purification of a thermostable laccase from leucaena leucocephala using a copper alginate entrapment...
TRANSCRIPT

P
Puo
ND
a
ARRAA
KCCDLL
1
ml[pbHcfmmatnibm
h1
ARTICLE IN PRESSG ModelRBI-10113; No. of Pages 9
Process Biochemistry xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Process Biochemistry
jo ur nal home p age: www.elsev ier .com/ locate /procbio
urification of a thermostable laccase from Leucaena leucocephalasing a copper alginate entrapment approach and the applicationf the laccase in dye decolorization
ivedita Jaiswal, Veda P. Pandey, Upendra N. Dwivedi ∗
epartment of Biochemistry, University of Lucknow, Lucknow 226007, UP, India
r t i c l e i n f o
rticle history:eceived 20 December 2013eceived in revised form 24 March 2014ccepted 2 April 2014vailable online xxx
eywords:elite chromatographyopper alginate
a b s t r a c t
Laccase from a tree legume, Leucaena leucocephala, was purified to homogeneity using a quick two-step procedure: alginate bead entrapment and celite adsorption chromatography. Laccase was purified110.6-fold with an overall recovery of 51.0% and a specific activity of 58.5 units/mg. The purified laccasewas found to be a heterodimer (∼220 kDa), containing two subunits of 100 and 120 kDa. The affinity oflaccase was found to be highest for catechol and lowest for hydroquinone, however, highest Kcat andKcat/Km were obtained for hydroquinone. Purified laccase exhibited pH and temperature optima of 7.0and 80 ◦C, respectively. Mn2+, Cd2+, Fe2+, Cu2+ and Na+ activated laccase while Ca2+ treatment increasedlaccase activity up to 3 mM, beyond which it inhibited laccase. Co2+, Hg2+, DTT, SDS and EDTA showed
ye decolorizationaccaseeucaena leucocephala
an inhibition of laccase activity. The Leucaena laccase was found to be fairly tolerant to organic solvents;upon exposure for 1 h individually to 50% (v/v) each of ethanol, DMF, DMSO and benzene, more than50% of the activity was retained, while in the presence of 50% (v/v) each of methanol, isopropanol andchloroform, a 40% residual activity was observed. The purified laccase efficiently decolorized syntheticdyes such as indigocarmine and congo red in the absence of any redox mediator.
© 2014 Elsevier Ltd. All rights reserved.
. Introduction
Laccases (benzenediol: oxygen oxidoreductase; EC 1.10.3.2) areulticopper oxidases that catalyze the oxidation of various pheno-
ic and non-phenolic aromatic compounds using molecular oxygen1]. They are widely distributed in nature and have been found inlants, insects, bacteria, fungi and crustaceans. Most laccases haveeen isolated and characterized extensively from fungal sources.owever, a considerably lesser number of laccases have beenharacterized from plant sources to date. The laccases reportedrom plants as well as fungi have been purified using traditional
ethods, such as salt or solvent precipitation and column chro-atography, which not only took longer time but also resulted in
considerably lower yield of the purified enzyme [2–4]. Therefore,he industrial application of laccase demands its bulk production,ecessitating the development of efficient, quick and econom-
Please cite this article in press as: Jaiswal N, et al. Purification
a copper alginate entrapment approach and the application of
http://dx.doi.org/10.1016/j.procbio.2014.04.002
cal purification methods. In this study, we report an alginateead entrapment method for Leucaena laccase purification thatay have industrial applications. The alginate bead entrapment
∗ Corresponding author. Tel.: +91 522 2740132; fax: +91 522 2740132.E-mail address: [email protected] (U.N. Dwivedi).
ttp://dx.doi.org/10.1016/j.procbio.2014.04.002359-5113/© 2014 Elsevier Ltd. All rights reserved.
method for the purification of enzymes, directly from crude extract,has recently been used as an attractive, quick and economicalmethod for purifying other enzymes, such as �- and �-amylases,glucoamylase, pectinase, phospholipase and �-galactosidase [5].
Alginate, a copolymer of �-d-mannuronic acid (M) and �-l-guluronic acid (G) residues, has been exploited most commonly forentrapment purposes in the form of calcium alginate beads. How-ever, besides calcium, various other divalent cations have also beenshown to possess an affinity for alginate in the following order:Pb2+ > Cu2+ > Cd2+ > Ba2+ > Sr2+ > Ca2+ > Co2+ > Ni2+ > Zn2+ > Mn2+.Furthermore, the affinity of a metal-alginate bead toward theentrapped enzyme is dependent upon the nature and compositionof the metal as well as its concentration in the bead [6]. Thus, a cal-cium alginate bead will have a higher affinity for a Ca2+-containingenzyme than for another metal-containing enzyme. One suchexample is the purification of �-amylase, a calcium containingmetalloenzyme, using calcium alginate beads [5]. Therefore, thisproperty has been exploited in the present study for purifyinglaccase, a copper containing metalloenzyme, using copper alginate
of a thermostable laccase from Leucaena leucocephala usingthe laccase in dye decolorization. Process Biochem (2014),
beads. Though copper alginate beads have been used for theimmobilization of laccase [7–9], it has not yet been used for thepurification of laccase or any other enzyme, to the best of ourknowledge.

ING ModelP
2 iochem
rthtiati[tsbird
abm[robddssdomtFttfa
fwtrtTtd
2
2
cL
2
[p3t3ct
ARTICLERBI-10113; No. of Pages 9
N. Jaiswal et al. / Process B
Laccases isolated from various sources exhibited diversity withegards to their size and subunit composition. Multimeric, homote-rameric, heterodimeric as well as monomeric forms of laccasesave been reported, ranging in subunit molecular weight from 36o 175 kDa [2,10–12]. Though the pH optima for plant laccases var-ed from neutral to alkaline range [11–13], while most of the fungalnd bacterial laccases exhibited an acidic pH optima [14,15]. Theemperature optima of laccases have been reported to vary widelyn the range of 25–90 ◦C, depending on the source of the laccase16–18]. Fungal laccases usually have a lower thermal stabilityhan bacterial laccases [19]. Recently, a laccase from Trametes hir-uta showed an optimum temperature of 85 ◦C [20]. Laccases haveeen inhibited by a number of ions and compounds, such as metal
ons (e.g., Ca2+, Mg2+, Co2+, Mn2+, Cd2+, Zn2+, Hg2+, etc.), sulfhydryleagents, EDTA, dithiothreitol, hydroxyglycine, kojic acid, thiourea,etergents, etc. [1].
Laccases have applications in various industrial processes, suchs textile dye bleaching, pulp bleaching, effluent detoxification,ioremediation of contaminating environmental pollutants, enzy-atic conversion of chemical intermediates, and organic synthesis
1]. Therefore, for these industrial applications, laccases that areesistant to extreme conditions of pH, temperature, high salt, andrganic solvents, are highly desirable. Furthermore, laccases haveecome an attractive option (in contrast to peroxidases) for theecolorization of synthetic dyes from industrial wastes [21], as theyo not require expensive H2O2 as a co-substrate and have broaderubstrate specificity. In the presence of mediators, the substratepecificity of laccases can be further enhanced, leading to the oxi-ation of more complex substrates [22]. Thus, laccases are capablef oxidizing a wide variety of aromatic compounds, such as ortho-,eta- or para- substituted phenols; diamines; aromatic amines and
hiols; and inorganic compounds, such as iodine, Mo(CN)84−, and
e(CN)64− [1,19]. Several microbial laccases have been assessed for
heir potential application in dye decolorization [23,24], however,he reports on dye decolorization by plant laccases is scanty. There-ore, there is a need to search for potential plant laccases with anbility to degrade dyes.
In the present paper, we describe a rapid purification protocolor laccase from the leaves of Leucaena leucocephala, a tree legumehich is of significance to the fodder and the paper and pulp indus-
ries in India. In addition, purified laccase was characterized withegards to the effects of temperature, pH, substrates, various effec-ors, and organic solvents and the ability to oxidize industrial dyes.o the best of our knowledge, this paper is the first report of a pro-ocol for the purification of a laccase and its effectiveness in dyeecolorization.
. Experimental
.1. Plant material
Fresh green and young leaves from a 5 years old tree of L. leu-ocephala, which was growing in the garden of the University ofucknow, Department of Biochemistry, were used as plant material.
.2. Enzyme assay and protein estimation
Laccase activity was assayed as described by Matijosyte et al.25]. The reaction mixture contained Tris–HCl buffer (100 mM,H 7.0), catechol (10 mM) and a suitable enzyme aliquot. After0 min of incubation at 37 ◦C, the increase in absorbance (due
Please cite this article in press as: Jaiswal N, et al. Purification
a copper alginate entrapment approach and the application of
http://dx.doi.org/10.1016/j.procbio.2014.04.002
o oxidation of catechol to o-benzoquinone) was measured at90 nm using UV–vis spectrophotometer (Elico SL-177). A parallelontrol containing all the ingredients of the assay system, excepthe enzyme, was used as blank and those without substrate
PRESSistry xxx (2014) xxx–xxx
were used as control. Enzyme activity was expressed in terms ofunits. One unit of enzyme activity was defined as the amount ofenzyme required to produce 1 �mol of o-benzoquinone in 1 minunder the specified conditions (ε = 1260 M−1 cm−1). Similarly,enzyme activity using substrates hydroquinone and ABTS, weredetermined by measuring increase in absorbance at 390 nm (ε forp-benzoquinone = 2240 M−1 cm−1), and 420 nm (ε for ABTS+ freeradical = 36,000 M−1 cm−1), respectively.
Protein concentration was estimated by the Bradford dye bind-ing method using bovine serum albumin as the standard [26].
2.3. Extraction from Leucaena leaves
A 30% crude extract was prepared by homogenizing 40 g of Leu-caena leaves in 120 ml of Tris–HCl buffer (100 mM, pH 7.5) using anice-cold blender. Solid PVP (polyvinylpyrrolidone, insoluble; 0.1%(w/v)) and 7 mM �-mercaptoethanol were added at the time ofextraction. The homogenate was centrifuged at 8500 × g for 30 minat 4 ◦C using a Sigma 4K15 centrifuge. The clear supernatant (crudeextract) was subjected to further purification. All of the operationswere performed at 4 ◦C, unless otherwise specified.
2.4. Purification of laccase
2.4.1. Entrapment in copper alginate beadsFor the purification of laccase, a copper alginate-mediated
entrapment (affinity precipitation) method, modified from thatdescribed by Prakash and Jaiswal [5], was used. Crude extract(100 ml) was mixed with 200 ml of sodium alginate (2%, w/v) andkept at 4 ◦C for 30 min with occasional hand swirling. Copper algi-nate beads (containing the entrapped enzyme) were prepared byadding the enzyme–sodium alginate mixture in a drop-wise man-ner to 1 l of a pre-cooled CuSO4 solution (100 mM) with continuousgentle hand swirling. The copper alginate beads formed using thismethod were allowed to harden for 1 h. Afterwards, the beads werewashed twice with Tris–HCl buffer (100 mM, pH 7.5, 200 ml perwash). Entrapped enzymes were eluted from the beads with 100 mlof Tris–HCl buffer (100 mM, pH 7.5) containing 1 M NaCl usingconstant agitation at 100 rpm for 30 min at 30 ◦C. The eluate wascollected by decanting. The eluted enzyme preparation was dia-lyzed overnight against the Tris–HCl buffer (100 mM, pH 7.5) withtwo to three changes. The dialyzed preparation was used for furtherpurification.
2.4.2. Celite adsorption chromatographyTwenty grams of Celite 545 (diatomaceous earth, obtained from
Sigma–Aldrich) was mixed with 250 ml of distilled water, boiled for5 min, and subsequently celite was allowed to settle at room tem-perature. The supernatant was decanted. The celite was washed 3–4times with distilled water to remove the fine particles. The washedcelite was suspended in Tris–HCl buffer (100 mM, pH 7.5) andpacked in a glass column (1.5 cm × 50 cm). The dialyzed enzymepreparation (100 ml) was applied on the top of the celite column.The pass-through fractions (5 ml) containing laccase activity werepooled and concentrated using Centricon with a 50 kDa-molecularmass-cut-off. The concentrated enzyme preparation was stored at4 ◦C for further use.
2.5. Native PAGE and in-gel activity staining
Native polyacrylamide gel-electrophoresis (7.5%; PAGE) was
of a thermostable laccase from Leucaena leucocephala usingthe laccase in dye decolorization. Process Biochem (2014),
performed and visualized using silver staining. In-gel activity stain-ing of the laccase was performed by immersing the gel (after 7.5%native PAGE) in a catechol solution (50 mM) containing Tris–HClbuffer (100 mM, pH 7.5) until brown-colored bands appeared.

ING ModelP
iochem
2
gcuplacelwoov
2
ctTSr(
2
gewaob6sTafnrwiu
2
Mo7ic
2
(flaemc
entrapment method (affinity precipitation) has been exploited suc-cessfully for the first time to purify laccase from a plant source.
To date, laccases isolated from various sources, such as plants,bacteria and fungi, have been purified using traditional multi-step
Fig. 1. (A) Native-PAGE analysis and silver staining of L. leucocephala laccase dur-
ARTICLERBI-10113; No. of Pages 9
N. Jaiswal et al. / Process B
.6. Native molecular weight determination
The native molecular weight of purified laccase was investi-ated by gel filtration chromatography [27] using a Sephadex G-200olumn. Blue dextran was used to determine the unoccupied vol-me (Vo) of the column. Catalase (240 kDa), �-amylase (200 kDa),hosphorylase B (97.4 kDa), bovine serum albumin (67 kDa) and
ysozyme (14.3 kDa), were used as standard proteins (1.0 mg/ml)nd, were applied onto the column. The amount of protein in theolumn eluent was estimated using Bradford’s method [26]. Thelution volume (Ve) of each standard protein as well as the purifiedaccase was measured. The molecular weight of the purified laccase
as calculated from a calibration curve obtained by plotting the logf the molecular weight of the standard proteins against the ratiof the elution volumes of the standard proteins and the unoccupiedolume of the column (Ve/Vo).
.7. Subunit molecular weight determination
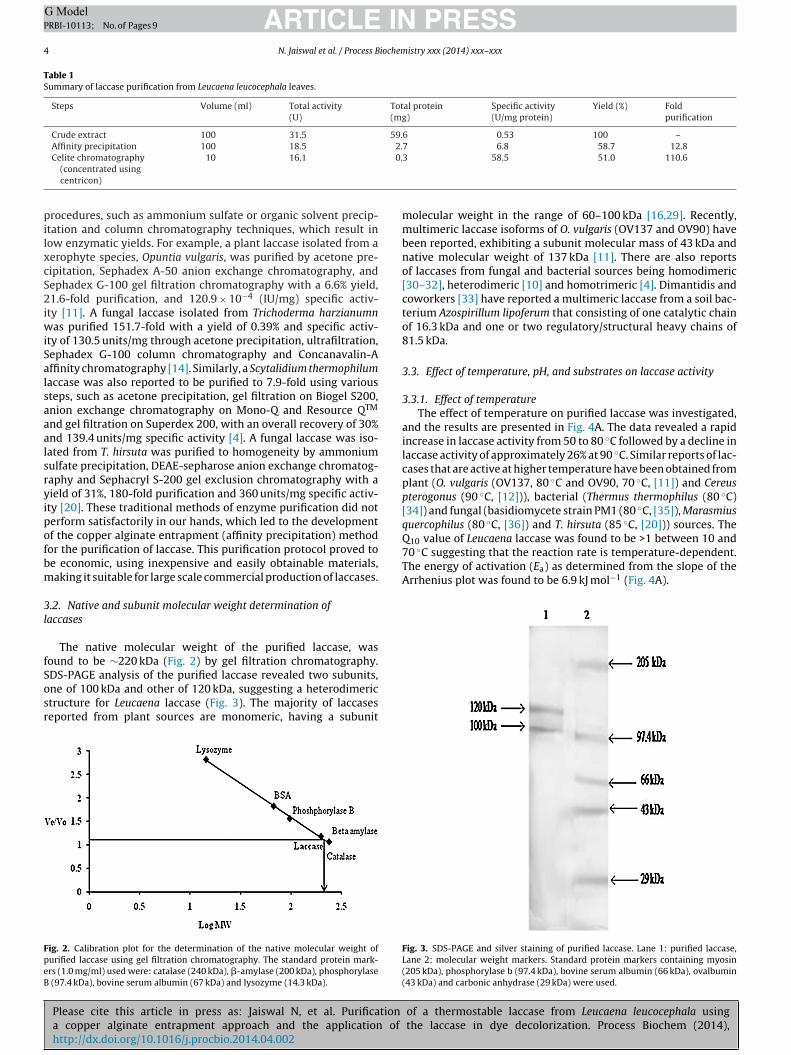
For subunit molecular weight determination, SDS-PAGE wasonducted in a mini electrophoresis chamber (Bio-Rad) at roomemperature using a 7.5% resolving and 3% stacking gel inris–glycine running buffer (pH 8.8) at 100 V for 90 min [28].tandard protein markers containing myosin (205 kDa), phospho-ylase b (97.4 kDa), bovine serum albumin (66 kDa), ovalbumin43 kDa) and carbonic anhydrase (29 kDa) were used.
.8. Effect of temperature, pH and substrates
Laccase activity at various temperatures (10–90 ◦C) was investi-ated by incubating the whole assay system at pH 7.0 as describedarlier in Experimental section. The thermostability of the enzymeas determined by incubating the enzyme at 80 ◦C and then
ssaying the activity at various time intervals. Similarly, the effectf pH on laccase activity was determined using 100 mM of variousuffers at different pH values (such as sodium acetate, pH 6.0 and.5; Tris–HCl, pH 7.0, 7.5, 8.0 and 8.5; sodium borate, pH 9.0 andodium carbonate, pH 9.5) under standard assay conditions at 37 ◦C.he pH stability of the enzyme was determined by estimating thectivity after incubating the enzyme at different pH values (6.0–9.0)or 24 h. The effect of phenolic (catechol and hydroquinone) andon-phenolic (ABTS) substrates was investigated at concentrationsanging from 0 to 50 mM at 37 ◦C and pH 7.0. Km and Vmax valuesere obtained by non-linear regression of a plot of enzyme activ-
ty vs substrate concentration (hyperbolic Michaelis–Menten plot)sing GraphPad Prism software.
.9. Effect of various metal ions, SDS, DTT, and EDTA
The effect of varying concentrations of different effectors (Na+,g2+, Ca2+, Mn2+, Cu2+, Co2+, Fe2+, Cd2+, Hg2+, DTT, SDS, and EDTA)
n laccase activity was studied as described above (at 37 ◦C and pH.0) by performing the activity assay using catechol as the substrate
n the presence and absence of individual effectors at specifiedoncentrations in the reaction mixture.
.10. Effect of organic solvents
The stability of Leucaena laccase in the presence of various polarmethanol, ethanol, isopropanol, dimethyl sulfoxide and dimethyluoride) and non-polar organic solvents (benzene and chloroform)
Please cite this article in press as: Jaiswal N, et al. Purification
a copper alginate entrapment approach and the application of
http://dx.doi.org/10.1016/j.procbio.2014.04.002
t 20% and 50% concentrations (v/v) was studied by incubating thenzyme in the respective solvents for 1 h at 37 ◦C, and subsequentlyeasuring the activity of enzyme aliquots under the standard assay
onditions (at 37 ◦C and pH 7.0).
PRESSistry xxx (2014) xxx–xxx 3
2.11. Effect on dye decolorization
Synthetic industrial dyes, namely indigocarmine (�max 610 nm)and congo red (�max 500 nm), were used for investigating the effi-cacy of decolorization by purified laccase. Stock solutions (1 mg/ml)of these dyes were prepared in distilled water and diluted to therequired concentration and then used for the decolorization assay.The reaction mixture (3 ml) contained Tris–HCl (100 mM, pH 7.0),a dye solution of a specified concentration and an appropriateamount of enzyme. A reaction mixture without enzyme was alsorun. After incubation for 6 h at 37 ◦C, the change in the absorbancewas measured spectrophotometrically using UV–vis spectropho-tometer (Elico SL-177). The effects of varying enzyme amounts(1.4–22 �g) and dye concentrations (10–250 �g/ml) on dye decol-orization were also investigated.
2.12. Statistical analysis
The experiments were performed in triplicates, and the meanand standard deviation were calculated accordingly.
3. Results and discussion
3.1. Purification of laccase
Laccase was purified to homogeneity from the leaves of Leucaenausing copper alginate bead entrapment followed by celite chro-matography (Table 1). The homogeneity of the purified enzyme wasestablished using native PAGE, where a single band was obtained(Fig. 1A). The purified laccase was found to be catalytically activeas established through in-gel activity staining (Fig. 1B). The laccasewas purified to 110.6-fold with an overall recovery of 51.0% anda specific activity of 58.5 units/mg. Thus, the copper alginate bead
of a thermostable laccase from Leucaena leucocephala usingthe laccase in dye decolorization. Process Biochem (2014),
ing purification. Lane 1: crude extract, Lane 2: eluate obtained after salt elutionof the entrapped enzyme from the copper alginate beads, Lane 3: purified laccaseobtained after celite chromatography. (B) In-gel activity staining of purified lac-case. The gel after Native-PAGE was immersed in 50 mM catechol solution until thebrown-colored band appeared.
ARTICLE IN PRESSG ModelPRBI-10113; No. of Pages 9
4 N. Jaiswal et al. / Process Biochemistry xxx (2014) xxx–xxx
Table 1Summary of laccase purification from Leucaena leucocephala leaves.
Steps Volume (ml) Total activity(U)
Total protein(mg)
Specific activity(U/mg protein)
Yield (%) Foldpurification
Crude extract 100 31.5 59.6 0.53 100 –Affinity precipitation 100 18.5 2.7 6.8 58.7 12.8
0.
pilxcS2iwiSalsaaalsryipofbm
3l
fSosr
FpeB
Q10 value of Leucaena laccase was found to be >1 between 10 and70 ◦C suggesting that the reaction rate is temperature-dependent.The energy of activation (Ea) as determined from the slope of theArrhenius plot was found to be 6.9 kJ mol−1 (Fig. 4A).
Celite chromatography(concentrated usingcentricon)
10 16.1
rocedures, such as ammonium sulfate or organic solvent precip-tation and column chromatography techniques, which result inow enzymatic yields. For example, a plant laccase isolated from aerophyte species, Opuntia vulgaris, was purified by acetone pre-ipitation, Sephadex A-50 anion exchange chromatography, andephadex G-100 gel filtration chromatography with a 6.6% yield,1.6-fold purification, and 120.9 × 10−4 (IU/mg) specific activ-
ty [11]. A fungal laccase isolated from Trichoderma harzianumnas purified 151.7-fold with a yield of 0.39% and specific activ-
ty of 130.5 units/mg through acetone precipitation, ultrafiltration,ephadex G-100 column chromatography and Concanavalin-Affinity chromatography [14]. Similarly, a Scytalidium thermophilumaccase was also reported to be purified to 7.9-fold using variousteps, such as acetone precipitation, gel filtration on Biogel S200,nion exchange chromatography on Mono-Q and Resource QTM
nd gel filtration on Superdex 200, with an overall recovery of 30%nd 139.4 units/mg specific activity [4]. A fungal laccase was iso-ated from T. hirsuta was purified to homogeneity by ammoniumulfate precipitation, DEAE-sepharose anion exchange chromatog-aphy and Sephacryl S-200 gel exclusion chromatography with aield of 31%, 180-fold purification and 360 units/mg specific activ-ty [20]. These traditional methods of enzyme purification did noterform satisfactorily in our hands, which led to the developmentf the copper alginate entrapment (affinity precipitation) methodor the purification of laccase. This purification protocol proved toe economic, using inexpensive and easily obtainable materials,aking it suitable for large scale commercial production of laccases.
.2. Native and subunit molecular weight determination ofaccases
The native molecular weight of the purified laccase, wasound to be ∼220 kDa (Fig. 2) by gel filtration chromatography.
Please cite this article in press as: Jaiswal N, et al. Purification
a copper alginate entrapment approach and the application of
http://dx.doi.org/10.1016/j.procbio.2014.04.002
DS-PAGE analysis of the purified laccase revealed two subunits,ne of 100 kDa and other of 120 kDa, suggesting a heterodimerictructure for Leucaena laccase (Fig. 3). The majority of laccaseseported from plant sources are monomeric, having a subunit
ig. 2. Calibration plot for the determination of the native molecular weight ofurified laccase using gel filtration chromatography. The standard protein mark-rs (1.0 mg/ml) used were: catalase (240 kDa), �-amylase (200 kDa), phosphorylase
(97.4 kDa), bovine serum albumin (67 kDa) and lysozyme (14.3 kDa).
3 58.5 51.0 110.6
molecular weight in the range of 60–100 kDa [16,29]. Recently,multimeric laccase isoforms of O. vulgaris (OV137 and OV90) havebeen reported, exhibiting a subunit molecular mass of 43 kDa andnative molecular weight of 137 kDa [11]. There are also reportsof laccases from fungal and bacterial sources being homodimeric[30–32], heterodimeric [10] and homotrimeric [4]. Dimantidis andcoworkers [33] have reported a multimeric laccase from a soil bac-terium Azospirillum lipoferum that consisting of one catalytic chainof 16.3 kDa and one or two regulatory/structural heavy chains of81.5 kDa.
3.3. Effect of temperature, pH, and substrates on laccase activity
3.3.1. Effect of temperatureThe effect of temperature on purified laccase was investigated,
and the results are presented in Fig. 4A. The data revealed a rapidincrease in laccase activity from 50 to 80 ◦C followed by a decline inlaccase activity of approximately 26% at 90 ◦C. Similar reports of lac-cases that are active at higher temperature have been obtained fromplant (O. vulgaris (OV137, 80 ◦C and OV90, 70 ◦C, [11]) and Cereuspterogonus (90 ◦C, [12])), bacterial (Thermus thermophilus (80 ◦C)[34]) and fungal (basidiomycete strain PM1 (80 ◦C, [35]), Marasmiusquercophilus (80 ◦C, [36]) and T. hirsuta (85 ◦C, [20])) sources. The
of a thermostable laccase from Leucaena leucocephala usingthe laccase in dye decolorization. Process Biochem (2014),
Fig. 3. SDS-PAGE and silver staining of purified laccase. Lane 1: purified laccase,Lane 2: molecular weight markers. Standard protein markers containing myosin(205 kDa), phosphorylase b (97.4 kDa), bovine serum albumin (66 kDa), ovalbumin(43 kDa) and carbonic anhydrase (29 kDa) were used.

ARTICLE IN PRESSG ModelPRBI-10113; No. of Pages 9
N. Jaiswal et al. / Process Biochemistry xxx (2014) xxx–xxx 5
Fig. 4. (A) Effect of temperature on the activity of L. leucocephala laccase. The enzyme was incubated at different temperature (10–90 ◦C) and activity was measured unders of L. ler
ic8svbamise
3
oprpiretttt
Flc
tandard assay conditions. The inset shows the Arrhenius plot. (B) Thermal stability
elative to the control (0 min), which was set at 100%.
Time-dependent thermostability of Leucaena laccase was alsonvestigated at 80 ◦C (Fig. 4B). The data revealed that Leucaena lac-ase was activated (by approximately 45%) upon pre-incubation at0 ◦C for up to 50 min, after which there was a decline in activation,uggesting that the enzyme was fairly thermostable. Similar acti-ation upon pre-incubation of laccases at higher temperatures haseen reported from a number of fungal sources [35,37]. The hyper-ctivation of the Leucaena laccase observed in the present studyight be attributed to interactions between the active site copper
ons as well as ionic and hydrogen bonding interactions, leading totabilization of more active conformation, as suggested by Hildent al. [37].
.3.2. Effect of pHThe effect of pH on the purified Leucaena laccase revealed pH
ptima of 7.0 with a steep decline in both acidic as well as basicH ranges (Fig. 5A). Laccase stability was also investigated and itevealed that the enzyme retained more than 80% of its activity atH values ranging from 6.0 to 9.0 within 24 h at 37 ◦C (Fig. 5B). Sim-
lar to our observations, pH optima of approximately 7.0 have beeneported for laccase from R. vernicifera and R. succedanea using cat-chol as a substrate [13]. The pH optimum of the Leucaena laccase in
Please cite this article in press as: Jaiswal N, et al. Purification
a copper alginate entrapment approach and the application of
http://dx.doi.org/10.1016/j.procbio.2014.04.002
he neutral range shows the potential of the enzyme for its applica-ion in the biobleaching industry, as these processes require neutralo alkaline conditions [38]. Recently, laccase isoforms isolated fromwo xerophytic plant species, C. pterogonus and O. vulgaris, have
ig. 5. (A) Effect of pH on the activity of L. leucocephala laccase. The activity was assayeeucocephala laccase. The enzyme was incubated for 24 h at different pH (6.0–9.0) and actonditions. Percent relative activity represents enzyme activity calculated by setting the
ucocephala laccase at 80 ◦C. Percent relative activity represents the enzyme activity
been found to show an optimum pH of 10 [17]. However, bacte-rial and fungal laccases are reported to exhibit acidic pH optima,e.g., A. lipoferum (6.0, [33]), Streptomyces cyaneus and Trametes ver-sicolor (3–5, [39]), Pleurotus ostreatus (3.5, [40]). The Melanocarpesalbomyces laccase exhibited an atypical pH optimum of 7.0 for phe-nolic substrates [41]. Some alkaline laccases have been reportedfrom a fungus, Myrothecium verrucaria (9.0, [42]), and bacteria,Streptomyces coelicolor (9.4, [43]) and Thermobifida fusca (8.0, [44]).
3.3.3. Effect of substrates on laccase activityThe effects of various polyphenolic (catechol and hydroquinone)
and non-phenolic (ABTS) substrates on the purified enzyme wereinvestigated and various kinetic parameters were calculated whichis presented in Table 2. Km and Vmax values were found to be 1.24,1.92, and 6.86 mM and 0.063, 0.088, and 0.351 �M min−1 ml−1,for catechol, ABTS and hydroquinone, respectively. Kcat of theenzyme for these substrates were found to be in the order: hydro-quinone (14.03 min−1) > ABTS (3.53 min−1) > catechol (2.52 min−1).However, the order of catalytic efficiency of the purified lac-caase for the substrates (as presented by Kcat/Km) was foundto be in the order: hydroquinone (2.05 mM−1 min−1) > catechol(2.03 mM−1 min−1) > ABTS (1.83 mM−1 min−1). As evident from the
of a thermostable laccase from Leucaena leucocephala usingthe laccase in dye decolorization. Process Biochem (2014),
Km values, catechol exhibited the highest affinity, while hydro-quinone exhibited the least affinity for laccase. Km of catechol forlaccase from various plant sources has been found to vary widely;for example, Km values of 3.13, 15 and 45 mM have been reported
d at different pH (6.0–9.0) under standard assay conditions. (B) pH stability of L.ivity assayed using a suitable pre-incubated enzyme aliquot under standard assay
activity, at optimum pH, as 100%.

ARTICLE IN PRESSG ModelPRBI-10113; No. of Pages 9
6 N. Jaiswal et al. / Process Biochemistry xxx (2014) xxx–xxx
Table 2Kinetic properties of L. leucocephala laccase with various phenolic and non-phenolic substrates.
Substrates Km (mM) Vmax (�M min−1 ml−1) Kcat (min−1) Kcat/Km (mM−1 min−1)
faeol(vKtso
voi
3
(c
TA
Catechol 1.24 0.063
ABTS 1.92 0.088Hydroquinone 6.86 0.351
or laccases from Amorphophallus campanulatus, Rhus succedaneand R. vernicifera, respectively [13,45]. Thus, the Leucaena laccasexhibited a lower Km value compared with laccases isolated fromther plants. Furthermore, based on the ability of the Leucaenaaccase to efficiently oxidize both o- and p-diphenol substratescatechol and hydroquinone, respectively), the purified enzyme isalidated as a laccase, as suggested by Ferrar and Walker [46]. Thecat value of L. leucocephala laccase for ABTS (3.53 min−1) was lowerhan those found for several other laccases, such as those of T. hir-uta (197 s−1 [20]), Pleurotus pulmonarius (1520 s−1 [31]), and P.streatus (244.32 s−1 [40]).
A comparison of the physico-chemical properties, namely Km forarious substrates, pH, temperature optima, and molecular weight,f L. leucocephala laccase, with those of laccases reported from var-ous plant, bacterial, and fungal sources are summarized in Table 3.
.4. Effect of various metal ions, SDS, DTT, and EDTA
Please cite this article in press as: Jaiswal N, et al. Purification
a copper alginate entrapment approach and the application of
http://dx.doi.org/10.1016/j.procbio.2014.04.002
The effects of different effectors, such as metal ions, detergentsSDS), reducing agents (DTT), and chelating agents (EDTA) on Leu-aena laccase activity were investigated and are shown in Table 4.
able 3 comparison of physico-chemical properties of purified L. leucocephala laccase with othe
Organism Substrates with Km Optimutemp. (
PlantsLeucaena leucocephala Catechol, 1.24 mM
ABTS, 1.92 mMHydroquinone, 6.86 mM
80
Opuntia vulgaris(OV137) 2,6-DMP, 2.2 mM 80
(OV90) 2,6-DMP, 2.2 mM 70
Cereus pterogonus(CP137) 2,6-DMP, 2.1 mM 90
(CP90) 2,6-DMP, 2.1 mM 90
(CP43) 2,6-DMP, 2.1 mM 60
Rhus vernicifera Catechol, 45 mM 40
Rhus succedanea Catechol, 15 mM 50
Morus alba 4-Methylcatechol, 6 mM 45
BacteriaThermus thermophilus – 92
Azospirillum lipoferum Syringaldazine, 34.65 �M –
Streptomyces psammoticus Pyrogallol, 0.25 mMABTS, 0.39 mM
45
Streptomyces ipomoea ABTS, 0.40 mM2,6-DMP, 4.27 mM
60
Thermobifida fusca – 60
FungiTrametes hirsuta ABTS, 0.07 mM
DMP, 0.2 mM85
Trichoderma harzianum ABTS, 180 �MGuaiacol, 60 �M
85
Ganoderma lucidum ABTS, 77 �MGuaiacol, 217 �M
50
Pleurotus pulmonarius ABTS, 210 �MGuaicacol, 550 �M
45
Marasmius quercophilus ABTS, 50 mMSyringaldazine, 7.7 mM
80
Pleurotus ostreatus ABTS, 46.51 mMDMP, 400 mMGuaiacol, 100 mMo-Dianisidine, 23.52 mM
50
2.52 2.033.53 1.83
14.03 2.05
Mn2+, Cd2+, and Na+ activated laccase in a concentration-dependentmanner (0.1–10 mM). However, Mg2+ did not have any effect, whileCa2+ exhibited activation of laccase up to 3 mM, beyond whichit exhibited an inhibition of laccase activity. Fe2+ and Cu2+ acti-vated laccase in a concentration-dependent manner up to 1 mM. Aconcentration-dependent inhibition of laccase activity with Hg2+
and Co2+ up to 1 mM was observed. The effect of metals, suchas Fe2+, Cu2+, Hg2+ and Co2+, on laccase activity at concentra-tions higher than 1 mM could not be determined because of theinterference of high concentrations of these metal salts with colordevelopment. Similar to our results, the addition of Mn2+, Cd2+, Cu2+
and Fe2+ have been reported to increase the activity of laccases fromO. vulgaris [17], P. ostreatus [19], and Streptomyces psammoticus [3].The involvement of four copper ions, distributed at the three dif-ferent copper centers via type-1 (T1) or blue copper center, type-2(T2) or normal copper and type-3 (T3) or coupled binuclear coppercenters, exhibiting characteristic UV/vis and electron paramagnetic
of a thermostable laccase from Leucaena leucocephala usingthe laccase in dye decolorization. Process Biochem (2014),
resonance (EPR) spectra, have been suggested during the catalysisof laccase [1]. Thus, the increase in activity with divalent metal ionsmight be due to their competition with Cu2+ in the electron trans-port system, leading to a positive cooperative relationship between
r reported plant, bacterial and fungal laccases.
m◦C)
OptimumpH
Mol. wt.(kDa)
Ref.
7.0 220 Present study
10.0 137 [11]10.0 90
10.0 137 [12]10.0 9010.0 43
7.0 – [13]7.0 – [13]7.0 62–64 [16]
– 53 [34]6.0 179.3 [33]8.5 43 [3]
5.0 79 [24]
8.0 73.3 [44]
2.4–2.5 90 [20]
4.5 79 [14]
4.5 62 [18]
4.0–5.5 46 [31]
6.2 65 [36]
4.5 68.4 [40]

ARTICLE IN PRESSG ModelPRBI-10113; No. of Pages 9
N. Jaiswal et al. / Process Biochemistry xxx (2014) xxx–xxx 7
Table 4Effect of various effectors on purified laccase activity. Percent relative activity represents enzyme activity relative to control (without any effector) which was taken ashundred percent. The values presented as the mean ± SD of triplicate tests.
Effectors % Relative activity
Concentration (mM)
0 0.1 0.5 1 3 5 10
Mn2+ 100 110 ± 1.7 149 ± 1.5 195 ± 2.0 376 ± 2.2 403 ± 2.0 431 ± 2.5Fe2+ 100 128 ± 1.6 178 ± 1.4 186 ± 1.7 * * *
Cd2+ 100 105 ± 1.5 114 ± 1.5 128 ± 1.8 159 ± 2.4 189 ± 2.2 189 ± 2.2Na+ 100 116 ± 1.3 129 ± 1.7 142 ± 1.7 148 ± 2.0 154 ± 1.5 159 ± 1.6Cu2+ 100 112 ± 1.3 123 ± 1.2 130 ± 1.4 * * *
Ca2+ 100 105 ± 1.8 112 ± 1.3 126 ± 1.6 139 ± 1.8 124 ± 2.0 104 ± 2.1Mg2+ 100 100 100 100 100 100 100Hg2+ 100 85 ± 1.7 66 ± 1.6 50 ± 1.8 * * *
Co2+ 100 70 ± 1.4 52 ± 1.4 37 ± 1.3 * * *
DTT 100 170 ± 1.8 147 ± 1.4 72 ± 2.1 33 ± 1.5 9 ± 1.5 0SDS 100 84 ± 1.5 71 ± 1.8 65 ± 1.7 53 ± 1.6 42 ± 2.3 11 ± 2.0
5 ± 1.8 0 0 0
h the color development during enzyme activity assay and hence the effect could not bem
tapibc
tdcwtlSabuaic[Stictartcp
3
Dsaqttwhzr
Fig. 6. Organic solvent stability of L. leucocephala laccase. The enzyme was incu-bated for 1 h with 20% and 50% (v/v) of organic solvents at 37 ◦C and subsequently
EDTA 100 91 ± 1.8 38 ± 1.5
* salts of Fe2+, Cu2+, Hg2+ and Co2+ at concentrations beyond 1 mM interfered witeasured.
he enzyme and substrate, as suggested by Chao et al. [47]. Thectivation of laccase by Cu2+ may be due to the filling of type-2 cop-er binding sites with Cu2+ ions [14]. The inhibition by the metal
ons Hg2+ and Co2+ have also been reported [14,48], and the reasonehind which might be due to the amino acid residue modifications,onformational changes or copper chelation [49].
DTT activated laccase at lower concentrations, while it inhibitedhe enzyme at concentrations above 0.1 mM in a concentration-ependent manner. SDS inhibited laccase activity at all of theoncentrations tested in a concentration-dependent manner. EDTAas found to strongly inhibit laccase in such a way that at concen-
rations above 1 mM, the enzyme was completely inhibited. It isikely that at lower concentration of DTT, only a limited number of
S bonds is split (may be the only one which supports the enzymective structure), providing the enzyme with some optimal flexi-ility and activity. However, at higher concentrations of DTT thenique three-dimensional conformation of the enzyme might haveffected, thus resulting in an inhibition in enzyme activity [50]. Thenhibition by SDS and EDTA was found almost similar to the lac-ases isolated from peach [29], S. cyaneus [51], and S. psammoticus3]. The disulfide reducing agent, DTT, and the anionic detergent,DS, might have also caused a conformational change in the pro-ein, resulting in the inhibition of the enzyme activity. The strongnhibition of the enzyme by EDTA showed that the purified Leu-aena laccase was highly sensitive to copper chelation, resulting inhe conformational change of the protein and the loss of enzymectivity. It has been reported that the type-2 Cu2+ can be reversiblyemoved from the protein with the chelating reagent EDTA, suchhat the copper-depleted protein is enzymatically inactive, indi-ating that the type-2 Cu2+ of laccase has a functional role in therotein [52].
.5. Effect of organic solvents on the stability of laccase
The effects of various polar (methanol, ethanol, isopropanol,MF, DMSO) and non-polar (benzene and chloroform) organic
olvents on the stability of Leucaena laccase have been investigatednd are presented in Fig. 6. The Leucaena laccase was found to beuite stable in presence of 20% (v/v) of all the organic solventsested. The enzyme was found to retain more than 80% activity inhe presence of all of the organic solvents except isopropanol, in
Please cite this article in press as: Jaiswal N, et al. Purification
a copper alginate entrapment approach and the application of
http://dx.doi.org/10.1016/j.procbio.2014.04.002
hich it retained approximately 60% activity. In the presence ofigher concentrations (50% (v/v)) of ethanol, DMF, DMSO and ben-ene, laccase retained more than 50% activity, while the enzymeetained 42, 37 and 37% activity in the presence of methanol,
the activity was assayed using a suitable aliquot of pre-incubated enzyme understandard assay conditions. Percent residual activity represents the enzyme activityrelative to the control (without any organic solvent), which was taken as 100%.
isopropanol and chloroform, respectively. In the literature, thereare few reports of organic solvent-tolerant laccases from plant,Rhus vernicifera [53]; white-rot fungus, Ganoderma fornicatum[54]; and bacteria, Bacillus licheniformis [47] and T. fusca [44].Laccases tolerant to organic solvents appear to be quite attractivefor their industrial applications, such as in the bioremediationof industrial waters contaminated with organic solvents, and inorganic synthesis and chiral resolution.
3.6. Effect of laccase on dye decolorization
The ability of Leucaena laccase to oxidize industrial dyes wasinvestigated to demonstrate its industrial applicability. The effectof enzyme concentration on the dye decolorization revealedthat with increasing concentration of laccase, decolorization ofindigocarmine (10 �g/ml) and congo red (10 �g/ml) increasedprogressively and the complete decolorization of both the dyeswere achieved within 6 h at 11 and 22 �g of enzyme, respectively(Fig. 7A). The rate of decolorization (oxidation) of the two dyes by
of a thermostable laccase from Leucaena leucocephala usingthe laccase in dye decolorization. Process Biochem (2014),
the Leucaena laccase was also investigated at various dye concen-trations (10–250 �g/ml) and the data are shown in Fig. 7B. Thoughthe rate of dye decolorization was found to increase with dye con-centration but it was not directly proportional to the concentration

ARTICLE IN PRESSG ModelPRBI-10113; No. of Pages 9
8 N. Jaiswal et al. / Process Biochemistry xxx (2014) xxx–xxx
Fig. 7. (A) Effect of L. leucocephala laccase concentration on the decolorization of indigocarmine (10 �g/ml) and congo red (10 �g/ml) dyes. The reaction mixture (3 ml)c t 37 ◦C( ucoce( �g for
oowwo1o(tdtaachaAw
4
mpbrbwidpti
A
aaUiBNEf
[
[
[
[
[
[
[
[
[
[
[
ontaining dyes (30 �g) in Tris–HCl (100 mM, pH 7.5) buffer was incubated for 6 h afor congo red). (B) Rate of decolorization of indigocarmine and congo red dyes by L. le10–250 �g/ml) in the reaction mixture keeping the enzyme amount 11 �g and 22
f the dye. Thus, in case of indigocarmine, the rate of dye decol-rization at a dye concentration of 10 �g/ml was 5.4 �M/min/mghile at 250 �g/ml dye concentration, the rate of decolorizationas 33.8 �M/min/mg. Similarly, in case of congo red, the rate
f dye decolorization at a dye concentration of 10 �g/ml was.81 �M/min/mg while at 250 �g/ml dye concentration, the ratef decolorization was 11.3 �M/min/mg. The rate of decolorizationoxidation) of indigocarmine was about three times faster thanhat of congo red at all dye concentrations. The variation in theecolorization efficiency of the two dyes might be attributed tohe structural variation of the dyes, as suggested by Nyanhongond coworkers [55]. Laccases from fungi, such as Panus rudis [23]nd T. versicolor [15], have been reported to decolorize the indigo-armine dye in the presence of ABTS. The purified Leucaena laccasead an advantage of decolorizing the tested dyes (indigocarminend congo red) without any additional redox mediators, such asBTS. Thus, the Leucaena laccase possesses an additional advantageith regards to its application in textile industries.
. Conclusion
A quick purification protocol using copper alginate bead entrap-ent with celite chromatography has been successfully utilized to
urify laccase to homogeneity. The purified laccase was found toe a heterodimeric protein showing a pH optimum in the neutralange and thermostability up to 80 ◦C. The enzyme was found toe active with both phenolic and non-phenolic substrates. Laccaseas potentially activated by Mn2+, Cd2+, Fe2+, Cu2+ and Na+ and
nhibited by Co2+, Hg2+, DTT, SDS and EDTA, in a concentration-ependent manner. The enzyme was tolerant toward a number ofolar and non-polar organic solvents. The remarkable decoloriza-ion ability of the enzyme suggests that it may have great potentialn the decolorization of effluents of dyes and in textile industries.
cknowledgements
Financial support from UGC, New Delhi, India, in the form of Dr. D. S. Kothari Post doctoral Fellowship to N.J. is gratefullycknowledged. Department of Higher Education, Government ofttar Pradesh, India under the Centre of Excellence in Bioinformat-
cs, Department of Biotechnology, Government of India under the
Please cite this article in press as: Jaiswal N, et al. Purification
a copper alginate entrapment approach and the application of
http://dx.doi.org/10.1016/j.procbio.2014.04.002
IF Scheme, New Delhi and Department of Science and Technology,ew Delhi, under Promotion of University Research and Scientificxcellence (DST-PURSE) program are also gratefully acknowledgedor providing infrastructure facilities.
[
and the change in color was measured at 610 nm (for indigocarmine) and 500 nmphala laccase. The change in color was measured upon varying the dye concentration
indigocarmine and congo red, respectively.
References
[1] Dwivedi UN, Singh P, Pandey VP, Kumar A. Structure–function relationshipamong bacterial, fungal and plant laccases. J Mol Cat B: Enzym 2011;68:117–28.
[2] Ranocha P, McDougall G, Hawkins S, Sterjiades R, Borderies G, Stewartz D, et al.Biochemical characterization, molecular cloning and expression of laccases –a divergent family-in poplar. Eur J Biochem 1999;259:485–95.
[3] Niladevi KN, Jacob N, Prema P. Evidence for a halotolerant alkaline laccasein Streptomyces psammoticus: purification and characterization. Proc Biochem2008;43:654–60.
[4] Younes SB, Sayadi S. Purification and characterization of a novel trimericand thermotolerant laccase produced from the ascomycete Scytalidium ther-mophilum strain. J Mol Catal B: Enzym 2011;73:35–42.
[5] Prakash O, Jaiswal N. A highly efficient and thermostable �-amylase from soy-bean seeds. Biotechnol Appl Biochem 2010;57:105–10.
[6] Smidsrod O. Some physical properties of alginates in solution and in the gelstate. Report No. 34. NTH Trykk: Norwegian Institute of Seaweed Research;1973.
[7] Palmieri G, Giardina P, Desiderio B, Marzullo L, Giamberini M, Sannia G. A newenzyme immobilization procedure using copper alginate gel: application to afungal phenol oxidase. Enzyme Microb Technol 1994;16:151–8.
[8] Brandi P, Annibale AD, Galli C, Gentili P, Pontes ASN. In search for practicaladvantages from the immobilization of an enzyme: the case of laccase. J MolCatal B: Enzym 2006;41:6–69.
[9] Phetsom J, Khammuang S, Suwannawong P, Sarnthima R. Copper-alginateencapsulation of crude laccase from Lentinus polychorus Lev. and their effec-tiveness in synthetic dyes decolorizations. J Biol Sci 2009;9:573–83.
10] Lisov AV, Zavarzina AG, Zavarzin AA, Leontievsky AA. Laccases produced bylichens of the order Peltigerales. FEMS Microbiol Lett 2007;275:46–52.
11] Kumar GN, Srikumar K. Thermophilic laccase from xerophyte species Opuntiavulgaris. Biomed Chromatogr 2011;25:707–11.
12] Kumar GN, Srikumar K. Isolation, purification, and characterization of ther-mophilic laccase from the xerophyte Cereus pterogonus. Chem Nat Comp2012;48:451–6.
13] Omura T. Studies on laccases of lacquer trees. I. Comparison of laccases fromRhus vernicifera and Rhus succedanea. J Biochem 1961;50:264–72.
14] Sadhasivam S, Savitha S, Swaminathan K, Lin F-H. Production, purification andcharacterization of mid-redox potential laccase from a newly isolated Tricho-derma harzianum WL1. Proc Biochem 2008;43:736–42.
15] Wong YX, Yu J. Laccase-catalyzed decolorization of synthetic dyes. Water Res1999;33:3512–20.
16] Kocabas DS, Ogel ZB, Bakir U. Screening of tree leaves as annual renewablegreen biomass for phenol oxidase production and biochemical characterizationof mulberry (Morus alba) leaf phenol oxidases. World J Microbiol Biotechnol2011;27:701–7.
17] Kumar GN, Srikumar K. Molecular and computational approaches to character-ize thermostable laccase gene from two xerophytic plant species. Appl BiochemBiotechnol 2013, http://dx.doi.org/10.1007/s12010-013-0611-6.
18] Sharma KK, Shrivastava B, Sastry VRB, Sehgal N, Kuhad RC. Middle-redox poten-tial laccase from Ganoderma sp.: its application in improvement of feed formonogastric animals. Sci Rep 2013, http://dx.doi.org/10.1038/srep01299.
19] Baldrian P. Fungal laccases-occurrence and properties. FEMS Microbiol Rev2006;30(2):215–42.
20] Haibo Z, Yinglong Z, Feng H, Peiji G, Jiachuan C. Purification and characterization
of a thermostable laccase from Leucaena leucocephala usingthe laccase in dye decolorization. Process Biochem (2014),
of a thermostable laccase with unique oxidative characteristics from Trameteshirsuta. Biotechnol Lett 2009;31:837–43.
21] Abadulla E, Tzanov T, Costa S, Robra KH, Paulo AC, Gubitz GM. Decolorizationand detoxification of textile dyes with a laccase from Trametes hirsuta. ApplEnviron Microbiol 2000;66:3357–62.

ING ModelP
iochem
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[a thermostable, pH-stable and organic solvent-tolerant Ganoderma fornicatum
ARTICLERBI-10113; No. of Pages 9
N. Jaiswal et al. / Process B
22] Bourbonnais R, Paice M. Oxidation of non-phenolic substrates. An expandedrole for laccase in lignin biodegradation. FEBS Lett 1990;267:99–102.
23] Zhang M, Wu F, Wei Z, Xiao Y, Gong W. Characterization and decolorizationability of a laccase from Panus rudis. Enzyme Microbiol Technol 2006;39:92–7.
24] Molina-Guijarro JM, Perez J, Munoz-Dorado J, Guillen F, Moya R, Hernandez M,et al. Detoxification of azo dyes by a novel pH-versatile, salt-resistant laccasefrom Streptomyces ipomoea. Int Microbiol 2009;12:13–21.
25] Matijosyte I, Isabel WCEA, Simon de V, Roger AS. Preparation and use ofcross linked enzyme aggregates (CLEAs) of laccases. J Mol Cat B: Enzym2010;62:142–8.
26] Bradford MM. Rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principle of protein-dye binding. AnalBiochem 1976;72:248–54.
27] Andrews P. Estimation of the molecular weights of proteins by sephadex gel-filtration. Biochem J 1964;91:222–33.
28] Laemmli UK. Cleavage of structural proteins during the assembly of the headof bacteriophage T4. Nature 1970;227:680–5.
29] Alba CM, de Forchetti SM, Horacio AT. Phenoloxidases of peach (Prunus persica)endocarp: its relationship with peroxidases and lignifications. Physiol Plant2000;109:382–7.
30] Min KL, Kim YH, Kim YW, Jung HS, Hah YC. Characterization of a novel laccaseproduced by the wood-rotting fungus Phellinus ribis. Arch Biochem Biophys2001;392:279–86.
31] De Souza CGM, Peralta RM. Purification and characterization of the main laccaseproduced by the white rot fungus Pleurotus pulmonarius on wheat bran solidstate medium. J Basic Microbiol 2003;43:278–86.
32] Ng TB, Wang HX. A homodimeric laccase with unique characteristics fromthe yellow mushroom Cantharellus cibarius. Biochem Biophys Res Commun2004;313:37–41.
33] Diamantidis G, Effosse A, Potier P, Bally R. Purification and characterization ofthe first bacterial laccase in the rhizospheric bacterium Azospirillum lipoferum.Soil Biol Biochem 2000;32:919–27.
34] Miyazaki K. A hyperthermophilic laccase from Thermus thermophilus HB27.Extremophiles 2005;9:415–25.
35] Coll PM, Fernandez-Abalos JM, Villaneuva JR, Santamaria R, Perez P. Purificationand characterization of a phenoloxidase (laccase) from the lignin-degradingbasidiomycete PM1 (CECT 2971). Appl Environ Microbiol 1993;59:2607–13.
36] Farnet AM, Criquet S, Pocachard E, Gil G, Ferre E. Purification of a new isoformof laccase from a Marasmius quercophilus strain isolated from a cork oak litter(Quercus suber L.). Mycologia 2002;94:735–40.
37] Hilden K, Hakala TK, Lundell T. Thermotolerant and thermostable laccases.Biotechnol Lett 2009;31:1117–28.
38] Heinzkill M, Bech L, Halkier T, Schneider P, Anke T. Characterization of laccases
Please cite this article in press as: Jaiswal N, et al. Purification
a copper alginate entrapment approach and the application of
http://dx.doi.org/10.1016/j.procbio.2014.04.002
and peroxidases from wood-rotting fungi (family Coprinaceae). Appl EnvironMicrobiol 1998;64:1601–6.
39] Margot J, Bennati-Granier C, Maillard J, Blánquez P, Barry DA, Holliger C. Bac-terial versus fungal laccase: potential for micropollutant degradation. AMBExpress 2013;3:63.
[
PRESSistry xxx (2014) xxx–xxx 9
40] Patel H, Gupte S, Gahlout M, Gupte A. Purification and characterization ofan extracellular laccase from solid-state culture of Pleurotus ostreatus HP-1.3 Biotech 2013, http://dx.doi.org/10.1007/s13205-013-0129-1.
41] Kiiskinen L-L, Viikari L, Kruus K. Purification and characterization of a novellaccase from the ascomycete Melanocarpus albomyces. Appl Microb Biotechnol2002;59:198–204.
42] Sulistyaningdyah WT, Ogawa J, Tanaka H, Maeda C, Shimizu S. Characteriza-tion of alkaliphilic laccase activity in the culture supernatant of Myrotheciumverrucaria 24G-4 in comparison with bilirubin oxidase. FEMS Microbiol Lett2004;230:209–14.
43] Machczynski MC, Vijgenboom E, Samyn B, Canters GW. Characterization ofSLAC: a small laccase from Streptomyces coelicolor with unprecedented activity.Protein Sci 2004;13:2388–97.
44] Chen C-Y, Huang Y-C, Wei C-M, Meng M, Liu W-H, Yang C-H. Properties of thenewly isolated extracellular thermo-alkali-stable laccase from thermophilicactinomycetes, Thermobifida fusca and its application in dye intermediates oxi-dation. AMB Express 2013;3:49.
45] Paranjpe PS, Karve MS, Padhye SB. Characterization of tyrosinase and accom-panying laccase from Amorphophallus campanulatus. Ind J Biochem Biophys2003;40:40–5.
46] Ferrar PH, Walker JRL. Inhibition of diphenol oxidases: a comparative study. JFood Biochem 1996;20:15–30.
47] Chao L, Lin C, Rui L, Yanru L, Guoji D. Effect of common metal ions on the rateof degradation of 4-nitrophenol by a laccase-Cu2+ synergistic system. J EnvironManag 2012;113:1–6.
48] Forootanfar H, Faramarzi MA, Shahverdi AR, Yazdi MT. Purification andbiochemical characterization of extracellular laccase from the ascomyceteParaconiothyrium variabile. Bioresour Technol 2011;102:1808–14.
49] Gianfreda L, Xu F, Bollag J-M. Laccases: a useful group of oxidoreductiveenzymes. Bioremed J 1999;3:1–25.
50] Mozhaev VV. Engineering stability of enzymes in systems with organic solvents.In: Ballesteros A, Plou FJ, Iborra JL, Halling PJ, editors. Stability and stabilizationof biocatalysts. Amsterdam: Elsevier; 1998. p. 355–63.
51] Arias ME, Arenas M, Rodriguez J, Soliveri J, Ball AS, Hernandez M. Kraft pulpbiobleaching and mediated oxidation of a nonphenol substrate by laccase fromStreptomyces cyaneus CECT3335. Appl Environ Microbiol 2003;69:1953–8.
52] Li J, McMillin DR. The removal of the type-2 copper from Rhus vernicifera laccase.Biochim Biophys Acta 1992;1160:239–45.
53] Wan Y-Y, Lu R, Xiao L, Du Y-M, Miyakoshi T, Chen C-L, et al. Effects of organicsolvents on the activity of free and immobilized laccase from Rhus vernicifera.Int J Biol Macromol 2010;47:488–95.
54] Huang W-T, Tai R, Hseu R-S, Huang C-T. Overexpression and characterization of
of a thermostable laccase from Leucaena leucocephala usingthe laccase in dye decolorization. Process Biochem (2014),
laccase in Pichia pastoris. Proc Biochem 2011;46:1469–74.55] Nyanhongo GS, Gomes J, Gubitz GM, Zvauya R, Reed JS, Steiner W. Decoloriza-
tion of textile dyes by laccase from a newly isolated strain of Trametes modesta.Water Res 2002;36:1449–56.