post-thaw aging affects activity of lactate dehydrogenase
TRANSCRIPT

NOTE
Post-Thaw Aging Affects Activity of Lactate Dehydrogenase
BAKUL S. BHATNAGAR,1 SARAH J. NEHM,1 MICHAEL J. PIKAL,1,2 ROBIN H. BOGNER1,2
1School of Pharmacy, U2092, University of Connecticut, Storrs, Connecticut, 06269
2Institute of Materials Science, University of Connecticut, Storrs, Connecticut, 06269
Received 22 September 2004; revised 31 January 2005; accepted 31 January 2005
Published online in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/jps.20345
ABSTRACT: Freeze-thawing is routinely used to study freezing-induced irreversibleprotein denaturation in the formulation characterization and development of lyophilizedproteins. In most cases, the temperature profiles of the samples are not fully monitoredduring freeze-thawing and therefore, the sample thermal histories are largely unknown.The objective of this study was to develop experimental protocols for the study ofisothermal protein degradation using a temperature-step apparatus. Freeze-thawexperiments were performed at a freezing rate of 108C/min and various thawing rates(0.5–3.38C/min) using a temperature-step apparatus. In our efforts to design validationstudies, we encountered anomalies in the recovered enzyme activity data of an enzyme,lactate dehydrogenase at the end of freeze-thawing. The effect of thawing rate wasstudied to explain the variability in the data. In addition, post-thaw ‘‘aging’’ of freshlyfrozen and thawed samples was performed at 58C to reduce the variability in therecovered enzyme activity. Results from these experiments implicate the use of aging ofdilute multimeric enzymes at the end of freeze-thawing to control the variability inenzyme assays. � 2005 Wiley-Liss, Inc. and the American Pharmacists Association J Pharm Sci
94:1382–1388, 2005
Keywords: aging; freeze-drying; freeze-thawing; ice nucleation; lyophilization; pre-formulation; protein formulation; stability
INTRODUCTION
Freezing and thawing can cause irreversibledamage to proteins, which is manifested by a lossof structure, conformation, or biological activity.1
Different stresses such as cold denaturation, iceformation, and freeze concentration develop as aresult of the physical changes that occur onfreezing.2–13 Freeze-thawing is routinely used asa tool to understand the combined damagingeffects of these different stresses on protein
stability during pre-formulation characterizationand formulation development of freeze-dried pro-teins. It is, perhaps, the most direct approachto determine the extent of freezing-inducedirreversible protein denaturation, where theamount of damage at the end of freeze-thawcycle can be quantified by means of a suitableassay. As a result, numerous examples ofsuch studies can be found in the literature wherethe effects of either rate of freezing and/orthawing or of freeze-thaw cycling have beendocumented. Examples of the enzymes studiedinclude L-alcohol dehydrogenase,14 a-amylase,15
L-asparaginase,16,17 catalase,18–20 b-galactosi-dase,21,22 glutamate dehydrogenase,23 L-lactatedehydrogenase23–29, and phosphofructokinase.30
1382 JOURNAL OF PHARMACEUTICAL SCIENCES, VOL. 94, NO. 6, JUNE 2005
Correspondence to: Robin H. Bogner (Telephone: 860-486-2136; Fax: 860-486-4998; E-mail: [email protected])
Journal of Pharmaceutical Sciences, Vol. 94, 1382–1388 (2005)� 2005 Wiley-Liss, Inc. and the American Pharmacists Association

A variety of approaches are used to producefreeze-thawing of protein solutions (Table 1).However, difficulties in controlling heat transferduring these processes have made results fromthe studies difficult to interpret. Therefore, therehas been a continued interest in simplifying themethodology and resolving the effects of freez-ing from those of thawing on stability. The vastnumber of approaches described in the literaturehighlights the complicated nature of freeze-thawstudies. While most investigators are careful tocontrol the temperatures to which the samplecontainers are subjected, there are very few thatmonitor the thermal profiles of the samples them-selves. If the variable heat transfer effects thatoccur during cooling, freezing, or thawing are notfully monitored, it could potentially lead to dif-ficulties in comparing various studies.
In our efforts to develop appropriate experi-mental protocols for the study of isothermal,irreversible protein degradation, we encounteredintriguing stability behavior where greater activ-ity retention was observed for frozen enzymesamples thawed at faster heating rates whencompared to that of the unfrozen, starting solu-tions. The objective of this note is to describe theeffect of thawing rate on the recovery of enzymaticactivity of frozen lactate dehydrogenase (LDH),and then to discuss the impact of aging post-thawing on LDH activity. The data presented hereare part of validation studies for the use of atemperature-step apparatus in the study of iso-thermal protein degradation.
EXPERIMENTAL
Materials
Ammonium sulfate-precipitated LDH (EC1.1.1.27, M4 isozyme suspension in 3.2M ammo-nium sulfate) derived from rabbit muscle waspurchased from Sigma Chemical Co. (St. Louis,MO). Sodium citrate and citric acid were obtainedfrom Fisher Scientific (Pittsburgh, PA). Double-distilled water filtered through 0.22 mM filter wasused for the preparation of all solutions. A 0.05Mcitrate buffer was prepared and filtered priorto use.
Enzyme Preparation and LDH CatalyticActivity Assay
LDH was dialyzed against 0.05M citrate buffer(pH 7.4) at 48C overnight before use. The proteinconcentration was determined by a Bradfordassay.31 Unless otherwise noted, all experi-ments employed a total LDH concentration of5 mg/mL. The catalytic activity of LDH wasdetermined by monitoring the rate of decreasein the absorbance of NADH following the reduc-tion of pyruvate to lactate at 340 nm at 258Cusing an LDH assay kit (Sigma Chemical Co.).For all experiments, the activities of the ex-perimental samples were compared to those ofunfrozen, untreated LDH. In all cases, theenzyme activity of at least four samples wasdetermined.
Table 1. Schematic of Various Possible Steps During Freeze-Thawing of Enzymes
Reference
Step ICooling Slow (<18C/min) 4,14,15,20,23,29,36,37
Intermediate (1–108C/min) 14,20,28,29,36,37
Rapid (10–9008C/min) 4,16,20,23,25,26,29,36,38,39
Step IIFreezing Without seeding 4,14,16,20,23,26,27
With seeding 20,29,32
Step IIIIsothermal hold
Step IVThawing Slow (<18C/min) 4,14–16,20,23,26–28,38–40
Intermediate (1–58C/min) 20,29,37
Rapid (>58C/min) 16,20,23,27,29,38
Step VAGING for time ‘‘t’’ following
thawingPresent study
POST-THAW AGING OF LACTATE DEHYDROGENASE 1383
JOURNAL OF PHARMACEUTICAL SCIENCES, VOL. 94, NO. 6, JUNE 2005

Freeze-Thawing Experimental Setup
During cooling in a traditional freezer or a freeze-dryer, a drug solution spends a finite time ateach temperature as it is lowered to the desiredsubzero setpoint. Therefore, the degradation dataobtained do not reflect isothermal behavior. Fur-thermore, the generated data becomes moredifficult to interpret if the kinetics of instabilityis of the same order of magnitude as the rate ofcooling. A rapid temperature-step to the subzerotemperature of interest enables one to bypass theslow cooling that solutions experience in a con-ventional cooling step in a freeze-dryer and can beuseful in the collection of data on the kineticsof isothermal, irreversible degradation of labileproteins.
In this study, freezing and thawing of LDHwasperformed in a specially fabricated assembly con-sisting of a highly polished nickel-plated copperplate with hemispherical sample wells (0.26 mLvolume) carved into the plate (Figure 1). Thenickel-plated copper plate was embedded in ahollow Plexiglas block, which had an inlet and anoutlet for the flow of an ethylene glycol:watermixture underneath the plate and through theblock. At the inlet and the outlet of the block, thedevice was connected to a 58C bath and a subzerotemperature programmable bath (initial bath tem-perature¼�6.58C) through single switch valves.All exposed surfaces of the temperature-step
assembly and the rubber tubing were insulatedto minimize heat loss to the surroundings and toensure an efficient temperature-step. This setupenabled us to produce a ‘‘rapid’’ temperature-stepfrom 5.0� 0.38C to �5.0� 0.38C. The sampleswere covered with a Plexiglas lid and the sampletemperature was monitored in one sample well(dummy sample) using a copper-constantan ther-mocouple connected to a temperature readoutdevice (Omega Engineering, Stamford, CT). Ther-mocouples were not placed in all sample wells asthey can act as a nucleation site for ice crystals.
At the beginning of an experiment, only the flowfrom the 58C bath occurred through the Plexiglasblock. The enzyme samples were placed in thesample wells and allowed to cool to 58C. The flowthrough the Plexiglas block was then switchedfrom the 58C bath to the subzero temperature bathusing the single-switch valves. A change of theflow from one bath to the other caused a rapidtemperature-step within the enzyme samples inthe wells. All temperature-steps were completewithin 45–60 s. No freezing occurred duringthe temperature-step. Two types of samples werestudied using the temperature-step assembly,(1) unfrozen and (2) frozen. An unfrozen samplewas used to ensure that the temperature-stepitself did not contribute to a loss in the catalyticactivity of LDH. In this case, a temperature-stepfrom 58C to �58C was followed by immediatelywarming the sample to 58C at 28C/min. For the
Figure 1. Schematic of the temperature-step assembly used to produce freezing andthawing of LDH solutions.
1384 BHATNAGAR ET AL.
JOURNAL OF PHARMACEUTICAL SCIENCES, VOL. 94, NO. 6, JUNE 2005
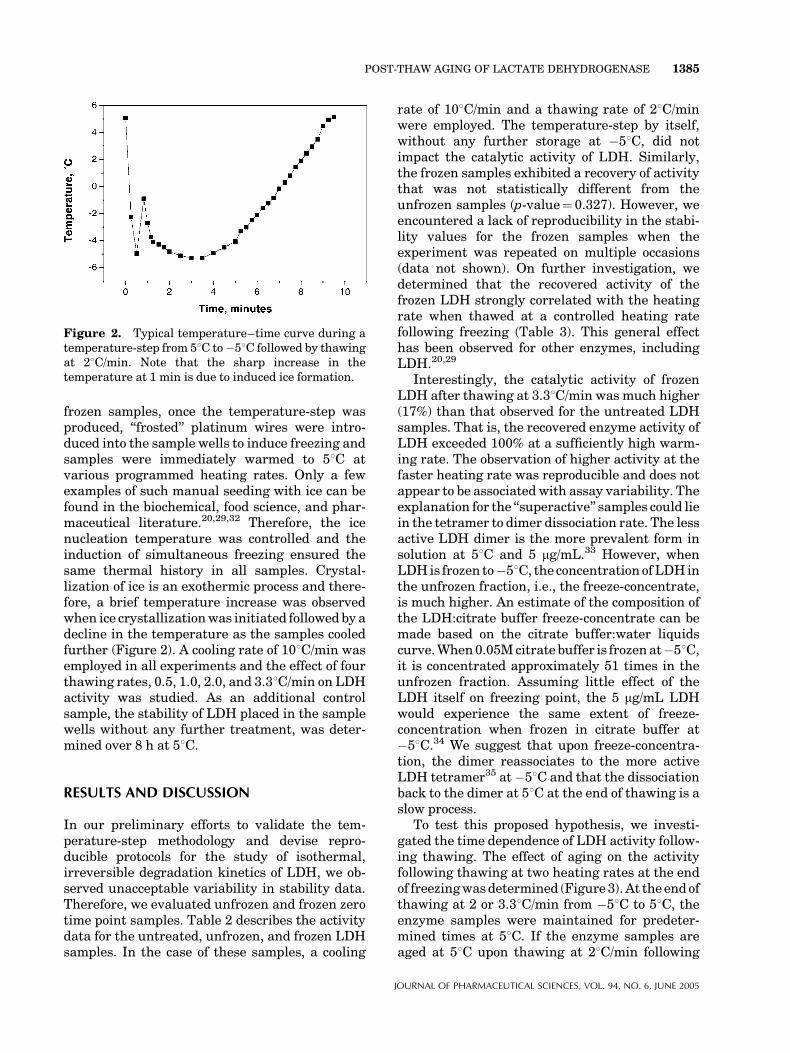
frozen samples, once the temperature-step wasproduced, ‘‘frosted’’ platinum wires were intro-duced into the sample wells to induce freezing andsamples were immediately warmed to 58C atvarious programmed heating rates. Only a fewexamples of such manual seeding with ice can befound in the biochemical, food science, and phar-maceutical literature.20,29,32 Therefore, the icenucleation temperature was controlled and theinduction of simultaneous freezing ensured thesame thermal history in all samples. Crystal-lization of ice is an exothermic process and there-fore, a brief temperature increase was observedwhen ice crystallizationwas initiated followed by adecline in the temperature as the samples cooledfurther (Figure 2). A cooling rate of 108C/min wasemployed in all experiments and the effect of fourthawing rates, 0.5, 1.0, 2.0, and 3.38C/min on LDHactivity was studied. As an additional controlsample, the stability of LDH placed in the samplewells without any further treatment, was deter-mined over 8 h at 58C.
RESULTS AND DISCUSSION
In our preliminary efforts to validate the tem-perature-step methodology and devise repro-ducible protocols for the study of isothermal,irreversible degradation kinetics of LDH, we ob-served unacceptable variability in stability data.Therefore, we evaluated unfrozen and frozen zerotime point samples. Table 2 describes the activitydata for the untreated, unfrozen, and frozen LDHsamples. In the case of these samples, a cooling
rate of 108C/min and a thawing rate of 28C/minwere employed. The temperature-step by itself,without any further storage at �58C, did notimpact the catalytic activity of LDH. Similarly,the frozen samples exhibited a recovery of activitythat was not statistically different from theunfrozen samples (p-value¼ 0.327). However, weencountered a lack of reproducibility in the stabi-lity values for the frozen samples when theexperiment was repeated on multiple occasions(data not shown). On further investigation, wedetermined that the recovered activity of thefrozen LDH strongly correlated with the heatingrate when thawed at a controlled heating ratefollowing freezing (Table 3). This general effecthas been observed for other enzymes, includingLDH.20,29
Interestingly, the catalytic activity of frozenLDH after thawing at 3.38C/min was much higher(17%) than that observed for the untreated LDHsamples. That is, the recovered enzyme activity ofLDH exceeded 100% at a sufficiently high warm-ing rate. The observation of higher activity at thefaster heating rate was reproducible and does notappear to be associated with assay variability. Theexplanation for the ‘‘superactive’’ samples could liein the tetramer to dimer dissociation rate. The lessactive LDH dimer is the more prevalent form insolution at 58C and 5 mg/mL.33 However, whenLDHis frozen to�58C, the concentration ofLDHinthe unfrozen fraction, i.e., the freeze-concentrate,is much higher. An estimate of the composition ofthe LDH:citrate buffer freeze-concentrate can bemade based on the citrate buffer:water liquidscurve.When0.05Mcitrate buffer is frozenat�58C,it is concentrated approximately 51 times in theunfrozen fraction. Assuming little effect of theLDH itself on freezing point, the 5 mg/mL LDHwould experience the same extent of freeze-concentration when frozen in citrate buffer at�58C.34 We suggest that upon freeze-concentra-tion, the dimer reassociates to the more activeLDH tetramer35 at�58C and that the dissociationback to the dimer at 58C at the end of thawing is aslow process.
To test this proposed hypothesis, we investi-gated the time dependence of LDH activity follow-ing thawing. The effect of aging on the activityfollowing thawing at two heating rates at the endof freezingwasdetermined (Figure3).At the endofthawing at 2 or 3.38C/min from �58C to 58C, theenzyme samples were maintained for predeter-mined times at 58C. If the enzyme samples areaged at 58C upon thawing at 28C/min following
Figure 2. Typical temperature–time curve during atemperature-step from 58C to�58C followed by thawingat 28C/min. Note that the sharp increase in thetemperature at 1 min is due to induced ice formation.
POST-THAW AGING OF LACTATE DEHYDROGENASE 1385
JOURNAL OF PHARMACEUTICAL SCIENCES, VOL. 94, NO. 6, JUNE 2005

freezing, the decrease in the activity to 100% takesless time (&15 min). In contrast, it takes 60 min toreach 100% activity in samples thawed at thefaster heating rate (3.38C/min) followed by agingat 58C. The decrease in activity on aging is asurprising result as an earlier observation onuntreated LDH showed no change in activity ofsolutions stored at 58C for 8 h (Table 4). It appearsthat the heating rate employed during thawinginfluences the distribution of LDH subunits andfurther aging the enzyme samples restores thedimer–tetramer equilibrium to a more dimer richand less active state.
To test the hypothesis that LDH exists as adimer at 5 mg/mL at 58C, we explored the use oflight scattering to determine themolecular weightof LDH samples of various concentrations. How-ever, we found that the light scattering signal wasweak and unreliable at low concentrations (i.e.,�50 mg/mL). An attempt was also made to usefluorescence spectroscopy to determine whetherthe LDH dimer is the more prevalent form at a5 mg/mL by determining the surface hydrophobi-city of LDH in the presence of a fluorescent probe,
1-anilinonaphthalene-8-sulfonate (1,8-ANS) as afunction of LDH concentration. However, the fluo-rescence signal was unreliably low at the LDHconcentration we needed in these experiments.While we were unable to obtain conclusive corro-boration to support our hypothesis that freeze-concentration induces the formation of the moreactive tetramer, which slowly dissociates uponthawing, the aging studies are consistentwith thathypothesis.
CONCLUSIONS
The effect of freezing and thawing rates onenzyme activity has been widely explored in theliterature. However, the effect of post-thaw aginghas not been previously investigated. Most com-monly pH change or urea/guanidium HCl areused to mediate enzyme dissociation, which inturn further leads to a change in activity. Enzymeactivity can also be altered by dilution of buffer,
Table 2. Percent Activity Remaining FollowingCooling After a Temperature-Step From 58C to �58Cand Thawing at 28C/min*
Sample Percent Remaining Activity
Untreateda 100 (5)Unfrozenb 109 (2)Frozenc 110 (5)
n� 3 (SD).*Activity is measured relative to the starting solution.aRepresents the activity of LDH sample not subjected to any
treatment in the temperature-step assembly.bRepresents the activity of LDH sample maintained unfro-
zen following the temperature-step and thawed immediately.cRepresents the activity of LDH sample frozen by seeding
with ice following the temperature-step and thawed immedi-ately.
Table 3. Percent Activity Remaining in FrozenSamples Following Thawing at Different HeatingRates After a Temperature-Step From 58C to �58C*
Heating Rate, 8C/min Percent Remaining Activity
0.5 70 (2)1.0 92 (5)2.0 107 (6)3.3 117 (2)
n� 3 (SD).*Activity is measured relative to the starting solution.
Figure 3. Percent activity remaining of frozen LDHsamples as a function of aging time following thawing at2.08C/min and 3.38C/min after a temperature-step from58C to �58C.
Table 4. Percent Activity Remaining at �58C forUntreated Samples in the Temperature-Step Assemblyin the Absence of any Freezing or Temperature-Step*
Time, h Percent Remaining Activity
0 100 (1)1 100 (2)2 100 (2)8 102 (3)
n� 3 (SD).*Activity is measured relative to the starting solution.
1386 BHATNAGAR ET AL.
JOURNAL OF PHARMACEUTICAL SCIENCES, VOL. 94, NO. 6, JUNE 2005

formulation components, and the protein itselfduring thawing. It is established by this work thataging affects activity and we postulate that thistime dependence arises from a shift of the equi-librium between the tetramer and dimer species.The time dependence of the tetramer to dimertransition can impact the reproducibility of theenzyme assay. Therefore, the aging of dilutemultimeric enzymes can be used to reduce vari-ability in assay results.
ACKNOWLEDGMENTS
The authors thank Dr. Devendra (Davy) Kaloniafor his helpful discussions. The support of theNational Science Foundation Center for Pharma-ceutical Processing Research (NSF CPPR) forBakul Bhatnagar and the University of Connecti-cut/Pfizer Summer Research Fellowships Pro-gram for Sarah Nehm are acknowledged.
REFERENCES
1. Pikal MJ. 2004. Mechanisms of protein stabiliza-tion during freeze-drying and storage: The relativeimportance of thermodynamic stabilization andglassy state relaxation dynamics. In: Freeze-drying/lyophilization of pharmaceutical and biolo-gical products, drugs and the pharmaceuticalsciences. New York: Marcel Dekker, pp. 63–107.
2. Jaenicke R, Zavodszky P. 1990. Proteins underextreme physical conditions. FEBS Lett 268:344–349.
3. Privalov PL. 1990. Cold denaturation of proteins.Crit Rev Biochem Mol Biol 25:281–305.
4. Chang BS, Kendrick BS, Carpenter JF. 1996.Surface-induced denaturation of proteins duringfreezing and its inhibition by surfactants. J PharmSci 85:1325–1330.
5. Kreilgaard L, Jones LS, Randolph TW, Frokjaer S,Flink JM, et al. 1998. Effect of Tween 20 on freeze-thawing and agitation-induced aggregation of re-combinant human factor XIII. J Pharm Sci 87:1597–1603.
6. Strambini GB, Gabellieri E. 1996. Proteins infrozen solutions: Evidence of ice-induced partialunfolding. Biophys J 70:971–976.
7. Gabellieri E, Strambini GB. 2003. Perturbationof protein tertiary structure in frozen solutionsrevealed by 1-anilino-8-naphthalene sulfonate fluo-rescence. Biophys J 85:3214–3220.
8. Izutsu K-I, Kojima S. 2000. Freeze-concentrationseparates proteins and polymer excipients into
different amorphous phases. Pharm Res 17:1316–1322.
9. Parker R, Ring SG. 1995. A theoretical analysis ofdiffusion-controlled reactions in frozen solutions.Cryo-Letters 16:197–208.
10. Heller MC, Carpenter JF, Randolph TW. 1996.Effects of phase separating systems on lyophilizedhemoglobin. J Pharm Sci 85:1358–1362.
11. Randolph TW. 1997. Phase separation of excipientsduring lyophilization: Effects on protein stability. JPharm Sci 86:1198–1203.
12. Pincock RE. 1969. Reactions in frozen systems. AccChem Res 2:97–103.
13. Gomez G, Pikal MJ, Rodriguez-Hornedo N. 2001.Effect of initial buffer composition on pH changesduring far-from-equilibrium freezing of sodiumphosphate buffer solutions. Pharm Res 18:90–97.
14. Tamiya T, Okahashi N, Sakuma R, Aoyama T,Akahane T, Matsumoto JJ. 1985. Freeze denatura-tion of enzymes and its prevention with additives.Cryobiology 22:446–456.
15. Whittam JH, Rosano HL. 1973. Effects of thefreeze-thaw process on a-amylase. Cryobiology 10:240–243.
16. Izutsu K-I, Yoshioka S, Terao T. 1994. Stabilizingeffect of amphiphilic excipients on the freeze-thawing and freeze-drying of lactate dehydrogen-ase. Biotechnol Bioeng 43:1102–1107.
17. Jameel F, Kalonia D, Bogner R. 1995. The effect ofhetastarch on the stability of L-asparaginase duringfreeze-thaw cycling. PDA J Pharm Sci Techno49:127–131.
18. Kiermeier F. 1947. Influence of freezing on theactivity of enzymes. Biochem Z 318:275–296.
19. Shikama K, Yamazaki I. 1961. Denaturation ofcatalase by freezing and thawing. Nature 190:83–84.
20. Fishbein WN, Winkert JW. 1977. Parameters ofbiological freezing damage in simple solutions:Catalase. I. The characteristic pattern of intracel-lular freezing damage exhibited in a membranelesssystem. Cryobiology 14:389–398.
21. Izutsu K-I, Yoshioka S, Terao T. 1994. Effect ofmannitol crystallinity on the stabilization ofenzymes during freeze-drying. Chem Pharm Bull42:5–8.
22. Pikal-Cleland KA, Rodriguez-Hornedo N, AmidonGL, Carpenter JF. 2000. Protein denaturationduring freezing and thawing in phosphate buffersystems: Monomeric and tetrameric b-galactosi-dase. Arch Biochem Biophys 384:398–406.
23. Chilson OP, Costello LA, Kaplan NO. 1965. Effectsof freezing on enzymes. Fed Proc 24:55–65.
24. Soliman FS, Van den Berg L. 1971. Factorsaffecting freezing damage of lactic dehydrogenase.Cryobiology 8:73–78.
25. Nema S, Avis KE. 1993. Freeze-thaw studies of amodel protein, lactate dehydrogenase, in the pre-
POST-THAW AGING OF LACTATE DEHYDROGENASE 1387
JOURNAL OF PHARMACEUTICAL SCIENCES, VOL. 94, NO. 6, JUNE 2005

sence of cryoprotectants. J Parenter Sci Technol 47:76–83.
26. Anchordoquy TJ, Carpenter JF. 1996. Polymersprotect lactate dehydrogenase during freeze-dryingby inhibiting dissociation in the frozen state. ArchBiochem Biophys 332:231–238.
27. Anchordoquy TJ, Izutsu K-I, Randolph TW,Carpenter JF. 2001. Maintenance of quaternarystructure in the frozen state stabilizes lactatedehydrogenase during freeze-drying. Arch BiochemBiophys 390:35–41.
28. Jiang S, Nail SL. 1998. Effect of process conditionson recovery of protein activity after freezing andfreeze-drying. Eur J Pharm Biopharm 45:249–257.
29. Cao E, Chen Y, Cui Z, Foster PR. 2003. Effect offreezing and thawing rates on denaturation ofproteins in aqueous solutions. Biotechnol Bioeng82:684–690.
30. Carpenter JF, Hand SC, Crowe LM, Crowe JH.1986. Cryoprotection of phosphofructokinase withorganic solutes: Characterization of enhancedprotection in the presence of divalent cations. ArchBiochem Biophys 250:505–512.
31. Bradford MM. 1976. A rapid and sensitive methodfor the quantitation of microgram quantities ofprotein utilizing the principle of protein-dye bind-ing. Anal Biochem 72:248–254.
32. Brandts JF, Fu J, Nordin JH. 1970. Low tem-perature denaturation of chymotrypsinogen inaqueous solution and in frozen aqueous solution.
In: Wolstenholme GE, O’Connor M, editors. Thefrozen cell, A Ciba foundation symposium. London:J & A Churchill, pp. 189–212.
33. Yamamoto S, Storey KB. 1988. Dissociation-association of lactate dehydrogenase isozymes:Influences on the formation of tetramers versusdimers of M4-LDH and H4-LDH. Int J Biochem20:1261–1265.
34. Weast RC, editor. 1974. Handbook of chemistry andphysics. 55th ed. Ohio: CRC Press, D-200 p.
35. Girg R, Rudolph R, Jaenicke R. 1983. The dimericintermediate on the pathway of reconstitution oflactate dehydrogenase is enzymatically active.FEBS Lett 163:132–135.
36. Greiff D, Kelly RT. 1966. Cryotolerance of enzymes.I. Freezing of lactic dehydrogenase. Cryobiology2:335–341.
37. Eckhardt BM, Oeswein JQ, Bewley TA. 1991.Effect of freezing on aggregation of human growthhormone. Pharm Res 8:1360–1364.
38. Lippert K, Galinski EA. 1992. Enzyme stabilizationby ectoine-type compatible solutes: Protectionagainst heating, freezing and drying. Appl Micro-biol Biotechnol 37:61–65.
39. Miller DP, Anderson RE, De Pablo JJ. 1998.Stabilization of lactate dehydrogenase followingfreeze-thawing and vacuum-drying in the presenceof trehalose and borate. Pharm Res 15:1215–1221.
40. Nema S, Avis KE. 1993. Loss of LDH activityduring membrane filtration. J Parenter Sci Technol47:16–21.
1388 BHATNAGAR ET AL.
JOURNAL OF PHARMACEUTICAL SCIENCES, VOL. 94, NO. 6, JUNE 2005