plant growth promoting bacteria and mycorrhizal …
TRANSCRIPT

Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
1
PLANT GROWTH PROMOTING BACTERIA AND
MYCORRHIZAL ON VARIETIES OF Sorghum spp.
GERMINATION UNDER STRESSING ABIOTIC CONDITIONS †
[BACTERIAS PROMOTORAS DEL CRECIMIENTO VEGETAL
Y MICORRIZAS SOBRE LA GERMINACION EN VARIEDADES
DE Sorghum spp. BAJO CONDICIONES DE ESTRESES
ABIÓTICOS]
Ramón Jaime Holguin Peña1, Juan Manuel Vargas López2,
Guadalupe Amanda López Ahumada2, Francisco Rodríguez Félix2,
Alberto Olguín Moreno2, Carlos Borbón Morales3,
Juan Pedro López Córdova4 and Edgar Omar Rueda-Puente4*
1 Centro de Investigaciones Biológicas del Noroeste, Mar Bermejo No.
195, Col. Playa Palo Santa Rita, La Paz, Baja California Sur CP 23090. 2 Universidad de Sonora, Departamento de Investigación y Posgrado en
Alimentos, Colonia centro Hermosillo, Sonora, México CP 83000. 3 Centro de Investigación en Alimentación y Desarrollo AC,
Coordinación de Desarrollo Regional, Departamento de Economía.
Carretera a la Victoria Km. 0.6, Sonora, México. CP 83304. 4 Universidad de Sonora, Departamento de Agricultura y Ganadería,
Carretera a Bahía de Kino, km 20.5, Hermosillo, Sonora, México, 83000.
Email. [email protected]
* Corresponding author
SUMMARY
Background. Mexico currently ranks first in Sorghum production. Notwithstanding, soils in a productive
area of Sonora are too poor in organic matter with saline intrusion and high temperatures. Farmers apply
synthetic fertilizers but the indiscriminate use increase salinity. Under dry and arid zones, plant growth
promoting bacteria (PGPB) and arbuscular mycorrhizal fungi (AMF) signifies a substitute to obtain
nutrients. Objective. New and productive varieties of Sorghum in Sonora desert (Silo Miel 370 VCB (370),
Sorgo Silo Miel 315 VC (315), Sorgo Silo Miel II (II) and Camino 526 Croplan winfield (526)) were
evaluated inoculated with a species PGPB and AMF. Methodology. Under salinity, temperature regime
(night/ day) in germination stage, tests were carried out considering the inoculation with PGPB and AMF;
growth variables were measured like a percentage and rate, plant height, root length, and produced biomass
(fresh and dry matter). Results. The cultivar 526 was the most outstanding with the PGPB and AMF
evaluated; at the seedling stage, there were significant differences between the treatments for plant height,
root length, fresh and dry weight, considering NaCl levels and temperature regimes. Implication. Studies
of the association of PGPB and AMF with the Sorghum cv. 526 are recommended under field conditions.
Conclusion. This study is the first step to obtain an ideal variety of Sorghum, which is a principal crop in
dry arid and desert zones of México. This kind of study promotes beneficial microorganisms as a resource
to increase the development and productivity of plants.
Keywords: temperature; salinity; basic crops.
RESUMEN
Antecedentes. Actualmente, México ocupa el primer lugar en producción de sorgo. No obstante, los suelos
en un área productiva de Sonora son demasiado pobres en materia orgánica con intrusión salina y altas
temperaturas. Los agricultores aplican fertilizantes sintéticos, pero el uso indiscriminado aumenta la
salinidad. En zonas áridas, las bacterias promotoras del crecimiento vegetal (PGPB) y los hongos
micorrícicos arbusculares (AMF) representan un sustituto para obtener nutrientes en los cultivos. Objetivo.
Se evaluaron las variedades nuevas y productivas del desierto de Sonora (Silo Miel 370 VCB (370), Sorgo
Silo Miel 315 VC (315), Sorgo Silo Miel II (II) y Camino 526 Croplan winfield (526)) al inocularse con
una especie de PGPB y AMF. Metodología. Bajo condiciones de salinidad, un régimen de temperatura
(noche / día) en la etapa de germinación, se realizaron pruebas considerando la inoculación con la PGPB y
† Submitted August 27, 2019 – Accepted January 6, 2020. This work is licensed under a CC-BY 4.0 International License.
ISSN: 1870-0462.

Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
2
el AMF; las variables de crecimiento se midieron como porcentaje y tasa de germinación, altura de la
plántula, longitud de la raíz y biomasa producida (materia fresca y seca). Resultados. El cultivar 526 fue
el más sobresaliente con la PGPB y el AMF evaluados; en la etapa de plántula existieron diferencias
significativas entre los tratamientos para la altura de la planta, longitud de la raíz, el peso fresco y seco
considerado los niveles de NaCl y los regímenes de temperatura. Implicaciones. Estudios de la asociación
de PGPB y AMF con el sorgo var. 526 se recomiendan en condiciones de campo. Conclusión. Este estudio
es el primer paso para obtener una variedad ideal de sorgo, que es un cultivo principal en zonas secas, áridas
y desérticas de México. Este tipo de estudio promueve microorganismos benéficos como un recurso para
aumentar el desarrollo y la productividad de las plantas.
Palabras clave: temperatura; salinidad; cultivo básico.
INTRODUCTION
Sorghum (Sorghum bicolor) is one principal
cereal crop in many countries; it contains high
levels of minerals and vitamins, i.e. those of
group B are listed thiamine, riboflavin, and
niacin (López-Ortíz et al. 2011). This richness
makes it an important antioxidant food. While it
is also rich in iron, calcium, phosphorus, zinc and
insoluble fiber (de Morais et al. 2017). Sorghum
(subsp. bicolor) belongs to the family Gramineae
and it is sowed in countries of America (Mexico,
Argentina, and the USA), Africa (where the
Sorghum is originated), Asia and Oceania
(Ethiopia, Sudan; Chine; Australia,
respectively). Sorghum is a protein-rich cereal
and stands out for containing a lower caloric
intake, with fewer carbohydrates and fats.
Besides, it is a gluten-free food, so it is optimal
for celiac people. Besides, it has antidiarrheal and
homeostatic properties. Currently, new markets
for Sorghum are developed producing novel
functional foods. Mexico currently ranks first in
Sorghum production with 11 million tons per
year. Likewise, our country has reached the
world record in yield of Sorghum of up to 17 tons
per hectare, reflecting that this crop is one of the
most productive crops.
Sonora and Baja California Peninsula are some
of the aridest states of Mexico, with 80 mm
average annual precipitation (Rueda et al. 2004).
The main problems facing agriculture are a
declining supply of water and a decrease in the
quality of the water. While the total world
population today demands proportionally more
and more food, that total is expected to exceed
9,000 million by 2050. In the agricultural sector,
many adverse factors reduce productivity, i.e.
water extraction in excess, use of inappropriate
fertilizers, which have caused increases in the
salinity of agriculture soil (SAGAR 1981; Salim
1989; Shalaby et al. 1993; Sangakkara et al.
1995; Abdullah et al. 2001; Rashid et al. 1999).
In this sense in dry arid and desert zones, Novo
production alternatives are developing
concerning the selection and evaluation of salt-
tolerant plants that already are adapted to salty
areas (Flowers and Yao 1995; Flowers 2004).
Studies with a salt-tolerant plant have been
carried out in an agronomic, physiological,
biochemical, and molecular sense. However, in
recent years, the number of works that address
tolerance to salinity from a germinations stage,
point of view has increased considerably, to
search responses under different conditions,
because the variability is extensive (Abdus et al.
1999; Acosta-Motos et al. 2017; Miranda et al.
2017; Chen et al. 2018; Yan et al. 2018; Dilnur
et al. 2019; Hernández 2019; Zhong et al. 2019).
However, although there are many Sorghum
varieties with a high viable seed, the germination
and establishment are poor under the soils of the
Sonora region (Zhang et al. 2019). In this sense,
farmers due the climatic conditions are favorable
to produce Sorghum, they invest in agro-inputs
and technology, so in the Mexican Sonora state,
the Sorghum has a wide distribution along with
the central parts of Sonora state (Zhang et al.
2019).
Moreover, the productivity of Sorghum is limited
by the absence of accessible nitrogen (de Morais
et al. 2017; SAGAR 1981). This situation
disturbs its development and reproduction
possible (Juarez and Harrison 2018).
Traditionally, growers apply artificial fertilizers
to recompense for soil nitrogen deficit. However,
unselective use of these nourishments forces the
increasing salinity and severely injuries soil
microorganism's structure and configuration
(Jefferies 1977; Kapulnik et al. 1981; Banwari
and Rao 1990; Nahid and Gomah 1991).
Few studies on plant growth-promoting bacteria
(PGPB) have been published of Sorghum spp
(Akhavan et al. 1991; Isopi et al. 1995; Luigi et
al. 1998; Compant et al. 2010), but not in
association with mycorrhizas diversity within the
rhizosphere. Concerning varieties sowed in the
Sonora desert, the studies are null. So is very
important to increase the number of known salt-
tolerant, nitrogen-fixing bacteria and arbuscular
mycorrhizal (Hamdi 1999; Whipps 2000).
On the other hand, several principal stressing
factors as a temperature (Adams et al. 2001;

Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
3
Aksouh et al. 2006; Luo 2011), and salinity
(Pichu 2010; Kroes and Supit 2011) has been
proved in glicophytes, which regulate some
stages of the plants (germination, pre-flowering,
flowering and mature physiologic) and interact
within the soil interface (Ungar 1995; Laynez-
Garsaball et al. 2007). The osmotic and matrix
potential in saline soils decreases the temperature
that is effective for seed germination (Hegarty
1978). The adverse factor such as salinity
interacting with another abiotic factor such as
temperature, determine high germination
percentages (Khan and Ungar 1998; Rivers and
Weber 1971; Philipupiya and Ungar 1984;
Keiffer and Ungar 1995; Khan and Ungar 1996;
Khan and Ungar 1998; Gulzar et al. 2000); it is
worth mentioning that, the ability of plants to
counteract adverse factors is conditioned by
multiple biochemical and physiological
pathways that help to better capture cellular
water; physiological protection of chloroplasts,
maintaining a homeostasis of the ions. Plants to
detoxify radicals under salinity conditions
activate to essential routes such as the synthesis
of osmotically active metabolites, specific
proteins and certain free radical elimination
enzymes that maintain a balance and flow of ions
and water; they also favor and contribute a
function in the elimination of oxygen radicals or
chaperones; some species accumulate
methylated metabolites, whose function is
osmoprotectant and eliminate free radicals.
Photorespiration is a physiological phenomenon
which it is altered under abiotic conditions (Asish
and Anath 2005). In the case of halophyte seeds,
they can be viable for prolonged periods when
they are in hypersaline conditions, allowing them
to germinate when the conditions of salinity in
the soil and water are reduced (Khan et al. 1976;
Woodell 1985; Khan and Ungar 1997; Khan and
Ungar 1999).
Notwithstanding the foregoing, it is important to
indicate that among salinity tolerant plants, there
is variability to respond positively and negatively
to adverse conditions varying conditions such as
temperature and salinity (Khan and Ungar 1997;
Gul and Weber 1999). The present study defines
the influence of the inoculation with the PGPB
(Bacillus amyloliquefaciens) and the AMF
(Glomus intraradices) under different conditions
of salinity and discontinuous temperature
regimes on the seed germination of varieties of
Sorghum bicolor.
MATERIALS AND METHODS
Effect on germination and seedling growth of
four varieties of Sorghum spp. by inoculants
in NaCl conditions
Seeds of four varieties of Sorghum spp. were
collected, from production areas of Sonora,
Northwest of Mexico. The collected varieties
were Silo Miel 370 VCB (370), Sorgo Silo Miel
315 VC (315), Sorgo Silo Miel II (II) and
Camino 526 CROPLAN WINFIELD (526).
Largest and health seeds were carefully chosen
and with no observable destruction. Seeds of
each variety were disinfected in sodium
hypochlorite (3% active chlorine) for 10 s, and
washed with sterilized and distilled water and
dried with sterile paper. The halo bacterium B.
amyloliquefaciens (Accession number
KJ433614), on N-free OAB agar medium
according to (Renganathan et al. 2019), with
three different concentrations of NaCl (0.25 M)
was replicated. Later, at a concentration of 108
cells.mL-1, seeds were inoculated with bacteria
treatments according to (Carrillo et al. 1998). At
the same time and according to (Strullu and
Romand 1986; Declerck et al. 1998; Bécard and
Piché 1992; Bécard and Fortin 1988), and using
Allium schoenoprasum was reproduced Glomus
intraradices. The inoculum obtained was from a
sample of AM 120 Granite Seed Company (Lehi,
UT 84043). Furthermore, seeds were treated with
the AMF-G. intraradices according to (Rueda-
Puente et al. 2018), using a concentration of 120
propagules.cm3. Into the sterilized Petri dishes,
germination tests were performed, each Petri
dish with a cotton layer substrate (150 x 15 mm)
covering the bottom of the dish. Dishes were
moistened with uniform NaCl solution (0, 0.06,
or 0.12 M). All seeds varieties considering its
treatment and separated form were done inside a
growth chamber at 27°C ± 0.5°C and 35% ± 1%
RH, with continuous white light. Every four
days, 20 mL of the appropriate solution was
added to each dish. When the radicle was at least
2 mm long from seeds, were considered
germinated. Final percentage germination was
measured after 25 days; also the number of
germinated seeds was recorded daily
(germination rate). Using the next formula, the
germination rate was calculated (Maguire 1962):
M=n1/t1 + n2/t2 +…n25/t25
where n1, n2, … n25 are the numbers of
germinated seeds at times t1, t2, … t25 in days.
The hierarchic experiment of a randomized
design included three factors (varieties,
inoculation, and salinity), with five replicates of
50 seeds. The first factor (variety) had four levels
(varieties: 370, 315, II and 526); three
bioinnoculations: without inoculation,
inoculation with mycorrhiza arbuscular (Glomus
intraradices) and inoculation with the bacterium
Bacillus amyloliquefaciens were the second
factor; and the three concentrations of NaCl (0,
0.06, and 0.12 M) were the third factor. The three

Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
4
studied factors combination turned out 36
treatments. Arcsine transformation was realized
to obtain percentage germination (56), with
three-way analyses of variance (ANOVA).
Germination rates were obtained by sums of
germinations per day and were not transformed
before analysis. Duncan’s multiple range test at
P = 0.05 was carried out to obtain significant
differences between means of treatments. Data
were analyzed using the Statistical Analysis
System (Sokal and James 1962). Seedling
growth of four varieties under all conditions was
measured by dry and fresh weights on 30 days
after inoculation (dai); other variables calculated
were root length and height and dry weight.
Colony-forming units (CFU) was a measurement
in the root system from seedlings considering
Carrillo et al. (Bashan et al. 2007). In all
variables least significant differences (LSD)
were obtained by Duncan’s Multiple Range Test
at P = 0.05. All statistical analysis was done with
SAS (2004).
Estimation of Bacillus amyloliquefaciens, and
Glomus intraradices, and NaCl on germination
and seedling growth of the superior variety
526 of Sorghum spp
As a second experiment, variety 526 was carried
out to evaluate the effect of temperature on
germination alternating temperature regimes of
5-15 (night-day), 10-20, 15-25, 20-30, and 25-
35°C, based on a 24-hr cycle of 12 hr (Growth
chambers with Sylvania cool white fluorescent
lamps, 25 μmol m-2 s-1, 400-750 nM). Seeds
were submitted at the same treatment
(disinfected and inoculated with PGPB and AMF
treatments under NaCl solutions (0, 0.06, and
0.12 M) as it was indicated in the first
experiment. Variables as percentage
germination, germination rates, root length,
height, and dry and fresh weights were evaluated
as previously cited in the first phase. All data
were analyzed using the Statistical Analysis
System (Maguire 1962; SAS 2004). A hierarchic
experiment of randomized was designed with
three factors (inoculation, temperature regime,
and salinity); same three levels as a 1st
experiment were analyzed: a) without inoculants,
with inoculant PGPB (B. amyloliquefaciens), and
AMF (Glomus intraradices); three
concentrations of NaCl (0, 0.06, and 0.12 M), and
c: five temperature regimes (5-15, 10-20, 15-25,
20-30, and 25-35 °C, based on a 24-hr cycle of
12 hr).
RESULTS
Germination and seedling growth of four
Sorghum varieties
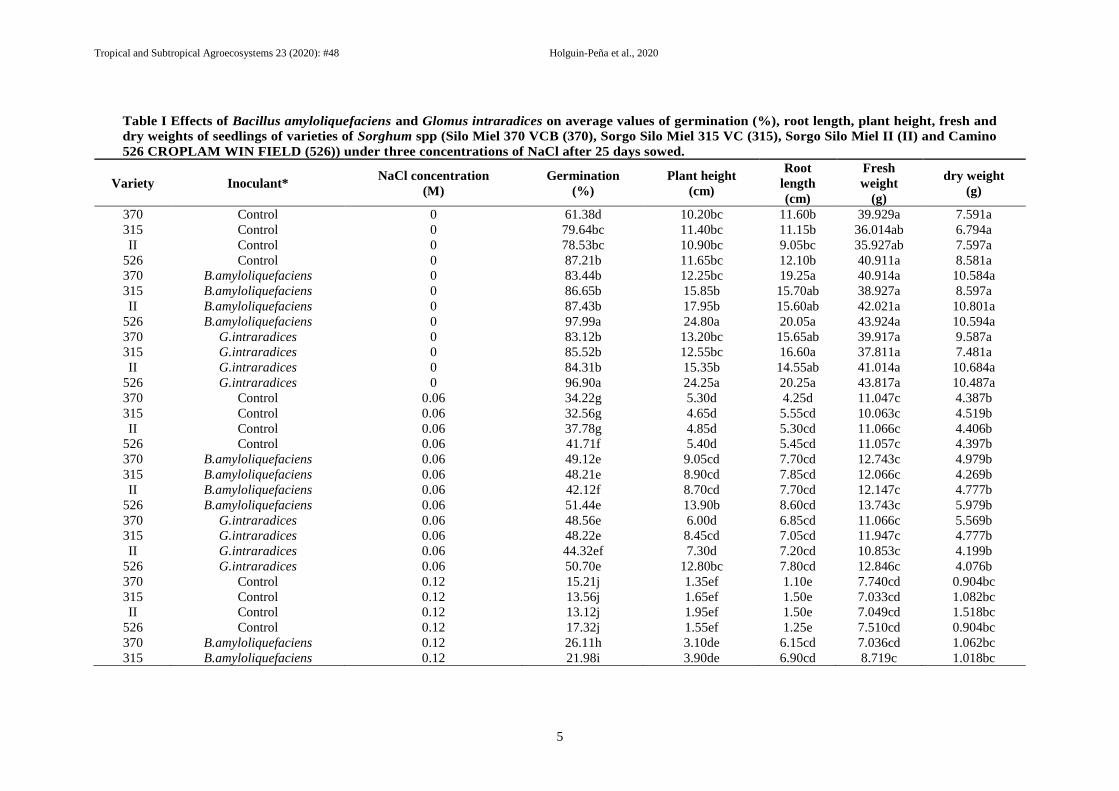
The germination percentage was significantly
modified by biotreatments studied in this
experiment (B. amyloliquefaciens and Glomus
intraradices), under salinity conditions (Table I).
When salinity was higher (0.12 M NaCl),
inhibition of germination was observed in all
varieties non inoculated (< 80%), compared with
those inoculated, sticking out the 526 variety
with B. amyloliquefaciens. However, when
salinity conditions were reduced at 0 M NaCl),
germination percentage was increasing until 97%
in 526 Var. + with both beneficial
microorganisms. The contrary was observed in
varieties non biotreated, obtaining significant
germination results in 76% at 0 M, and obtaining
high values the 526 var. Similar behavior was
showed in 0.06 M of NaCl, where 526 var.,
showed the highest values in germination with
and even without Plant Growth promoting
Bacteria (PGPB) and arbuscular mycorrhizal
fungi (AMF). According to the germination rate,
results (data no-showed) of all varieties with bio-
inoculants studied, showed a total germination
(after inoculation and sowed – AIS-), among 8 to
17 days =d= (at 0 M NaCl = 9d with a 99.50%;
at 0.06 M = 11d with 55%, and 0.12 M NaCl =
17 d with no more than 43%), while those non
inoculated showed germination total rate till
11dAIS to 20d; at 0M with a 82.35%; at 0.06
with 43% in 16d AIS, and no more than 42% at
0.12 M NaCl in 20d, respectively. PGPB and
AMF influenced plant growth of all varieties,
during early seedling with significant differences
among treatments for plant height, root length,
fresh and dry weight (Table I). These results
evidenced the potential on 526 var. to be selected
and used in the next step. All high values were
obtained at 0 M of NaCl with B.
amyloliquefaciens and Glomus intraradices, and
highlighting the same bionoculants at 0.06 and
0.12 M vs treatments without beneficial
microorganisms.
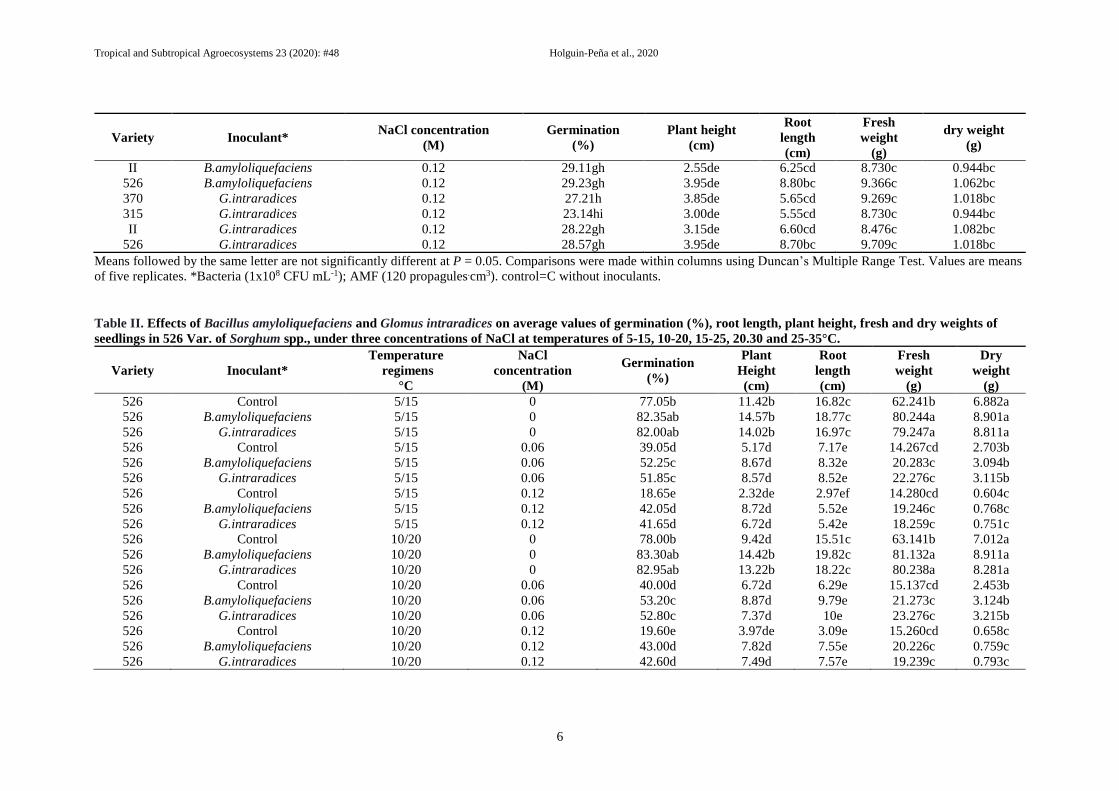
Evaluation of B. amyloliquefaciens and
Glomus intraradices inoculants, NaCl levels,
and temperature regimes on germination and
seedling growth of 526 variety
After inoculation with PGPB and AM during the
germination stage under NaCl levels and
temperature regimes, results showed significant
differences among treatments (Table II). Optimal
germination of 526 var. in 12/12 hr light/dark
conditions occurred under all salinity conditions,
were obtained at 0 M NaCl (P>0.05), and in these
all treatments whit bio inoculants (P>0.05), but
no significant between temperatures 5/15, 10/20,
15/25, and 25/35 °C (Table II).
Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
5
Table I Effects of Bacillus amyloliquefaciens and Glomus intraradices on average values of germination (%), root length, plant height, fresh and
dry weights of seedlings of varieties of Sorghum spp (Silo Miel 370 VCB (370), Sorgo Silo Miel 315 VC (315), Sorgo Silo Miel II (II) and Camino
526 CROPLAM WIN FIELD (526)) under three concentrations of NaCl after 25 days sowed.
Variety Inoculant* NaCl concentration
(M)
Germination
(%)
Plant height
(cm)
Root
length
(cm)
Fresh
weight
(g)
dry weight
(g)
370 Control 0 61.38d 10.20bc 11.60b 39.929a 7.591a
315 Control 0 79.64bc 11.40bc 11.15b 36.014ab 6.794a
II Control 0 78.53bc 10.90bc 9.05bc 35.927ab 7.597a
526 Control 0 87.21b 11.65bc 12.10b 40.911a 8.581a
370 B.amyloliquefaciens 0 83.44b 12.25bc 19.25a 40.914a 10.584a
315 B.amyloliquefaciens 0 86.65b 15.85b 15.70ab 38.927a 8.597a
II B.amyloliquefaciens 0 87.43b 17.95b 15.60ab 42.021a 10.801a
526 B.amyloliquefaciens 0 97.99a 24.80a 20.05a 43.924a 10.594a
370 G.intraradices 0 83.12b 13.20bc 15.65ab 39.917a 9.587a
315 G.intraradices 0 85.52b 12.55bc 16.60a 37.811a 7.481a
II G.intraradices 0 84.31b 15.35b 14.55ab 41.014a 10.684a
526 G.intraradices 0 96.90a 24.25a 20.25a 43.817a 10.487a
370 Control 0.06 34.22g 5.30d 4.25d 11.047c 4.387b
315 Control 0.06 32.56g 4.65d 5.55cd 10.063c 4.519b
II Control 0.06 37.78g 4.85d 5.30cd 11.066c 4.406b
526 Control 0.06 41.71f 5.40d 5.45cd 11.057c 4.397b
370 B.amyloliquefaciens 0.06 49.12e 9.05cd 7.70cd 12.743c 4.979b
315 B.amyloliquefaciens 0.06 48.21e 8.90cd 7.85cd 12.066c 4.269b
II B.amyloliquefaciens 0.06 42.12f 8.70cd 7.70cd 12.147c 4.777b
526 B.amyloliquefaciens 0.06 51.44e 13.90b 8.60cd 13.743c 5.979b
370 G.intraradices 0.06 48.56e 6.00d 6.85cd 11.066c 5.569b
315 G.intraradices 0.06 48.22e 8.45cd 7.05cd 11.947c 4.777b
II G.intraradices 0.06 44.32ef 7.30d 7.20cd 10.853c 4.199b
526 G.intraradices 0.06 50.70e 12.80bc 7.80cd 12.846c 4.076b
370 Control 0.12 15.21j 1.35ef 1.10e 7.740cd 0.904bc
315 Control 0.12 13.56j 1.65ef 1.50e 7.033cd 1.082bc
II Control 0.12 13.12j 1.95ef 1.50e 7.049cd 1.518bc
526 Control 0.12 17.32j 1.55ef 1.25e 7.510cd 0.904bc
370 B.amyloliquefaciens 0.12 26.11h 3.10de 6.15cd 7.036cd 1.062bc
315 B.amyloliquefaciens 0.12 21.98i 3.90de 6.90cd 8.719c 1.018bc
Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
6
Variety Inoculant* NaCl concentration
(M)
Germination
(%)
Plant height
(cm)
Root
length
(cm)
Fresh
weight
(g)
dry weight
(g)
II B.amyloliquefaciens 0.12 29.11gh 2.55de 6.25cd 8.730c 0.944bc
526 B.amyloliquefaciens 0.12 29.23gh 3.95de 8.80bc 9.366c 1.062bc
370 G.intraradices 0.12 27.21h 3.85de 5.65cd 9.269c 1.018bc
315 G.intraradices 0.12 23.14hi 3.00de 5.55cd 8.730c 0.944bc
II G.intraradices 0.12 28.22gh 3.15de 6.60cd 8.476c 1.082bc
526 G.intraradices 0.12 28.57gh 3.95de 8.70bc 9.709c 1.018bc
Means followed by the same letter are not significantly different at P = 0.05. Comparisons were made within columns using Duncan’s Multiple Range Test. Values are means
of five replicates. *Bacteria (1x108 CFU mL-1); AMF (120 propagules.cm3). control=C without inoculants.
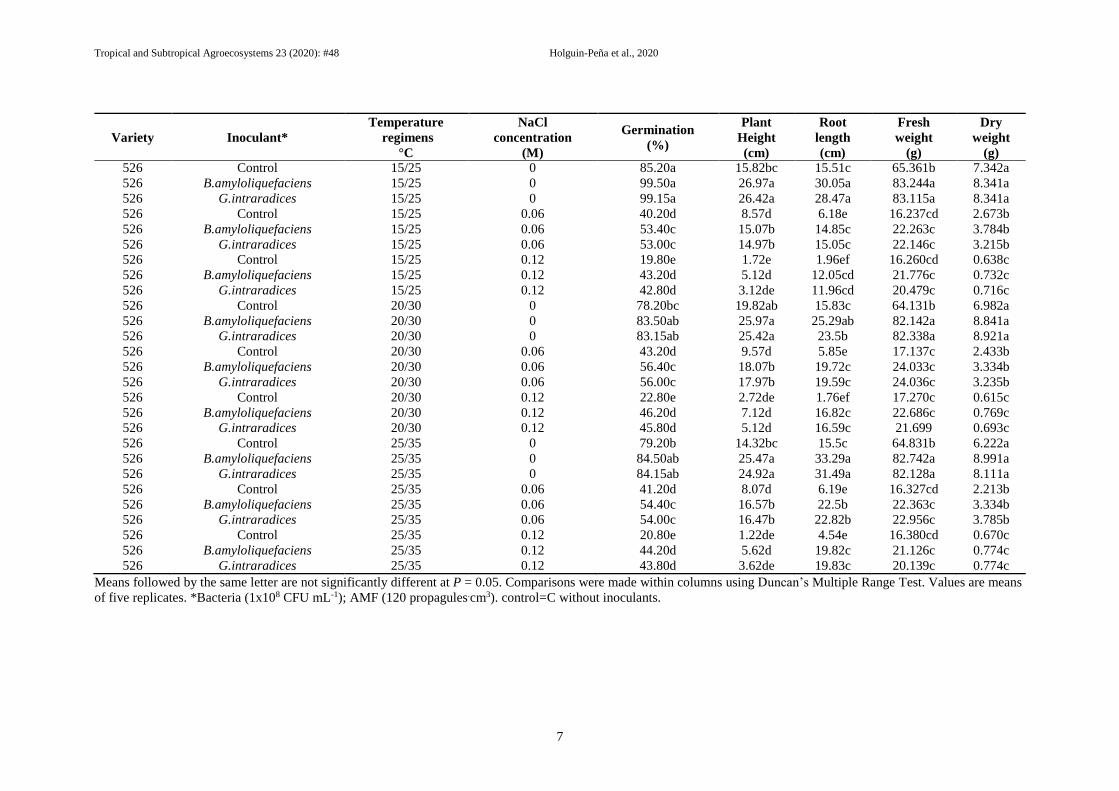
Table II. Effects of Bacillus amyloliquefaciens and Glomus intraradices on average values of germination (%), root length, plant height, fresh and dry weights of
seedlings in 526 Var. of Sorghum spp., under three concentrations of NaCl at temperatures of 5-15, 10-20, 15-25, 20.30 and 25-35°C.
Variety Inoculant*
Temperature
regimens
°C
NaCl
concentration
(M)
Germination
(%)
Plant
Height
(cm)
Root
length
(cm)
Fresh
weight
(g)
Dry
weight
(g)
526 Control 5/15 0 77.05b 11.42b 16.82c 62.241b 6.882a
526 B.amyloliquefaciens 5/15 0 82.35ab 14.57b 18.77c 80.244a 8.901a
526 G.intraradices 5/15 0 82.00ab 14.02b 16.97c 79.247a 8.811a
526 Control 5/15 0.06 39.05d 5.17d 7.17e 14.267cd 2.703b
526 B.amyloliquefaciens 5/15 0.06 52.25c 8.67d 8.32e 20.283c 3.094b
526 G.intraradices 5/15 0.06 51.85c 8.57d 8.52e 22.276c 3.115b
526 Control 5/15 0.12 18.65e 2.32de 2.97ef 14.280cd 0.604c
526 B.amyloliquefaciens 5/15 0.12 42.05d 8.72d 5.52e 19.246c 0.768c
526 G.intraradices 5/15 0.12 41.65d 6.72d 5.42e 18.259c 0.751c
526 Control 10/20 0 78.00b 9.42d 15.51c 63.141b 7.012a
526 B.amyloliquefaciens 10/20 0 83.30ab 14.42b 19.82c 81.132a 8.911a
526 G.intraradices 10/20 0 82.95ab 13.22b 18.22c 80.238a 8.281a
526 Control 10/20 0.06 40.00d 6.72d 6.29e 15.137cd 2.453b
526 B.amyloliquefaciens 10/20 0.06 53.20c 8.87d 9.79e 21.273c 3.124b
526 G.intraradices 10/20 0.06 52.80c 7.37d 10e 23.276c 3.215b
526 Control 10/20 0.12 19.60e 3.97de 3.09e 15.260cd 0.658c
526 B.amyloliquefaciens 10/20 0.12 43.00d 7.82d 7.55e 20.226c 0.759c
526 G.intraradices 10/20 0.12 42.60d 7.49d 7.57e 19.239c 0.793c
Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
7
Variety Inoculant*
Temperature
regimens
°C
NaCl
concentration
(M)
Germination
(%)
Plant
Height
(cm)
Root
length
(cm)
Fresh
weight
(g)
Dry
weight
(g)
526 Control 15/25 0 85.20a 15.82bc 15.51c 65.361b 7.342a
526 B.amyloliquefaciens 15/25 0 99.50a 26.97a 30.05a 83.244a 8.341a
526 G.intraradices 15/25 0 99.15a 26.42a 28.47a 83.115a 8.341a
526 Control 15/25 0.06 40.20d 8.57d 6.18e 16.237cd 2.673b
526 B.amyloliquefaciens 15/25 0.06 53.40c 15.07b 14.85c 22.263c 3.784b
526 G.intraradices 15/25 0.06 53.00c 14.97b 15.05c 22.146c 3.215b
526 Control 15/25 0.12 19.80e 1.72e 1.96ef 16.260cd 0.638c
526 B.amyloliquefaciens 15/25 0.12 43.20d 5.12d 12.05cd 21.776c 0.732c
526 G.intraradices 15/25 0.12 42.80d 3.12de 11.96cd 20.479c 0.716c
526 Control 20/30 0 78.20bc 19.82ab 15.83c 64.131b 6.982a
526 B.amyloliquefaciens 20/30 0 83.50ab 25.97a 25.29ab 82.142a 8.841a
526 G.intraradices 20/30 0 83.15ab 25.42a 23.5b 82.338a 8.921a
526 Control 20/30 0.06 43.20d 9.57d 5.85e 17.137c 2.433b
526 B.amyloliquefaciens 20/30 0.06 56.40c 18.07b 19.72c 24.033c 3.334b
526 G.intraradices 20/30 0.06 56.00c 17.97b 19.59c 24.036c 3.235b
526 Control 20/30 0.12 22.80e 2.72de 1.76ef 17.270c 0.615c
526 B.amyloliquefaciens 20/30 0.12 46.20d 7.12d 16.82c 22.686c 0.769c
526 G.intraradices 20/30 0.12 45.80d 5.12d 16.59c 21.699 0.693c
526 Control 25/35 0 79.20b 14.32bc 15.5c 64.831b 6.222a
526 B.amyloliquefaciens 25/35 0 84.50ab 25.47a 33.29a 82.742a 8.991a
526 G.intraradices 25/35 0 84.15ab 24.92a 31.49a 82.128a 8.111a
526 Control 25/35 0.06 41.20d 8.07d 6.19e 16.327cd 2.213b
526 B.amyloliquefaciens 25/35 0.06 54.40c 16.57b 22.5b 22.363c 3.334b
526 G.intraradices 25/35 0.06 54.00c 16.47b 22.82b 22.956c 3.785b
526 Control 25/35 0.12 20.80e 1.22de 4.54e 16.380cd 0.670c
526 B.amyloliquefaciens 25/35 0.12 44.20d 5.62d 19.82c 21.126c 0.774c
526 G.intraradices 25/35 0.12 43.80d 3.62de 19.83c 20.139c 0.774c
Means followed by the same letter are not significantly different at P = 0.05. Comparisons were made within columns using Duncan’s Multiple Range Test. Values are means
of five replicates. *Bacteria (1x108 CFU mL-1); AMF (120 propagules.cm3). control=C without inoculants.
Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
8
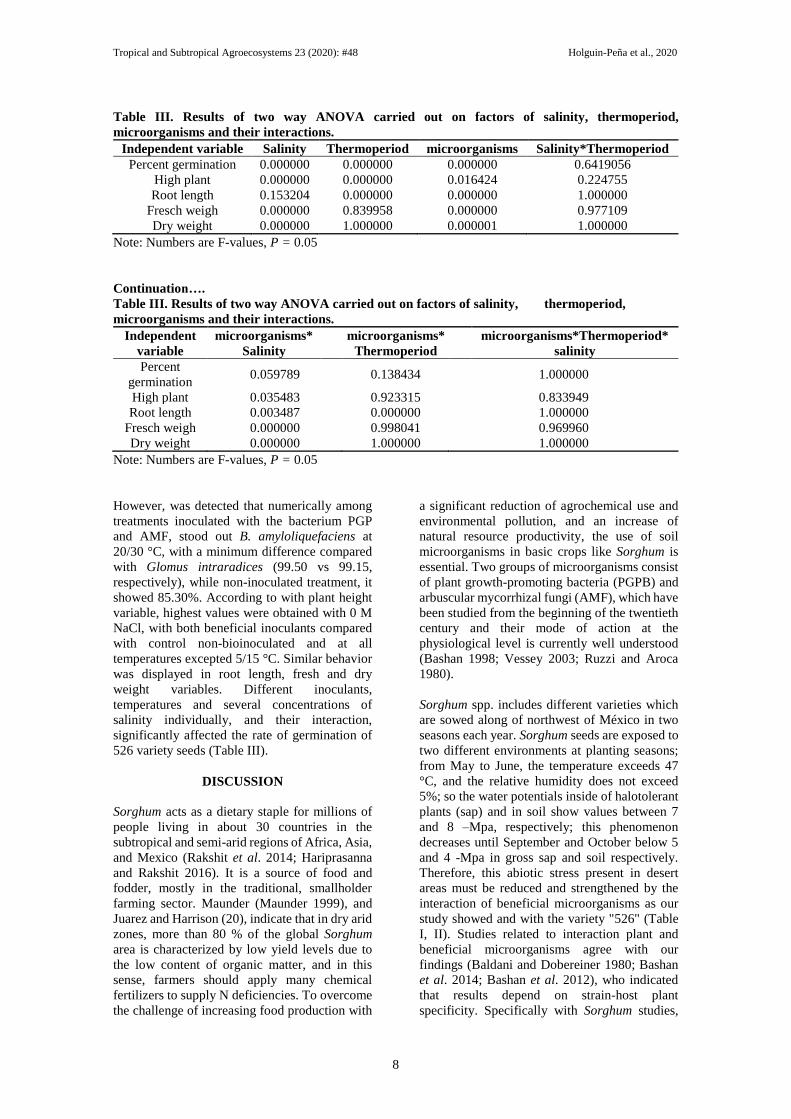
Table III. Results of two way ANOVA carried out on factors of salinity, thermoperiod,
microorganisms and their interactions.
Independent variable Salinity Thermoperiod microorganisms Salinity*Thermoperiod
Percent germination 0.000000 0.000000 0.000000 0.6419056
High plant 0.000000 0.000000 0.016424 0.224755
Root length 0.153204 0.000000 0.000000 1.000000
Fresch weigh 0.000000 0.839958 0.000000 0.977109
Dry weight 0.000000 1.000000 0.000001 1.000000
Note: Numbers are F-values, P = 0.05
Continuation….
Table III. Results of two way ANOVA carried out on factors of salinity, thermoperiod,
microorganisms and their interactions.
Independent
variable
microorganisms*
Salinity
microorganisms*
Thermoperiod
microorganisms*Thermoperiod*
salinity
Percent
germination 0.059789 0.138434 1.000000
High plant 0.035483 0.923315 0.833949
Root length 0.003487 0.000000 1.000000
Fresch weigh 0.000000 0.998041 0.969960
Dry weight 0.000000 1.000000 1.000000
Note: Numbers are F-values, P = 0.05
However, was detected that numerically among
treatments inoculated with the bacterium PGP
and AMF, stood out B. amyloliquefaciens at
20/30 °C, with a minimum difference compared
with Glomus intraradices (99.50 vs 99.15,
respectively), while non-inoculated treatment, it
showed 85.30%. According to with plant height
variable, highest values were obtained with 0 M
NaCl, with both beneficial inoculants compared
with control non-bioinoculated and at all
temperatures excepted 5/15 °C. Similar behavior
was displayed in root length, fresh and dry
weight variables. Different inoculants,
temperatures and several concentrations of
salinity individually, and their interaction,
significantly affected the rate of germination of
526 variety seeds (Table III).
DISCUSSION
Sorghum acts as a dietary staple for millions of
people living in about 30 countries in the
subtropical and semi-arid regions of Africa, Asia,
and Mexico (Rakshit et al. 2014; Hariprasanna
and Rakshit 2016). It is a source of food and
fodder, mostly in the traditional, smallholder
farming sector. Maunder (Maunder 1999), and
Juarez and Harrison (20), indicate that in dry arid
zones, more than 80 % of the global Sorghum
area is characterized by low yield levels due to
the low content of organic matter, and in this
sense, farmers should apply many chemical
fertilizers to supply N deficiencies. To overcome
the challenge of increasing food production with
a significant reduction of agrochemical use and
environmental pollution, and an increase of
natural resource productivity, the use of soil
microorganisms in basic crops like Sorghum is
essential. Two groups of microorganisms consist
of plant growth-promoting bacteria (PGPB) and
arbuscular mycorrhizal fungi (AMF), which have
been studied from the beginning of the twentieth
century and their mode of action at the
physiological level is currently well understood
(Bashan 1998; Vessey 2003; Ruzzi and Aroca
1980).
Sorghum spp. includes different varieties which
are sowed along of northwest of México in two
seasons each year. Sorghum seeds are exposed to
two different environments at planting seasons;
from May to June, the temperature exceeds 47
°C, and the relative humidity does not exceed
5%; so the water potentials inside of halotolerant
plants (sap) and in soil show values between 7
and 8 –Mpa, respectively; this phenomenon
decreases until September and October below 5
and 4 -Mpa in gross sap and soil respectively.
Therefore, this abiotic stress present in desert
areas must be reduced and strengthened by the
interaction of beneficial microorganisms as our
study showed and with the variety "526" (Table
I, II). Studies related to interaction plant and
beneficial microorganisms agree with our
findings (Baldani and Dobereiner 1980; Bashan
et al. 2014; Bashan et al. 2012), who indicated
that results depend on strain-host plant
specificity. Specifically with Sorghum studies,

Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
9
our results are strengthened with those found by
Sarig et al. (Sarig et al. 1992), Issopi et al.
(1995), James et al. (1997), Stein et al. (1997),
they obtained favourable results with the
inoculation of beneficial microorganisms like a
vesicular-arbuscular mycorrhizas and
Acetobacter diazotrophicus; Herbaspirillum, as
a endophytic diazotroph colonizing vascular
tissue in leaves; Azospirillum brasilense
inoculation on growth dynamics and hydraulic
conductivity; and the contribution of nitrogen
fixation by Azoarcus sp. BH72 in Sorghum,
respectively. However, disagree with Arsac et al.
(1990), Okon and Labandera (1994), who stated
that the effect is not strain-dependent among
different plant species.
Our study shows that low temperatures reduce
germination; the contrary was observed in
germination percentage with an increase in
temperature (Table 2). For its part, while the
salinity was reduced, the germination was
increased. At 15 °C and under 0.12 M NaCl, our
526 var. germinated and showed tolerance to
salinity; however the germination of this seed
was more effective when we inoculated PGPB;
results agreeing with others results in others
crops (Khan 1991); Khan and Weber (1986),
reported maximum salinity tolerance ranges
from 200 to 1720 mM NaCl in halophytes such
as S. bigelovii, S. europaea, S. stricta, Cressa
cretica, and Suaeda moquinii. These reports
indicate that 526 variety could be classified into
the salt-tolerant one during germination.
In this 2nd phase, the temperature increasing
germination percentage in 2 and 13% at 20/30
and 15/25, respectively, compared with the 1st
phase using 27 °C in a constant form (at 0 M
NaCl and both bio-inoculants). Moreover, the
same variety in the other variables (plant height,
root length, fresh and dry weight variables), its
values were significant vs control non inoculated
when seeds were inoculated with PGPB and
AMF. Considering a relationship between all the
variables with the different temperatures and
salinity studied, some plants showed variability
in their length and weight in their roots and whole
plant. This is attributed to the capacity of the
plant to tolerate in its interior ions and the
production of glycophytes and osmolytes, which
are have a function of maintaining an ionic
homeostasis between several tissues and
intracellular compartmentalization; effects
visualized in desert species of the Great Basin,
subtropical maritime deserts of Pakistan,
Haloxylon recurvum, Zygophyllum simplex and
Arthrocnemum macrostachyum (Khan and
Ungar 1997; Khan 1991; Gul, B.; Weber 1999),
respectively.
All varieties studied in this study and especially
“526” obtained, was benefit using beneficial
microorganisms by B. amyloliquefaciens and G.
intraradices abilities. Other studies indicate
positive effects using plant growth-promoting
microorganisms (Afek et al. 1990), results that
coincide with those obtained in the present study,
appreciating that the PGPB of this study and
experiencing it with var. 526 Sorghum, positively
was affected. These effects are possible to the
participation of responsible substances in the
growth of plants (Afek et al. 1990; Puente
Bashan 1993; Goodfriend et al. 2000; Díaz et al.
2001). Similarly, some studies support G.
intraradices in the promotion of plant
development (Melgares et al. 2004), results that
are consistent with the findings in this evaluation.
Similar results were obtained for other plants and
beneficial microorganisms (90). However, other
assays although they were carried out with other
plants and other beneficial microorganisms, also
some inhibitive effects on germination were
observed (Díaz et al. 2001).
Plants exposed to saline stress change their
environment, and one of the first vegetative
phases is germination. In this first stage, osmotic
stress begins to manifest itself, mainly by
stopping the emergence of the radicle and its
subsequent roots. The recoil is associated with an
accumulation of ionic stress that generates
phytotoxicity, and showing an imbalance in
nutrient intake and absorption (Asish and Anath
2005), oxidative stress at the subcellular level,
mediated by reactive oxygen species (ROS)
(Acosta-Motos et al. 2017), mainly. These
responses to salinity in plants tolerant to NaCl,
implement adaptations so as not to be affected by
salinity and carry out their survival. There are
mechanisms inside the cells that promote an
adaptation that is reflected in their morphology,
physiology, biochemistry and molecular
(Acosta-Motos et al. 2017). In the germination
stage, among the first existing metabolic actions
to tolerate salt, it is determined by multiple routes
that influence in water retention and / or
acquisition, protecting organelles and
maintaining homeostasis. Given this situation,
that the seeds or new plants are subject, the
beneficial microorganisms as a PGPB or AMF,
release bio substances that generate hormonal
changes within the plants, the production of
volatile organic, benefiting in the taking of
nutrients and a tolerance to abiotic stress such as
salinity (Ruzzi and Aroca 2015). Same authors
indicate that oxidative stress in different phases
in the plant is linked to the electron transport
chain in membranes of mitochondria and
chloroplast during ATP production. Therefore,

Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
10
numerous energetically demanding
physiological processes like germination and
seedling development are associated with
oxidative stress (Bailly 2004). It is well known
that mitochondria are the principal subcellular
compartment of oxygen consumption and the
principal source of reactive oxygen species
(ROS) (Puntarulo et al. 1998). Most of the ATP
required for early stages of development comes
from mitochondria; therefore, generation of ROS
during this process is mainly associated to
cellular respiration. At later stages of
development, photophosphorylation in
chloroplast becomes the main source of ATP to
cover the energy requirement for proper
development; then, chloroplast also becomes a
primary source of ROS (Asada 2006), and it is
possible that the release of hormones by
beneficial microorganisms inside and attached to
plants, the production of volatile organic
compounds, help to improve nutrient intake and
improve tolerance to abiotic stress (García et al.
2018).
CONCLUSIONS
This study is the first step to obtain an ideal
variety of Sorghum, which is a principal crop in
dry arid and desert zones of México. This kind of
study promotes beneficial microorganisms as a
resource to increase the development and
productivity of plants. Moreover, studies with the
interaction of B. amyloliquefaciens and G.
intraradices with 526 var. of Sorghum are
necessary to decide the magnitude to which these
observations can be observed and reproduced
under one production system.
Acknowledgments Thanks to personal technicians of Laboratories in
CIBNOR. In memorial of Dr. Rogelio Ramirez
SerranoT at CIBNOR, QEPD.
Funding. We thank to Universidad de Sonora by
Project “Effect of promoting halobacteries of
vegetable growth in the ecophysiology and
production of the Sorghum crop under saline
conditions”, key: USO313006248.
Conflict of interest. The authors declared no
conflict of interest.
Data availability. Data are available with the
corresponding author
([email protected]) upon reasonable
request
Compliance with ethical standards. There was
strict adherence to the Global code of conduct for
research in resource-poor settings following the
Convention on Biological Diversity and
Declaration of Helsinki.
REFERENCES
Abdullah, Z., Khan, M., Flowers, J. 2001. Causes
of Sterility in Seed set of Rice under
Salinity Stress. Journal Agronomy and
Crop Science. 187: 25-32.
https://doi.org/10.1046/j.1439-
037X.2001.00500.x
Abdus, S., Hollington P.A., Gorham, J., Wyn, J.
and Gliddon, C. 1999. Physiological
genetics of salt tolerance in wheat
(Triticum aestivum L): Performance of
wheat varieties, inbred lines and
reciprocal F1 hybrids under saline
conditions. Journal Agronomy and
Crop Science. 183: 145-56.
https://doi.org/10.1046/j.1439-
037x.1999.00361.x
Acosta-Motos, J., Ortuño, M., Bernal-Vicente,
A., Diaz-Vivancos, P., Sanchez-Blanco,
M. and Hernandez, J. 2017. Plant
Responses to Salt Stress: Adaptive
Mechanisms. Agronomy. 7:18-22.
https://doi.org/10.3390/agronomy7010
018
Adams, S., Cockshull, K. and Cave, C. 2001.
Effect of temperature on the growth and
development of tomato fruits. Annals
Botany. 88: 869-877.
https://doi.org/10.1006/anbo.2001.152
4
Afek, U., Rinaldelli, A., Menge, J., Jonson, E.
and Pond, E. (1990). Mycorrhizal
species, root age, and position of
mycorrhizal inoculum influence
colonization of cotton, onion, and
pepper seedlings. Journal American
Society Horticulture Science. 115(6):
938- 942.
Akhavan, K., Campbel, W., Jurinak, J.and
Dudley I. 1991. Effects of CaSO4,
CaCl2, and NaCl on leaf nitrogen,
nodule weight, and acetylene reduction
activity in Phaseolus vulgaris L. Arid
Soil Research and Rehabilitation. 5: 97-
103.
Aksouh-Harradj, N., Campbell, L. and Mailer,
R. 2006. Canola response to high and
moderately high temperature stresses
during seed maturation. Canadian
Journal Plant Science. 86: 967-980.
10.3389/fpls.2018.01224
Arsac, J., Lamothe, C., Mulard, D. and Fages, J..
1990. Growth enhancement of maize

Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
11
(Zea mays L) through Azospirillum
lipoferum inoculation: effect of plant
genotype and bacterial concentration.
Agronomie. 10: 640-654.
https://hal.archives-ouvertes.fr/hal-
00885325/document
Asada, K. 2006. Production and scavenging of
active oxygen in chloroplast and their
functions.
Plant Physiology.141:391-396. DOI:
https://doi.org/10.1104/pp.106.082040
Asish, K. and Anath, B. 2005. Salt tolerance and
salinity effects on plants: a review.
Ecotoxicology and Environmental
Safety. 60(3): 324-349.
https://doi.org/10.1016/j.ecoenv.2004.0
6.010
Bailly, C. 2004. Active oxygen species and
antioxidants in seed biology. Seed
Science Reserach. 14: 93-107. DOI:
https://doi.org/10.1079/SSR2004159
Baldani, V. and Dobereiner, L.1980. Host- plant
specificity in the infection of cereals
with Azospirillum spp. Soil Biology and
Biochemestry. 12: 443-439. DOI:
10.1016/0038-0717(80)90021-8
Banwari, I. and Rao, V. 1990. Effect of
Azospirillum brasilense on growth and
nitrogen content of Cynodon dactylon
under different moisture regimens.
Indian Journal of Plant Physiology. 33:
210-213.
Bashan, Y. 1998. Inoculants of plant growth-
promoting bacteria for use in
agriculture. Biotechnology Advances.
16(4): 729-770.
https://doi.org/10.1016/S0734-
9750(98)00003-2
Bashan, Y. de-Bashan, L. and Prabhu, S. 2014.
Advances in plant growth-promoting
bacterial inoculant technology:
formulations and practical perspectives
(1998–2013). Plant Soil. 378: 123-145.
https://doi.org/10.1007/s11104-013-
1956-x
Bashan, Y., Khaosaad, T., Salazar, B., Ocampo,
J., Wiemken, A. and Oehl Horst
Vierheilig, F. 2007. Mycorrhizal
characterization of the boojum tree,
Fouquieria columnaris, an endemic
ancient tree from the Baja California
Peninsula, Mexico. Trees. 21: 329-335.
http://www.bashanfoundation.org/gma
web/pdfs/mycorrhizaecirio.pdf
Bashan, Y., Salazar, B., Moreno, M., Lopez, B.
2012. Linderman, R.G. Reforestation of
eroded
desert soil with native trees: effects of
inoculation with plant growth-
promoting microorganisms, limited
amounts of compost and water and plant
density. Journal Environmental
Management.102: 26-36.
10.1016/j.jenvman.2011.12.032
Bécard, G. and Fortin, J. 1988. Early events of
vesicular-arbuscular mycorrhiza
formation on Ri T-DNA transformed
roots. New phytologist.108: 211-218.
https://doi.org/10.1111/j.1469-
8137.1988.tb03698.x
Bécard, G. and Piché, Y. 1992. Establishment of
vesicular-arbuscular mycorrhiza in root
organ culture: review and proposed
methodology. Methods in
Microbiology. 24: 89-108.
https://doi.org/10.1016/S0580-
9517(08)70089-8
Carrillo, A., Puente, M., Castellanos, T. and
Bashan, Y. 1998. Aplicaciones
Biotecnológicas de Ecología
Microbiana. In: Manual de Laboratorio
(Eds) Pontificia Universidad Javeriana,
Santa Fe de Bogotá, Colombia and
Centro de Investigaciones Biológicas
del Noroeste, La Paz, B.C.S., México.
139 p.
Chen, C., Wang, C., Liu, Z., Liu, X., Zou, L., Shi,
J., Chen, S., Chen, J. and Tan, M. 2018.
Variations in physiology and multiple
bioactive constituents under salt stress
provide insight into the quality
evaluation of Apocyni Veneti Folium.
International Journal of Molecular
Sciences. 19: 30-42.
https://doi.org/10.3390/ijms19103042
Compant, S., Clément, C. and Sessitsch, A. 2010.
Plant growth-promoting bacteria in the
rhizo and endosphere of plants: their
role, colonization, mechanisms
involved and prospects for utilization.
Soil Biology and Biochemistry. 42:
669–678.
https://doi.org/10.1016/j.soilbio.2009.1
1.024
de Morais, C., Pinheiro, S., Martino, H. and
Pinheiro-Sant'Ana, H. 2017. Sorghum
(Sorghum bicolor L.): Nutrients,
bioactive compounds, and potential
impact on human health. Critical
Reviews in Food Science and Nutrition.

Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
12
57(2): 372-390.
10.1080/10408398.2014.887057
Declerck, S., Strullu, D. and Plenchette, C. 1998.
Monoxenic culture of the intraradical
forms of Glomus sp. isolated from a
tropical ecosystem: a proposed
methodology for germplasm collection.
Mycologia. 90: 579-585.
https://doi.org/10.1080/00275514.1998
.12026946
Díaz, V., Ferrera, C., Almaraz, S. and Alcántar,
G. 2001. Inoculation of plant growth-
promoting bacteria in lettuce. Terra. 19:
327-335.
https://www.academia.edu/34530222/I
noculation_of_Plant_Growth-
promoting_Bacteria_in_Lettuce
Dilnur, T., Peng, Z., Pan, Z., Palanga, K., Jia, Y.
and Gong Du, X. 2019. Association
analysis of salt tolerance in Asiatic
cotton (Gossypium arboretum) with
SNP markers. Critical Reviews in Food
Science and Nutrition. 20: 21-68.
https://www.mdpi.com/1422-
0067/20/9/2168
Flowers, T.J. 2004. Improving crop salt
tolerance. Journal of Experimental
Botany. 55: 307-319.
https://doi.org/10.1093/jxb/erh003
Flowers, T. and Yao, A. 1995. Breeding for
salinity resistance in crop plants-where
next. Australian Journal of Plant
Physiology. 23: 667-667.
https://link.springer.com/article/10.100
7/s11738-997-0039-0
García, H., Ruíz, A., Preciado, R., Nieto, G.,
Murillo, A., Ruiz, E., Domínguez, R.
and Rueda Puente, E. 2018. Growth and
oil yield parameters of the Capsicum
annuum var. aviculare associated to the
beneficial bacterium Bacillus
amyloliquefaciens and Azospirillum
halopraeferens under field conditions.
Biotecnia. XX(2): 59-64.
https://biotecnia.unison.mx/index.php/
biotecnia/article/view/599
Goodfriend, W., Olsen, M. and Frye, R. 2000.
Soil microfloral and microfaunal
response to Salicornia bigelovii
planting density and soil residue
amendment. Plant and Soil. 1: 23-32.
Gul, B. and Weber, D. 1999. Effect of salinity,
light, and thermoperiod on the seed
germination of AlIenrolfea occidentalis.
Canadian Journal of Botany. 77: 1-7.
Gulzar, S., Gulzar, M., Khan, M., Khan, I.,
Ungar, I. and Ungar, A. (2000). Effect
of salinity and temperature on the
germination of Urochondra setulosa
(Trin.) C. E. Hubbard. Seed Science and
Technology 29(1).
Hamdi, H. 1999. Rhizobium-Legume symbiosis
and Nitrogen fixation under severe
conditions and in arid climate.
Microbiology and Molecular Biology
Reviews. 63: 968-989.
https://www.ncbi.nlm.nih.gov/pmc/arti
cles/PMC98982/
Hariprasanna, K. and Rakshit, S. 2016.
Economic Importance of Sorghum. In:
Rakshit S., Wang YH. (eds) The
Sorghum Genome. Compendium of
Plant Genomes. Springer, Cham. DOI
https://doi.org/10.1007/978-3-319-
47789-3_1
Hegarty, T. 1978. The physiology of seed
hydration and dehydration and the
relation between water stress and the
control of germination: a review. Plant
Cell and Environment 1: 101-119.
https://doi.org/10.1111/j.1365-
3040.1978.tb00752.x
Hernández, J. 2019. Salinity Tolerance in Plants:
Trends and Perspectives. International
Journal of Molecular Sciences. 5: 10-
20. pii: E2408.
Isopi, R., Fabbri, P., Del-Gallo, M. and Puppi, G.
1995. Dual inoculation of Sorghum
bicolor (L.) Moench ssp. bicolor with
vesicular arbuscular mycorrhizas and
Acetobacter diazotrophicus. Symbiosis.
18: 43-55. http://agris.fao.org/agris-
search/search.do?recordID=US954700
7
James, E., Olivares, F., Baldani, J. and
Dobereiner, J. 1997. Herbaspirillum, an
endophytic diazotroph colonizing
vascular tissue in leaves of Sorghum
bicolor L. Moench. Journal of
Experimental Botany. 48: 785-797.
Jefferies, R. 1977. Growth responses of coastal
halophytes to inorganic nitrogen.
Journal of Ecology. 65: 847-865.
http://agris.fao.org/agris-
search/search.do?recordID=GB197802
57247
Juarez, B. and Harrison, T. 2018. Mexico, Grain
and Feed Annual, Slight Changes in
Production as Grain Imports Continue
Upward Trend Global Agricultura
Information Network. USDA.

Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
13
https://apps.fas.usda.gov/newgainapi/a
pi/report/downloadreportbyfilename?fi
lename=Grain%20and%20Feed%20An
nual_Mexico%20City_Mexico_3-7-
2018.pdf
Kapulnik, Y. Okon, J., Kigel, J., Nur, N. and Y.
Henis Y. 1981. Effects of temperature,
nitrogen fertilization and plant age on
nitrogen fixation by Setaria italica
inoculated with Azospirillum brasilense
(strain cd). Plant Physiology. 68: 340-
343.
https://www.ncbi.nlm.nih.gov/pmc/arti
cles/PMC427487/
Keiffer, C. and Ungar, L.1995. Germination
responses of halophyte seeds exposed to
prolonged hypersaline conditions. In:
Khan. M.A. and Ungar. l.A. (Eds),
Biology of Salt Tolerant Plants.
Department of Botany, University of
Karachi. Pakistan. 2-45.
Khan, M. and Weber, D. 1986. Factors
influencing seed germination in
Salicornia. American Journal of
Botany. 73: 1163-1167.
https://www.jstor.org/stable/2443795?s
eq=1
Khan, M. 1991. Studies on germination of
Cressa cretica L. seeds. Pakistan
Journal of Weed Science Research.1:
89-98.
Khan, M., Khan, I. and Khizar, T. 1976. Plant
growth regulators from species
differing in salt tolerance as affected by
soil salinity. Plant and Soil. 45: 269-
276.
Khan, M. and Ungar, L. 1998. Germination of
salt tolerant shrub Suaeda fruticosa
from Pakistan: Salinity and temperature
responses. Seed Science and
Technology. 26: 657-667.
Khan, M. and Ungar, L. 1997. Germination
responses of the subtropical annual
halophyte Zygophyllum simplex. Seed
Science and Technology. 25: 83-91.
http://agris.fao.org/agris-
search/search.do?recordID=CH970020
7
Khan, M. and Ungar L. 1996. Influence of
salinity and temperature on the
germination of Haloxylon recurvum.
Annals of Botany. 78: 547-551.
10.1006/anbo.1996.0159
Khan, M. and Ungar, L. 1998. Seed germination
and dormancy of Polygonum aviculare
L. as influenced by salinity,
temperature, and gibberellic acid. Seed
Science and Technology. 26: 107-117.
Khan, M. and Ungar, L.1999. Seed germination
and recovery of Triglochin maritima
from salt stress under different
thermoperiods. Great Basin Naturalist.
59: 144-150.
Kroes, J. and Supit, J. 2011. Impact analysis of
drought, water excess and salinity on
grass production in The Netherlands
using historical and future climate data.
Agriculture, Ecosystems and
Environment. 144(1): 370-381
https://doi.org/10.1016/j.agee.2011.09.
008
Laynez-Garsaball, J., Méndez, N. and Mayz-
Figueroa, J. 2007. Crecimiento de
plántulas a partir de tres tamaños de
semilla de dos cultivares de maíz (Zea
mays L.), sembrados en arena y regados
con tres soluciones osmóticas de
sacarosa. Idesia (Arica). 25(1): 21-36.
https://dx.doi.org/10.4067/S0718-
34292007000100003
López-Ortiz N., Tique, M., Pérez-Lavalle, L. del
S. 2011. Contribución al estudio del
sorgo (Sorghum bicolor (L.) Moench)
para nutrición humana. Perspectivas en
Nutrición Humana. 13: 33-44.
http://www.scielo.org.co/pdf/penh/v13
n1/v13n1a4.pdf
Luigi, Ch. 1998. Annamaria, B.; Tabacchioni,
S.; Dalmastri, C. Inoculation of
Burkholderia cepacia, Pseudomonas
fluorescens and Enterobacter sp. on
Sorghum bicolor: Root colonization
and plant growth promotion of dual
strain inocula. Soil Biology and
Biochemistry. 30(1): 81-87.
https://doi.org/10.1016/S0038-
0717(97)00096-5
Luo, Q. 2011. Temperature thresholds and crop
production: a review. Climatic Change.
109: 583-593.
https://doi.org/10.1007/s10584-011-
0028-6
Luo, S., Xu, T. and Chen, L. 2012. Endophyte-
assisted promotion of biomass
production and metal-uptake of energy
crop sweet Sorghum by plant-growth-
promoting endophyte Bacillus sp.
SLS18. Applied Microbiology and
Biotechnology. 93: 1745-1749.
https://doi.org/10.1007/s00253-011-
3483-0

Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
14
Maguire, J. 1962. Speed of germination-aid in
selection and evaluation for seedling
emergence and vigour. Crop Science. 2:
176-177.
https://www.scirp.org/(S(i43dyn45teex
jx455qlt3d2q))/reference/ReferencesPa
pers.aspx?ReferenceID=1017323
Maunder, A.B. 1999. History of cultivar
development in the United States: From
‘Memoirs of A. B. Maunder—Sorghum
Breeder’. In: Smith CW, Frederiksen
RA (eds) Sorghum: origin, history,
technology and production. Wiley, New
York, USA. p. 191-223
Melgares, J., González, D., and Gutiérrez A.,
Honrubia, M. and Morte, A. 2004.
Efectos del hongo endomicorrícico
Glomus intraradices en el
cultivoecológico de lechuga tipo
Iceberg In: VI Congreso de la Sociedad
Española de Agricultura Ecológica.
Almería. 300 p.
Miranda, R.S., Alvarez-Pizarro, J., Costa, J.,
Paula, S., Pirsco, J. and Gomes-Filho, E.
2017. Putative role of glutamine in the
activation of CBL/CIPK signaling
pathways during salt stress in Sorghum.
Plant Signal Behaviour. 12: 136-1075.
https://doi.org/10.1080/15592324.2017
.1361075
Nahid, S. and A. Gomah. 1991. Response of
wheat to dual inoculation with Va-
mycorrhiza and Azospirillum fertilized
with NPK and irrigated with sewage
effluent. Arid Soil Research and
Rehabilitation. 5: 83-96.
Okon, Y. Labandera, G. 1994. Agronomic
applications of Azospirillum: and
evaluation of 20 years worlwide field
inoculation. Soil Biology and
Biochemistry. 26: 1591-1601.
https://doi.org/10.1016/0038-
0717(94)90311-5
Philipupiya, J. and Ungar, L. 1984. The effect of
seed dimorphism on the germination
and survival of Salicornia europaea L.
populations. American Journal of
Botany. 71(1): 542-549.
https://doi.org/10.1002/j.1537-
2197.1984.tb12539.x
Pichu, R. 2010. Soil processes affecting crop
production in salt-affected soils.
Functional Plant Biology. 7(1): 613-
620. https://doi.org/10.1071/FP09249
Puente, M. and Bashan, Y. 1993. Effect of
inoculation with Azospirillum
brasilense strains on germination and
seedling growth of the giant columnar
cardon cactus (Pachycereus pringlei).
Symbiosis. 1: 49-60.
Puente, M., Holguin, G., Glick, B. and Bashan,
Y. 1999. Root-surface colonization of
black mangrove seedlings by
Azospirillum halopraeferens and
Azospirillum brasilense in seawater.
FEMS Microbiology Ecology. 29 (1):
283-292.
https://doi.org/10.1111/j.1574-
6941.1999.tb00619.x
Puntarulo, S., Sanchez, R. and Boveris, A. 1988.
Hydrogen peroxide metabolism in
soybean embryonic axes at the onset of
germination. Plant Physiology. 86(1):
626-630. DOI:
https://doi.org/10.1104/pp.86.2.626
Rakshit, S., Hariprasanna, K., Gomashe, S.,
Ganapathy, K., Das, I., Ramana, O.,
Dhandapani, A., Patil, J. 2014. Changes
in area, yield gains, and yield stability
of Sorghum in major Sorghum-
producing countries, 1970 to 2009.
Crop Science. 54(1): 1571-1584.
Rashid, A., Qureshi, R., Hollington, P. and Wyn,
J. 1999. Comparative responses of
wheat (Triticum aestivum L.) cultivars
to salinity at the seedling stage. Journal
of Agronomy and Crop Science. 182
(1): 199-207.
https://doi.org/10.1046/j.1439-
037x.1999.00295.x
Renganathan, P.J., Borboa-Flores, E., Rosas-
Burgos, J. Cárdenas-López, J., Ortega-
García, J. and Rueda-Puente E. 2019.
Phaseolus acutifolius en asociación con
Bacillus amyloliquefaciens y
Azospirillum halopraeferens bajo
condiciones de salinidad. Biotecnia.
1(1): 127-132.
https://biotecnia.unison.mx/index.php/
biotecnia/article/view/875
Rivers, W. and Weber W. 1971. The influence of
salinity and temperature on seed
germination in Salicornia bigelovii.
Physiologia Plantarum. 24(1): 73-75.
https://doi.org/10.1111/j.1399-
3054.1971.tb06719.x
Rozema, J. 1975. The influence of salinity,
inundation and temperature on the
germination of some halophytes and
non-halophytes. Planta Acta OEcol.
10(1): 341-345.
https://doi.org/10.1111/j.1399-
3054.1971.tb06719.x

Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
15
Rueda-Puente, E., Castellanos, T., Troyo, E., and
De León, J. 2004. Effect of Klebsiella
pneumoniae and Azospirillum
halopraeferens on the growth and
development of two Salicornia
bigelovii genotypes. Australian journal
of experimental agriculture. 44 (1): 65-
74. https://doi.org/10.1071/EA03012
Rueda-Puente, E., Bianciotto, O., Saifolah, F.,
Omid, Z., López, E., Hernández-
Montiel, G., Murillo, A. and Sediles, G.
2018. Plant Growth-Promoting Bacteria
Associated to the Halophyte Suaeda
maritima (L.) in Abbas, Iran. ©
Springer Nature Switzerland AG. B.
Gul et al. (eds.), Sabkha Ecosystems,
Tasks for Vegetation Science VI,
https://doi.org/10.1007/978-3-030-
04417-6_18
Ruzzi, M. and Aroca, R. 20015. Plant growth-
promoting rhizobacteria act as
biostimulants in horticulture. Scientia
Horticulturae. 196(1): 124-134.
http://www.bashanfoundation.org/contr
ibutions/Aroca-R/2015.-Aroca-SH.pdf
Salim, M. 1989. Effects of NaCl and KCl salinity
on growth and ionic relations of red
kidney beans (Phaseolus vulgaris L.).
Journal Agronomy and Crop
Science.163 (1): 338-344.
https://doi.org/10.1111/j.1439-
037X.1989.tb00776.x
Sangakkara. U., Meylemans, B. and Van
Damme, P. 1995. Impact of Different
Weed Types on Growth and Yield of
Mungbean (Vigna radiata L. Wilczek).
Journal Agronomy and Crop Science.
175: 1-5. http://agris.fao.org/agris-
search/search.do?recordID=DE962037
7
Sarig, S., Okon, Y. and Blum, A. 1992. Effect of
Azospirillum brasilense inoculation on
growth dynamics and hydraulic
conductivity of Sorghum bicolor roots.
Journal of Plant Nutrition. 15(1): 805-
819.
SAS. 2004. SAS Institute. SAS/STAT user’s
guide. Version 6.12 SAS, Institute,
Cary, N.C. U.S.A.
Secretaria de Agricultura, Ganadería y
Desarrollo Rural SAGAR. 1981.
Alternativas alimentarias en la cuenca
del Golfo de California. In: VI simposio
sobre el medio ambiente del Golfo de
California. Halffer G. (Ed) Secretaria de
Agricultura y Recursos Hidráulicos.
Hermosillo, Sonora, México. 334 p.
Shalaby, E., Epstein, E. and Qualset, C. 1993.
Variation in Salt Tolerance among
Some Wheat and Triticale Genotypes.
Journal Agronomy and Crop
Science.171: 298-304. 10.1111/j.1439-
037X.1993.tb00144.x
Sokal, R. and James, R. 1988. Biometry. In: The
principles and practice of statistics in
biological research. 3rd edn. Freeman
and Co, San Francisco. 435 p.
https://www.amazon.com.mx/Biometry
-Principles-Practice-Statistics-
Biological/dp/0716724111
Stein, T., Hayen, N. and Fendrik, S. 1997.
Contribution of BNF by Azoarcus sp.
BH72 in Sorghum vulgare. Soil Biology
and Biochemistry. 29: 969-971.
http://agris.fao.org/agris-
search/search.do?recordID=US201302
874180
Strullu, D. and Romand, C. 1986. Méthode
d’obtention d’endomycorhizes à
vésicules et arbuscules en conditions
axéniques. Comptes Rendus de l
Académie des Sciences - Series III. 530
p.
Ungar, L. 1995. Seed germination and seed-bank
ecology of halophytes. In Seed
Development and Germination, (eds. J,
Kigel and G. Galili) Marcel and Dekker
Incorporation. New York. 221 p.
Vessey, J.K. 2003. Plant growth promoting
rhizobacteria as biofertilizers. Plant and
Soil. 255: 571-583.
https://doi.org/10.1023/A:1026037216
893
Whipps, J. 2000. Microbial interactions and
biocontrol in the rhizosphere. Journal of
Experimental Botany.15(1): 487-511.
https://doi.org/10.1093/jexbot/52.suppl
_1.487
Woodell, S. 1985. Salinity and seed germination
patterns in coastal plants. Vegetation.
61(1): 223-229.
https://www.jstor.org/stable/20146251
?seq=1
Yan, K., Bian, T., He, W., Han, G., Lv, M., Guo
M. and Lu, M. 2018. Root abscisic acid
contributes to defending photoinibition
in jerusalem artichoke (Helianthus
tuberosus L.) under salt stress.
International Journal of Molecular
Sciences. 19(1): 39-34.
https://doi.org/10.3390/ijms19123934
Zhang, Y., Liu, L., Chen, B., Qin, Z., Xiao, Y.,
Zhang, Y., Yao, R., Liu, H. and Yang,

Tropical and Subtropical Agroecosystems 23 (2020): #48 Holguin-Peña et al., 2020
16
H. 2019. Progress in understanding the
physiological and molecular responses
of Populus to salt stress. International
Journal of Molecular Sciences. 20(1):
1312-1342. 10.3390/ijms20061312
Zhong. M., Wang, Y., Zhang, Y., Shu, S., Sun, J.
and Guo O.S. 2019. Overexpression of
transglutaminase from cucumber in
tobacco increases salt tolerance through
regulation of photosynthesis.
International Journal of Molecular
Sciences. 20(1): 894-910.
10.3390/ijms20040894