prohead and dna-gp3-dependent atpase activity of the dna
TRANSCRIPT

.I. No/. Hid. (1987) 197, 229-236
Prohead and DNA-gp3-dependent ATPase Activity of the DNA Packaging
Protein gp16 of Bacteriophage 429
Peixuan Guot, Charlene Peterson and Dwight Anderson
Departments of Microbiology and Dentistry tTn,iversity of Minnesota, Minneapolis, MN 55455. C’.S.A.
(Received 12 January 1987, and in revised form 7 May 1987)
The ATPase activity of the DNA packaging protein gp16 (gene product 16) of bacteriophage 429 was studied in the completely defined in-vitro assembly system. ATP was hydrolyzed to ADP and Pi in the packaging reaction that included purified proheads, DNA-gp3 and gp16. Approximately one molecule of ATP was used in the packaging of 2 base-pairs of 429 DNA, or 9 x lo3 ATP molecules per virion. The hydrolysis of ATP by gpl6 was both prohead and DNA-gp3 dependent. gp16 contained both the “A-type” and bhe “B-type” ATP-binding consensus sequences (Walker et al., 1982) and the predicted secondary structure for ATP binding. The A-type sequence of gp16 was “basic-hydrophobic region-G-X,-G-X-G-K-S-X,-hydrophobic”, and similar sequences were found in the phage DNA packaging proteins gpA of lambda, gp19 of T7 and gp17 of T4. Having both the ATP- binding and potential magnesium-binding domains, all of these proteins probably fumtion as ATPases and may have common prohead-binding capabilities. The 429 protein gp3. oovalently bound to the DNA, may be analogous in fun&ion to proteins gpNu1 of lambda and gpl of 421 that bind the DNA.
1. Introduction
The tailed, double-stranded DNA bacteriophages purportedly have a common mechanism of genome encapsidation (for reviews, see Earnshaw & Casjens, 1980: Casjens, 1985). Commonalities of the DNA packaging machines of several of these phages include the use of a pair of non-capsid proteins and ATP to effect translocation of the viral DNA into a preformed protein shell. We have summarized briefly the attributes of these systems and provided the rationale for use of the Bacillus subtilis bacteriophage 429 to study the DNA packaging mechanism (Guo et al., 1987); most significantly, 429 offers an efficient, completely defined in-vitro assembly system (Guo et al.. 1986).
With the defined $29 assembly system, we have demonstrated the initial sequential events in DNA- gp3$ packaging. showing that the packaging protein gpl6 first binds to, and is modified by, the prohead (Guo et aZ., 1987). The modified gp16 then reacts with DNA-gp3. resulting in a second
t Permanent address: South China Agricultural IYnivrrsit*v. (iuangzhou (Canton). The People’s Republic ot’ China.
1 Abbreviation used: gp. gene product,.
modification of gpl6 that promotes trapping of ATP and ATP hydrolysis. Surprisingly, the DNA- gp3 is converted to an aggregated form, even though 429 DNA replication, which is primed by gp3 covalently bound at the 5’ termini, produces monomers (for a review, see Geiduschek & Ito, 1982). This is mindful of the “headful” packaging process of other phages that excise genomes nucleolytically from concatemers composed of covalently linked replicative products.
The energetias of the DNA packaging process may also hold general significance. The ability to transform free energy into motion, a ubiquitous property of biological systems, is manifest in the translocation of viral nucleic acid into a protein shell. In the present paper, we show directly that ATP is hydrolyzed to ADP and Pi when all components of the defined in-vitro packaging system are present’, including proheads, DNA-gp3 and the DNA packaging protein gpl6: no pair of reactants is sufficient. The gpl6 has the ATP- binding consensus sequences (Walker et al., 1982) and may have the potential magnesium-binding domain (Berg, 1986). Therefore, the gpl6 by itself (after modification by the prohead and the DNA- gp3) or the prohead/gpl6/DNA-gp3 complex must constitute the 429 ATPase. Finally. we provide
0022.-2836!87/180229-08 $03.00/O 229
0 1987 Academic Press Limited
230 P. Guo et al.
quantification of ATP hydrolysis with purified components from which the bulk of contaminating ATPases has been removed; approximately one ATP molecule is used in the in-vitro packaging of two base-pairs of 429 DNA.
2. Materials and Methods
(a) Chemicals and isotopes
[c(-35S]ATP (adenosine 5’-[a-35S]thiotriphosphate) and [y-32P]ATP (adenosine 5’-[y-32P]triphosphate) were from Amersham Corp.
(b) In-vitro DNA-gp3 packaging in the dejned system
The purification of gp16, DNA-gp3 and proheads, and the methods of in-vitro assembly in the defined system, have been described (Guo et al., 1986. 1987).
(c) Sequence search, alignment and secondary structure prediction
Search and alignment of ATP-binding and magnesium- binding sequences of bacteriophage DNA packaging proteins was by the computer program PEP of TntelliGenetics (IntelliGenetics, Inc.) and visual inspection. The secondary structure of the gp16 sequence was predicted by a computer program of Cambridge- Bask, according to the method of Garnier et al. (1978).
(d) ATPase assays
In the defined in-vitro assembly system, volumes of reaction buffer, proheads, gpl6 and [3H]DNApgp3 were generally mixed in the proportions 3 : 3 : 7 : 10 (Guo et al., 1986). Mixtures were incubated on Parafilm for various times at ambient temperature. When one or more constituents were omitted (Table l), they were replaced with the appropriate volume(s) of buffer or salt solution. [y-32P]ATP or [c(-3’S]ATP was added at 42@Zi/ml and 600 @i/ml, respectively, and the reaction was initiated by the addition of gpl6. A reaction with dog kidney ATPase (Sigma Chem. Co.) was used as a control. At 4 min and 30 min, samples of the reaction mixtures were spotted on to PEI-cellulose (J. T. Baker Chem. Co.) and dried. The chromatogram was soaked in methanol for 5 min, redried, and run in 1 M-formic acid/05 M-lithium chloride. Autoradiograms were produced on Kodak XAR-5 X-ray film and used to localize the labeled ATP and ADP or Pi spots on the chromatograms; these areas were excised and assayed by liquid scintillation counting.
At the same time, filled heads containing L3H]DiVA gp3 were assayed by sucrose density gradient centrifuga- tion as a measure of packaging efficiency. A sample of the reaction mixture was loaded onto a 5% to 20% linear sucrose density gradient containing TMS buffer (Guo et al., 1987) and centrifuged in the SW55 rotor at 35,000 revs/min for 30 min at 20°C; filled heads sedi- mented to fractions 4 to 6 (Guo et al., 1986. 1987).
3. Results
(a) ATP is hydrolyzed to ADP and Pi in the complete packaging reaction
Hydrolysis of radiolabeled ATP in the defined in- vitro DNA-gp3 packaging system was assayed
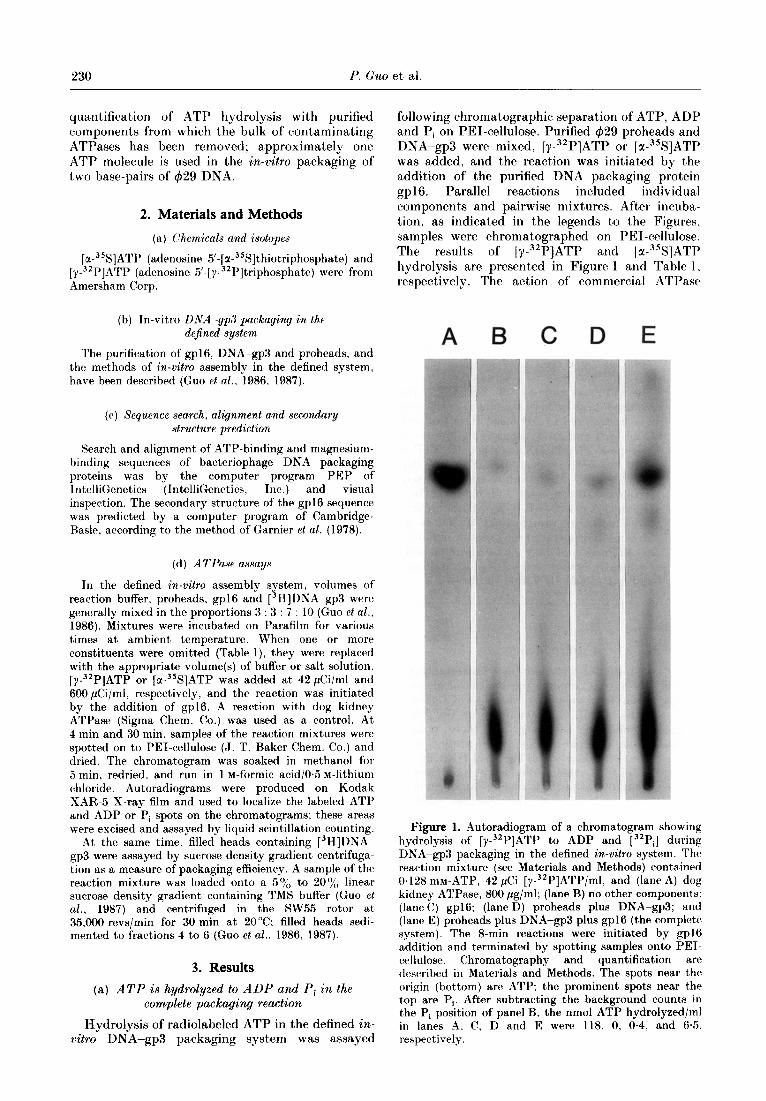
following chromatographic separation of ATP, ADP and Pi on PEI-cellulose. Purified 429 proheads and DNA-gp3 were mixed, [y-32P]ATP or [z-~%]ATP was added, and the reaction was initiated by the addition of the purified DNA packaging protein .wl6. Parallel reactions included individual components and pairwise mixtures. After incuba- tion, as indicated in the legends to the Figures, samples were chromatographed on PEI-cellulose. The results of [Y-~‘P]ATP and [u-~%]ATP hydrolysis are presented in Figure 1 and Table 1. respectively. The action of commercial ATPase
A B C D E
Figure 1. Autoradiogram of a chromatogram showing hydrolysis of [y-32P]ATP to ADP and [32Pi] during DNA-gp3 packaging in the defined in-vitro system. The reaction mixture (see Materials and Methods) contained 0.128 mM-ATP, 42 &i [y-32P]ATP/ml, and (lane A) dog kidney ATPase, 800 pg/ml; (lane B) no other component,s; (IaneC) gpl6; (lane D) proheads plus DNA-gp3; and (lane E) proheads plus DPjA-gp3 plus gp16 (the complete system). The 8-min reactions were initiated by gp16 addition and terminated by spotting samples onto PET- cellulose. Chromatography and quantification are described in Materials and Methods. The spots near the origin (bottom) are ATP; the prominent spots near the top are Pi. After subtracting the background counts in the Pi position of panel B, the nmol ATP hydrolyzed/ml in lanes A. C, D and E were 118, 0, 0.4, and 65. respectively.

Prohead- and DNA-gpd-dependent -4 TPase 231
marked t.he J’osition of Pi (Fig. 1, lane A). (‘hromatography of the [Y-~~P]ATP alone is shown in lane H. and lanes C and 0 show that gpl6 alone or proheads plus Dh’A-gp3 produced little Pi over this background. Pi production by the complete reaction &lixture is illust,rated in lane E. After subtracting t,he counts at the Pi position in t’he ~Y-“~P]ATP a,lone (lane B), the nmoles ATP/ml, hydrolyzed by the gpl6 alone, proheads plus DNA- gp3, and the complete reaction mixture were 0 (lane (‘). 04 (lane D) and 6.5 (lane E), respectively, while 118 nmoles ATP/ml were hydrolyzed by the c~ommercial ATPase (lane A). To quantify [ 3H]DSA~-gp3 packaging in the complete system (lanr E). conversion of label to filled heads was analyzed by sucrose density gradient centrifugation (see (:uo et (11.. 1!)87). In the experiment of Figure I, 7.70,, (4.3 x lO”,‘ml) of the 13H]DNA-gp3 added in the reaction was packaged into filled heads in eight minutes. and 3.9 X 1015 ATP molecules/ml were used in J)a(akaging the 18 x 103 base-pair genome. The results were comparable when the conversion of la-35S]ATP to 135S]ADP was analyzed by the same chromatography methods (Table 1). In addition, all of the individual components were studied, and a comparison of the results of reactions 1: 4, 5 and 7 demonstrated that the ATPase activity of gp16 was both Jjrohead and DNA-gp3 dependent. The prohead requirement support,ed the conclusion that gpl6 was moditied by the prohead (Guo et al., 1987). The prohead plus DNA-gp3 mixture (reaction 6) showed more ATP hydrolysis than proheads or D&A-gp3 alone, but less than one- fourth t)hr activit>y of t’he complete mixture (react.ion i). In the complete system, 3.2 x IO” DSA -gp3 complexes/ml were packaged, and 3.1 x JO15 XTP molecules/ml were hydrolyzed. In the cxperinrc~nt,s wit,h IY-~~P]ATP and [M-~‘S]ATP, 0.60 and 0.54 ATP molecules, respectively, were used in the J)ackaging of 1 base-pair of 429 DNA.
(I)) gpl6 contains both “il-type” and “B-type” .-I TP-binding consensus sequences
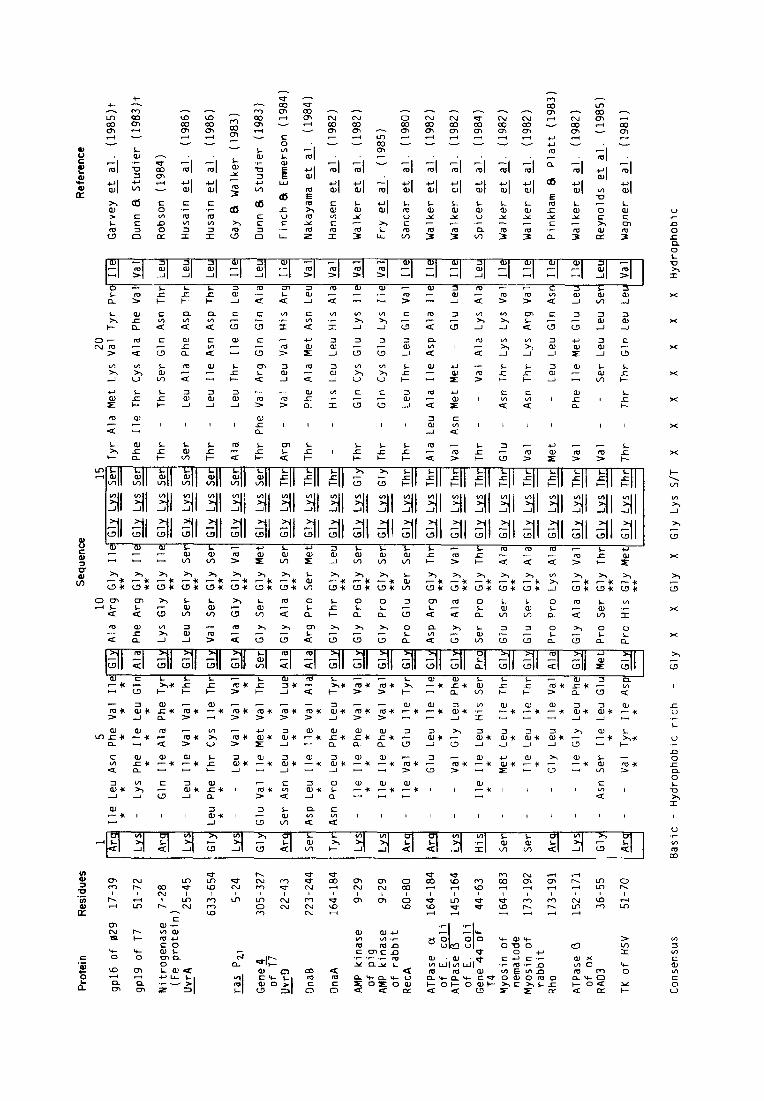
W:alker ut ccl. (190%) reported that two types of seyuences are Jmasentj in a number of ATPases and suggested that these sequences may constitute part of the nucltaotide-binding domain. The “A-type” sequence consist,s of G-X,-GK(T)-X,-I/V, preceded bv a basic amino acid, where X can be any amino a&d. The “B-type” sequence contains the common sequence R/K-X,-(:-X,-L-hydrophobic4 followed by an aspartica acid residue. Some ATPases have both sequen(*es, and others have either the A- or the B-type. Both sequences purportedly contribute to nucleotide binding: some of the residues in the A- type srqu~n~e make contacts with bot)h the adenine ring and t.hc phosphates, while the B-type sequence forms the back of the nucleotide pocket (Fry et al., 1985. 19X6). LVe searched for t’hese sequences and found that both the A-type and the B-type were well c~onservt~d in t’he DNA packaging protein gp16 of 429 (Figs 2. 3 and 4).
The common secondary structure of t hr Jjeptide sequence for nucleotide binding has been predictrd (Risler et al.. 1981: Krauhs et al.. 1981: Walker rt al., 1982: Husain et al.. 1986). It csomprisrs H fl- sheet, a flexible j-turn and an a-helix. Recently. Fry et al. (1985, 1986) studied the MgATP-binding site of adenylate kinase by nucalear magnetic resonance and X-ray diffraction studies. The three- dimensional structure of the J)&ein and the positions of MgATP and the ATP-binding site have been described in detail. Three segments of the adenylate kinase sequence have been c~harac~terixrcl. Segment 1 (positions 8 t’o 14 in Fig. 2). t(~rminstt~tl by a lysine, is a glycine-rich flexible IooJ~ that. ma) control access to the ATP-binding site I)>, changing its conformation. The -h’Hi group of this lysinc could be positioned very close to the z-phosJ)hate.
The secondary structure of segment 1 in gJ) I6 was predict,ed to be a p-sheet followed by a flexible IooJ) in t,he glycine-rich region (Fig. 3). This J)r(sdic*tion agrees with the secondary structure in other ATI’- binding proteins (Fig. 3). Alt,hough the glycine-ricah region (residues 24 to 30) of gpl6 in Figure 3 was marked as a p-sheet and P-t’urn structure, this region scored very low in computecl J)otjential for either a-helix or P-sheet structure. suggesting a flexible loop similar t,o t’hat predicted in ot,her ATT’- binding proteins (Risler et al., 1981: Krauhs pf n/.. 1981; Walker et al., 1982: Husain rf nl.. J 986: Fry rf al.. 1985, 1986).
Segment 2 of the adenylatc kinase sequence includes residues 27 to 31, LVS-Jl~-\~al -His-J,ys (Fig.2 does not depict this whole sequencr). This segment is an a-helix containing two hydrophobic residues that interact with the adeninr-ribostl of ATP and a lysine that might bind t,o the p- and r- phosphate of ATP. gpJ6 also contained t ht. similar a-helix His -Glu -I,eu-Val -\‘a1 -1,~s at rcsidut~s 76 to -.__ 81 (Garvey et al.. 1985).
Segment 3 of adenylate kinase, a 13.tyJ)e ATI’- binding consensus sequence depicted bb- M’alkrr pf al. (1982). is a hydrophobic strand of [I-pleated sheet’, terminated by an asJJart,atr. t hxt is thr
Table 1
Reaction (‘omponentst ATP hydrolyzed
(nnrol/ml)
I gpl6 04!)*0~1~ 2 DNA/gp3 0.1 I *0.15 3 Proheads 0.3” * 04~2 4 gp I6 + DNA/gp3 0.37*0.14 5 gp I6 + proheads 0~54 + 0.24 6 DNA/gp3 + proheads 124+0.;1 7 gp16 + DNA/gp3 + proheads 5 I ” + w.w -
t (Components of the defined system in reaction buffer containing 0.118 mwunlabeled ATP and 600 p(‘i lw3%]ATf’/ml were incubated for 10min at ambient temperature. Samples spotted on cellulose-PEI were chromatographed to separate labeled ADP and ATP as described in Materials and Methods. The complete mixture (gp16+ DNA/gp% +proheads) was done in triplicate, while other mixtures were done in ttupliatr
Prot
ein
Res
idue
s
gp16
of
02
9 17
-39
gp19
of
T7
51
-72
Nitr
ogen
ase
7-28
(F
e pr
otei
n)
Uvr
A 25
-45
-
633-
654
ras
Pzl
- 5-
24
Gen
e4
of
TT
Uvr
D
-
305-
327
22-4
3
Dna
B 22
3-24
4
Dna
A 16
4-18
4
AMP
kina
se
9-29
of
pi
g AM
P ki
nase
9-
29
of
rabb
it R
ecA
60-8
0
ATPa
se
CL
164-
184
of
E.
coli
ATPa
Fe
R
145-
164
of
E.
coli
Gen
e-44
of
44
-63
T4
Myo
sin
of
nem
atod
e M
yosi
n of
ra
bbit
Rho
164-
183
173-
192
173-
191
152-
171
36-5
5
51-7
0
1 5
10
15
20
Arq‘
Ile
Le
u As
n Ph
e Va
l I1
Al
a Ar
g G
ly
Ile
G&
& Se
r Ty
r Al
a M
et
Lys
Val
Tyr
Pro
Ile
* *
* *
* **
---
&-
Lys
Phe
Ile
Leu
Gl
Phe
Arg
Gly
Ile
a
& Se
r Ph
e Ile
Th
r C
ys
Ala
Phe
Val
Val
* *
* **
---
Arq
- G
ln
Ile
Ala
Phe
Ty
Lys
Gly
G
ly
Ile
u &
Ser
Thr
- Th
r Se
r G
ln
Asn
Thr
Leu
* *
* **
---
b -
Leu
Ile
Val
Val
Th
Leu
Ser
Gly
Se
r G
lx
Ls
Ser
Ser
- Le
u Al
a Ph
e As
p Th
r Le
u *
* *
* **
---
Gly
Le
u Ph
e Th
r C
ys
Ile
Th
Val
Ser
Gly
Se
r G
& &
Ser
Thr
- Le
u Ile
As
n As
p Th
r Le
u *
* *
**
---
LJ-
- Le
u Va
l Va
l Va
Al
a G
ly
Gly
Va
l G
ly
Ls
Ser
Ala
- Le
u Th
r Ile
G
ln
Leu
Ile
* *
* *
**
---
Gly
G
lu
Val
Ile
Met
Va
l Th
G
ly
Ser
Gly
M
et
Gly
Ls
Se
r Th
r Ph
e Va
l Ar
g G
ln
Gln
Al
a Le
u *
* *
* **
---
Arq
Ser
Asn
Leu
Leu
Val
Lu
Gly
Al
a G
ly
Ser
a &
Thr
Arg
- Va
l Le
u Va
l H
is
Arg
Iie
* *
* *
**
---
Ser
Asp
Leu
Ile
Ile
Val
Al
Arg
Pro
Ser
Met
m
&
Thr
Thr
- Ph
e Al
a M
et
Asn
Leu
Val
* *
* *
---
Tyr
Asn
Pro
Leu
Phe
Leu
Ty
Gly
Th
r G
ly
Leu
Gly
Lf
i Th
r -
- H
is
Leu
Leu
His
Al
a Va
l *
* *
* **
---
& -
Ile
Ile
Phe
Val
Va
Gly
Pr
o G
ly
Ser
G&
I,rr
Gly
Th
r -
Gln
C
ys
Glu
Ly
s Ile
Va
l *
* *
* *
**
--
& -
Ile
Ile
Phe
Val
Va
Gly
Pr
o G
ly
Ser
G&
& G
ly
Thr
- G
in
Cys
G
lu
Lys
Ile
Val
* *
* *
* **
--
Arq
- Ile
Va
l G
lu
Ile
Ty
Pro
Glu
Se
r Se
r =
& Th
r Th
r -
Leu
Thr
Leu
Gln
Va
l I l
e *
* *
---
Arc-
-
Glu
Le
u Ile
11
As
p Ar
g G
ly
Thr
Q
& Th
r Al
a Le
u Al
a Ile
As
p Al
a Ile
Ile
*
* *
**
---
&-
- Va
l G
ly
Leu
Ph
Gly
Al
a G
ly
Val
Q
& Th
r Va
l As
n M
et
Met
-
Glu
Le
u Ile
*
* *
**
---
His
-
Ile
Ile
Leu
His
Se
Se
r Pr
o G
ly
Thr
Gly
Ls
Th
r Th
r -
- Va
l Al
a Ly
s Al
a Le
u *
* *
**
---
Ser
- -
Met
Le
u Ile
Th
G
lu
Ser
Gly
Al
a G
J &
Thr
Glu
-
Asn
Thr
Lys
Lys
Val
Ile
* *
* **
---
Ser
- -
Ile
Leu
Ile
Th
Glu
Se
r G
ly
Ala
Gly
LQ
Th
r Va
l -
Asn
Thr
Lys
Arg
Val
Ile
* *
* **
---
Arq-
-
Gly
Le
u Ile
Va
Pr
o Pr
o Ly
s Al
a 9
& Th
r M
et
- -
Leu
Leu
Gln
As
n Ile
*
* *
---
&-
- Ile
G
ly
Leu
Ph
* *
* G
ly
Ala
Gly
Va
l m
&
Thr
Val
- Ph
e Ile
M
et
Glu
Le
u Ile
**
---
Gly
-
Asn
Ser
Ile
Leu
Gl
* *
Pro
Ser
Gly
Th
r Q
w
Thr
Val
- -
Ser
Leu
Leu
Ser
Leu
**
---
Arq-
-
Val
Tyr
Ile
Asp
* *
* Pr
o H
is
Gly
M
et
Gly
L~
J Th
r Th
r -
Thr
Thr
Gin
Le
u Le
u Va
l **
---
Spic
er
et
al.
(198
4)
--
Wal
ker
et
al.
(198
2)
Wal
ker
et
al.
(198
2)
--
Pink
ham
8
Plat
t (1
983)
ATPa
se
0 of
ox
R
AD3
TK
of
HSV
Wal
ker
et
al.
(198
2)
--
Rey
nold
s et
al
. (1
985)
--
Wag
ner
et
al.
(198
1)
--
Con
sens
us
Basi
c -
Hyd
roph
obic
ric
h -
Gly
X
X G
ly
X G
ly
Lys
S/T
X X
X X
X X
X H
ydro
phob
ic
Sequ
ence
R
efer
ence
Gar
vey
et
al.
(198
5)t
Dun
n 8
Stud
ier
(198
3)t
Rob
son
(198
4)
Hus
ain
et
al.
(198
6)
--
Hus
ain
et
al.
(198
6)
--
Gay
&
Wal
ker
(198
3)
Dun
n 8
Stud
ier
(198
3)
Finc
h &
Emm
erso
n (1
984)
Nak
ayam
a et
al
. (1
984)
--
Han
sen
et
al.
(198
2)
--
Wal
ker
et
al.
(198
2)
--
Fry
et
al.
(198
5)
--
Sanc
ar
et
al.
(198
0)
--
Wal
ker
et
al.
(198
2)
--
Wal
ker
et
al.
(198
2)

Prohead- and DNA-gp3-dependent A TPase 233
Protein
Adenylate kinase
Residues Sequence and secondary structure Reference
9-21 K I I FVVGGPGSGK Walker et u/. (1982)
Nw?w~~-
UtifA 25-37 KLIVVTGLSGSGK Husain et a/. (1986)
R-I?“-
.!/vrA 633-645 LFTCITGVSGSGK Husain et al. (1966)
,w-
gpl6 of +29 18-30 ILNFVIGARG IGK This work
Figure 3. Predicted secondary st,ructurr of segment, 1 that may contribute to the nucleotide binding function. Serondary structures with homology are shown for comparison. w----- ,&sheet: - ----- random c~)il; n----- p-turn.
triphosphat.ebinding sit.e (Fry et nE., 1986). Its forms the back of the nucleotide pocket. This segment was well conserved in gp16 (Fig. 4) and was also computed to be the b-pleated sheet.
((a) Align,ment of homologous sequences in the DYA packaging proteins of various
bacteriophages
Many phages specify two non-capsid proteins t,hat fun&on in DNA packaging. These include gpNu1 and gpA of lambda, gp16 and gp17 of T4, gpl8 and gpl9 of T3 and T7, gp2 and gp3 of P22, and gpM and gpP of P2. In 429, gp16 is required, and a second essent,ial protein, gp3 (Bjornsti et al., 1983), is covalently bound to the 5’ ends of the DNA. All of t,hese systems may be basically similar.
Because of the complexity of the DNA packaging process, little is known about the functions of these protein pairs. We have demonstrated that the ATPase activity of gp16 is DNA-gp3-dependent. gp16 contains both t,he A-type and B-type ATP- binding consensus sequences of ATPases and the potential magnesium-binding domain. Searching the sequences of DNA packaging proteins of other phages, we found the A-type ATP-binding consensus sequence in gpA of la.mbda, gp17 of T4 and gp19 of T7. Figure 5 shows the homologies of these sequences.
(d) gplh’ of &29 as ,u)ell as DIVA packaging proteins of other ph,ages con&in potential magnesium-binding
domains
Recently, Miller et al. (1985) and Berg (1986) predicted that closely positioned Cz and/or His -- residues can form a potential metal-binding domain
in nucleic acid-binding proteins. These were classi- fied into five groups (Berg, 1986). The metal- binding domain in gpA of lambda is His -X,-His -X2- Cys-X2-Cys (Berg. 1986). gp2 of the lambda relative -1 is highly homologous t,o gpA of lambda. We searched for the above met,al-binding domain in the sequence of gp2 (Miller 8t Feiss, 1985) and found that the domain was oonserved. except, that t,he second His of the domain was changed tSo Tyr. The magnesium-binding domain in gp17 of T4 has been predicted to be His-X,-His-Xi6-Cys-Xs- -- His-X,-His, with the His-X,-His residues at 7 posrtionsx2 to 385 and=1 tom (Powell, I)., Franklin, J., Arisaka, F. & Mosig? G., unpublished results).
The only two cysteine residues in gp16 are proximate (Garvey et al., 1985), and the reductant dithiothreitol is required during denaturation of gp16 by guanidinium chloride (Quo et al., 1986). Thus the -SH groups of the only two cysteine residues in gp16 must be protected to maintain DNA packaging function. Moreover, the require- ment of magnesium for 429 DNA-gp3 packaging has been demonstrated (Bjornsti et a/., 1981). In the defined 429 DNA-gp3 packaging system, three proteins are candidates for magnesium binding: the gpl9 of the portal vertex (connector), the gp3 covalently linked to the DNA termini, and the DNA packaging protein gp16. There arc no closely linked Cys and/or His doma,ins in the published sequences of gpl0 (Garcia et al.. 1984) and gp3 (Escarmis b Salas, 1982). Therefore the Cys-Be-Cys .Y-- domain in the gp16 sequence at positions 153 to 155, possibly together with the His residues at -- positions 268 and 298 (Garvey et ~1.. 19X5), may bc
Figure 2. (lomparison of A-type sequences for ATP binding of the @9 DNA packaging protein gplB and ot,hrr proteins. The ronsmsus sequence of Walker et al. (1982) is in boxes. The identical amino acids are double underlined. \vhile the similar ones (Miyata et al., 1979) are single underlined. Note that there is a region rich in hydrophobic amino acids (*) between the basic amino acid and the glycine residue, and that there is anot.her conserved glyckw wsidue (**). TK, thymidine kinase; HSV, herpes simplex virus.
t Search for consensus sequence is our work.

234 P. Guo et al
Protein
gp16 of 029
ATPase I3 of ox
ATPase I3 of E. fl
ATPase a of E. coli
ATP/ADP translocase
ras Pzl
Adenylate kinase
Phosphofructokinase
Residues
248-256
250-258
235-243
273-281
283-291
37-46
-
G
G
G
G
G
G
G
G -
F *
Q
R
E
A
E
Q
I *
Sequence
L *
V *
V *
A
V *
C
T
G
G
L *
L *
L *
L *
L *
L *
L *
V *
L *
L *
I *
V *
L *
L *
V *
W *
F *
F *
I *
L *
D
L *
V *
V *
I *
V *
Y *
Y *
I *
Y *
I *
-
D
D
D
D
D
Figure 4. Comparisons of segment 3 sequences (B-type sequences) proposed to be the triphosphate-binding site. The sequence of gene 16 of 429 is from Garvey et aE. (1985). Comparisons involving the other 7 proteins are from Fry el al. (1986). The identical amino acids are in solid boxes. The hydrophobic ones are marked with an asterisk.
responsible for magnesium binding. This is con- sistent with the ATP-binding consensus sequence in
sequences, the Cys and/or His residues are generally
gp16 and the role of gpl6 as a prohead and DNA- separated by two amino acids; the only two Cys
-
gp3-dependent ATPase. Moreover, we found that residues in gp16 are separated by a single ammo
gp19 of phage T7 contained the A-type ATP- acid (Garvey et al., 1985).
binding consensus sequence at residues 51 to 72 (Fig. 5) and a potential magnesium-binding domain 4. Discussion His-X,-His at residues 343 to 346 (Dunn & Studier, -- The ATPase activity of the 429 DNA packing 1983). In most of the magnesium-binding protein gp16 is both prohead and DNA-gp3-
Protein Residues
gp16 of 029 17-39
gp19 of T7 51-72
gpA of h 484-505
gp17 of T4 154-174
293-314
Seauence Reference
RILNFVI
3 /
ARG ** *** ===
KKF-ILQ FRG * * * ==
RVI-PIK ASV * * * =
RMTVCNLS-ZQ * * * *
HFY-DI * *
TiAV * *
TY :M K V Y PI Garvey et al. (1985)
Dunn & Studier (1983)
Daniels et al. (1983) --
Powell et a 1. (unpublished results) --
Powel 1 et al. (unpublished results) --
Figure 5. Comparison of the A-type ATP-binding sequence in gp16 of (629 with similar sequences in gp19 of T7, gpA of I and gp17 of T4. The Walker et al. (1982) consensus sequence for ATP binding is in solid boxes. The identical amino acids are double underlined, while the similar ones (Miyata et aE., 1979) are single underlined. The amino acids in the, hydrophobic region are marked with an asterisk.

Prohead- and DNA-gp3-dependent ATI’ase 235
dependent) (Fig. 1, Table I; Guo et al., 1987). gp16 contains both the A-type and B-type ATP-binding consensus sequences of ATPases (Figs 2 to 4) and a potential magnesium-binding domain, and therefore must be the ATPase or a,n integral part of the ATPase. The prohead (probably the connector) and the gp3 of DNA-gp3 must either induce modifica- tion of gpl6 to produce the ATPase activity or constitute parts of the active enzyme. If the latter is true, the procapsid is converted from a passive shell to part of an active enzyme that translocates DNA into its cramped interior.
DNA-dependent ATPase activity of the lambda trrminase and the T3 gp18/gpl9 complex has been demonst,rated (Gold & Reeker, 1983; Hamada et al., 1984). Concerning the DNA-gp3 dependence of gl~16. is this related t’o DNA or gp3 or both? The question may he difficult to answer definitively because gp3 is covalently linked to 429 DNA; prot,einase or 1)Nase treatment leaves a few amino acids or nucleotides, respectively, and these may be sufficient to bestow the changes in the enzyme needed for ATPase activity.
Prohead dependence of t,he 429 ATPase activity is apparent)lT a, new observat,ion, although phage T3 ATPase actrvity in vitro was reported to be prohead stimulated (Hamada et al.. 1984). The gp16 used here was purified from Escherichia coli harboring a plasmid containing gene 26, and therefore the gp16 had not reacted with proheads. On the contrary, part’ially purified terminase, the bacteriophage lambda DNA packaging complex composed of the prot,eins gpr\‘ul and gpA. was demonstrated to have DNA-dependent ATPase activity in vitro without the prohead (Gold & Becker, 1983): the terminase was isolated from a lysogen with a lambda deletion- substitution so t,hat, prohead constituents were not produced.
Segment 1 of the ATP-binding consensus sequence (Fig. 3) is a glycine-rich flexible loop that is purported to be diagnostic of nucleotide-binding proteins (Walker et nl., 1982). However, it, appears to havtb a function other than simply holding a nuclrotidr (M&Lughlin et al., 1984; Sachsenheimer & Schulz. 1977: Fry et al., 1985, 1986). The function of this loop most likely involves its ability to undergo a conformational change in response either to substrate binding or to an interaction with another protein or domain. Fry et al. (1986) indicated three possible roles for a conformational change in segment 1: (1) control of accessibility to the sabstrat~e-binding site, (2) modification of binding sit,r affinities. and (3) relocation of catalytic groups t’oward the reaction center of the bound suhstratr. Segment 1 of the DNA packaging protein gpl6 of 429 was a flexible loop that manifested a very low score in cbomputed potential for either /3- sheet) or a-helix st’ructure. We have demonstrated t’hat gp16 undergoes conformational changes or moditication in the DNA-gp3 packaging process (Guo et nl.. 1987). Modification by the prohead permits binding to DNA-gp3, and DNA-gp3 binding ac*tuates a second change that permits
trapping of ATP. ATP binding and hydrolysis stabilize the prohead/gplG/DNA-gp3 complex and result in t,he formation of DNA aggregates and packaging. The non-hydrolyzable ATP analog (y-S)ATP permits an unstable interaction between gp16 and DNA-gp3. Because the modification of segment, 1 involves nucleotide binding, the last change(s) of gpl6 might be related t’o the function of segment 1. Conformational change of the segment 1 loop of gpl6 triggered 1~.y l)NA-gp3 binding may have made the MgATP-binding site accessible. Another conformational change of seg- ment 1 after MgATP binding may have stabilized the gplS/DNA-gp3 complex.
The two non-capsid DNA packaging prot’eins of various bacteriophages apparently co-operate to link the prohead to the DNA packaging substrate. cut monomers of DNA from concatemrrs and aid in processive packaging (for reviews. see Earnshaw & Casjens, 1980; Casjens, 1985). Terminase of lambda, composed of the int#erac*t.ing polypept~idrs gpNu1 and gpA, presents a linear array of functional domains (for a review. see l+iss. 1986). but the two proteins may still have separatck roles among the multiple functions. (‘onsi(lering the protein pairs of various phages, on(h prot,ein ma? bind the prohead and function as an ATPasr. while the other binds the DNA. The domain for prohead binding is at, the carboxy terminus of ~IJA of lambda (Frackman et al.. 1984). gpf9 of ‘1’7 purportedly int,eracts with the minor prohead protein gp8 situated at the prohead portal vertex (Nakasu et nl.. 1983; Hamada rt nl.. 1984). and genetic evidence suggests that, gp16 of 1’4 interacts with the connector (Hsiao & Black, 1977: Klack & Showe, 1983). We have demonstrated the binding of gp16 of 429 to the prohead (Guo pf ~1.. 1987). Moreover, t’erminase of lambda has a I)NL\\- dependent ATPase activity ((iold & f%xker, 1983). and we noticed that gpA of lambda. gpl6 of @!j. gpl9 of T7 and gp17 of T4 all contain the ATI’- binding consensus sequence. Finally. potent,ial magnesium-binding domains were identified in gp.A of lambda (Berg, 1986) and gpl6 of 429 (this
paper), and we found a His-X,-His domain in gp19 of T7 that may have a comparable function (~tris paper). A potential magnesium-binding domain has been identified as well as gpl7 of T4 (Powell, D., Franklin, J., Arisaka, F. & Mosig. (:. unpublished results). On these bases we speculate that gpA of lambda, gp2 of 421, gpl9 of T3 and Ti, gp17 of T4. and gpl6 of $29 havtb common properties of prohead binding and ATPast> activit? (Table 2). Considering the function of t.he second protein of each pair, the DNA-binding activity of gpNu1 of lambda and gpl of $21 have been demonstrated clearly (for a review. sc~ F&s, 1986). gp3 of 429 is covalently bound to t hta 5’ t ermini of the DNA. Both gp16 a;d gpf 7 of T-t are purported to be DNA-binding proteins (131ack KT Show?. 1983). Inclusion of gp18 of T3/T7 in t,his group. however. is speculat,ive.
Approximately one ATT’ was usecl in t h(>

236 P. Guo et al.
Table 2 Provisional relationship of gp3 and gp16 of 429 to
other phage DNA pa&aging proteins
Bacteriophage
I#229
qJt1 T4 T3 T7
Group 1: prohead-binding
magnesium-binding Group 2: DNA-dependent ATPase DNA-binding
gP16 gP3 gp A gpNu1 gP2 gPl gP17 gp16 gP19 gp181 gP19 gp18?
packaging of two base-pairs of 429 DNA, or 9 x 103 ATP molecules per virion. An expenditure of one ATP molecule to package 1 base-pair is estimated to account for about 5% of the total energy required for production of the lambda virion (Hendrix, 1978), and thus the DNA packaging requirement is not so extravagant. By comparison, helicases active in E. coli DNA replication hydrolyze two ATP molecules per base-pair broken (Kornberg et al., 1978; Yarranton & Gefter, 1979).
We acknowledge the technical help of Stephen Erickson, Marv-Ann Gomes, Jesdawan Wichitwechkam and Shelley Grimes. Sequence alignments were supervised by Hershel Ginsberg of the Sun Computer Cent,er of the University of Minnesota. Beat Keller of the University of Basle. Switzerland, helped with the gp16 secondary structure prediction. We thank Gisela Mosig for providing the T4 gene 17 sequence and location of the potential magnesium-binding domain prior to publication. We are grateful to Carol Church for typing the manuscript and to Shuang Yong Xu for helpful discussion. The work was supported by grant DE-03606 from the National Tnstitutes of Health.
References Berg, J. M. (1986). Science, 232, 485-487. Bjornsti, M. A.. Reilly, B. E. & Anderson, D. 1,. (1981).
Proc. Nat. Acad. Sci., U.S.A. 78, 5861-5865. Bjornsti, M. A., Reilly, B. E. & Anderson. 1). L. (1983).
J. Virol. 45, 383-396. Black, L. W. & Showe, M. (1983). In Bacteriophage T4
(Matthews, C.. Kutter, E., Mosig, G. & Berget, P.. eds), pp. 219-245, American Society for Micro- biology, Washington, DC.
Casjens, S. (1985). In Virus Structure and Assembly (Casjens, S., ed.), pp. 75-147, Jones & Bartlett, Portola Valley, CA.
Daniels, D. L., Schroeder, J. L., Szybalski, W., Sanger, F., Coulson, A. R.. Hong, G. F., Hill, D. F., Petersen. G. B. & Blattner. F. R. (1983). In Lambda II (Hendrix, R. W.. Roberts, J. W., Stahl. F. W. & Weisberg, R. A., eds), pp. 519-676, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Dunn, J. J. & Studier, F. W. (1983). J. MOE. Biol. 166. 477-535.
Earnshaw, W. C. & Casjens, S. R. (1980). Cell. 21, 319- 331.
Escarmis, C. & Salas, M. (1982). Nucl. Acids Res. 10. 578555789.
Feiss, M. (1986). Trends Genet. 2, 100-104. Finch. P. W. & Emmerson, P. T. (1984). Nucl. Acids Res.
12, 5789-5799.
Frackman, S.. Siegele, D. A. & Feiss, M. (1984). J. Mol. BioZ. 180, 283-300.
Fry, D. C.. Kuby, S. A. & Mildvan, A. S. (1985). Biochemistry, 24, 4680-4694.
Fry, D. C.. Kuby, S. A. & Mildvan, A. S. (1986). Proc. Xat. Acad. Sci., U.S.A. 83, 907-911.
Garcia. J. A., Mendez, E. & Salas. M. (1984). Gene. 30. 87-98.
(iarnier, J.. Osguthorpe. 1). J. & Robson. B. (1978). .J. Mol. BioZ. 120, 97-120.
Garvey, K. <J.. Saedi. M. S. 8: Ito. J. (1985). Gene, 40, 31 l-316.
Gay. X. J. & Walker, J. E. (1983). Nature (London), 301, 262-264.
Geiduschek, E. I’. & Ito, J. (1982). In Molecular Biology of the Lh.%~~i (Dubnau, D.. ed.), pp. 203-245, Academic Press, New York.
Gold, M. & Becker, A. (1983). J. BioZ. Chem. 258, 14619-- 14625.
Guo. P., Grimes. S. & Anderson, D. (1986). Proc. Nat. Acad. Sci., U.9.A. 83, 3505-3509.
Guo, P., Peterson, C. & Anderson, D. (1987). J. Mol. RioZ. 197, 219-228.
Hamada. K.. Fujisawa, H. & Minagawa. T. (1984). virozogy, 139. 251-259.
Hansen, E. B.. Hansen, F. G. & Von Meyenburg, K. (1982). NucZ. Acid Res. 10, 7373-7385.
Hendrix, R’. W. (1978). Proc. Nat. Acad. Rci., I’.R.A 75, 4779-4783.
Hsiao. C. I,. & Black. L. W. (1977). Proc. Nat. Acad. Sci., 1 :.S.A. 74, 3652-3656.
Husain. I.. Houten, B. V., Thomas, D. C. & Sancar. A. (1986). J. Biol. Chem. 261, 4895-4901.
Kornberg, A., Scott, J. F. & Bertsch. L. L. (1978). J. RioZ. Chem. 253, 3298-3304.
Krauhs. E., Little, M., Kenpt, T.. Warbinek. R. H.. Ade, W. & Pongtingl. H. (1981). Proc. Nat. Acad. Sci., I’X.A. 78. 4156-.4160.
,&Laughlin, I’. J.. Stuart. 1). I.. Klein, H. W.. Oikonomakos, N. G. & Johnson. L. N. (1984). Biochemistry. 23. 5862-5873.
Miller. A. J.. McLachlan, A. D. & Klug, A. (1985). EMRO J. 4, 1609-1614.
Miller. G. & Feiss. M. (1985). J. Mol. BioZ. 185, 246-249. Miyata, T., Miyazawa. S. & Yasunaga, T. (1979). J. Mol.
Evol. 12, 219-236. Nakasu. R.. Fujisawa, H. & Minagawa, T. (1983).
Virology, 127, 124-133. Nakayama. N.. Arai. N., Bond, M. W., Kaziro. Y. & Arai,
K. (1984). J. Riol. Chem. 259, 97p101. Pinkham. ,J. 1,. & Platt. T. (1983). NucZ. Acids Res. 11,
3531L3545. Reynolds. P.. Higgins, D. R., Prakash, L. & Prakash. S.
(1985). NucZ. Acids Res. 13, 2357-2372. Risler, ,J. L.. Zelwer. C. & Brunie, S. (1981). Nature
(London). 292, 384-386. Robson. R. L. (1984). FEBS Letters, 173, 394-398. Sachsenheimer, W. & Schulz, G. E. (1977). J. Mol. BioZ.
114, 23-36. Sancar, A.. Stachelek, C., Konigsberg, W. & Rupp, W. D.
(1980). Proc. Nut. Acud. Sci., U.S.A. 77, 2611-2615. Spicer, E. K., Nossal, N. G. & Williams, K. R. (1984).
J. BioZ. Chem. 259, 15425-15432. Wagner, M. J., Sharp, J. A. & Summers, W. C. (1981).
Proc. Nat. Acad. Sci., U.S.A. 78, 1441-1445. Walker, tJ. E., Sarasate, M., Runswick, M. J. & Gay, N. J.
(1982). EMBO J. 1, 945-951. Yarranton, 0. T. & Gefter, M. L. (1979). Proc. Nut. Acad.
Sri.. I,‘.N.A. 76, 1658-1662. Edited by A. Klug