plac8-dependent and inducible no synthase … · plac8-dependent and inducible no...
TRANSCRIPT

of August 30, 2018.This information is current as
Genital Tract Infections from theChlamydia muridarum
Synthase-Dependent Mechanisms Clear -Dependent and Inducible NOPlac8
Raymond M. Johnson, Micah S. Kerr and James E. Slaven
http://www.jimmunol.org/content/188/4/1896doi: 10.4049/jimmunol.1102764January 2012;
2012; 188:1896-1904; Prepublished online 11J Immunol
MaterialSupplementary
4.DC1http://www.jimmunol.org/content/suppl/2012/01/11/jimmunol.110276
Referenceshttp://www.jimmunol.org/content/188/4/1896.full#ref-list-1
, 29 of which you can access for free at: cites 42 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2012 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Plac8-Dependent and Inducible NO Synthase-DependentMechanisms Clear Chlamydia muridarum Infections from theGenital Tract
Raymond M. Johnson,* Micah S. Kerr,* and James E. Slaven†
Chlamydia trachomatis urogenital serovars replicate predominantly in genital tract epithelium. This tissue tropism poses a unique
challenge for host defense and vaccine development. Studies utilizing the Chlamydia muridarum mouse model have shown that
CD4 T cells are critical for clearing genital tract infections. In vitro studies have shown that CD4 T cells terminate infection by
upregulating epithelial inducible NO synthase (iNOS) transcription and NO production. However, this mechanism is not critical,
as iNOS-deficient mice clear infections normally. We recently showed that a subset of Chlamydia-specific CD4 T cell clones could
terminate replication in epithelial cells using an iNOS-independent mechanism requiring T cell degranulation. We advance that
work using microarrays to compare iNOS-dependent and iNOS-independent CD4 T cell clones. Plac8 was differentially expressed
by clones having the iNOS-independent mechanism. Plac8-deficient mice had delayed clearance of infection, and Plac8-deficient
mice treated with the iNOS inhibitor N-monomethyl-L-arginine were largely unable to resolve genital tract infections over 8 wk.
These results demonstrate that there are two independent and redundant T cell mechanisms for clearing C. muridarum genital
tract infections: one dependent on iNOS, and the other dependent on Plac8. Although T cell subsets are routinely defined by
cytokine profiles, there may be important subdivisions by effector function, in this case CD4Plac8. The Journal of Immunology,
2012, 188: 1896–1904.
Chlamydia trachomatis serovars D–K are sexually trans-mitted bacterial pathogens that cause urethritis, endo-metritis, salpingitis, epididymitis, and pelvic inflam-
matory disease. In women, infections ascend into the upperreproductive tract causing scarring responsible for ectopic preg-nancies and infertility. Aggressive public health measures basedon antibiotic treatment of source cases and their sexual partnersare partially counterproductive due to deleterious effects on herdimmunity (1–3). There is a consensus among researchers andpublic health experts that a Chlamydia vaccine is likely necessaryto reduce the incidence of C. trachomatis infections and themedical resources committed to treating them. A critical compo-nent of rational vaccine development is defining the immuneparameters that mediate/correlate with protective immunity.For viral vaccines, neutralizing Abs are the immune parameter
that correlates with protective immunity against primary infec-
tions. Existing data from the Chlamydia muridarum mouse modelsuggest that immunity to urogenital serovars of Chlamydia iscomplex. Passive transfer of Abs from immune mice has littleeffect on primary infections of the genital tract (4), andB cell-deficient mice clear primary genital tract infections similar towild-type controls (5). Clearance of primary genital tract infectionsis dependent on T cell immunity, and T cell immunity is sufficientto clear the genital tract (6). MHC class I Ag presentation is dis-pensable for protective host immunity whereas MHC class II isabsolutely required (7). Recent data in experimental mouse modelsargue for a supportive role for Abs in vaccine-generated immunityin the genital tract (8, 9). A large body of research has identifiedCD4 T cells of the Th1 subset as the critical parameter for pro-tective host immunity (6). The protective Th1 subset generated byvaccination appears to be multifunctional CD4 T cells that secreteIFN-g and TNF-a, with or without IL-2 production (10).In vitro studies using a Chlamydia-specific CD4 T cell clone ca-
pable of terminating C. muridarum replication in epithelial cellsidentified a probable mechanism for sterilizing CD4 T cell-mediatedimmunity that was dependent on inducible NO synthase (iNOS),IFN-g, and T cell/epithelial cell contact via LFA-1/ICAM-1 (11–14).However, follow-up studies showed that iNOS knockout mice werenot compromised in clearance of C. muridarum genital tract infec-tions (15, 16), and that IFN-g knockout mice cleared 99.9% of C.muridarum from the genital tract with near normal kinetics (17, 18).Furthermore, mice deficient in the known T cell killing mechanismsperforin and Fas ligand/Fas were also able to clear C. muridarumgenital tract infections with normal kinetics (19). Additionalknockout mice and Ab depletion studies have shown that mice de-ficient in TNF-a receptors (20), IL-12 (18, 20), or IL-1b (21) havevarying degrees of compromised C. muridarum genital tract infec-tion clearance. However, cytokine deficiencies have pleiotropiceffects on host innate and adaptive immunity and do not identify spe-cific effector mechanisms responsible for terminating Chlamydia re-plication in reproductive tract epithelium.
*Department of Medicine, Indiana University School of Medicine, Indianapolis,IN 46202; and †Department of Biostatistics, Indiana University School of Medicine,Indianapolis, IN 46202
Received for publication September 27, 2011. Accepted for publication December 5,2011.
This work was supported by National Institutes of Health Grant R01AI070514(to R.M.J.).
The microarray data presented in this article have been submitted to the Gene Ex-pression Omnibus database (www.ncbi.nlm.nih.gov/geo) under accession numberGSE32128.
Address correspondence and reprint requests to Dr. Raymond M. Johnson, Divisionof Infectious Diseases, Department of Medicine, Indiana University School ofMedicine, 635 Barnhill Drive, MS 224, Indianapolis, IN 46202. E-mail address:[email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: EB, elementary body; IFU, inclusion-forming unit;iNOS, inducible NO synthase; MLA, N-monomethyl-L-arginine; PAO, phenyl arsenicoxide; RMA, robust multiarray analysis; SPG, sucrose-phosphate-glutamic acid.
Copyright� 2012 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/12/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1102764
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Our laboratory recently identified an iNOS-independent mech-anism for terminating C. muridarum replication in epithelial cells(22). Using a panel of Chlamydia-specific CD4 T cell clones weshowed that a subset of clones could terminate C. muridarumreplication in the presence of the iNOS inhibitor N-monomethyl-L-arginine (MLA), whereas a separate clone was completelyinhibited by MLA as previously reported in the literature. TheiNOS-independent CD4 T cell clones were also relatively inde-pendent of IFN-g and could be partially blocked using phenyl ar-senic oxide (PAO), an inhibitor of T cell degranulation. Potentblocking of the iNOS-independent CD4 clones required both aninhibitor of degranulation and an iNOS inhibitor, suggesting twoindependent mechanisms capable of terminating C. muridarumreplication in infected epithelial cells. All of the Chlamydia-specificCD4 T cell clones were Th1, and they made IFN-g when activatedand were capable of recognizing infected epithelial cells (23).To further investigate these findings, we compared two iNOS-
independent CD4 T cell clones to two iNOS-dependent CD4T cell clones using gene expressionmicroarrays.Microarray analysisidentified a candidate second mechanism dependent on Plac8 forterminating C. muridarum replication in epithelial cells that wasthen investigated in vivo utilizing an existing Plac8 knockoutmouse. Plac8 knockout mice were significantly compromised intheir ability to clear C. muridarum genital tract infections.
Materials and MethodsMice
Four- to five-week-old female C57BL/6J and B6.129S6-Plac8tm1Bhk/J(stock no. 009598) mice were purchased from The Jackson Laboratory(Bar Harbor, ME). All mice were housed in Indiana University–PurdueUniversity Indianapolis specific pathogen-free facilities. The Indiana Uni-versity–Purdue University Indianapolis Institutional Animal Care and Uti-lization Committee approved all experimental protocols.
Cells, T cell clones, and bacteria
C57epi.1 epithelial cells and Chlamydia-specific CD4 T cell clones uvmo-1, uvmo-2, uvmo-4, and spl4-10 were derived from C57BL/6 mice andcultured as previously described (23). Mycoplasma-free C. muridarum(Nigg), previously known as C. trachomatis strain mouse pneumonitis, wasgrown in McCoy cells as previously described (23).
Cytokine ELISAs
T cell clones (5 3 104) were activated in 96-well tissue culture plates byimmobilized anti-CD3 mAb 145-2c11 (BD Biosciences, San Jose, CA),0.5 mg/ml in PBS overnight at 4◦C (washed once), in RPMI 1640 mediacontaining 1 hg/ml recombinant murine IL-7 (R&D Systems, Minneapolis,MN). Relative levels of IFN-g, TNF-a, IL-2, and IL-10 in culture super-natants were determined by ELISA using capture and biotinylated mAbpairs with recombinant murine standards according to the manufacturers’protocols (IFN-g ELISA, XMG1.2; IL-2 ELISA, 5H4/1A12 [Pierce-Endogen; Rockford, IL]; TNF-a ELISA, TN3-19.12/C1150-14; IL-10ELISA, JES5-2A5/SXC-1 [BD Biosciences]). Detection was accomplishedwith streptavidin-HRP (BD Biosciences) and tetramethylbenzidine sub-strate (Sigma-Aldrich, St. Louis, MO).
Gene expression microarray analysis
Chlamydia-specific CD4 T cell clones uvmo-2, uvmo-3, uvmo-4, and spl4-10 were purified by Ficoll-Hypaque (Histopaque 1083; Sigma-Aldrich) atthe end of their culture cycle and then grown for 3 d in their usual culture
media including growth factors, without Ag stimulation. On day 3, totalRNAwas isolated from each T cell clone using a protocol that included anRNase-free DNase I treatment step (RNeasy; Qiagen, Valencia, CA). Withassistance from the Indiana University Center for Medical Genomics, geneexpression patterns were analyzed using the Affymetrix Mouse ST 1.0Array that analyzes 28,853 murine genes. Samples were labeled using thestandard Affymetrix protocol for the Whole Transcript Sense Target La-beling and Control Reagents kit according to the Affymetrix user manual,that is, the GeneChip Whole Transcript Sense Target Labeling AssayManual. Individual labeled samples were hybridized to the Mouse Gene1.0 ST GeneChips for 17 h and then washed, stained and scanned with thestandard protocol using Affymetrix GeneChip Operating System. TheGeneChip Operating System was used to generate data (CEL files). Arrayswere visually scanned for abnormalities or defects. CEL files wereimported into Partek Genomics Suite (Partek, St. Louis, MO). Robustmultiarray analysis (RMA) signals were generated for the core probe setsusing the RMA background correction, quantile normalization, and sum-marization by median polish. Summarized signals for each probe set werelog2 transformed. These log-transformed signals were used for principalcomponents analysis, hierarchical clustering, and signal histograms todetermine whether there were any outlier arrays. Untransformed RMAsignals were used for fold change calculations. Data were analyzed usinga one-way ANOVA using log2-transformed signals for all four CD4 T cellclones, and contrasts were made comparing iNOS-dependent uvmo-4 andspl14-10 individually to the combined expression of the iNOS-independentT cell clones uvmo-2 and uvmo-3. Fold changes were calculated using theuntransformed RMA signals. Genes up- or downregulated 5-fold with pvalues of ,0.001 for either uvmo-4 or spl4-10 compared with uvmo-2/uvmo-3 were considered in the final analysis (Supplemental Table I).
RT-PCR analysis
Chlamydia-specific CD4 T cell clones uvmo-2, uvmo-3, uvmo-4, and spl4-10 were purified by Ficoll-Hypaque at the end of their culture cycle andthen grown for 3 d in their usual culture media including growth factors,without Ag stimulation. On day 3, total RNAwas isolated from each T cellclone using a protocol that included an RNase-free DNase I treatmentstep. Specific mRNA gene reverse transcription and amplification wereperformed using AMV reverse transcriptase/Tfl DNA polymerase ina one-step system (AccessQuick RT-PCR; Promega, Madison, WI).Amplification conditions were 1) 48˚C for 45 min; 2) 95˚C for 2 min; 3)95˚C for 30 s; 4) 56˚C for 20 s; 5) 72˚C for 30 s; 6) go to step 3 for37 times; 7) 72˚C for 7 min; and 8) hold at 4˚C using an MJ Research J200PCR machine. All gene analyses were done on the same set of RNAsamples. PCR primers, amount of total RNA in amplification, and expectedproduct sizes are shown in Table I. RT-PCR reactions were run on 2%agarose gels with ethidium bromide; images are inverted for presentation.The “no RT” controls generated no RT-PCR products (data not shown).PCR primers used are shown in Table I.
Epithelial cell infections, Chlamydia replication, and titration
C57epi.1 cell monolayers in 48-well plates were untreated or treated withIFN-g (10 hg/ml) for 10 h prior to infection. Wells were infected with 3inclusion-forming units (IFU) per cell. After addition of C. muridarum theplates were spun at 300 3 g for 30 min. Four hours postinfection, theinoculums were removed and CD4 T cell clones were added in RPMI1640 T cell medium. Thirty-two hours postinfection, wells were scraped,harvested with an equal volume of sucrose-phosphate-glutamic acid (SPG)buffer, and stored at 280˚C until C. muridarum titers were determinedon McCoy cell monolayers using an anti-Chlamydia LPS Ab andFITC-labeled goat anti-mouse IgG (Rockland Immunochemicals, Gil-bertsville, PA) as previously described (24).
Genital tract infections
One week prior to infection mice were treated with 2.5 mg medroxy-progesterone delivered s.c. (Depo-Provera; Pfizer Pharmaceuticals, New
Table I. RT-PCR primers
Gene Sense Primer (39 → 59) Antisense Primer (39 → 59)ProductSize (bp)
Total RNA inAmplification (hg)
Casd1 GGGAGATCAACCACTACTTCAG GACCCAATAAACATCACTAGTC 590 250Plac8 ATGGCTCAGGCACCAACAGTTA GAAAGCGTTCATGGCTCTCCTC 336 250Prfl TGGATGTGAACCCTAGGCCAGAG AAGTACTTCGACGTGACGCTCAC 515 100b-actin ATGGATGACGATATCGCTGCGC CGTACATGGCTGGGGTGTTGAA 400 100
The Journal of Immunology 1897
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

York, NY). Lightly anesthetized mice were infected vaginally with 53 104
IFU C. muridarum biovar Nigg in 10 ml SPG buffer. Mice were swabbedMonday, Wednesday, and Friday to monitor the clearance of C. muridarumfrom the genital tract as previously described (25).
Pathology scoring
A simple scoring system was used to assess macroscopic genital tractpathology. Hydrosalpinx was scored as 0, 1, or 2 reflecting no, unilateral, orbilateral hydrosalpinx, respectively. Mice have a bifed uterus (two uterinehorns) that was similarly scored 0, 1, or 2 for hydro-uterus. The maximumpathology score for an individual mouse was 4.
Degranulation experiments
CD4 T cell clone cells (5 3 104) were incubated in wells with media,10 mg/ml PMA (Sigma-Aldrich), and 500 ng/ml calcium ionophore A23187(Sigma-Aldrich), PMA and A23187 plus immobilized anti-CD3 (0.5 mg/ml;50 ml PBS overnight at 4˚C) (145-2C11; BD Biosciences), or media con-taining 0.1% Triton X-100 for 4 h at 37˚C. Culture supernatants were col-lected and levels of granzyme B were determined using a murine granzymeB ELISA (mouse granzyme B DuoSet; R&D Systems). To determine per-centage degranulation, the following equation was used: % degranulation ={[measured value in experimental well 2 spontaneous release (media)]/[Triton X-100 value (maximal release) 2 spontaneous release]} 3 100.
Statistical analyses
Summary figures for each experimental investigation are presented asmeans and SD or as “pooled” means and SEM. Figure legends indicate thenumber of independent experiments pooled to generate each figure. AStudent two-tailed t test was used to assess significance of experimentaldata. Homogeneity of variances was assessed using a folded F test. Sta-tistical analysis of Chlamydia shedding was performed using repeatedmeasures ANOVA to test for differences between groups over time. Alldata were verified to meet analytic assumptions. Nonparametric tests werealso performed and showed similar results. Analyses were performed usingSAS 9.2 (SAS Institute, Cary, NC).
ResultsCharacteristics of Chlamydia-specific CD4 T cell clones usedin gene expression microarray analyses
We previously characterized a panel of 10 Chlamydia-specific CD4T cell clones, including uvmo-2, uvmo-3, uvmo-4, and spl4-10 (22,23). The uvmo-2 and uvmo-3 T cell clones are potent terminatorsof C. muridarum replication, with or without IFN-g pretreatment ofepithelial monolayers, and they make significant amounts of IFN-gwhen activated by infected epithelial cells (23). They have twopotent mechanisms to terminate replication: one dependent oniNOS, and the other dependent on T cell degranulation. Significantreversal of the ability of uvmo-2 and uvmo-3 to terminate infectionrequired simultaneous use of two inhibitors: an iNOS-inhibitor(MLA) and an inhibitor of T cell degranulation (PAO) (22).Availability of the degranulation-dependent mechanism alloweduvmo-2 and uvmo-3 to terminate C. muridarum replication in thepresence of iNOS inhibitors (iNOS-independent). The uvmo-4 and
spl4-10 CD4 T cell clones make modest amounts of IFN-g whenactivated by infected epithelial cells, and they require IFN-gpretreatment of the epithelial monolayer to control C. muridarumreplication (23). The ability of spl4-10 to terminate C. muridarumreplication in epithelial cells is iNOS-dependent and was com-pletely reversed by iNOS inhibition with MLA (22). The mech-anism used by uvmo-4 to terminate C. muridarum replication inepithelial cells was not previously determined. Because uvmo-4required IFN-g pretreatment of epithelial cells to control C.muridarum replication, we hypothesized that it was dependent oniNOS, similar to spl4-10. C57epi.1 epithelial monolayers pre-treated 10 h with 10 hg/ml IFN-g in the absence or presence of 1mM MLA were infected with C. muridarum for 4 h, then cocul-tured without and with uvmo-4 at a T cell/epithelial cell ratio of∼0.75:1. Thirty-two hours after infection the wells were harvestedby addition of SPG buffer plus scraping. Replication of C. mur-idarum under each condition was quantified by plating recoveredIFU on McCoy monolayers (Fig. 1). The ability of uvmo-4 toblock .90% of C. muridarum recovery required IFN-g pretreat-ment of the epithelial monolayer. iNOS inhibition with MLA re-stored C. muridarum recovery from untreated epithelial cells andrestored C. muridarum recovery to .50% of control values inIFN-g–pretreated epithelial cells. Termination of C. muridarumreplication by uvmo-4 was dependent on iNOS production ofNO. Identification of uvmo-4 as an iNOS-dependent CD4 T cell
FIGURE 1. Uvmo-4 is an iNOS-dependent
CD4 T cell clone. C57epi.1 epithelial cells un-
treated (A) or pretreated for 10 h with IFN-g (B),
in the absence or presence of iNOS inhibitor
MLA, were infected with C. muridarum at 3
IFU/cell. Inocula were removed 4 h later and
infected epithelial cells were cocultured without
and with 1.53 105 uvmo-4T cells in the absence
of MLA (Media) or in the presence of MLA.
Thirty-two hours after infection wells were har-
vested and C. muridarum was quantified on
McCoymonolayers. Means and SD are from one
experiment done in triplicate; statistical com-
parisons were made to the no T cell control wells
set at 100%. *p , 0.05, **p , 0.005.
FIGURE 2. Multifunctional Chlamydia-specific CD4 T cell clones with
differing cytokine profiles. T cells (5 3 104) were cultured, without acti-
vation or activated by immobilized anti-CD3 mAb, in RPMI 1640 media
containing 1 hg/ml IL-7. Twenty hours later supernatants were collected
and analyzed for individual cytokines by ELISA. Cytokine production in
media alone was negligible for all clones (graphed but not easily visualized
to left of each colored or hatched bar). Aggregate means and SEM are from
two independent experiments; statistical comparisons were made to the
media control. Note that scale is linear to 30,000 and is logarithmic there-
after. *p , 0.05, **p , 0.005, ***p , 0.0005.
1898 Plac8 AND iNOS CLEAR CHLAMYDIA INFECTIONS
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

clone gave us the desired CD4 T cell panel for gene expressionmicroarray analyses, that is, two iNOS-dependent CD4 T cellclones (uvmo-4 and spl4-10) and two iNOS-independent CD4T cells clones (uvmo-2 and uvmo-3).Uvmo-2, -3, and -4 and spl4-10 would be classified as Th1 based
on IFN-g production, but there are significant differences in theircytokine profiles (Fig. 2). When activated via the TCR complex,uvmo-2, uvmo-3, and uvmo-4 produce TNF-a (50–100 hg/ml)and IL-2 (3–10 hg/ml) in addition to IFN-g; spl4-10 producessignificant amounts of IL-10 (14 hg/ml), lesser amounts of IFN-gand TNF-a, and a negligible amount of IL-2.To assess the role of cytokines in the anti-Chlamydia effector
mechanism of these four CD4 T cell clones, we activated eachT cell clone with UV-inactivated C. muridarum-pulsed irradiatedsplenocytes in media containing IL-2 (25 U/ml) and IL-7 (2 hg/ml) and harvested the conditioned media 48 h later. Control RPMI
1640 media and fresh cell-free–conditioned media from eachclone (40% v/v in RPMI 1640 media) were exchanged for theepithelial media covering monolayers of C57epi.1 epithelial cellsinfected 4 h earlier with 3 IFU C. muridarum per cell. Thirty-twohours after infection the wells were harvested by adding SPGbuffer and scraping. Replication of C. muridarum under eachcondition was quantified using McCoy monolayers (Fig. 3). Thefinal IFN-g concentration under each experimental condition wascomparable for all four clones, as was the relative inhibition of C.muridarum replication. Fresh T cell-conditioned media was 2–3logs more potent than recombinant murine IFN-g at 10 hg/ml,which causes only a 2- to 3-fold reduction in recovered IFU perwell in C57epi.1 cells (22). The cytokine-mediated inhibition ofreplication was almost completely reversed in the presence ofMLA, arguing that the mechanism of replication inhibition bycytokines in the absence of T cells was potent induction of epi-thelial iNOS and NO production.
Gene expression microarrays comparing iNOS-independentT cell clones to iNOS-dependent T cell clones
Our previous study showed that the most potent CD4 T cell cloneterminators of C. muridarum replication, uvmo-2 and uvmo-3,used induction of epithelial iNOS and a degranulation-depen-dent effector mechanism that was not functional in spl4-10. Wealso showed that uvmo-2 and uvmo-3 recognized and lysed in-fected epithelial cells late in replication cycle, which was too lateto catch C. muridarum in the noninfectious reticulate body stage(eclipse phase). Late recognition of infected epithelial cells byuvmo-2 and uvmo-3 argued against their termination mechanismbeing a function of earlier presentation of their nominal Ags(i.e., not early recognition). Because the uvmo-2 and uvmo-3could terminate replication in the presence of iNOS inhibitorMLA, we hypothesized that they had an unidentified T cell ef-fector mechanism. To gain insight into this unidentified degran-ulation-dependent effector mechanism, we performed gene ex-pression microarrays comparing iNOS-independent (uvmo-2 anduvmo-3) to iNOS-dependent (uvmo-4 and spl4-10) T cell clones.We recognized that microarray analyses of four T cell cloneswould not permit statistically significant conclusions about T cellmechanisms, but we postulated that the resulting data wouldsuggest testable hypotheses. T cell clones at the end of theirculture cycle were purified by Ficoll-Hypaque to remove cellulardebris, washed, and then cultured for 72 h in media with usual
FIGURE 3. The role of T cell cytokines and iNOS in controlling C.
muridarum replication in epithelial cells. Fresh T cell clone-conditioned
culture supernatant (40% v/v), without and with 1 mM MLA, was ex-
changed for the infecting inocula 4 h postinfection of C57epi.1 epithelial
monolayers with C. muridarum (3 IFU/cell). Thirty-two hours later the
wells were harvested and C. muridarum was quantified on McCoy mon-
olayers. Concentration of IFN-g in 40% conditioned media for each clone
is shown in brackets. Percentage inhibition of C. muridarum replication is
shown immediately above each bar. Black bars indicate no treatment; gray
bars indicate 1 mM MLA. Mean and standard deviations are from one
experiment done in triplicate; statistical comparisons were made between
untreated and MLA-treated wells for each clone. *p , 0.05, **p , 0.005,
***p , 0.0005.
FIGURE 4. Principal components analysis of
the gene expression microarray data for the four
CD4 T cell clones: uvmo-2, red; uvmo-3, blue;
uvmo-1, light purple; spl4-10, dark purple.
The Journal of Immunology 1899
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

growth cytokines. Total RNA was harvested and used for Affy-metrix mouse ST1.0 gene expression microarrays. Four indepen-dent RNA isolations for four independent microarray experimentswere done to minimize background noise. Fig. 4 shows principalcomponents analysis from the microarrays comparing the fourT cell clones. Interestingly, the two iNOS-independent CD4 T cellclones derived from different mice (uvmo-2 and uvmo-3) clus-tered tightly together and away from the iNOS-dependent clones(uvmo-4 and spl4-10), indicating that they had a similar expres-sion pattern for the genes that accounted for the greatest variabilityin the data set. In Supplemental Table I we list the 193 genes thatdiffered 65-fold comparing the individual expression patterns ofspl4-10 and uvmo-4 to the combined expression pattern of uvmo-2and uvmo-3. The genes that were the most intriguing to us and theprimary focus of this study were those potentially useful as bio-markers or possibly involved in an effector mechanism (Table II).Uvmo-2 and uvmo-3, the iNOS-independent terminators of C.muridarum replication, had high mRNA levels of Casd1 (CAS1domain containing 1) and Plac8 (placenta-specific 8), and lowlevels of perforin (Prf1). The iNOS-dependent T cell clonesuvmo-4 and spl4-10 had very low Casd1 and Plac8 levels andhigh levels of perforin. RT-PCR analysis confirmed the microarraydata for Casd1, Plac8, and perforin in iNOS-independent (uvmo-2and uvmo-3) and iNOS-dependent (uvmo-4 and spl4-10) CD4T cell clones (Fig. 5).
Plac8 knockout mice are compromised in their ability to clearC. muridarum genital tract infections
High-level expression of Plac8 by the iNOS-independent CD4T cell clones was particularly interesting because Plac8 knockoutmice were previously shown to have a defect in clearing Klebsiellapneumoniae peritoneal infections, a phenotype ascribed to a kill-ing defect in Plac8-deficient neutrophils (26). In the original
Plac8 knockout mouse study, investigation of tissue distributionshowed expression of Plac8 in bulk T cell populations by Westernblotting. To investigate whether Plac8 was important in host de-fense against C. muridarum genital tract infections, we challengedPlac8 knockout mice, comparing them to C57BL/6J control mice.C57BL/6J and Plac8 knockout mice were treated with medro-
xyprogesterone to synchronize estrous, then infected vaginallywith 5 3 104 IFU C. muridarum. The iNOS inhibitor MLA wasadded to the drinking water on day 52 and continued thereafter.Mice were swabbed thrice weekly through 80 d postinfection andthen assessed for genital tract pathology. Recovered IFU fromgenital swabs were titered on McCoy monolayers (Fig. 6). Plac8mice had a clear defect in clearance of C. muridarum from thegenital tract. C57BL/6J control mice cleared the infection by day26 whereas Plac8 knockout mice cleared at day 47. Clearancekinetics were identical through the first 3 wk infection beforediverging during clearance of the final ∼1000 IFU/swab from thegenital tract. Addition of iNOS inhibitor MLA to the drinkingwater postclearance on day 52 led to transient recovery of viableC. muridarum from Plac8 knockout mice on days 56 and 59, butnot from C57BL/6J mice at any time point. With MLA treatmentall the Plac8 knockout mice shed viable Chlamydia on at least 1 d,two mice on a single day (either day 56 or day 59), and four miceon both days. These results mirror previously published experi-ments done by Ramsey et al. (27) with iNOS knockout mice wherecyclophosphamide treatment (lymphocyte/neutrophil depletion)postclearance led to transient recovery of viable C. muridarumfrom iNOS knockout mice but not C57BL/6J control mice. Ourdata combined with those of Ramsey et al. argue that sterilizingimmunity in the genital tract is dependent on two separate mech-anisms: one mechanism dependent on iNOS, the other mechanismdependent on Plac8. Clearance of transient C. muridarum sheddingin iNOS and Plac8 knockout mice in postclearance experimentswith cyclophosphamide and MLA, respectively, argues for a thirdmechanism, possibly the Ab-dependent mechanism described byMorrison et al. (4, 28).To further investigate the role of Plac8 in clearance of geni-
tal tract infections, C57BL/6J and Plac8 knockout mice weretreated with medroxyprogesterone 1 wk prior to infection, continu-ous MLA was added to the drinking water 1 d prior to infection,and then the mice were infected vaginally with 5 3 104 IFU C.muridarum. Mice were swabbed thrice weekly through day 56postinfection; recovered IFU were titered on McCoy monolayers(Fig. 7). Plac8 knockout mice treated with MLA were markedlycompromised in their ability to clear C. muridarum compared withC57BL/6J mice under the same experimental conditions. Clear-ance kinetics diverged after day 14, with all C57BL/6J miceclearing the infection by day 40 compared with only one of eightPlac8 knockout mice clearing the infection by the day 56. Inaddition to the absolute difference in final clearance, there isa difference in the slope of the clearance curve, with the Plac8clearance curve being nearly flat from day 21 through the endof the experiment. Clearance of C. muridarum from one Plac8knockout mouse on the last day of the experiment raises thepossibility that Plac8 knockout mice treated with MLA would
Table II. Biomarker and mechanism genes
Gene SymbolMaximum
Average Signalp Value of uvmo-2/3 Versus uvmo-4
Fold Changeuvmo-2/3 Versus
uvmo-4p Value uvmo-2/3Versus spl4-10
Fold Changeuvmo-2/3 Versus
spl4-10
Casd1 9.15 1.58 3 10216 31.68 1.11 3 10216 35.15Plac8 11.30 2.33 3 10216 138.39 1.40 3 10216 171.60Prf1 11.17 1.30 3 10211 222.62 1.80 3 10212 239.88
FIGURE 5. RT-PCR confirmation of microarray data. mRNA levels for
Casd1, Plac8, perforin (Prf1), and b-actin were analyzed using an inde-
pendent total RNA isolation for each CD4 T cell clone. RT-PCR products
were run on a 2% ethidium bromide agarose gel; molecular mass markers
are on the left margin. Image shown is inverted for presentation purposes.
1900 Plac8 AND iNOS CLEAR CHLAMYDIA INFECTIONS
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

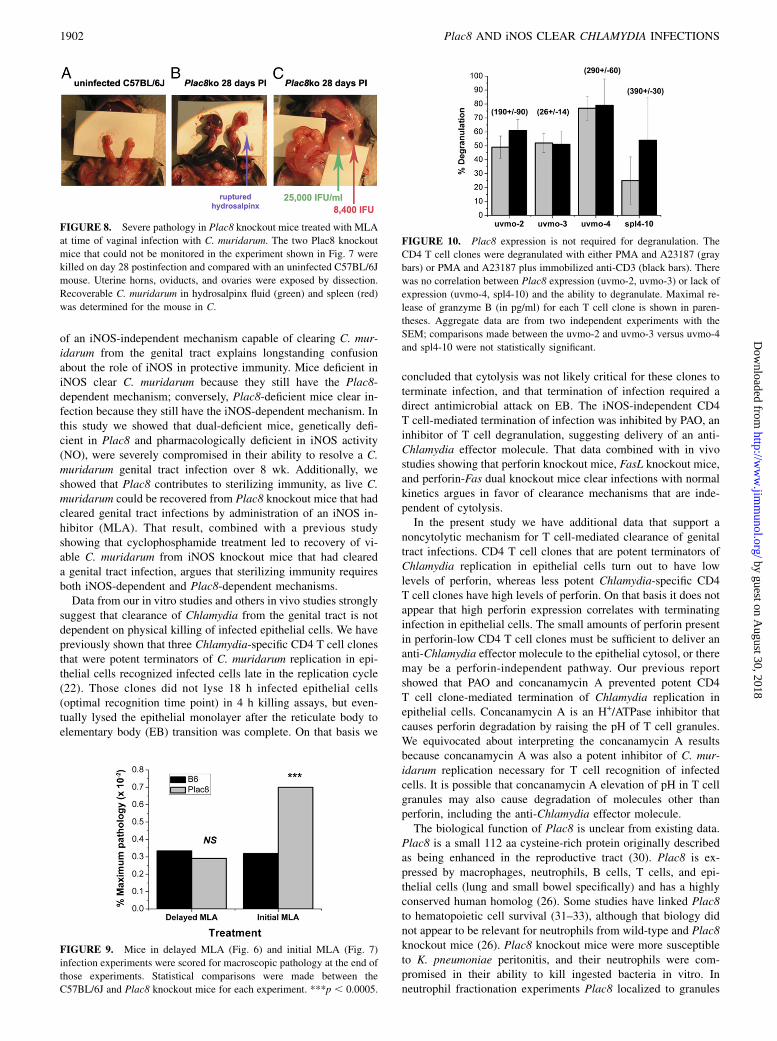
clear genital tract infections over a longer time course. Two Plac8knockout mice infected with continuous MLA drinking waterexperienced an inflammatory response that sealed the vaginalvault, making it impossible to do sequential monitoring of Chla-mydia shedding. This outcome is unique in our experience withthe C. muridarum mouse model. These mice were killed at day 28due to increasing abdominal girth. Both mice appeared to be wellby activity level but were judged to be at risk for acute decom-pensation. Gross reproductive tract pathology in these mice isshown in Fig. 8. Both Plac8 knockout mice had gross distortionof the genital tract anatomy with pan dilatation, hydrouterusor hemouterus, and severe hydrosalpinx. The luminal fluid andspleen were harvested from the second mouse and C. muridarumwas quantified on McCoy monolayers. Significant IFU were re-covered from the hydrosalpinx fluid (25,000 IFU/ml) and from thespleen (8,400 IFU), suggesting dissemination and extragenitaltract replication when both the iNOS-dependent and the Plac8-dependent mechanisms are compromised.Assessment of macroscopic genital tract pathology for the
experiments in Figs. 6 and 7 showed no difference betweenC57BL/6J mice and Plac8 knockout mice when MLA was ad-ministered on postinfection day 52 (delayed MLA), but a signifi-cant difference was shown when MLA was administered concur-rently with infection (initial MLA) (see Fig. 9).
CD4 T cell clones that do not express Plac8 have no apparentdefect in degranulation
Our previous study showed that the iNOS-independent mechanismused by uvmo-2 and uvmo-3 was dependent on T cell degranu-
lation. In neutrophils, Plac8 localizes to the granules. It is unclearfrom existing data whether Plac8 is in granules or regulatestrafficking of granules (see Discussion). With respect to degra-nulation-dependent mechanism used by uvmo-2 and uvmo-3, thetwo simplest explanations for Plac8 function would be either di-rectly as an antimicrobial effector molecule analogous to humangranulysin, or indirectly as a regulator of events in degranulation,that is, participation in delivery of an unidentified effector mole-cule. To test whether T cells that do not express Plac8 are deficientin degranulation, we compared the degranulation capacity of ourC57BL/6-derived Plac8+ CD4 T cell clones uvmo-2 and uvmo-3to our Plac82 CD4 T cell clones spl4-10 and uvmo-4. The T cellclones were degranulated with PMA/A23187 and PMA/A23187plus anti-CD3. Degranulation was measured as percentage ofgranzyme B release as measured by ELISA (Fig. 10). The fourCD4 T cell clones had a wide range of intracellular granzyme Blevels (26–390 pg/ml). There was no obvious defect in degranu-lation in any of the CD4 T cell clones or any correlation of de-granulation efficiency with presence or absence of Plac8 expres-sion. Plac8 does not appear to have a role in T cell degranulation.
DiscussionData presented in this study establish that there are two redundantmechanisms for clearance of C. muridarum from the genital tract.The first mechanism is dependent on T cell & IFN-g–mediated upregulation of epithelial iNOS transcription and NO production aspreviously described in the literature by Igietseme et al. (29). Thesecond mechanism is dependent on Plac8 and likely on T celldegranulation based on our in vitro studies (22). The existence
FIGURE 6. Plac8 is required for steril-
izing immunity. Wild-type C57BL/6J and
Plac8 female knockout mice in experimental
groups of six mice each were treated with
medroxyprogesterone and then infected vag-
inally 1 wk later with 5 3 104 IFU C. mur-
idarum. On day 52 (arrow), regular drinking
water was replaced with water containing
50 mM MLA (iNOS inhibitor). Genital tract
shedding was monitored through day 80 post-
infection. Data presented are from one ex-
periment; differences in the clearance kinet-
ics for C57BL/6J versus Plac8 knockout
mice were statistically significant with an
overall p value of 0.003.
FIGURE 7. Inhibition of iNOS at initiation of infection compromises clearance of C. muridarum from the genital tract of Plac8 knockout mice. Wild-
type C57BL/6J (11 mice) and Plac8 knockout (10 mice) female mice were treated with medroxyprogesterone and then infected vaginally 1 wk later with
5 3 104 IFU C. muridarum. The drinking water contained 50 mM MLA (iNOS inhibitor) on the day before infection through day 56 postinfection. Genital
tract shedding was monitored through day 56 postinfection. Two of the 10 Plac8 knockout mice could not be monitored due to inflammatory obstruction of
the vaginal vault; eight Plac8 knockout mice were included in the final data set. Aggregate data are from two experiments; differences in the clearance
kinetics for C57BL/6J versus Plac8 knockout mice were statistically significant with an overall p value of ,0.0001.
The Journal of Immunology 1901
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
of an iNOS-independent mechanism capable of clearing C. mur-idarum from the genital tract explains longstanding confusionabout the role of iNOS in protective immunity. Mice deficient iniNOS clear C. muridarum because they still have the Plac8-dependent mechanism; conversely, Plac8-deficient mice clear in-fection because they still have the iNOS-dependent mechanism. Inthis study we showed that dual-deficient mice, genetically defi-cient in Plac8 and pharmacologically deficient in iNOS activity(NO), were severely compromised in their ability to resolve a C.muridarum genital tract infection over 8 wk. Additionally, weshowed that Plac8 contributes to sterilizing immunity, as live C.muridarum could be recovered from Plac8 knockout mice that hadcleared genital tract infections by administration of an iNOS in-hibitor (MLA). That result, combined with a previous studyshowing that cyclophosphamide treatment led to recovery of vi-able C. muridarum from iNOS knockout mice that had cleareda genital tract infection, argues that sterilizing immunity requiresboth iNOS-dependent and Plac8-dependent mechanisms.Data from our in vitro studies and others in vivo studies strongly
suggest that clearance of Chlamydia from the genital tract is notdependent on physical killing of infected epithelial cells. We havepreviously shown that three Chlamydia-specific CD4 T cell clonesthat were potent terminators of C. muridarum replication in epi-thelial cells recognized infected cells late in the replication cycle(22). Those clones did not lyse 18 h infected epithelial cells(optimal recognition time point) in 4 h killing assays, but even-tually lysed the epithelial monolayer after the reticulate body toelementary body (EB) transition was complete. On that basis we
concluded that cytolysis was not likely critical for these clones toterminate infection, and that termination of infection required adirect antimicrobial attack on EB. The iNOS-independent CD4T cell-mediated termination of infection was inhibited by PAO, aninhibitor of T cell degranulation, suggesting delivery of an anti-Chlamydia effector molecule. That data combined with in vivostudies showing that perforin knockout mice, FasL knockout mice,and perforin-Fas dual knockout mice clear infections with normalkinetics argues in favor of clearance mechanisms that are inde-pendent of cytolysis.In the present study we have additional data that support a
noncytolytic mechanism for T cell-mediated clearance of genitaltract infections. CD4 T cell clones that are potent terminators ofChlamydia replication in epithelial cells turn out to have lowlevels of perforin, whereas less potent Chlamydia-specific CD4T cell clones have high levels of perforin. On that basis it does notappear that high perforin expression correlates with terminatinginfection in epithelial cells. The small amounts of perforin presentin perforin-low CD4 T cell clones must be sufficient to deliver ananti-Chlamydia effector molecule to the epithelial cytosol, or theremay be a perforin-independent pathway. Our previous reportshowed that PAO and concanamycin A prevented potent CD4T cell clone-mediated termination of Chlamydia replication inepithelial cells. Concanamycin A is an H+/ATPase inhibitor thatcauses perforin degradation by raising the pH of T cell granules.We equivocated about interpreting the concanamycin A resultsbecause concanamycin A was also a potent inhibitor of C. mur-idarum replication necessary for T cell recognition of infectedcells. It is possible that concanamycin A elevation of pH in T cellgranules may also cause degradation of molecules other thanperforin, including the anti-Chlamydia effector molecule.The biological function of Plac8 is unclear from existing data.
Plac8 is a small 112 aa cysteine-rich protein originally describedas being enhanced in the reproductive tract (30). Plac8 is ex-pressed by macrophages, neutrophils, B cells, T cells, and epi-thelial cells (lung and small bowel specifically) and has a highlyconserved human homolog (26). Some studies have linked Plac8to hematopoietic cell survival (31–33), although that biology didnot appear to be relevant for neutrophils from wild-type and Plac8knockout mice (26). Plac8 knockout mice were more susceptibleto K. pneumoniae peritonitis, and their neutrophils were com-promised in their ability to kill ingested bacteria in vitro. Inneutrophil fractionation experiments Plac8 localized to granules
FIGURE 8. Severe pathology in Plac8 knockout mice treated with MLA
at time of vaginal infection with C. muridarum. The two Plac8 knockout
mice that could not be monitored in the experiment shown in Fig. 7 were
killed on day 28 postinfection and compared with an uninfected C57BL/6J
mouse. Uterine horns, oviducts, and ovaries were exposed by dissection.
Recoverable C. muridarum in hydrosalpinx fluid (green) and spleen (red)
was determined for the mouse in C.
FIGURE 9. Mice in delayed MLA (Fig. 6) and initial MLA (Fig. 7)
infection experiments were scored for macroscopic pathology at the end of
those experiments. Statistical comparisons were made between the
C57BL/6J and Plac8 knockout mice for each experiment. ***p , 0.0005.
FIGURE 10. Plac8 expression is not required for degranulation. The
CD4 T cell clones were degranulated with either PMA and A23187 (gray
bars) or PMA and A23187 plus immobilized anti-CD3 (black bars). There
was no correlation between Plac8 expression (uvmo-2, uvmo-3) or lack of
expression (uvmo-4, spl4-10) and the ability to degranulate. Maximal re-
lease of granzyme B (in pg/ml) for each T cell clone is shown in paren-
theses. Aggregate data are from two independent experiments with the
SEM; comparisons made between the uvmo-2 and uvmo-3 versus uvmo-4
and spl4-10 were not statistically significant.
1902 Plac8 AND iNOS CLEAR CHLAMYDIA INFECTIONS
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

and was not found in the cytosol. SDS-PAGE eletrophoretic mo-bility suggested that Plac8 was not processed (26). Plac8 does nothave a conventional signal sequence. There is a potential signalpeptide cleavage site between aa 17 and 18, but only 7% of knownmammalian cleavage sites share these amino acid sequence char-acteristics (SignalP; http://www.cbs.dtu.dk/services/SignalP/). Ourdata and the existing Klebsiella data are most easily explained byone of two scenarios. Plac8 is either an antimicrobial effectormolecule analogous to human granulysin, or it is a critical com-ponent of the pathway that delivers or activates an anti-Chlamydiaeffector molecule in the cytosol of infected epithelial cells. In thisstudy we showed that Plac8 does not appear to have a role inT cell degranulation. It is attractive to speculate that reactivesulfhydryl groups on cysteine-rich Plac8 react with cysteine-richOmcA and OmcB in the Chlamydia EB outer membrane leadingto neutralization. Our data show that Plac8 is not ubiquitouslyexpressed by CD4 T cells. Limited by a sample size of 4, itappears that CD4 T cells that express Plac8, CD4Plac8, use bothiNOS-dependent and degranulation-dependent mechanisms forterminating C. muridarum replication in epithelial cells, whereasPlac82 CD4 T cells appear to be dependent on NO. It is plausiblethat dual effector Chlamydia-specific CD4Plac8 T cells are theoptimal protective subset in the genital tract.The susceptibility of Plac8 knockout mice to C. muridarum
genital tract infections is likely due to a defect in T cell immunitybecause 1) nude mice sufficient in neutrophils, Plac8, and iNOSare incapable of clearing primary C. muridarum primary genitaltract infections (34), 2) time to clearance in wild-type mice is notaffected by neutrophil depletion (35, 36), and 3) clearance is en-tirely dependent on MHC class II-restricted T lymphocytes (7).The details of how Plac8 is critical for cellular immunity toChlamydia remain to be determined. Interestingly, both iNOS- andPlac8-dependent mechanisms for terminating Chlamydia repli-cation in epithelial cells are not likely to be cytolytic mechanismsthat expose intracellular bacteria to extracellular effector mecha-nisms such as neutralizing Abs or neutrophils. Instead, bothmechanisms appear to kill Chlamydia within intact epithelial cells.In this “toxic bag” model, Chlamydia is trapped and neutralizedwithin the infected cells by a combination of NO and Plac8 ora Plac8-dependent effector. This type of mechanism is intellec-tually satisfying, as most CD4 T cells appear to recognize infectedepithelial cells after the noninfectious reticulate body to infectiousEB transition has occurred (23). Because Plac8 is also expressedby epithelial cells that are susceptible to infection, we speculatethat it exists in an inactive state in those cells. It is possible thatCD4 T cells not only deliver active Plac8 to infected cell cytosol,but that active Plac8 or other injected T cell molecules triggeractivation of epithelial Plac8 in an amplified final pathway.In this study CD4 T cell-conditioned media were a very potent
inhibitor of C. muridarum replication in murine epithelial cellsthrough an iNOS-dependent mechanism, being .2 logs morepotent than rIFN-g in our previous studies and those of others.This finding suggests that in vitro studies using rIFN-g (22, 37–39)may have significantly underestimated the contribution of cyto-kines to controlling C. muridarum replication in murine cells dueeither to lesser biologic activity of rIFN-g or, more likely, tocontributions of additional cytokines such as TNF-a to a cytokine-mediated anti-Chlamydia epithelial response. The iNOS promoteronce activated by IFN-g can be further upregulated by TNF-a(40). TNF-a has been shown to be an important cytokine duringgenital tract clearance (20) and in protective immunity induced byChlamydia T cell vaccines (10, 41). Scurlock et al. (42), in a re-cent paper focused on possible IL-17 contributions to Chlamydiaimmunity, showed that IFN-g and TNF-a in genital tract secre-
tions of C. muridarum-infected mice peaked on day 4 and wereessentially undetectable by day 14. Doxycycline experimentssuggest that adaptive immunity takes hold around day 10 postin-fection (43). We propose a working model in which early innateimmunity in a genital tract awash with cytokines including IFN-gand TNF-a clears 2–3 logs of C. muridarum during the first10 d infection, and that clearance of the last 3–4 logs C. mur-idarum by the adaptive cellular immune response can be accom-plished by either cytokine-dependent (iNOS) or Plac8-dependentmechanisms, whereas sterilizing cellular immunity requires bothiNOS- and Plac8-dependent mechanisms.Identifying the Plac8-dependent mechanism for clearing Chla-
mydia from the reproductive tract resolves longstanding confusionabout the role of iNOS in protective immunity to Chlamydiagenital tract infections by identifying a redundant second clearancemechanism. Investigating the CD4Plac8 T cell subset will likelycontribute to our understanding of protective immunity and hope-fully identify surrogate biomarkers for protective immunity thatprove useful during rational vaccine development. Casd1 is an un-characterized open reading frame predicted to be a transmembraneglycoprotein with an extracellular domain. We are investigatingCasd1 as a possible T cell accessory molecule, and potentially aconvenient biomarker for the CD4Plac8 T cell subset.
AcknowledgmentsCritical assistance with microarray data analysis was provided by Jeanette
McClintick and Howard Edenberg in the Indiana University Center for
Medical Genomics.
DisclosuresThe authors have no financial conflicts of interest.
References1. Brunham, R. C., B. Pourbohloul, S. Mak, R. White, and M. L. Rekart. 2005. The
unexpected impact of a Chlamydia trachomatis infection control program onsusceptibility to reinfection. J. Infect. Dis. 192: 1836–1844.
2. Brunham, R. C., and M. L. Rekart. 2008. The arrested immunity hypothesis andthe epidemiology of Chlamydia control. Sex. Transm. Dis. 35: 53–54.
3. Rekart, M. L., and R. C. Brunham. 2008. Epidemiology of chlamydial infection:are we losing ground? Sex. Transm. Infect. 84: 87–91.
4. Morrison, S. G., and R. P. Morrison. 2005. A predominant role for antibody inacquired immunity to chlamydial genital tract reinfection. J. Immunol. 175:7536–7542.
5. Su, H., K. Feilzer, H. D. Caldwell, and R. P. Morrison. 1997. Chlamydia tra-chomatis genital tract infection of antibody-deficient gene knockout mice. Infect.Immun. 65: 1993–1999.
6. Morrison, R. P., and H. D. Caldwell. 2002. Immunity to murine chlamydialgenital infection. Infect. Immun. 70: 2741–2751.
7. Morrison, R. P., K. Feilzer, and D. B. Tumas. 1995. Gene knockout mice es-tablish a primary protective role for major histocompatibility complex class II-restricted responses in Chlamydia trachomatis genital tract infection. Infect.Immun. 63: 4661–4668.
8. Farris, C. M., S. G.Morrison, and R. P. Morrison. 2010. CD4+ T cells and antibodyare required for optimal major outer membrane protein vaccine-induced immunityto Chlamydia muridarum genital infection. Infect. Immun. 78: 4374–4383.
9. Li, W., A. K. Murthy, M. N. Guentzel, J. P. Chambers, T. G. Forsthuber,J. Seshu, G. Zhong, and B. P. Arulanandam. 2010. Immunization witha combination of integral chlamydial antigens and a defined secreted proteininduces robust immunity against genital chlamydial challenge. Infect. Immun.78: 3942–3949.
10. Yu, H., K. P. Karunakaran, I. Kelly, C. Shen, X. Jiang, L. J. Foster, andR. C. Brunham. 2011. Immunization with live and dead Chlamydia muridaruminduces different levels of protective immunity in a murine genital tract model:correlation with MHC class II peptide presentation and multifunctional Th1cells. J. Immunol. 186: 3615–3621.
11. Igietseme, J. U. 1996. The molecular mechanism of T-cell control of Chlamydiain mice: role of nitric oxide. Immunology 88: 1–8.
12. Igietseme, J. U. 1996. Molecular mechanism of T-cell control of Chlamydia inmice: role of nitric oxide in vivo. Immunology 88: 1–5.
13. Igietseme, J. U., G. A. Ananaba, J. Bolier, S. Bowers, T. Moore, T. Belay, D. Lyn,and C. M. Black. 1999. The intercellular adhesion molecule type-1 is required forrapid activation of T helper type 1 lymphocytes that control early acute phase ofgenital chlamydial infection in mice. Immunology 98: 510–519.
The Journal of Immunology 1903
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

14. Igietseme, J. U., P. B. Wyrick, D. Goyeau, and R. G. Rank. 1994. An in vitromodel for immune control of chlamydial growth in polarized epithelial cells.Infect. Immun. 62: 3528–3535.
15. Igietseme, J. U., L. L. Perry, G. A. Ananaba, I. M. Uriri, O. O. Ojior,S. N. Kumar, and H. D. Caldwell. 1998. Chlamydial infection in inducible nitricoxide synthase knockout mice. Infect. Immun. 66: 1282–1286.
16. Ramsey, K. H., G. S. Miranpuri, C. E. Poulsen, N. B. Marthakis, L. M. Braune,and G. I. Byrne. 1998. Inducible nitric oxide synthase does not affect resolutionof murine chlamydial genital tract infections or eradication of chlamydiae inprimary murine cell culture. Infect. Immun. 66: 835–838.
17. Cotter, T. W., K. H. Ramsey, G. S. Miranpuri, C. E. Poulsen, and G. I. Byrne.1997. Dissemination of Chlamydia trachomatis chronic genital tract infection ing interferon gene knockout mice. Infect. Immun. 65: 2145–2152.
18. Perry, L. L., K. Feilzer, and H. D. Caldwell. 1997. Immunity to Chlamydiatrachomatis is mediated by T helper 1 cells through IFN-g-dependent and -in-dependent pathways. J. Immunol. 158: 3344–3352.
19. Perry, L. L., K. Feilzer, S. Hughes, and H. D. Caldwell. 1999. Clearance ofChlamydia trachomatis from the murine genital mucosa does not require perforin-mediated cytolysis or Fas-mediated apoptosis. Infect. Immun. 67: 1379–1385.
20. Perry, L. L., H. Su, K. Feilzer, R. Messer, S. Hughes, W. Whitmire, andH. D. Caldwell. 1999. Differential sensitivity of distinct Chlamydia trachomatisisolates to IFN-g-mediated inhibition. J. Immunol. 162: 3541–3548.
21. Prantner, D., T. Darville, J. D. Sikes, C. W. Andrews, Jr., H. Brade, R. G. Rank, andU. M. Nagarajan. 2009. Critical role for interleukin-1b (IL-1b) during Chlamydiamuridarum genital infection and bacterial replication-independent secretion of IL-1b in mouse macrophages. Infect. Immun. 77: 5334–5346.
22. Jayarapu, K., M. Kerr, S. Ofner, and R. M. Johnson. 2010. Chlamydia-specificCD4 T cell clones control Chlamydia muridarum replication in epithelial cellsby nitric oxide-dependent and -independent mechanisms. J. Immunol. 185:6911–6920.
23. Jayarapu, K., M. S. Kerr, A. Katschke, and R. M. Johnson. 2009. Chlamydiamuridarum-specific CD4 T-cell clones recognize infected reproductive tract epi-thelial cells in an interferon-dependent fashion. Infect. Immun. 77: 4469–4479.
24. Johnson, R. M. 2004. Murine oviduct epithelial cell cytokine responses toChlamydia muridarum infection include interleukin-12-p70 secretion. Infect.Immun. 72: 3951–3960.
25. Schachter, J. 1980. Chlamydiae (psittacosis-lymphogranuloma venereum-trachomagroup). In Manual of Clinical Immunology, 3rd Ed. E. H. Lennette, A. Balows,W. J. Hausler, Jr., and J. P. Truant, eds. American Society for Microbiology,Washington, D.C., p. 357–364.
26. Ledford, J. G., M. Kovarova, and B. H. Koller. 2007. Impaired host defense inmice lacking ONZIN. J. Immunol. 178: 5132–5143.
27. Ramsey, K. H., G. S. Miranpuri, I. M. Sigar, S. Ouellette, and G. I. Byrne. 2001.Chlamydia trachomatis persistence in the female mouse genital tract: induciblenitric oxide synthase and infection outcome. Infect. Immun. 69: 5131–5137.
28. Morrison, S. G., and R. P. Morrison. 2001. Resolution of secondary Chlamydiatrachomatis genital tract infection in immune mice with depletion of both CD4+
and CD8+ T cells. Infect. Immun. 69: 2643–2649.29. Igietseme, J. U., I. M. Uriri, R. Hawkins, and R. G. Rank. 1996. Integrin-
mediated epithelial-T cell interaction enhances nitric oxide production and in-creased intracellular inhibition of Chlamydia. J. Leukoc. Biol. 59: 656–662.
30. Galaviz-Hernandez, C., C. Stagg, G. de Ridder, T. S. Tanaka, M. S. Ko,D. Schlessinger, and R. Nagaraja. 2003. Plac8 and Plac9, novel placental-enriched genes identified through microarray analysis. Gene 309: 81–89.
31. Rogulski, K., Y. Li, K. Rothermund, L. Pu, S. Watkins, F. Yi, andE. V. Prochownik. 2005. Onzin, a c-Myc-repressed target, promotes survivaland transformation by modulating the Akt-Mdm2-p53 pathway. Oncogene 24:7524–7541.
32. Li, Y., K. Rogulski, Q. Zhou, P. J. Sims, and E. V. Prochownik. 2006. Thenegative c-Myc target onzin affects proliferation and apoptosis via its obligateinteraction with phospholipid scramblase 1. Mol. Cell. Biol. 26: 3401–3413.
33. Wu, S. F., Y. Huang, J. K. Hou, T. T. Yuan, C. X. Zhou, J. Zhang, andG. Q. Chen. 2010. The downregulation of onzin expression by PKCε-ERK2signaling and its potential role in AML cell differentiation. Leukemia 24: 544–551.
34. Rank, R. G., L. S. Soderberg, and A. L. Barron. 1985. Chronic chlamydialgenital infection in congenitally athymic nude mice. Infect. Immun. 48: 847–849.
35. Barteneva, N., I. Theodor, E. M. Peterson, and L. M. de la Maza. 1996. Role ofneutrophils in controlling early stages of a Chlamydia trachomatis infection.Infect. Immun. 64: 4830–4833.
36. Lee, H. Y., J. H. Schripsema, I. M. Sigar, C. M. Murray, S. R. Lacy, andK. H. Ramsey. 2010. A link between neutrophils and chronic disease manifes-tations of Chlamydia muridarum urogenital infection of mice. FEMS Immunol.Med. Microbiol. 59: 108–116.
37. Nelson, D. E., D. P. Virok, H. Wood, C. Roshick, R. M. Johnson,W. M. Whitmire, D. D. Crane, O. Steele-Mortimer, L. Kari, G. McClarty, andH. D. Caldwell. 2005. Chlamydial IFN-g immune evasion is linked to host in-fection tropism. Proc. Natl. Acad. Sci. USA 102: 10658–10663.
38. Coers, J., I. Bernstein-Hanley, D. Grotsky, I. Parvanova, J. C. Howard,G. A. Taylor, W. F. Dietrich, and M. N. Starnbach. 2008. Chlamydia muridarumevades growth restriction by the IFN-g-inducible host resistance factor Irgb10. J.Immunol. 180: 6237–6245.
39. Al-Zeer, M. A., H. M. Al-Younes, P. R. Braun, J. Zerrahn, and T. F. Meyer. 2009.IFN-g-inducible Irga6 mediates host resistance against Chlamydia trachomatisvia autophagy. PLoS ONE 4: e4588.
40. MacMicking, J., Q. W. Xie, and C. Nathan. 1997. Nitric oxide and macrophagefunction. Annu. Rev. Immunol. 15: 323–350.
41. Yu, H., X. Jiang, C. Shen, K. P. Karunakaran, J. Jiang, N. L. Rosin, andR. C. Brunham. 2010. Chlamydia muridarum T-cell antigens formulated withthe adjuvant DDA/TDB induce immunity against infection that correlateswith a high frequency of g interferon (IFN-g)/tumor necrosis factor alpha andIFN-g/interleukin-17 double-positive CD4+ T cells. Infect. Immun. 78: 2272–2282.
42. Scurlock, A. M., L. C. Frazer, C. W. Andrews, Jr., C. M. O’Connell, I. P. Foote,S. L. Bailey, K. Chandra-Kuntal, J. K. Kolls, and T. Darville. 2011. Interleukin-17 contributes to generation of Th1 immunity and neutrophil recruitment duringChlamydia muridarum genital tract infection but is not required for macrophageinflux or normal resolution of infection. Infect. Immun. 79: 1349–1362.
43. Su, H., R. Morrison, R. Messer, W. Whitmire, S. Hughes, and H. D. Caldwell.1999. The effect of doxycycline treatment on the development of protectiveimmunity in a murine model of chlamydial genital infection. J. Infect. Dis. 180:1252–1258.
1904 Plac8 AND iNOS CLEAR CHLAMYDIA INFECTIONS
by guest on August 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from