porphyrin biosynthesis in cell-free homogenates higher · plant physiol. (1970) 46, 543-549...
TRANSCRIPT

Plant Physiol. (1970) 46, 543-549
Porphyrin Biosynthesis in Cell-free Homogenates fromHigher Plants1
Received for publication February 24, 1970
CONSTANTIN A. REBEIZ,2 MouNIR ABou HAIDAR, AND MOUSTAPHA YAGHDepartment of Biological Sciences, Scientific and Agricultural Research Institute, Fanar, LebanonPAUL A. CASTELFRANCODepartment of Botany, University of California, Davis, California 95616
ABSTRACT
The porphyrin and phorbin biosynthetic activity of etio-lated cucumber (Cucumis sativus, L.) cotyledons was com-pared to that of cotyledonary homogenates. Etiolated coty-ledons incubated with 5-aminolevulinic acid accumulateprotoporphyrin, coproporphyrin, small amounts of Mgprotoporphyrin monoester, and trace amounts of uropor-phyrin. They also incorporate 4-14C-5-aminolevulinic acidinto free porphyrins, protochlorophyllide, protochlorophyl-lide phytyl ester, and Mg protoporphyrin monoester. Ho-mogenates incubated with 5-aminolevulinic acid likewiseaccumulate coproporphyrin, uroporphyrin, Mg copropor-phyrin, and trace amounts of protoporphyrin. They alsoincorporate 4-"C-8-aminolevulinic acid into Mg protopor-phyrin monoester, Mg coproporphyrin, and free porphyrins.However, the capacity to synthesize protochlorophyllideand protochlorophyllide phytyl ester is lost and the en-dogenous protochlorophylls gradually disappear. Mg proto-porphyrin monoester represents the terminal biosyntheticstep in this cell-free system.
Recently, Rebeiz et al. (28, 29) determined the time curves ofprotochlorophyllide and protochlorophyllide ester3 accumulationin etiolated cucumber cotyledons during etioplast biogenesis.They also reported a pronounced incorporation of 4-'C-Z-aminolevulinic acid into the tetrapyrrole moiety of phototrans-formable protochlorophyllide (28, 29). Kinetic analysis of specificradioactivities of the "4C-protochlorophyll pools suggested thatprotochlorophyllide and protochlorophyllide ester originate si-multaneously from a common precursor. The nature of thecommon precursor has not yet been determined and probably hasto await the elucidation of the terminal stages of protochlorophyllbiosynthesis between Mg protoporphyrin monomethyl ester andprotochlorophyll. The development of a cell-free system capableof protochlorophyll biosynthesis might facilitate such an under-
'This work was supported in part by Research Grant GM-07532from the United States Public Health Service.
2Present address: Department of Botany, University of California,Davis, California 95616.
3 The terminology proposed by Kirk and Tilney-Basset is followed inthis paper: the term "protochlorophyllide ester" denotes protochloro-phyllide phytyl ester, while the term "protochlorophyll" denotes themixture of protochlorophyllide and protochlorophyllide phytyl esterwhich accumulates in etiolated tissues.
standing. Hopefully, it would also shed some light upon theregulatory aspects of this problem.
Until now, most of the porphyrin biosynthetic investigations inplants have been carried out with unicellular algae and photo-synthetic bacteria.Rhodopseudomonas spheroides incubated with various sub-
strates accumulate a major fraction of coproporphyrin m; thelatter is accompanied sometimes by varying amounts of uropor-phyrins I and II (6, 10, 16). Rhodopseudomonas capsulata, incu-bated in the light with a mixture containing glycine and succinate,synthesizes large amounts of bacteriochlorophyll and two inter-mediates: coproporphyrinogen III and protoporphyrin mono-methyl ester (4). Richards and Rapoport have demonstrated, onthe other hand, that concentrated suspensions of Chlorobiumthiosulfatophilum-660 produce coproporphyrin III, uroporphyrinI and III, as well as porphyrins with 2, 3, 5, 6, and 7 carboxylgroups (30).
Carell and Kahn worked with isolated Euglena chloroplastsand reported that when the latter were incubated with ALA4 for10 to 18 hr they produced uroporphyrin and coproporphyrin, butno protoporphyrin (3). However, if the chloroplasts were dis-rupted they formed uroporphyrin, coproporphyrin, and proto-porphyrin (3).To our knowledge no one yet has investigated the protochloro-
phyll biosynthetic activity of a cell-free system from higher plants,although individual reactions of the metabolic pathway have beenstudied with crude homogenates (21).
This work reports on the tetrapyrrole biosynthetic activity ofetiolated cotyledons incubated with ALA and the effect of tissuedisorganization on the metabolic pathway of protochlorophyllbiosynthesis. A preliminary communication based on this re-search was presented to the First Science Meeting of the LebaneseAssociation for the Advancement of Sciences, Beirut, Lebanon, inNovember 1969 (25).
MATERIALS AND METHODS
Plant Material. Cucumber seed (Cucwnis sativus L. varietyAlpha green) were germinated in white sand at 28 C in completedarkness (22).
Chemicals and Radiochemicals. 14C-ALA was purchased fromN. V. Philips-Duphar, Amsterdam, Holland. The petroleumether used was a hydrocarbon fraction boiling from 60 to 80 C.
Preparation of the Crude Homogenate. Two grams of 4-day-oldetiolated cotyledons were macerated gently with mortar andpestle in 2 ml of 0.5 M sucrose, 0.2 M tris-HCl, pH 7.5, under a
4Abbreviations: ALA and 14C-ALA: 5-aminolevulinic and 414C-5-aminolevulinic acid.
543
www.plantphysiol.orgon January 8, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 467 1970
green safelight at 1 C. The slurry was filtered through four layersof cheesecloth. The resulting filtrate is called the crude homoge-nate fraction; it contains intact etioplasts and no detectable intactcells as evidenced by light and phase contrast microscopy. Totalproteins in this homogenate were determined by biuret as de-scribed previously (27).
Incubation of Etiolated Cotyledons with ALA. Three grams of4-day-old etiolated cotyledons were incubated in a total volumeof 5.55 ml of H20 containing 50,moles of K-ALA. The incuba-tion was carried for 16 to 20 hr in the dark at 28 C.
Incubation of Etiolated Cotyledons with "4C-ALA. Five eti-olated 4-day-old cotyledon pairs were incubated in a total volumeof1 ml of H20 containing 1,uc (20 m,umoles) of 14C-ALA. Theincubation was carried out in the dark at 28 C for periods of 30min to 18 hr.
Incubation of the Crude Homogenate with ALA. Ten millilitersof crude homogenate were incubated in the dark with 50,molesof ALA for 16 to 18 hr at 28 C. In a total volume of 10.65 ml,the incubation medium contained 50 Amoles of K-ALA, 10,umolesof MgCl2, 2 mmoles of tris-HCl (pH 7.5), 4 mmoles ofsucrose, and about 300 mg of proteins.
Incubation of the Crude Homogenate with14C-ALA. A volumeof 1.25 ml of crude homogenate was incubated in the dark with2,c (40 mMmoles) of14C-ALA at 28 C for periods of 20 min to2 hr. In a total volume of 1.35 ml the incubation medium con-tained 2,uc (40 m,umoles) 14C-ALA, 250,umoles of tris-HCl (pH7.5), 500,moles of sucrose, and about 38 mg of proteins.
Extraction of Uroporphyrin, Coproporphyrin, Protoporphyrin,and Protopheophytin. The extraction of porphyrins was monitoredby their red fluorescence under ultraviolet light of 366 nm. Threegrams of cotyledons or 5 ml of crude homogenate were blendedwith 50 ml of ethyl acetate:acetic acid (3:1, v/v) in a mechan-ically driven Ten-Broeck homogenizer. At this pH Mg phorbinsand Mg porphyrins lose their metal. The homogenate was subse-quently exposed to 40 ft-c of white fluorescent light at 30 C for30 min to oxidize porphyrinogens (33). After centrifugation, theresidue was washed twice with 10 ml of ethyl acetate:acetic acid(3:1, v/v) and the combined supernatant and washings werefractionated into uroporphyrin, coproporphyrin, and protopor-phyrin fractions by the method of Dresel and Falk (5). Proto-pheophytin remaining in the ether phase was extracted with 8 NHCI.
Extraction ofMg Porphyrins and Mg Phorbins. Mg porphyrinsand Mg phorbins were extracted in ether essentially as describedby Granick for Mg protoporphyrin monomethyl ester (9). Threegrams of cotyledons or 5 ml of the crude homogenate werehomogenized in 20 ml of acetone:0.1 N NH4OH (9:1, v/v) in anice-jacketed Ten-Broeck homogenizer. After centrifugation, thepellet was extracted with 10 ml of the acetone:NH40H mixture.The pooled supernatant and washings were extracted with 30 mlof petroleum ether to extract the /-carotene and protochlorophyl-lide ester. To the remaining acetone extract were added 1.2 ml ofa saturated NaCl solution and 0.3 ml of 0.5 M KH2PO4. Mgporphyrins and Mg phorbins were transferred to ether by re-peated ether extraction of the aqueous acetone phase. The etherextract was washed with 50 ml of cold H20 saturated withMgCO3. For the spectrophotometric determination of the totalprotochlorophyll content of the ether extract, the petroleum etherwash was omitted.
Extraction of '4C-Mg Porphyrins and '4C-Mg Phorbins fromWhole Cotyledons. Five pairs of '4C-labeled cotyledons werehomogenized in 10 ml of cold acetone containing MgCO3; the'4C-Mg porphyrins and '4C-Mg phorbins were extracted as de-scribed in a previous communication for '4C-protochlorophyllideand 14C-protochlorophyllide ester (29). After evaporation of theether extract, the residue containing the 14C-Mg porphyrins andphorbins was taken up in a minimal volume of benzene for
chromatography either on thin layers of Silica Gel H or onWhatman No. 3 MM paper.
Extraction of '4C-Porphyrins, '4C-Mg Porphyrins, and 14C-MgPhorbins from the Crude Homogenate. The 14C-Mg porphyrinsand 14C-Mg phorbins were extracted into ether from 1.35 ml ofcrude homogenate as described for the extraction of Mg por-phyrins and Mg phorbins and chromatographed either on thinlayers of Silica Gel H or on Whatman No. 3 MM paper. Allvolumes were adjusted to accomodate 1.35 ml of crude homoge-nate. The proteinaceous pellet and the aqueous acetone phasethat result from the extraction of the 14C-Mg porphyrins and4C-Mg phorbins were mixed, and the slurry was concentratedunder N2 to a thick paste. Eight milliliters of ethyl acetate :aceticacid (3:1, v/v) were then added, and the mixture was exposed to40 ft-c of white fluorescent light at 30 C for 30 min. The "4C-copro-porphyrin and 14C-protoporphyrin fractions were then extractedas described by Dresel and Falk (5), for chromatographicanalysis.
Conversion of Mg Porphyrins into Porphyrins. The Mg por-phyrin ether extract was thoroughly mixed with 0.5 to 1 ml ofaqueous 3 N HCI. The Mg porphyrins lost their metal and passedinto the aqueous acid phase. To transfer the porphyrins back toether, the acid phase was neutralized with solid sodium acetateand extracted several times with ether (12).
Hydrolysis of Mg Protoporphyrin Monoester. Mg protopor-phyrin monoester in ether was mixed with 0.5 to 1 ml of 7 N HCI.The ether was evaporated under N2, and the acid extract wasincubated in the dark for 20 hr at 4 C. To transfer the resultingprotoporphyrin and any unhydrolyzed protoporphyrin monoesterback into ether, the acid phase was neutralized with solid Naacetate and extracted several times with ether (12).
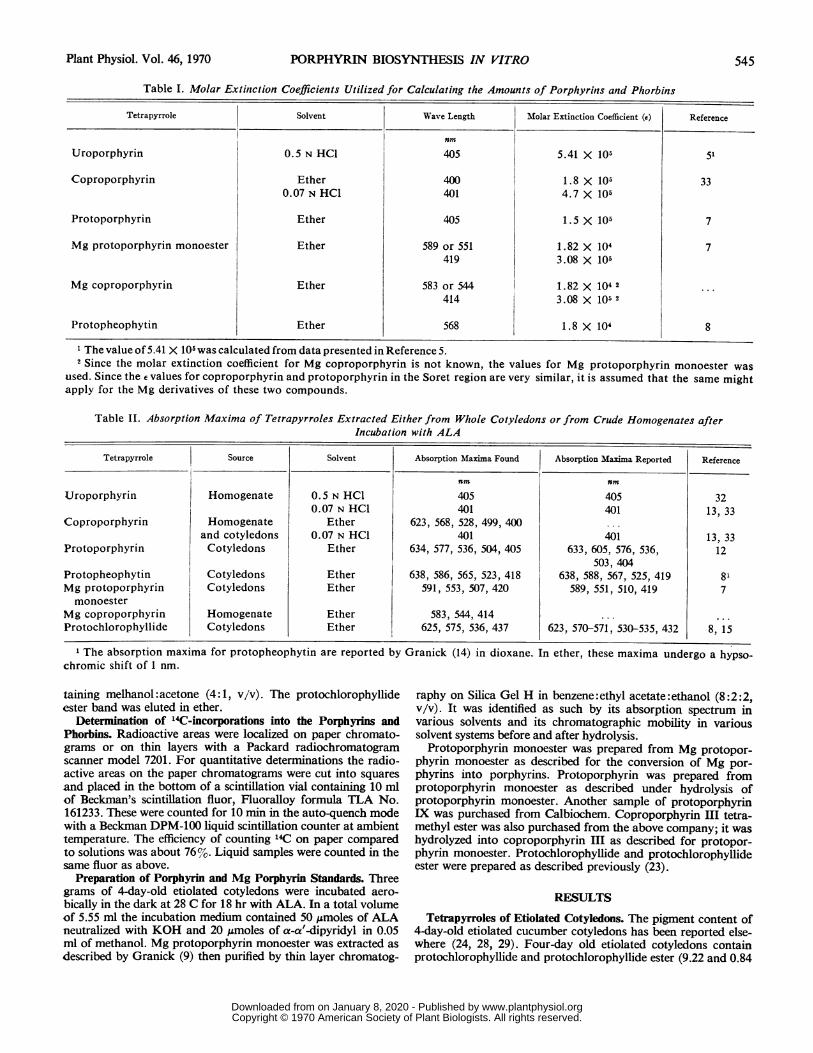
Spectrophotometric Determinations. Absorption spectra wererecorded with a Beckman model DB-G grating spectrophotom-eter coupled to a Beckman 10-inch linear log recorder. The wavelength calibration was checked against the double peak at 573and 586 nm of a Chance O.N. 12-glass Didymium filter. Inaddition the absorption maxima were corrected for the slightdiscrepancies between the actual wave length and the tracing ofthe recorder. A Unicam SP 500 spectrophotometer was utilizedfor all quantitative spectrophotometric determinations. The wavelength of the instrument was adjusted before use and all ab-sorbance readings were corrected for light scattering as reportedin a previous article (29). The amounts of porphyrins andphorbins were calculated from corrected absorbances and themolar extinction coefficients presented in Table I.
Separation of the Tetrapyrroles by Paper Chromatography.Free porphyrins were separated by ascending paper chromatog-raphy on Whatman No. 3 MM paper according to the number ofcarboxyl groups in lutidine:0.05 N NH40H (5:3.5, v/v) (9, 19,20) or in acetone:petroleum ether:acetic acid (3:7:0.01, v/v)(9). The chromatograms were developed in the dark at roomtemperature. Protoporphyrin monoester, Mg protoporphyrinmonoester, and Mg coproporphyrin were separated from oneanother and from contaminating coproporphyrin by ascendingchromatography on Whatman No. 3 MM paper in acetone:pe-troleum ether:acetic acid (3:7:0.01, v/v) at room temperaturein the dark (9).
Separation of Porphyrins and Phorbins on Thin Layers. Proto-chlorophyllide, protochlorophyllide ester, and Mg protopor-phyrin monoester were separated from one another and fromother porphyrins on thin layers of Silica Gel H. The pigmentswere quantitatively transferred to Silica Gel H plates in benzeneor ether, and the chromatograms were developed in benzene: ethylacetate:ethanol (8:2:2, v/v) at 4 C in the dark. After develop-ment and before drying, the "4C-Mg protoporphyrin monoesterand protochlorophyllide bands were scraped into a beaker con-
544 REBEIZ ET AL.
www.plantphysiol.orgon January 8, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.
PORPHYRIN BIOSYNTHESIS IN VITRO
Table I. Molar Extinction Coefficients Utilized for Calculating the Amounts of Porphyrins and Phorbins
Tetrapyrrole Solvent Wave Length Molar Extinction Coefficient (e) Reference
rnmUroporphyrin 0.5 N HCl 405 5.41 X 105 51
Coproporphyrin Ether 400 1.8 X 105 330.07 N HCl 401 4.7 X 105
Protoporphyrin Ether 405 1.5 X10| 7
Mg protoporphyrin monoester Ether 589 or 551 1.82 X 104 7419 3.08 X 105
Mg coproporphyrin Ether 583 or 544 1.82 X 104 2 ..
414 3.08 X 105 2
Protopheophytin Ether 568 1.8 X 104 8
1 The value of 5.41 X 105 was calculated from data presented in Reference 5.2 Since the molar extinction coefficient for Mg coproporphyrin is not known, the values for Mg protoporphyrin monoester was
used. Since the e values for coproporphyrin and protoporphyrin in the Soret region are very similar, it is assumed that the same mightapply for the Mg derivatives of these two compounds.
Table II. Absorption Maxima of Tetrapyrroles Extracted Either from Whlole Cotyledons or from Crude Homogenates afterIncubation with ALA
Tetrapyrrole Source Solvent Absorption Maxima Found Absorption Maxima Reported Reference
rnm rnmUroporphyrin Homogenate 0.5 N HCl 405 405 32
0.07 N HCI 401 401 13, 33Coproporphyrin Homogenate Ether 623, 568, 528, 499, 400 ...
and cotyledons 0.07 N HCl 401 401 13, 33Protoporphyrin Cotyledons Ether 634, 577, 536, 504, 405 633, 605, 576, 536, 12
503, 404Protopheophytin Cotyledons Ether 638, 586, 565, 523, 418 638, 588, 567, 525, 419 81Mg protoporphyrin Cotyledons Ether 591, 553, 507, 420 589, 551, 510, 419 7monoester
Mg coproporphyrin Homogenate Ether 583, 544, 414 ... ...
Protochlorophyllide Cotyledons Ether 625, 575, 536, 437 623, 570-571, 530-535, 432 8, 15
1 The absorption maxima for protopheophytin are reported by Granick (14) in dioxane. In ether, these maxima undergo a hypso-chromic shift of 1 nm.
taining melhanol:acetone (4:1, v/v). The protochlorophyllideester band was eluted in ether.
Determination of "4C-incorporations into the Porphyrins andPhorbins. Radioactive areas were localized on paper chromato-grams or on thin layers with a Packard radiochromatogramscanner model 7201. For quantitative determinations the radio-active areas on the paper chromatograms were cut into squaresand placed in the bottom of a scintillation vial containing 10 mlof Beckman's scintillation fluor, Fluoralloy formula TLA No.161233. These were counted for 10 min in the auto-quench modewith a Beckman DPM-100 liquid scintillation counter at ambienttemperature. The efficiency of counting 14C on paper comparedto solutions was about 76%. Liquid samples were counted in thesame fluor as above.
Preparation of Porphyrin and Mg Porphyrin Standards. Threegrams of 4-day-old etiolated cotyledons were incubated aero-bically in the dark at 28 C for 18 hr with ALA. In a total volumeof 5.55 ml the incubation medium contained 50 umoles of ALAneutralized with KOH and 20 jAmoles of a-a'-dipyridyl in 0.05ml of methanol. Mg protoporphyrin monoester was extracted asdescribed by Granick (9) then purified by thin layer chromatog-
raphy on Silica Gel H in benzene:ethyl acetate:ethanol (8:2:2,v/v). It was identified as such by its absorption spectrum invarious solvents and its chromatographic mobility in varioussolvent systems before and after hydrolysis.
Protoporphyrin monoester was prepared from Mg protopor-phyrin monoester as described for the conversion of Mg por-phyrins into porphyrins. Protoporphyrin was prepared fromprotoporphyrin monoester as described under hydrolysis ofprotoporphyrin monoester. Another sample of protoporphyrinIX was purchased from Calbiochem. Coproporphyrin III tetra-methyl ester was also purchased from the above company; it washydrolyzed into coproporphyrin III as described for protopor-phyrin monoester. Protochlorophyllide and protochlorophyllideester were prepared as described previously (23).
RESULTS
Tetrapyrroles of Etiolated Cotyledons. The pigment content of4-day-old etiolated cucumber cotyledons has been reported else-where (24, 28, 29). Four-day old etiolated cotyledons containprotochlorophyllide and protochlorophyllide ester (9.22 and 0.84
Plant Physiol. Vol. 46, 1970 545
www.plantphysiol.orgon January 8, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 46, 1970
F
XoCED
0MPE
Pide
/I\I \
I II
10
10x
0L
c)
'0
x
FIG. 1. Chromatographic mobility of the cotyledonary ether extract.The etiolated cotyledons were incubated for 18 hr with ALA and theether extract was chromatographed on Silica Gel H plates in benzene:ethyl acetate:ethanol (8:2:2, v/v) in presence of standard Mg proto-porphyrin monoester (MPE) and protochlorophyllide (Pide). 0:origin; F: front; I: ether extract; II: standard MPE and Pide; Xa:xanthophylls. The spots traced in broken lines indicate porphyrinscontaminating the ether extract.
Table III. Amounts of Tetrapyrroles and RF Values of the Copro-and Protoporphyrin Fractions Extracted from EtiolatedCucumber Cotyledons Incubated with 50 ,umoles of ALA
for 18 HrThe copro- and protoporphyrin fractions were separated as-
cendingly on Whatman No. 3 MM in 2,4-lutidine:0.05 N NH40H(5:3.5, v/v).
Tetrapyrrole Amount RF Values RF Values Refer-Obtained' Reported ence
m,mole/gfresh wt
Protochlorophyll2 10.81 ... ... ...
Mg protoporphyrin 0.18 .. ...
monoesterProtoporphyrin 8.36 0.77-0.83 0.81 20Coproporphyrin 2.62 0.50-0.58 0.54 20Uroporphyrin Trace
The range of RF values represents the spread of RF values forfive experiments.
2 Determined as protopheophytin.
mAmoles/g fresh weight, respectively). No porphyrins or Mgporphyrins are detected spectrophotometrically. The same is trueof excised cotyledons incubated with H20 for 18 hr. Theseobservations confirm the results of other workers (14) about thelack of accumulation of protochlorophyll biosynthesis inter-mediates in etiolated tissues.
Tetrapyrroles of Etiolated Cotyledons Incubated with ALA. Theether extract of cotyledons incubated for 18 hr with 50 jAmolesof ALA consists mainly of protochlorophyllide. The nature ofthe latter was confirmed by its pale green color under visiblelight, its absorption spectrum, and absorption maximum beforeacidification (Table II) as well as its "Oxorhodo" absorptionspectrum (11) after acidification.The ether extract also contained small quantities of Mg pro-
20 cm
FIG. 2. Radiochromatogram tracing of the cotyledonary ether ex-
tract cochromatographed with standard protochlorophyllide, proto-chlorophyllide ester, and Mg protoporphyrin monoester on Silica GelH in benzene:ethyl acetate:ethanol (8:2:2, v/v). The etiolated 4-day-old cotyledons were incubated in the dark at 28 C with 1 uc of 14C_ALA for: A: 20 min; B: 60 min; 0: origin; F: front; Xa: xanthophylls;C: carotene; MPE: Mg protoporphyrin monoester; Pide: protochloro-phyllide; Pide E: protochlorophyllide ester.
toporphyrin monoester separable from protochlorophyllide onSilica Gel H plates developed in benzene:ethyl acetate:ethanol(8:2:2, v/v). The identity of this compound was determined byits pink fluorescence under ultraviolet light as well as its absorp-tion spectrum and maxima between 500 and 600 nm (Table II).Its nature was further confirmed by a chromatographic mobilityon thin layers of Silica Gel H in benzene:ethyl acetate:ethanol(8:2:2) similar to standard Mg protoporphyrin monoester(Fig. 1).The porphyrins were extracted and fractionated according to
their acid affinity. Two major fractions were obtained: a "co-proporphyrin fraction" extractable in 0.1 N HCI and a "proto-porphyrin fraction" extractable in 3 N HCI. The latter possessesan "aetio" absorption spectrum in ether (11) and maxima simi-lar to those of protoporphyrin (Table II). Likewise, the copro-porphyrin fraction (5) exhibits an "aetio" absorption spectrumin ether, but the absorption maxima are different from those ofthe protoporphyrin fraction (Table II). Furthermore its Soretabsorption maximum in 0.1 N HC1 at 401 nm coincides withthat of coproporphyrin (13, 33).The nature of these free porphyrins was confirmed by their
paper chromatographic mobility in 2 ,4-lutidine:0.05 N NH4OH(5:3.5, v/v) (9, 19, 20) (Table III). Only a trace of uroporphyrincould be detected by its red fluorescence in the remaining aque-ous phase.
Table III presents the amounts in m,umoles per gram freshweight of cotyledons of the tetrapyrroles identified above.
Incorporation of 14C-ALA into the Tetrapyrroles of EtiolatedCucumber Cotyledons. Incorporation of 14C-ALA into por-phyrins and protochlorophyll became noticeable after 20 min ofincubation (Fig. 2A). After 60 min of incubation the radioactiveincorporations were already substantial as shown in Figure 2B.The latter also shows incorporation into Mg protoporphyrinmonoester. This is in contrast to the 24-hr incubations when noradioactivity could be detected in this compound (29). These
A
PideI0~~~~~~~~~~~
546 -REB-EIZ ETs AL.
www.plantphysiol.orgon January 8, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.

PORPHYRIN BIOSYNTHESIS IN VITRO
z0
40)
z< 50
H 60z
LJ 70-
00-
8c
90,/1
100 -700 600 500 400
X (mLLFIG. 3. Tracings of the absorption spectra of standard magnesium
protoporphyrin monoester in ether ( ) and of the ether extract ofthe crude homogenate incubated with ALA for 18 hr (---).
findings might indicate that, durng the early phase of incuba-tion with 1"C-ALA, the latter is rapidly converted into 14(KMgprotoporphyrin monoester, which accumulates until its laterconversion into protochlorophyll. This observation is also com-patible with the hypothesis formulated earlier (29) that proto-chlorophyll biosynthesis might be controlled at the Mg proto-porphyrin monoester level.The radioactivity at the origin of Figure 2, A and B, probably
represents l<: incorporation into protoporphyrin and copro-porphyrin since these two compounds accumulate in cotyledonsincubated with ALA (Table Ill).
Biosynthetic Activity of the Crude Homogenate Incubated withALA. After 18 hr of incubation in the absence of ALA, the crudehomogenate lost its original yellowish green color. Spectro-photometric determinations performed on the ether extractrevealed only trace amounts of protochlorophyll left. On theother hand, after 18 hr of incubation with ALA, the reactionmedium turned pink and emitted a strong red fluorescence underultraviolet light of 366 nm, indicating the accumulation of por-phyrins. The contribution of microorganisms borne by thecotyledonary surfaces to this tetrapyrrole biosynthesis was shownto be negligible. For an estimation of the bacterial contribution,6 g of whole etiolated cotyledons were incubated in 10 ml of thesame incubation medium used for the homogenate and underthe same conditions. At the end of incubation, the supernatantwas analyzed for porphyrins and phorbins synthesized by thebacterial population of the cotyledonary surfaces. No tetra-pyrroles were found.The ether extract of the reaction medium contained a com-
pound with two absorption bands of equal intensity between500 and 600 um, typical of metalloporphyrins (Fig. 3). However,its absorption maxima at 584, 544, and 414 nm were different
FIG. 4. Chromatography ofthe ether extract ofthe crude homogenateafter incubation with ALA for 18 hr. Whatman No. 3 MM paper wasused with the following solvents: A: in acetone:petroleum ether:acetic acid (3:7:0.01, v/v); B: in 2,4-lutidine:0.05 N NH4OH (5:3.5,v/v). The compounds represented by broken lines are present in traceamounts. I: ether extract before acidification; II: ether extract afteracidification; MPE: standard Mg protoporphyrin monoester; Co:standard coproporphyrin; P: standard protoporphyrin; 0: origin;F: front.
from those of magnesium protoporphyrin monoester at 591,553, and 420 nm (Fig. 3). Furthermore, the chromatographicmobility of this compound in acetone:petroleum ether:aceticacid (3:7:0.01, v/v) was different from that of standard Mgprotoporphyrin monoester (Fig. 4A). The chromatographicmobility of this metalloporphyrin in 2,4-lutidine :0.05 N NH40H(5:3.5, v/v) before and after acidification is presented in Figure4B. It is obvious that after acidification the RF value of thismetalloporphyrin decreases indicating the loss of a metal (Fig.4B). The resulting free porphyrin moves with an RF typical ofcoproporphyrin (Fig. 4B) and exhibits an absorption spectrumin ether typical of coproporphyrin (Table II). All these resultsindicate that the major compound in the ether extract of thereaction medium is a metallocoproporphyrin, most probablyMg coproporphyrin, in consideration of the large amounts ofMg2+ added to the reaction medium.The fractionation of the free porphyrins by the method of
Dresel and Falk (5) results in two major, red fluorescent frac-tions: a uroporphyrin fraction in sodium acetate and a copro-porphyrin fraction in 0.1 N HCI.The coproporphyrin nature of the 0.1 N HCI fraction was
confirmed as described in the preceding section by its absorp-tion spectrum in ether, its maxima in HCI and ether (Table II),and its chromatographic mobility in 2 ,4-lutidine :0.05 NNH40H(5:3.5, v/v).The uroporphyrin nature of the sodium acetate fraction was
confirmed by its chromatographic mobility in the 2,4-lutidine:0.05 N NH,OH system as well as its Soret absorption at 405nm in 0.5 N HCI. Both fractions were, however, slightly con-taminated by porphyrins, having probably six and five carboxylgroups, as evidenced from paper chromatography in 2 ,4-lutidine:0.05 N NH4OH (5:3.5 v/v). Only trace amounts of free proto-porphyrin could be detected in the reaction. The amounts oftetrapyrroles in the various fractions are presented in Table IV.
Biosynthetic Activity of the Crude Homogenate Incubated withYC-ALA. After 2 hr of incubation at 28 C in the presence of I
Plant Physiol. Vol. 46, 1970 547
www.plantphysiol.orgon January 8, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 46, 1970
Table IV. Tetrapyrroles Synthesized by the Crude HomogenateTen milliliters of crude homogenate were incubated in the dark
with 50.umoles of ALA for 16 hr at 28 C.
Tetrapyrrole Amount
mumoles /g protein
Uroporphyrin 38Coproporphyrin 57Mg coproporphyrin 28Protoporphyrin Trace
'0
MPE
F-e-Pide E- ---Xo--C- --Pide--- 020 cm
FIG. 5. Radiochromatogram tracing of the ether extract of the crudehomogenate incubated with 14C-ALA and cochromatographed onSilica Gel H in benzene:ethyl acetate :ethanol (8:2:2, v/v) with stand-ard protochlorophyllide, protochlorophyllide ester, and Mg proto-porphyrin monoester. The symbols are the same as in Figure 2.
,uc (20 m,moles) of 14C-ALA, the ether extract was chromato-graphed on Silica Gel H; both fast moving and slow movingtetrapyrroles became labeled (Fig. 5).The contribution of cotyledonary surface bacteria to the tetra-
pyrrole biosynthesis in this system was estimated; 800 mg ofwhole etiolated cotyledons were incubated in 1.35 ml of theincubation medium used for the homogenate and under thesame conditions. At the end of the incubation the cotyledonswere discarded and the reaction medium was analyzed for4C-porphyrins and "4C-phorbins. No radioactive tetrapyrroleswere found. Since a 2-hr incubation was sufficient for substan-tial incorporations into metallo and free porphyrins (Fig. 5), noneed was felt for longer incubations.The presence of a fast moving '4C-metalloporphyrin in the
ether extract of the reaction medium became apparent afterchromatography on Silica Gel H. Indeed, this compound ex-hibited a chromatographic mobility similar to standard Mg pro-toporphyrin monoester (Fig. 5). After elution from Silica GelH, the compound was mixed with standard Mg protoporphyrinmonoester and protoporphyrin monoester and chromatographedon paper in acetone: petroleum ether: acetic acid (3:7:0.01,v/v) before and after acidification. The results are presented inFigure 6. It is evident that the bulk of the radioactivity exhibitsthe same chromatographic mobility as Mg protoporphyrinmonoester and protoporphyrin monoester, respectively, beforeand after acidification (Fig. 6, A and B). After hydrolysis of theester group in 7 N HC1, most of the radioactivity moves on chro-matography with the hydrolysis products of the standards,protoporphyrin and small amounts of unhydrolyzed protopor-phyrin monoester (Fig. 6C). These results indicate that the'4C-metalloporphyrin present in the ether extract is "4C-Mg pro-toporphyrin monoester. The appearance of protoporphyrinmonoester after elution from Silica Gel H and subsequent chro-matography on paper (Fig. 6A) might be due to a loss of Mgduring elution and chromatography in the slightly acidic solvent
'0x
05;Ca_
Nl0
CE-
5
10
5
--W - - - 20 cm
FIG. 6. Radiochromatogram tracing of the 14C-metalloporphyrineluted from Silica Gel H, mixed with standard Mg protoporphyrinmonoester and protoporphyrin monoester and rechromatographed onWhatman No. 3 MM paper in acetone:petroleum ether:acetic acid(3:7:0.01, v/v). A: before acidification; B: after acidification; C: afterhydrolysis in 7 N HCI; PE: protoporphyrin monoester; P: protopor-phyrin. The other symbols are the same as in Figure 2.
Table V. '4C-Tetrapyrroles Synthesized by the Crude HomogenateA volume of 1.25 ml of crude homogenate was incubated in the
dark with 2 ,c (40 m,umoles) of 14C-ALA at 28 C for 2 hrs.
Tetrapyrrole 14C Incorporated
dpm/100 mg protein
Mg protoporphyrin monoester + 50,800protoporphyrin monoester
Protoporphyrin 1,490C3-porphyrin' 7,600Mg coproporphyrin 2,780Coproporphyrin2 12,400
1 Refers to a porphyrin with three carboxyl groups.2 Slightly contaminated with Mg coproporphyrin.
acetone:petroleum ether:acetic acid (3:7:0.01, v/v). The natureof the ester group has not been investigated, either in the stand-ard Mg protoporphyrin monoester or in the "4C-compound, butprobably consists of a methanol residue.
Incorporation of 14C into other tetrapyrroles was also investi-
10
5
c\J'0
0xC-)
A
<-PE-- -<-MPE--F 0
B
F <--PE--> 0
C
PE > 0F O
548 REBEIZ ET AL.
www.plantphysiol.orgon January 8, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.

PORPHYRIN BIOSYNTHESIS IN VITRO
gated. Beside Mg protoporphyrin monoester, smaller amountsof radioactivity were also incorporated into Mg coproporphyrin.Free porphyrins were also labeled, namely, coproporphyrin, pro-toporphyrin, and a porphyrin with three carboxyl groups. Theuroporphyrin pool was highly contaminated by 'IC-ALA, andno efforts were made to purify it. Table V presents the incor-poration of 14C-ALA into the pools mentioned above.
DISCUSSION
The ability of etiolated cotyledonary tissue to synthesize pro-tochlorophyll appears to be lost upon homogenization. How-ever, the crude, cell-free homogenate can still accumulate copro-porphyrin, uroporphyrin, and Mg coproporphyrin upon pro-longed incubation with ALA (Table IV). During this incubation,the endogenous protochlorophyll present in the extract is gradu-ally broken down by an unknown mechanism. If the incubationis carried out in the presence of radioactive ALA, it is possibleto demonstrate the formation of Mg protoporphyrin monoester(Table V). This compound has been postulated as an inter-mediate of protochlorophyll biosynthesis in higher plants (1, 2,4, 9, 12, 21, 31), but does not accumulate normally in wholecotyledonary tissue. Thus, Mg protoporphyrin monomethylester seems to constitute the last step of the protochlorophyllbiosynthetic pathway in this crude cell-free system. Recentresults also indicate that etioplasts isolated by differential cen-trifugation incorporate 14C-ALA into Mg protoporphyrinmonomethyl ester, although to lesser extent than the crudehomogenate (Rebeiz and Castelfranco, unpublished). Theaccumulation of Mg coproporphyrin might be attributed to achemical or a nonspecific enzymatic chelation of Mg2+ (17, 18)by the accumulated coproporphyrin. The occurrence of metallo-coproporphyrin (31), metallophorphyrins with three to eightcarboxyl groups (31), free porphyrins with three to seven car-boxyl groups (3, 30), and a free porphyrin with three carboxylgroups (33) have been reported in the literature.The biosynthetic activity of the crude cotyledonary homoge-
nate is considerably more limited than that of intact cotyledons.Nevertheless, it does show certain similarities with the biosyn-thetic activity of isolated Euglena chloroplasts, unicellular algae,and photosynthetic bacteria (3, 4, 6, 10, 16, 30), suggesting thatthe same biochemical pathway for the synthesis of chlorophyllprecursors which has been found in lower organisms is operatingalso in higher plants.
Recent results indicate that in the presence of the proper co-factors, glutathione, methyl alcohol, Mg'+, and inorganic phos-phate (26), the same crude homogenate is able to incorporate'4C-ALA into protochlorophyll. However, the incorporation of4C-ALA into "4C-protochlorophyll remains much lower than inthe intact cotyledons. This might suggest that structural dis-organization of enzymes and cofactor destruction and dilutionduring the homogenization interfere with the simultaneous andefficient biosynthesis of the lipid, protein, and pigment compo-nents of a specific organelle membrane.
LITERATURE CITED
1. BoGoRAD, L. 1965. Chlorophyll biosynthesis. In: T. W. Goodwin, ed., Chemistryand Biochemistry of Plant Pigments. Academic Press, New York. pp. 29-74.
2. BocoRAD, L. 1966. The blosynthesis of chlorophylls. In: L. P. Vernon and G. R.Seely, eds., The Chlorophylls. Academic Press, New York. pp. 481-510.
3. CARELL, E. F. AND J. S. KAHN. 1964. Synthesis of porphyrins byisolatedchloroplastsof Euglena. Arch. Biochem. Biophys. 180: 1-6.
4. COOPER, R. 1963. The biosynthesis of coproporphyrinogen, magnesium proto-
porphyrin monomethyl ester and bacteriochlorophyll by Rhodopseudomonascapsulata. Biochem. J. 89: 100-108.
5. DRESEL, E. I. B. AND E. FALK. 1956. Studies on the biosynthesis of blood pigments.2. Haem and porphyrin formation in intact chicken erythrocytes. Biochem. J. 63:72-79.
6. GIBSON, K. D., A. NEUBERGER, AND G. H. TMTs. 1962. Studies on the biosynthesisof porphyrin and bacteriochlorophyll by Rhodopseudomonas spheroides. 2. Theeffects of ethionine and threonine. Biochem. J. 83: 550-559.
7. GRANICK, S. 1948. Magnesium protoporphyrin as a precursor of chlorophyll inChlorella. J. Biol. Chem. 175: 333-342.
8. GRANicK, S. 1950. Magnesium vinyl pheoporphyrin as, another intermediate in thebiological synthesis of chlorophyll. J. Biol. Chem. 183: 713-730.
9. GRANsCK, S. 1961. Magnesium protoporphyrin monoester and protoporphyrinmonomethyl ester in chlorophyll biosynthesis. J. Biol. Chem. 236: 1168-1172.
10. HEATH, H. AND D. S. HOARE. 1959. The biosynthesis of porphyrins from porpho-bilinogen by Rhodopseudomonas spheroides. Biochem. J. 72: 14-22.
11. HOLT, A. S. 1965. Nature properties and distribution of chlorophylls. In: T. W.Goodwin, ed., Chemistry and Biochemistry of Plant Pigments. Academic Press,New York, pp. 3-28.
12. JONEs, D. T. G. 1963. The production of magnesium protoporphyrin monomethylester by Rhodopseudomonas spheroides. Biochem. J. 86: 429-432.
13. JoPE, E. M. AND J. R. P. O'BRIEN. 1945. Spectral absorption and fluorescence ofcoproporphyrin isomers I and III and the melting points of their methyl esters.Biochem. J. 39: 239-245.
14. KIRK, J. T. 0. AND R. A. E. TILNEY-BAsSr. 1968. In: The Plastids. Freeman, Lon-don and San Francisco. pp. 402-576.
15. KosKm, V. M. AND J. H. C. SMITH. 1968. The isolation and spectral absorptionproperties of protochlorophyll from etiolated barley seedlings. J. Amer. Chem.Soc. 70: 3558-3562.
16. LASCELLES, JUNE. 1956. The synthesis of porphyrins and bacteriochlorophyll by cellsuspensions of Rhodopseudomonas spheroides. Biochem. J. 62: 78-93.
17. Lnrrr, H. N. AND M. L. KELSsY. 1964. Incorporation of ZnII into protoporphyrinby extracts from barley. Fed. Proc. 23: 223.
18. NEUBERGER, A. AND G. H. TAIT. 1964. Studies on the biosynthesis of porphyrinand bacteriochlorophyll by Rhodopseudomonas spheroides. 5. Zinc-protopor-phyrin chelatase. Biochem. J. 90: 607-616.
19. NICHOLAS, R. E. H. AND C. RJMNGTON. 1949. Qualitative analysis of the porphy-rins by partition chromatography. Scand. J. Clin. Lab. Invest. 1: 12-18.
20. NICHOLAS, R. E. H. AND C. RIMINGTON. 1951. Paper chromatography of porphy-rins: some hitherto unrecognized porphyrins and further notes on the method.Biochem. J. 48: 306-309.
21. RADMER, R. J. AND L. BOGORAD. 1967. (-) S-Adenosyl-L-methionine-magnesiumprotoporphyrin methyl transferase, an enzyme in the biosynthetic pathway ofchlorophyll in Zea mays. Plant Physiol. 42: 463-465.
22. REBEIZ, C. A. 1967. Studies on chlorophyll biosynthesis in etiolated excised cotyle-dons of germinating cucumber at different stages of seedling development.Magon, Serie Scientifique 13: 1-21.
23. REBEIZ, C. A. 1968. The chloroplast pigments of etiolated and greening cucumbercotyledons. Magon, Serie Scientifique 21: 1-25.
24. REBEIz, C. A. 1968. Dark and light carotenoids accumulation in etiolated and green-ing cucumber cotyledons. Magon, Serie Scientifique 23: 1-10.
25. REBEIZ, C. A. AND M. ABou HAIDAR. 1969. Biosynthese du Mg protoporphyrinemonoester a partir de l'acide i-aminolevulinique par un systeme extracellulaire.First Science Meeting of the Lebanese Association for the Advancement ofScience, Beirut, Lebanon. pp. 21-22.
26. REBEIZ, C. A. AND P. CAsTELFRANCO. 1970. Protochlorophyllide biosynthesis in acell-free system from higher plants. Plant Physiol. 45: 5226.Annual Meetings, Bloomington, Inc. 46: 000-00
27. REBEIz, C. A., P. CASTELFRANCO, AND A. H. ENGELBRECHT. 1965. Fractionationand properties of an extra-mitochondrial enzyme system from peanuts catalyz-ing the ,-oxidation of palmitic acid. Plant Physiol. 40: 281-286.
28. REBEIz, C. A., M. YAGHI, AND M. ABou HAiDAR. 1969. Etioplast pigment accumulation in germinating cucumber cotyledons. XI International Botanical Congress,Seattle, Wash. p. 178.
29. RERaaz, C. A., M. YAGHI, M. ABou-HAIDAR, AND P. CAsTELFRANcO. 1970. Pro-tochlorophyll biosynthesis in cucumber (Cucumis sativus, L.) cotyledons. PlantPhysiol. 46: 57-63.
30. RIcHARDs, W. R. AND H. RAPOPORT. 1966. The biosynthesis of Chlorobium chloro-phylls-660. The isolation and purification of porphyrins from Chlorobium thio-sulfatophilum-660. Biochemistry 5: 1079-1089.
31. RIcHARDs, W. R. AND H. RAPOPORT. 1967. The biosynthesis of Chlorobium chloro-phylls-660. The production of magnesium protoporphyrin monomethyl ester,bacteriochlorophyll, and Chlorobium pheoporphyrins by Chlorobium thiosulfato-philum-660. Biochemistry 6: 3830-3839.
32. RIn,NGTON, C. AND S. L. SvEiNssoN. 1950. The spectrophotometric determinationof uroporphyrin. Scand. J. Clin. Lab. Invest. 2: 209-216.
33. SANO, S. AND G. GRANICK. 1961. Mitochondrial coproporphyrinogen oxidase andprotoporphyrin formation. J. Biol. Chem. 236: 1173-1180.
549Plant Physiol. Vol. 46, 1970
www.plantphysiol.orgon January 8, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.