prolonged recovery of ultraviolet b-irradiated skin in neuropsin (klk8)-deficient mice
TRANSCRIPT

Cutaneous Biology
Prolonged recovery of ultraviolet B-irradiated skin in neuropsin(KLK8)-deficient mice
T . K I R I H A R A , K . M A T S U M O T O - M I Y A I , Y . N A K A M U R A , T . S A D A Y A M A ,
S . Y O S H I D A * A N D S . S H I O S A K A
Division of Structural Cell Biology, Nara Institute of Science and Technology, 8916-5, Takayama, Ikoma, Nara, 630-0101, Japan
*Department of Anatomy, Asahikawa Medical College, Asahikawa, Japan
Accepted for publication 25 November 2002
Summary Background Neuropsin (KLK8), a serine protease of the kallikrein family, is thought to be involved
in the function of keratinocytes, i.e. migration, differentiation and desquamation. However, how
neuropsin participates is still unknown.
Objective To observe the epidermal function of serine protease in neuropsin-deficient mice.
Methods We irradiated the skin of neuropsin-deficient mice with ultraviolet light to induce acute
inflammation and compared the morphology with that of wild-type mice.
Results We observed a phenotypic change in the epidermis. An acute inflammatory dose of
ultraviolet light induced a marked increase in neuropsin mRNA expression in the skin. The signal
intensity of the mRNA expression was highest on day 2–3 after irradiation, when keratinocytes
were aligned irregularly in the recovery period. Morphological comparison between neuropsin – ⁄ –and + ⁄ + mice revealed that an irregular alignment of cells in the thickened epidermis was obvious
on day 2 after irradiation in the wild-type mice, whereas it was prolonged for at least 2 days in the
neuropsin-deficient mice. The stratum corneum of neuropsin-deficient mice was remarkably thicker
than that of the wild-type mice at 5, 14 and 21 days after irradiation. The increase, as a response to
this stimulus, in involucrin immunoreactivity, a marker for cell envelope assembly, was delayed in
the mutant mice.
Conclusions Thus, neuropsin might be involved early in the process of differentiation, such as in
the assembly of the cell envelope, but not in migration and desquamation.
Key words: epidermis, kallikrein family, keratinocyte, neuropsin, serine protease
The kallikreins constitute a subfamily of secreted serine
proteases of about 230 amino acid residues. There is a
large kallikrein multigene containing 13 different
kallikrein genes in rats,1,2 26 in mice,3,4 and at least
15 in humans.5,6 Some members of the kallikrein
family have been postulated to be involved in the
processing of precursor protein for physiologically
active proteins or peptides.4 However, recent studies
suggest that members of the family are involved not
only in the activation and processing of such precursor
proteins, but also in a variety of cellular events such as
neural plasticity,7 the terminal differentiation of kera-
tinocytes,8 and the regulation of myelin turnover.9
Neuropsin (KLK8 in mouse and human; also known
as human TADG-14 ⁄ ovasin and rat BSP-1) was first
cloned from the mouse brain by us in 199510 and since
has been shown to belong to the kallikrein multigene
family11,12 and to be involved in a variety of functions
in the brain, epidermis and uterus.7,8,13–16 Previous
studies suggested that epidermal neuropsin was impli-
cated in cornification or desquamation, because it is
related to the pathogenesis of hyperkeratosis upon
inflammation,8 and is seen in hyperkeratotic epidermal
diseases, such as psoriasis vulgaris, seborrhoeic kera-
tosis, lichen planus and squamous cell carcinoma.15
This notion is fully consistent with the observation thatCorrespondence: Dr Sadao Shiosaka.
E-mail: [email protected]
British Journal of Dermatology 2003; 149: 700–706.
700 � 2003 British Association of Dermatologists

there was no or only a weak expression of neuropsin
mRNA in the skin of individuals with basal cell
carcinoma.15 To examine neuropsin’s still unknown
function, we produced neuropsin-deficient mice and
studied them by irradiating their skin with ultraviolet
(UV) B light to induce acute inflammation. We then
compared the morphology with that of wild-type mice.
Here we observed a phenotypic change in the epidermis
of neuropsin-deficient mice and found that neuropsin is
involved in an earlier process of the terminal differen-
tiation rather than following cornification and desqu-
amation.
Materials and methods
Generation of neuropsin-deficient mice
Neuropsin-deficient mice were produced as described
elsewhere.17 Briefly, the complementary DNA sequence
of neuropsin was used to isolate genomic clones from a
kFIXII mouse 129 ⁄ Sv genomic library (Stratagene, La
Jolla, CA, U.S.A.). The targeting vector was constructed
by replacing the genomic sequences encoding exons
1–3 with a neomycin-resistance cassette. The ES cells
(1 · 107) were transfected by electroporation (Bio-Rad
Gene Pulsar; 230 V, 500 lF, 0Æ4 cm cuvette, Biorad,
Hercules, CA, U.S.A.) with a NotI-linearized targeting
construct, placed on feeder layers and then subjected to
positive–negative selection for 8 days in 400 lg mL)1
of G418 (Geneticin, Life Technologies Inc., Rockville,
MD, U.S.A.) and 2 lmol L)1 ganciclovir (a gift from
Syntex Incorporated, Tokyo, Japan). The generation of
chimeras and mutant mice was performed essentially
as described previously. The initial chimeric mice were
mated with C57BL ⁄ 6J. Three clones yielded heterozy-
gotes. Heterozygous F1 animals from 129 ⁄ Sv (foun-
der) · C57BL ⁄ 6J have been backcrossed to C57BL ⁄ 6J
animals through more than seven generations. All
animals used in the present study were treated in
accordance with the institutional guidelines for animal
welfare (Animal Centre Guidelines of the Nara Institute
of Science and Technology).
In situ hybridization histochemistry
Forty-two C57BL ⁄ 6J mice (three mice for each time
point; 7-week-old; Japan SLC, Shizuoka, Japan)
were used for in situ hybridization histochemistry to
detect neuropsin mRNA. The histochemistry was
performed according to Chen et al.10,18 and Kitayoshi
et al.8 Briefly, the sections were fixed for 20 min with
4% formaldehyde in 0Æ1 mol L)1 phosphate buffer
(pH 7Æ4), washed twice in phosphate buffer, and then
treated with 10 mg mL)1 of protease K (Roche Diag-
nostic GmbH, Mannheim, Germany) in 50 mmol L)1
Tris–HCl (pH 7Æ4) and 5 mmol L)1 ethylenediamine
tetraacetic acid. The sections were fixed again for
20 min as above and acetylated with 0Æ25% acetic
anhydride in 0Æ1 mol L)1 triethanolamine for 10 min.
After dehydration through an ascending alcohol
series, the sections were hybridized with 35S-labelled
cRNA probes10 at 55 �C overnight. The sections were
washed at 65 �C in 50% formamide, 2 · saline
sodium citrate buffer, and 5 mmol L)1 dithiothreitol.
Subsequently, they were treated with RNase A then
washed again at high stringency. After dehydration
through an ascending alcohol series, they were
immersed in Kodak NTB2 emulsion and exposed for
3 weeks.
Quantification of hybridization signals
Hybridization signals were quantified with randomly
selected areas that contain 10 consecutive cells of the
stratum granulosum under a bright-field microscope.
Forty-two areas in total (three sections; one section
per animal) were counted for each time point. The
specificity of the hybridization signals was checked
with a sense hybridization probe. The average grain
density of the background in the stratum granulosum
was 0Æ44 ± 0Æ13 grains per cell and the value was
almost identical to the silver grains on the glass slide
using an anti-sense hybridization probe (Fig. 1a-
0–21). Areas with a grain density at least four times
that of the background were counted as positive. The
number of silver grains per cell was indicated
(Fig. 2).
Ultraviolet B irradiation and histology
Each of 24 neuropsin – ⁄ – and + ⁄ + mice (three mice for
each time point; 8–11 weeks old) was used to examine
the histology of UVB-irradiated skin. A bank of six
fluorescent sunlamps (ATTO, DT-20CMP, Tokyo,
Japan), which emit rays of between 280 and 320 nm
with a peak at 312 nm, was used. After the dorsal skin
was shaved, an acute (54 s) single exposure of UVB
(360 J cm)2) was irradiated on to the dorsal skin of the
mice according to several studies.19–21 Nonirradiated
(control ¼ 0 days) or UVB-irradiated skin from mice
which had survived 1, 2, 3, 5, 7, 15 and 21 days after
irradiation, was cryostat-sectioned 10 lm thick,
N E U R O P S I N ( K L K 8 ) I N U V B - I R R A D I A T E D S K I N 7 0 1
� 2003 British Association of Dermatologists, British Journal of Dermatology, 149, 700–706

Figure 1. Time course of the changes in neuropsin mRNA expression and epidermal morphology after ultraviolet (UV) irradiation. UV irradiation
induced a marked increase in hybridization signals for neuropsin mRNA in the skin of C57BL ⁄ 6J (C57) mice. Skin samples were prepared at 0
(nonirradiated control ¼ a-0) and 1, 2, 3, 5, 7 or 21 days after UV irradiation (a-1, a-2, a-3, a-5, a-7 or a-21, respectively). UV induced
morphological changes were compared between wild-type [NP + ⁄ +; b-0–21] and neuropsin-deficient [NP – ⁄ –; c-0–21] mice over the same period.
Healing was apparently delayed in the NP – ⁄ – mouse skin. Scale bar ¼ 50 lm. White lines represent the surface of the epidermis. Abbreviations:
d, dermis; ep, epidermis; sb, stratum basale; sc, stratum corneum; sg, stratum granulosum; ss, stratum spinosum.
7 0 2 T . K I R I H A R A et al.
� 2003 British Association of Dermatologists, British Journal of Dermatology, 149, 700–706
mounted on glass slides, and stored at )80 �C until use.
Two sets of serial tissue sections were produced, one
was subjected to haematoxylin–eosin staining for
morphological observation and the other to immunoh-
istochemistry for involucrin, a marker protein of
cornified envelope.
Immunohistochemistry for involucrin and neuropsin
The immunofluorescent techniques used for involucrin
and neuropsin were in accord with Ishida-Yamamoto
and Iizuka22 and Inoue et al., respectively.14 Six
micrometre thick cryostat sections of frozen samples
were air-dried and fixed with acetone at 4 �C for
10 min. After preincubation with 5% bovine serum
albumin in phosphate-buffered saline (PBS) for 1 h at
room temperature, the sections were incubated with
anti-involucrin antibody (Berkeley Antibody Company,
Richmond, CA, U.S.A.; 1 : 1000) or antineuropsin
monoclonal antibody (MabB5, MBL Co., Nagoya,
Japan; 1 : 1000) diluted with 5% bovine serum albu-
min in PBS overnight at 4 �C. After a wash, the
sections were incubated with fluorescein isothiocya-
nate (FITC)-labelled antibody (TAGO, Camarillo, CA,
U.S.A.) diluted with PBS (1 : 1500) overnight at 4 �C.
Fluorescent images were acquired by computer
through a charged couple device (CCD) camera
(Hamamatsu Photonics, Hamamatsu, Japan) and ana-
lysed using WinRoof v3.3 image analysis software
(Mitani Co., Fukui, Japan).
Data analyses
The unpaired t-test was performed for statistical ana-
lyses using StatViewJ-5Æ0 software (SAS institute, Cary,
NC, U.S.A.).
Results
Brief irradiation of shaved dorsal skin with UVB light
induced a transient increase in neuropsin mRNA
expression in the epidermis of the C57BL ⁄ 6J mouse
(Fig. 1a-1–7). The expression of the hybridization
signal was still low at 1 day after irradiation (Fig. 1a-1)
but became prominent at 2 days (Fig. 1a-2), when the
keratinocytes aligned irregularly, and peaked at 3 days
(Fig. 1a-3). Thereafter, the signal gradually faded
(Fig. 1a-5,7) with a return to the control level 21 days
after irradiation (Fig. 1a-21, cf. Fig. 1a-0). A low level
of neuropsin mRNA is constitutively expressed in
normal skin (Fig. 1a-0).8,14 There was no change of
signal expression in the skin of the control group of
mice which were shaved and survived without irradi-
ation (control group).
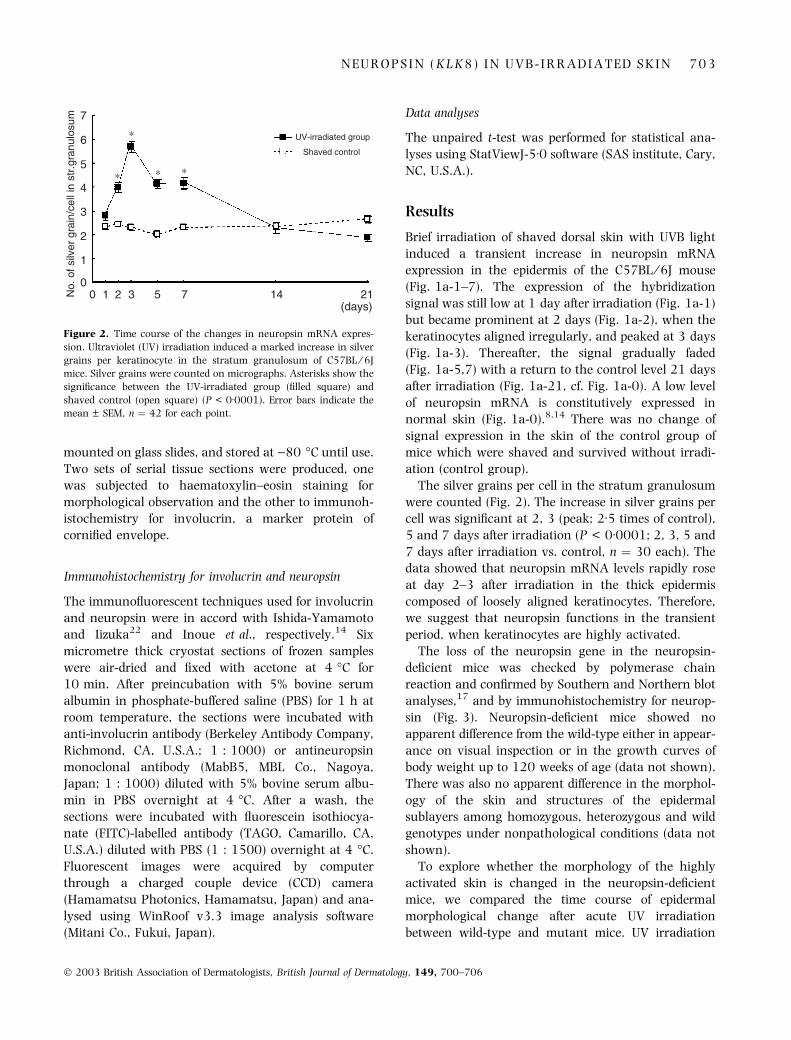
The silver grains per cell in the stratum granulosum
were counted (Fig. 2). The increase in silver grains per
cell was significant at 2, 3 (peak; 2Æ5 times of control),
5 and 7 days after irradiation (P < 0Æ0001; 2, 3, 5 and
7 days after irradiation vs. control, n ¼ 30 each). The
data showed that neuropsin mRNA levels rapidly rose
at day 2–3 after irradiation in the thick epidermis
composed of loosely aligned keratinocytes. Therefore,
we suggest that neuropsin functions in the transient
period, when keratinocytes are highly activated.
The loss of the neuropsin gene in the neuropsin-
deficient mice was checked by polymerase chain
reaction and confirmed by Southern and Northern blot
analyses,17 and by immunohistochemistry for neurop-
sin (Fig. 3). Neuropsin-deficient mice showed no
apparent difference from the wild-type either in appear-
ance on visual inspection or in the growth curves of
body weight up to 120 weeks of age (data not shown).
There was also no apparent difference in the morphol-
ogy of the skin and structures of the epidermal
sublayers among homozygous, heterozygous and wild
genotypes under nonpathological conditions (data not
shown).
To explore whether the morphology of the highly
activated skin is changed in the neuropsin-deficient
mice, we compared the time course of epidermal
morphological change after acute UV irradiation
between wild-type and mutant mice. UV irradiation
7
6
5
4
3
2
1
0
No.
of s
ilver
gra
in/c
ell i
n st
r.gr
anul
osum
0 1 2 3 5 7 14 21(days)
Shaved control
UV-irradiated group*
* * *
Figure 2. Time course of the changes in neuropsin mRNA expres-
sion. Ultraviolet (UV) irradiation induced a marked increase in silver
grains per keratinocyte in the stratum granulosum of C57BL ⁄ 6J
mice. Silver grains were counted on micrographs. Asterisks show the
significance between the UV-irradiated group (filled square) and
shaved control (open square) (P < 0Æ0001). Error bars indicate the
mean ± SEM, n ¼ 42 for each point.
N E U R O P S I N ( K L K 8 ) I N U V B - I R R A D I A T E D S K I N 7 0 3
� 2003 British Association of Dermatologists, British Journal of Dermatology, 149, 700–706

induced evident morphological changes in the wild-
type mice (Fig. 1b-0–21) consistent with the findings
previously published.23,24 Proliferated cells were
loosely and irregularly aligned in the stratum spinosum
and granulosum 2 days after UV irradiation (Fig. 1b-2)
and parakeratosis was also extensive in the stratum
corneum in this period (Fig. 1b-2, white arrows). The
epidermis became thickest 3 days after irradiation and
the parakeratosis disappeared (Figs 1b-3 and 3a). The
prickle and granular cells were aligned in an orderly
manner in close contact with adjacent cells in this
period. However, neuropsin-deficient mice exhibited
different morphological changes during UV-induced
skin injury and recovery (Fig. 1c-2–5). The acute
morphological changes prior to 2 days after irradi-
ation were identical in wild-type and mutant mice
(Fig. 1b-1,2, 1c-1,2). However, the period in which
keratinocytes were aligned loosely was evidently pro-
longed in the neuropsin-deficient mice (Fig. 1c-2,3).
Parakeratosis was not only more severe, but also lasted
much longer in the mutant than the wild-type mice
(Fig. 1c-2,3, white arrows, cf. Fig. 1b-2,3). Thus, the
results strongly suggested that neuropsin is involved in
the dynamic period, when cells are actively aligning
and assembling prior to cornification.
To examine this issue further, the thickness of the
entire epidermis (stratum basale, stratum granulosum,
stratum spinosum and stratum corneum) was meas-
ured and compared between wild-type and mutant
mice (Fig. 4a; cf. Figs 1b-0–21 and 1c-0–21). The
highest values for epidermal thickness both in wild-type
and in neuropsin-deficient mice were obtained at
3 days after irradiation (Fig. 4a). The decrease there-
after was significantly faster at 5–7 days postirradia-
tion in the mutant mice (Fig. 4a, day 5–7; P < 0Æ02 at
day 7 and 14; wild vs. mutant). The thickness of the
stratum corneum differed significantly between the two
(Fig. 4b). The stratum corneum of neuropsin-deficient
mice was remarkably thicker at 5, 14 and 21 days
after the irradiation (Fig. 4b, P < 0Æ001; P < 0Æ001,
P < 0Æ02, respectively; cf. Fig. 1b-5 and 1c-5, 1b-21,
1c-21; wild vs. mutant). The remaining three layers
(stratum basale, stratum granulosum and stratum
spinosum) did not change significantly except on
day 7 after the irradiation, when they were thinner
(P < 0Æ005). This may have been caused by a delay of
differentiation in these layers. Therefore, desquamation
in the mutant might be completely impaired at least up
to day 21.
To analyse the function of neuropsin further, invo-
lucrin, a marker for the initiation of cell envelope
assembly,25 was immunostained to compare mutant
with wild-type mouse skin (Fig. 4c). Involucrin immu-
noreactivity transiently increased and reached a peak
3 days after irradiation in the wild-type mice (Fig. 4c, a
black arrow). However, the peak of the transient
increase of immunoreactivity was markedly delayed
in the mutant mice (Fig. 4c, grey arrow; P < 0Æ05 at
day 3 and P < 0Æ01 at day 7, wild type vs. mutant).
Discussion
This study clearly demonstrated that the skin of
neuropsin-deficient mice was more vulnerable to an
acute inflammatory dose of UVB than that of wild-type
mice. A deficiency of neuropsin caused marked pro-
longation of the recovery period after UVB irradiation,
i.e. thickening of the entire epidermis, thickening of the
Figure 3. Confirmation of deficiency of neuropsin. The deficiency of
neuropsin was confirmed with immunohistochemistry using
antineuropsin antibody. No immunofluorescence was seen in back
skin of the neuropsin (NP) – ⁄ – mouse (a), although clear fluores-
cence (white arrow) appeared in the epidermis of the NP + ⁄ + mouse
(b). Asterisks in (a) and (b) indicate auto fluorescence of the hair.
Scale bar ¼ 50 lm, ep; epidermis.
7 0 4 T . K I R I H A R A et al.
� 2003 British Association of Dermatologists, British Journal of Dermatology, 149, 700–706

stratum corneum and transient irregular alignment of
cells. The increase in involucrin immunoreactivity as a
response to stimulus was delayed in the mutant. As
involucrin is an essential protein for initiating assembly
of the cornified cell envelope, this delay in the increase
in immunoreactive involucrin after UVB irradiation
represents the prolongation of epidermal differentiation
in the mutant. Therefore, neuropsin might be involved
early in the process of differentiation, such as during
the assembly of the cell envelope in the stratum
spinosum and granulosum, by cleaving matrix or cell
adhesion protein(s) that organize the epidermal struc-
ture.26,27 In contrast, neuropsin might not be involved
in the proliferation and migration of keratinocytes,
because there was no or only a weak expression of
neuropsin mRNA in the skin of an individual with
basal cell carcinoma15 and the migrating epithelial
tongue during incisional wounding.8 All together,
neuropsin might be involved in the early process of
differentiation, such as in the assembly of the cell
envelope, but not in migration and desquamation.
Acknowledgments
The authors thank Dr A.Ishida-Yamamoto (Depart-
ment of Dermatology, Asahikawa Medical College) for
critical reading of the manuscript. This work was
supported in part by a grant from the Ministry of
Science, Sport, Culture and Technology.
References
1 Chao L, Gerald W, Chao J. Characterization of rat kallikrein-like
multigene family and its expression in the submandibular gland.
Adv Exp Med Biol 1986; 198: 189–94.
Figure 4. Time course of epidermal morphological change after UV
irradiation. (a) Comparison of changes in the thickness of the epi-
dermis (lm) between neuropsin (NP) + ⁄ + and NP – ⁄ – mice. In the
NP + ⁄ + mouse, the decrease after a transient increase in epidermal
thickness was smoother than that in the NP – ⁄ – mouse. Asterisks
indicate significance at that time point between NP + ⁄ + and NP – ⁄ –
mice (P < 0Æ05). (b) Comparison of changes in thickness of the
stratum corneum between NP + ⁄ + and NP – ⁄ – mice. The stratum
corneum was significantly thicker in the NP – ⁄ – mice than NP + ⁄ +
mice as represented by asterisks (P < 0Æ001 at day 5 and 14, and
P < 0Æ02 at day 21). (c) Comparison of changes in combined thick-
ness of the stratum basale to granulosum between NP + ⁄ + and
NP – ⁄ – mice. The strata were significantly thinner in the NP – ⁄ –
mice than NP + ⁄ + mice only at day 7 as represented by asterisks
(P < 0Æ005). (d) Comparison of the relative content of
immunoreactive involucrin between NP + ⁄ + and NP – ⁄ – mice.
Relative contents were measured using a charged couple device
(CCD) camera and the image analysed as the total intensity of
fluorescein isothiocyanate labelled involucrin in each tissue section
(Materials and methods). The greatest density of immunoreactivity
was seen at 3 days after the irradiation in NP + ⁄ + mice, whereas it
was delayed to 5 days after the irradiation in NP – ⁄ – mice. Asterisks
show the significance between density at 3 days (P < 0Æ05) and
5 days (P < 0Æ01) after the irradiation in NP + ⁄ + and NP – ⁄ – mice.
Error bars indicate the mean ± SEM. n ¼ 12–14 for each point.
N E U R O P S I N ( K L K 8 ) I N U V B - I R R A D I A T E D S K I N 7 0 5
� 2003 British Association of Dermatologists, British Journal of Dermatology, 149, 700–706

2 MacDonald R, Southard E, Kroon E. Discrete tissue-specific
expression of members of the tissue kallikrein multigene family of
the rat. J Biol Chem 1995; 271: 13684–90.
3 Murray SR, Chao J, Lin FK et al. Kallikrein multigene families and
the regulation of their expression. J Cardiovasc Pharmacol 1990;
15: S7–16.
4 van Leeuwen B, Evans B, Tregear G et al. Mouse glandular kal-
likrein genes. J Biol Chem 1986; 251: 5529–35.
5 Diamandis EP, Yousef GM, Luo LY et al. The new human kallik-
rein gene family: implications in carcinogenesis. Trends Endocrinol
Metab 2000; 11: 54–60.
6 Harvey TJ, Hooper JD, Myers SA et al. Tissue-specific expression
patterns and fine mapping of the human kallikrein (KLK) locus on
proximal 19q13.4. J Biol Chem 2000; 275: 37397–406.
7 Komai S, Matsuyama T, Matsumoto K et al. Neuropsin regu-
lates an early phase of Schaffer-collateral long-term potentia-
tion in the murine hippocampus. Eur J Neurosci 2000; 12:
1479–86.
8 Kitayoshi H, Inoue N, Kuwae K et al. Effect of 12-O-tetradecanoyl-
phorbol ester and incisional wounding on neuropsin mRNA and
its protein expression in murine skin. Arch Dermatol Res 1999;
291: 333–8.
9 Scarisbrick IA, Blaber SI, Lucchinetti CF et al. Activity of a newly
identified serine protease in CNS demyelination. Brain 2002;
125: 1283–96.
10 Chen ZL, Yoshida S, Kato K et al. Expression and activity-
dependent changes of a novel limbic-serine protease gene in the
hippocampus. J Neurosci 1995; 15: 5088–97.
11 Yoshida S, Hirata A, Inoue N et al. Assignment of the neuropsin
gene (Prss19) to mouse chromosome band 7B4 by in situ
hybridization. Cytogenet Cell Genet 2000; 88: 97–8.
12 Yoshida S, Taniguchi M, Hirata A et al. Sequence analysis and
expression of human neuropsin cDNA and gene. Gene 1998; 213:
9–16.
13 Matsumoto-Miyai K, Kitagawa R, Ninomiya A et al. Dividualiza-
tion induces expression and activation of an extracellular prote-
ase neuropsin in mouse uterus. Biol Reprod 2002, 67: 1414–18.
14 Inoue N, Kuwae K, Ishida-Yamamoto A et al. Expression of neu-
ropsin in the keratinizing epithelial tissue—immunohistochemical
analysis of wild-type and nude mice. J Invest Dermatol 1998; 110:
923–31.
15 Kuwae K, Matsumoto-Miyai K, Yoshida S et al. Epidermal
expression of serine protease, neuropsin (KLK8) in normal and
pathologic skin. Mol Pathol 2002; 55: 235–41.
16 Tomizawa K, He X, Yamanaka H et al. Injury induces neuropsin
mRNA in the central nervous system. Brain Res 1999; 824:
308–11.
17 Hirata A, Yoshida S, Inoue N et al. Abnormalities of synapses and
neurons in the hippocampus of neuropsin-deficient mice. Mol Cell
Neurosci 2001; 17: 600–10.
18 Chen ZL, Momota Y, Kato K et al. Expression of neuropsin mRNA
in the mouse embryo and the pregnant uterus. J Histochem
Cytochem 1998; 46: 313–20.
19 Saade NE, Nasr IW, Massaad CA et al. Modulation of ultraviolet-
induced hyperalgesia and cytokine upregulation by interleukins
10 and 13. Br J Pharmacol 2000; 131: 1317–24.
20 Sluyter R, Halliday GM. Enhanced tumor growth in UV-irradiated
skin is associated with an influx of inflammatory cells into the
epidermis. Carcinogenesis 2000; 21: 1801–7.
21 Gasparro FP, Brown DB. Photobiology 102: UV sources and
dosimetry—the proper use and measurement of �photons as a
reagent�. J Invest Dermatol 2000; 114: 613–15.
22 Ishida-Yamamoto A, Iizuka H. Differences in involucrin immu-
nolabeling within cornified cell envelopes in normal and psoriatic
epidermis. J Invest Dermatol 1995; 104: 391–5.
23 Danno K, Horio T. Formation of UV-induced apoptosis relates to
the cell cycle. Br J Dermatol 1982; 107: 423–8.
24 Fitzpatrick TB, Johnson RA, Wolff K et al. Color Atlas and Synopsis
of Clinical Dermatology, 4th edn. New York: McGraw-Hill, 2001.
25 Steinert PM, Marekov LN. Initiation of assembly of the cell
envelope barrier structure of stratified squamous epithelia. Mol
Biol Cell 1999; 10: 4247–61.
26 Shimizu C, Yoshida S, Shibata M et al. Characterization of
recombinant and brain neuropsin, a plasticity-related serine
protease. J Biol Chem 1998; 273: 11189–96.
27 Tani N, Matsumoto K, Ota I et al. Effects of fibronectin cleaved by
neuropsin on cell adhesion and migration. Neurosci Res 2001; 39:
247–51.
7 0 6 T . K I R I H A R A et al.
� 2003 British Association of Dermatologists, British Journal of Dermatology, 149, 700–706