.5. The extract (1 g/25 ml, 30,000 units) was applied to a column of heparin-Sepharose (2.5 X 8 cm, 40-ml bed volume). After a rinse with 160 ml of equilibration buffer, a gradient (500 ml) of 0.1-3.0 M NaCl was applied. Fractions (8 ml) were collected and measured for absorbance (O), conductivity (---), and growth factor activity (0). About 20,000 units were recovered. Inset, the peak of growth factor activity (fraction 54) was analyzed by SDS-PAGE and silver stain. Lane I, CDGF Lane 2, molecular weight markers.
against deionized double-distilled HzO, lyophilized, and stored a t -20 "C. The recovery of growth factor activity from columns of heparin-Sepharose ranged from 3040%. Dialyzed fractions were used to estimate recovery.
HPLC-Size Exclusion Chromatography-The molecular weight of CDGF was estimated by HPLC-size exclusion chromatography at room temperature (Beckman Model 332 gradient liquid chromatog- raphy system) on TSK 2000 columns (60 cm long X 7.5 mm inner diameter, Altex) equilibrated with 0.6 M NaCl and 0.02 M Tris-HCI, pH 7.0. The flow rates were 0.5 ml/min and fractions of 0.8 ml were collected and tested for growth factor activity. TSK 2000 columns were calibrated with Blue Dextran (2 X lo'), serum albumin (68,000), ovalbumin (43,000), a-chymotrypsinogen (25,700), and lysozyme (14,300).
Immobilization of Glycosaminoglycans-Immobilized chondroitin sulfate, immobilized hyaluronic acid, and immobilized heparin were prepared. Slurries (10 ml) of Affi-Gel 102 (Bio-Rad) were adjusted to pH 4.7-5.0 with 0.1 M HCI. About 50 mg/lO ml of Hz0 of chondroitin sulfate ABC (grade 111, mixed isomers, contains chondroitin 4-sulfate, chondroitin 6-sulfate, and dermatan sulfate, Sigma), hyaluronic acid (grade I, Sigma), or heparin (Hepar Industries, Franklin, Ohio) were added to each slurry followed by 50 mg of l-ethyl-3-(3-dimethylami- nopropy1)carbodiimide HCI (Bio-Red). The reactions proceeded with constant stirring for 2 h at room temperature (12). The slurries were centrifuged at 1500 X g and the supernatants were assayed for unbound glycosaminoglycan (13). Greater than 70% of the glycos- aminoglycans were bound, resulting in preparations of 7 mg of gly- cosaminoglycan/ml of packed Affi-GellO2.
Sodium Dodecyl Sulfate-Polyacrylumide Gel Electrophoresis- CDGF was analyzed by SDS-PAGE (14) followed by silver stain visualization (15). Molecular weight markers containing ovalbumin (43,000), a-chymotrypsinogen (25,700), lactalbumin (18,400), lyso- zyme (14,300), and trypsin inhibitor (6,200) were purchased from Bethesda Research Laboratories. CDGF was eluted from polyacryl- amide gels as follows. CDGF and molecular weight markers were electrophoresed by SDS-PAGE. A lane containing molecular weight markers and a lane containing CDGF were fixed and stained with silver. Another lane containing molecular weight standards (10 pg each) was cut out and stained rapidly by fixing with 50% trichloroa- cetic acid (15 min), staining with 0.25% Coomassie Blue (20 rnin), and destaining with 10% acetic acid (15 min). The rest of the polyacrylamide gel which contained CDGF was refrigerated while the molecular weight markers were being stained. Using the rapidly stained molecular weight markers as a guide (especially the M, 18,400 lactalbumin marker), the polyacrylamide gel containing CDGF was cut into 6 segments. The 6 segments were diced finely, suspended in phosphate-buffered saline, and incubated overnight at room temper- ature. The eluates were dialyzed exhaustively (Mr 6,000-8,000 cutoff) against double-distilled deionized Hz0 and tested for growth factor activity.
Measurement of Growth Factor Activity-Growth factor activity was determined by measuring the ability of samples to stimulate the incorporation of 3H-thymidine into the DNA of confluent quiescent BALB/c 3T3 cells (3). The growth factor assay was carried out in 96- well microtiter plates (Costar). Each well contained about 20,000 3T3 cells incubated in 250 pl of Dulbecco's Modified Eagle's Medium
Cartilage-deriued Growth Factor 2401
(Gibco) supplemented with 10% calf serum (Colorado Serum Co., Denver, C O ) . One unit of growth factor activity is defined as the amount of growth factor in 250 ~1 necessary to stimulate half-maximal DNA synthesis in a 36-48-h incubation period (about 50,000-75,000 cpm). For assays of column fractions, a constant volume of each fraction was chosen so that the peak fraction displayed 0.5-1.5 units of growth factor activity (about 25,000 to 100,000 cpm).
RESULTS
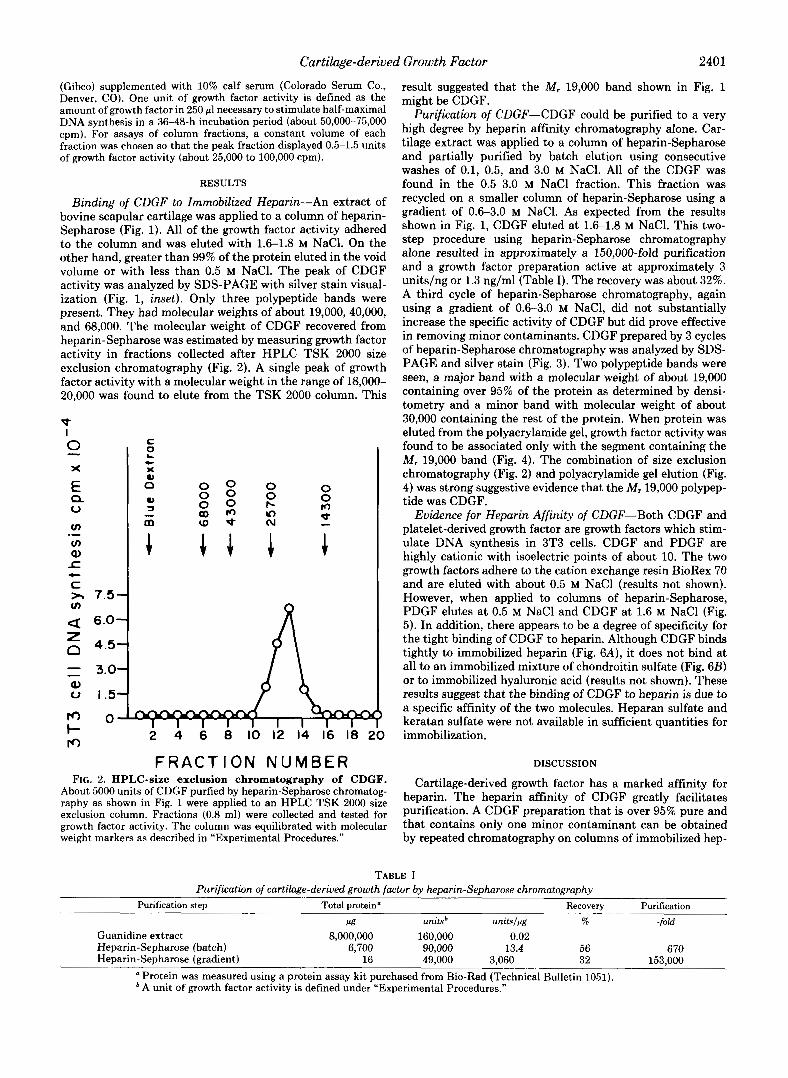
Binding of CDGF to Immobilized Heparin-An extract of bovine scapular cartilage was applied to a column of heparin- Sepharose (Fig. 1). All of the growth factor activity adhered to the column and was eluted with 1.6-1.8 M NaC1. On the other hand, greater than 99% of the protein eluted in the void volume or with less than 0.5 M NaC1. The peak of CDGF activity was analyzed by SDS-PAGE with silver stain visual- ization (Fig. 1, inset). Only three polypeptide bands were present. They had molecular weights of about 19,000,40,000, and 68,000. The molecular weight of CDGF recovered from heparin-Sepharose was estimated by measuring growth factor activity in fractions collected after HPLC TSK 2000 size exclusion chromatography (Fig. 2). A single peak of growth factor activity with a molecular weight in the range of 18,000- 20,000 was found to elute from the TSK 2000 column. This
P I 0 - X
E 0 a
m m a -c c UJ
.-
t
)r 7.5-
Q: 6.0- z 0 4.5- - - 3.0-
0 I .5-
0-
a
I- rc)
C z c X
2 4 6 8 IO 12 14 16 18
F R A C T I O N NUMBER FIG. 2. HPLC-size exclusion chromatography of CDGF.
About 5000 units of CDGF purfied by heparin-Sepharose chromatog- raphy as shown in Fig. 1 were applied to an HPLC TSK 2000 size exclusion column. Fractions (0.8 ml) were collected and tested for growth factor activity. The column was equilibrated with molecular weight markers as described in “Experimental Procedures.”
result suggested that the M, 19,000 band shown in Fig. 1 might be CDGF.
Purification of CDGF-CDGF could be purified to a very high degree by heparin affinity chromatography alone. Car- tilage extract was applied to a column of heparin-Sepharose and partially purified by batch elution using consecutive washes of 0.1, 0.5, and 3.0 M NaCl. All of the CDGF was found in the 0.5-3.0 M NaCl fraction. This fraction was recycled on a smaller column of heparin-Sepharose using a gradient of 0.6-3.0 M NaC1. As expected from the results shown in Fig. 1, CDGF eluted at 1.6-1.8 M NaCl. This two- step procedure using heparin-Sepharose chromatography alone resulted in approximately a 150,000-fold purification and a growth factor preparation active at approximately 3 units/ng or 1.3 ng/ml (Table I). The recovery was about 32%. A third cycle of heparin-Sepharose chromatography, again using a gradient of 0.6-3.0 M NaC1, did not substantially increase the specific activity of CDGF but did prove effective in removing minor contaminants. CDGF prepared by 3 cycles of heparin-Sepharose chromatography was analyzed by SDS- PAGE and silver stain (Fig. 3). Two polypeptide bands were seen, a major band with a molecular weight of about 19,000 containing over 95% of the protein as determined by densi- tometry and a minor band with molecular weight of about 30,000 containing the rest of the protein. When protein was eluted from the polyacrylamide gel, growth factor activity was found to be associated only with the segment containing the M , 19,000 band (Fig. 4). The combination of size exclusion chromatography (Fig. 2) and polyacrylamide gel elution (Fig. 4) was strong suggestive evidence that the M , 19,000 polypep- tide was CDGF.
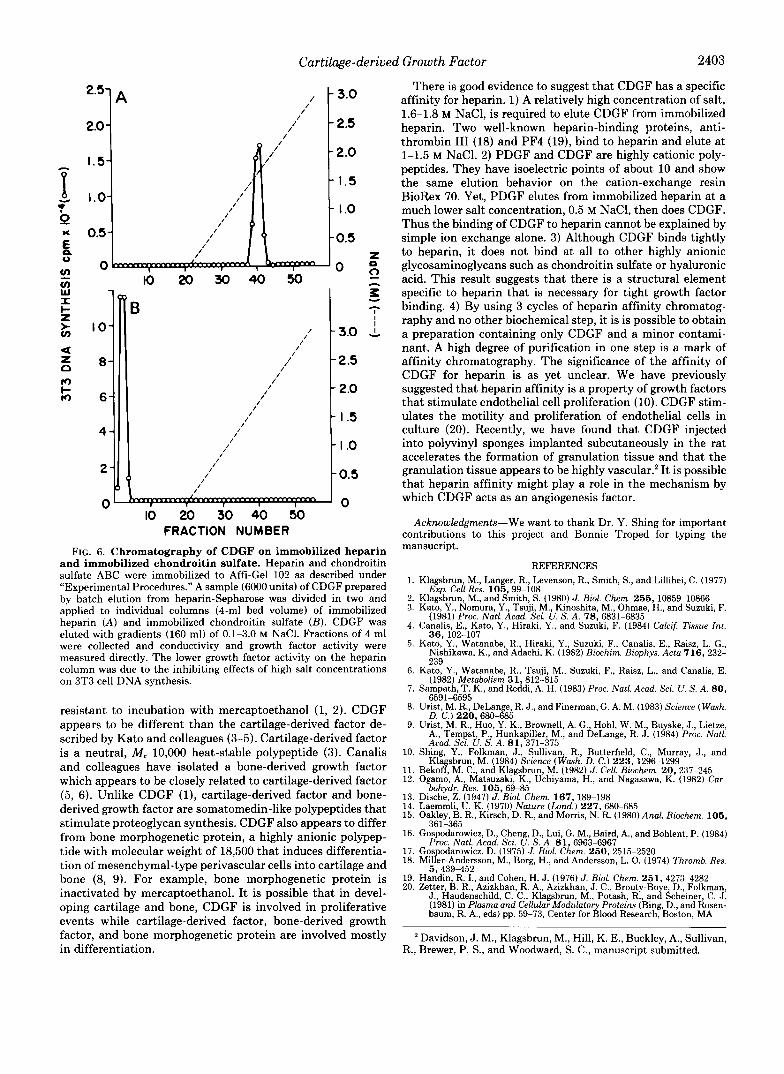
Evidence for Heparin Affinity of CDGF-Both CDGF and platelet-derived growth factor are growth factors which stim- ulate DNA synthesis in 3T3 cells. CDGF and PDGF are highly cationic with isoelectric points of about 10. The two growth factors adhere to the cation exchange resin BioRex 70 and are eluted with about 0.5 M NaCl (results not shown). However, when applied to columns of heparin-Sepharose, PDGF elutes at 0.5 M NaCl and CDGF at 1.6 M NaCl (Fig. 5). In addition, there appears to be a degree of specificity for the tight binding of CDGF to heparin. Although CDGF binds tightly to immobilized heparin (Fig. 6A), it does not bind at all to an immobilized mixture of chondroitin sulfate (Fig. 6B) or to immobilized hyaluronic acid (results not shown). These results suggest that the binding of CDGF to heparin is due to a specific affinity of the two molecules. Heparan sulfate and keratan sulfate were not available in sufficient quantities for immobilization.
DISCUSSION
Cartilage-derived growth factor has a marked affinity for heparin. The heparin affinity of CDGF greatly facilitates purification. A CDGF preparation that is over 95% pure and that contains only one minor contaminant can be obtained by repeated chromatography on columns of immobilized hep-
TABLE I Purification of cartilage-derived growth factor by heparin-Sephnrose chromatography
Purification step Total protein“ Recovery Purification
M unitsb unitsjpg % -fold Guanidine extract 8,000,000 160,000 0.02 Heparin-Sepharose (batch) 6,700 90,000 13.4 56 6 70 Heparin-Sepharose (gradient) 16 49,000 3,060 32 153,000
Protein was measured using a protein assay kit purchased from Bio-Rad (Technical Bulletin 1051). ’ A unit of growth factor activity is defined under “Experimental Procedures.”

2402 Cartilage-derived Growth Factor
SDS PAGE
43 kDa -
2 5 . 7 kDa -
I8.4kDa“-
1 4 . 3 k D a - D 6 . 2 kDa --
FIG. 3. SDS-PAGE analysis of CDGF purified by batch elu- tion and gradient elution heparin-Sepharose chromatogra- phy. A 1 M guanidine HCI cartilage extract (8 g/265 ml, 160,000 units) was applied to a column of heparin-Sepharose (160 ml). The column was washed consecutively with 0.1 M NaCl (400 ml), 0.5 M NaCl (300 ml), and 3.0 M NaCl (300 ml) in Tris-HCI buffer. The 0.5- 3.0 M NaCl fraction (90,000 units) was dialyzed against 0.01 M Tris- HCI, pH 7.5, so as to obtain a solution of 0.6 M NaCl and applied to a heparin-Sepharose column (12-ml bed volume). A gradient (600 ml) of 0.6-3.0 M NaCl was applied and CDGF was eluted at 1.6-1.8 M NaCI. After dialysis, CDGF (50,000 units) was applied to another heparin-Sepharose column (10-ml bed volume, 200 ml of a 0.6-3.0 M NaCl gradient) and recovered (30,000 units). About 3,000 units were analyzed by SDS-PAGE and silver stain.
arin as the only biochemical step. Total purification of CDGF can be achieved by the method used previously in our labo- ratory to purify a chondrosarcoma-derived growth factor to apparent homogeneity, namely a combination of cation-ex- change chromatography and heparin affinity chromatography (10). Heparin affinity chromatography followed by size exclu- sion chromatography also yields pure CDGF. CDGF purified by heparin affinity chromatography has a molecular weight of about 19,000 and is active in stimulating DNA synthesis in 3T3 cells a t about 1-2 ng/ml. It is slightly larger than its tumor-derived counterpart which has a molecular weight of about 18,000 (10).
In our original purification of CDGF, we isolated a M, 16,400 growth factor that was active a t about 1-5 pg/ml (2). There are several possibilities for the apparent discrepancies in specific activities and molecular weight between our origi- nal and present studies. First of all, it is possible that the polypeptide we originally designated as CDGF was a contam- inant and that the actual CDGF was not detected because the highly sensitive silver stain technique was not available a t that time. Alternatively, the harsh chemicals such as guani- dine hydrochloride and urea that were used in the original purification scheme might have resulted in a growth factor with greatly reduced specific activity. Proteolytic activity during the lengthy purification procedure might have ac- counted for the lower molecular weight. A recent report indi- cates that pituitary fibroblast growth factor purified by hep- arin affinity chromatography has a molecular weight of about 16,000 (16). Fibroblast growth factor (purified originally by
W z ‘ 3
pr) 8 r
I I I I I
M.W. CDGF
GEL ELUTION
GEL SEGMENT FIG. 4. Elution of CDGF from polyacrylamide gels. CDGF
was prepared as described in Fig. 3.9000 units were analyzed by SDS- PAGE. A lane containing CDGF (3000 units) was fixed and stained (upper, lane 2). The other lanes containing CDGF were not fixed but were cut into 6 segments (bottom). Each segment was eluted and growth factor activity was measured as described under “Experimen- t a l Procedures.”
3.0 2.5
2.0 g
1.0 i 1.5
1
0.5
.. 0 I O 20 30 40 50 60 70 so 90 100
FRACTION NUMBER FIG. 5. Heparin-Sepharose chromatography of CDGF and
PDGF. Platelet-derived growth factor was obtained from Dr. H. Antoniades, Harvard Medical School, Boston, MA. Extracellular matrix containing CDGF was prepared by collagenase digest of bovine scapular cartilage as described under “Experimental Procedures.” CDGF (40 ml of extracellular matrix, 1.1 g, 10,000 units) and PDGF (5,000 units) were applied to individual columns of heparin-Sepharose (6-ml bed volume, 1 X 8 cm). Growth factor was eluted with a gradient of 320 ml of 0.1-3.0 M NaCl in 0.01 M Tris-HCI, pH 7.0. Fractions of 3 ml were collected and growth factor activity was measured. PDGF, 0 , CDGF, 0.
acid extraction at pH 3.5, ammonium sulfate precipitation, cation-exchange, and size exclusion chromatography) was reported to have a molecular weight of about 13,400 (17). It is possible that in the purification of CDGF and fibroblast growth factor the use of heparin affinity chromatography results in a larger molecular weight form.
It is of interest to compare CDGF to growth and maturation factors in cartilage and bone isolated by other laboratories. CDGF is a cationic polypeptide that is heat-labile but is
Cartilage-derived Growth Factor 2403
2.1” 2.0
I O
8
6
4
2
C
z 0 0
IO 20 30 40 50 ” - - A
z I I
3.0
2.5
2 .o
I .5
I .o
0.5
0 IO 20 30 40 SO
FRACTION NUMBER FIG. 6. Chromatography of CDGF on immobilized heparin
and immobilized chondroitin sulfate. Heparin and chondroitin sulfate ABC were immobilized to Affi-Gel 102 as described under “Experimental Procedures.” A sample (6000 units) of CDGF prepared by batch elution from heparin-Sepharose was divided in two and applied to individual columns (4-ml bed volume) of immobilized heparin (A) and immobilized chondroitin sulfate (B) . CDGF was eluted with gradients (160 ml) of 0.1-3.0 M NaC1. Fractions of 4 ml were collected and conductivity and growth factor activity were measured directly. The lower growth factor activity on the heparin column was due to the inhibiting effects of high salt concentrations on 3T3 cell DNA synthesis.
resistant to incubation with mercaptoethanol (1, 2). CDGF appears to be different than the cartilage-derived factor de- scribed by Kat0 and colleagues (3-5). Cartilage-derived factor is a neutral, M , 10,000 heat-stable polypeptide (3). Canalis and colleagues have isolated a bone-derived growth factor which appears to be closely related to cartilage-derived factor (5, 6). Unlike CDGF ( l ) , cartilage-derived factor and bone- derived growth factor are somatomedin-like polypeptides that stimulate proteoglycan synthesis. CDGF also appears to differ from bone morphogenetic protein, a highly anionic polypep- tide with molecular weight of 18,500 that induces differentia- tion of mesenchymal-type perivascular cells into cartilage and bone (8, 9). For example, bone morphogenetic protein is inactivated by mercaptoethanol. I t is possible that in devel- oping cartilage and bone, CDGF is involved in proliferative events while cartilage-derived factor, bone-derived growth factor, and bone morphogenetic protein are involved mostly in differentiation.
There is good evidence to suggest that CDGF has a specific affinity for heparin. 1) A relatively high concentration of salt, 1.6-1.8 M NaC1, is required to elute CDGF from immobilized heparin. Two well-known heparin-binding proteins, anti- thrombin I11 (18) and PF4 (19), bind to heparin and elute at 1-1.5 M NaC1. 2) PDGF and CDGF are highly cationic poly- peptides. They have isoelectric points of about 10 and show the same elution behavior on the cation-exchange resin BioRex 70. Yet, PDGF elutes from immobilized heparin at a much lower salt concentration, 0.5 M NaCl, then does CDGF. Thus the binding of CDGF to heparin cannot be explained by simple ion exchange alone. 3) Although CDGF binds tightly to heparin, it does not bind at all to other highly anionic glycosaminoglycans such as chondroitin sulfate or hyaluronic acid. This result suggests that there is a structural element specific to heparin that is necessary for tight growth factor binding. 4) By using 3 cycles of heparin affinity chromatog- raphy and no other biochemical step, it is is possible to obtain a preparation containing only CDGF and a minor contami- nant. A high degree of purification in one step is a mark of affinity chromatography. The significance of the affinity of CDGF for heparin is as yet unclear. We have previously suggested that heparin affinity is a property of growth factors that stimulate endothelial cell proliferation (10). CDGF stim- ulates the motility and proliferation of endothelial cells in culture (20). Recently, we have found that CDGF injected into polyvinyl sponges implanted subcutaneously in the rat accelerates the formation of granulation tissue and that the granulation tissue appears to be highly vascular.* It is possible that heparin affinity might play a role in the mechanism by which CDGF acts as an angiogenesis factor.
Acknowledgments-We want to thank Dr. Y. Shing for important contributions to this project and Bonnie Troped for typing the mansucript.
REFERENCES 1. Klagsbrun, M., Langer, R., Levenson, R., Smith, S., and Lillihei, C. (1977)
2. Klagsbrun, M., and Smith, S. (1980) J. Biol. Chem. 255,10859-10866 3. Kato, Y., Nornura, Y., Tsuji, M., Kinoshita, M., Ohmae, H., and Suzuki, F.
(1981) Proc. Natl. Acad. Sci. U. S. A. 78, 6831-6835 4. Canalis, E., Kato, Y., Hiraki, Y., and Suzuki, F. (1984) Calcif. Tissue Int.
3 6 , 102-107 5. Kato, Y., Watanabe, R., Hiraki, Y., Suzuki, F., Canalis, E., Raisz, L. G.,
Nishikawa, K., and Adachi, K. (1982) Biochim. Biophys. Acta 716 , 232-
Exp. Cell Res. 105,99-108
6. Kato, Y., Watanabe, R., Tsuji, M., Suzuki, F., Raisz, L., and Canalis, E.
7. Sampath, T. K., and Reddi, A. H. (1983) Proc. Natl. A d . Sci. U. S. A. 80,
239
(1982) Metabolism 31,812-815
8.
9.
10.
11. 12.
13. 14. 15.
Urist, M. R., DeLange, R. J., and Finerman, G . A. M. (1983) Science (Wash. 6591-6595
Urist, M. R., Huo, Y. K., Brownell, A. G., Hohl, W. M., Buyske, J. , Lietze, D. C.) 220,680-685
A., Tempst, P., Hunkapiller, M., and DeLange, R. J. (1984) Proc. Natl. Acad. Sci. U. S. A. 81,371-375
Shing, Y., Folkman, J., Sullivan, R., Butterfield, C., Murray, J., and Klagsbrun, M. (1984) Science (Wash. D. C.) 2 2 3 , 1296-1299
Bekoff, M. C., and Klagsbrun, M. (1982) J. Cell. Btochem. 20, 237-245 Ogamo, A,, Matsuzaki, K., Uchiyama, H., and Nagasawa, K. (1982) Car-
Dische, Z. (1947) J. Bid. Chem. 167,189-198 bohydr. Res. 105,69-85
Laemmli, U. K. (1970) Nature (Lond.) 227,680-685 Oakley, B. R., Kirsch, D. R., and Morris, N. R. (1980) And. Biochem. 1 0 5 ,
261 -2KR 16. Gospodarowicz, D., Cheng, D., Lui, G. M., Baird, A., and Bohlent, P. (1984)
17. Gospodarowicz, D. (1975) J. Blol. Chem. 250,2515-2520 18. Miller-Anderson, M., Borg, H., and Andersson, L. 0. (1974) Thromb. Res.
l _ _ _”
Proc. Natl. Acad. Sci. U. S. A. 81,6963-6967
5. 439-452 19. Handin, R. I., and Cohen, H. J. (1976) J. Biol. Chem. 251, 4273-4282 20. Zetter, B. R., Azizkhan, R. A,, Azizkhan, J. C., Brouty-Boye, D., Folkman,
J., Haudenschild, C. C., Klagsbrun, M., Potash, R., and Scheiner, C. J. (1981) in Plasma and Cellular Modulatory Proteins (Bing, D., and Rosen- baum, R. A,, eds) pp. 59-73, Center for Blood Research, Boston, MA
Davidson, J. M., Klagsbrun, M., Hill, K. E., Buckley, A,, Sullivan, R., Brewer, P. S., and Woodward, S. C., manuscript submitted.