pharmacologic inhibition of jak-stat signaling promotes ...pharmacologic inhibition of jak-stat...
TRANSCRIPT

R E S EARCH ART I C L E
CL IN I CAL MED IC INE
1Department of Dermatology, Columbia University, NewYork, NY 10032, USA. 2Departmentof Systems Biology, Columbia University, New York, NY 10032, USA. 3Department ofGenetics and Development, Columbia University, New York, NY 10032, USA.*Present address:DepartmentofBioengineering, ImperialCollegeLondon, LondonSW72AZ,UK.†Corresponding author. E-mail: [email protected]
Harel et al. Sci. Adv. 2015;1:e1500973 23 October 2015
2015 © The Authors, some rights reserved;
exclusive licensee American Association for
the Advancement of Science. Distributed
under a Creative Commons Attribution
NonCommercial License 4.0 (CC BY-NC).
10.1126/sciadv.1500973
Pharmacologic inhibition of JAK-STAT signalingpromotes hair growth
Sivan Harel,1 Claire A. Higgins,1* Jane E. Cerise,1 Zhenpeng Dai,1 James C. Chen,1,2Raphael Clynes,1 Angela M. Christiano1,3†
Dow
Several forms of hair loss in humans are characterized by the inability of hair follicles to enter the growth phase(anagen) of the hair cycle after being arrested in the resting phase (telogen). Current pharmacologic therapieshave been largely unsuccessful in targeting pathways that can be selectively modulated to induce entry intoanagen. We show that topical treatment of mouse and human skin with small-molecule inhibitors of the Januskinase (JAK)–signal transducer and activator of transcription (STAT) pathway results in rapid onset of anagen andsubsequent hair growth. We show that JAK inhibition regulates the activation of key hair follicle populations suchas the hair germ and improves the inductivity of cultured human dermal papilla cells by controlling a molecularsignature enriched in intact, fully inductive dermal papillae. Our findings open new avenues for exploration ofJAK-STAT inhibition for promotion of hair growth and highlight the role of this pathway in regulating the acti-vation of hair follicle stem cells.
nlo
on April 25http://advances.sciencem
ag.org/aded from
INTRODUCTION
Several hair growth disorders are characterized by the inability to re-enter the growth phase of the hair cycle (anagen) due to hair follicle(HF)miniaturization in the case of androgenetic alopecia or immunedysfunction in the case of alopecia areata (AA). Current pharmaco-logical therapy for androgenetic alopecia is primarily focused on theprevention of further hair loss. However, the search for pharmaco-logic agents to restart the hair cycle has been largely unsatisfactory.Recently, we demonstrated that pharmacological inhibition of theJanus kinase (JAK)–signal transducer and activator of transcription(STAT) pathway promotes rapid hair regrowth in AA in both miceand humans (1). Unexpectedly, during the course of our studies onmice with AA, we observed that topical treatment with JAK-STATinhibitors resulted in an unusually robust hair growth, suggesting alocalized effect on initiation of the hair cycle. Here, we show that phar-macological inhibition of JAK-STAT signaling initiates the hair cyclein normal mice and promotes growth of hair follicles in humans.
, 2020
RESULTS
JAK-STAT inhibition results in rapid onset of hair growth inmiceWe first treated C57/B6 mice in telogen on half of the dorsal back witheither vehicle control (negative control, left side), a sonic hedgehog (Shh)agonist previously shown to promote anagen initiation (2) (positivecontrol), or several small-molecule inhibitors of the JAK-STAT path-way, including tofacitinib (JAK1/3 > JAK2 > TYK2) (3) and ruxolitinib(JAK1/2>Tyk2> JAK3) (4–7) (right side). As expected, entry into anagenwas evident within 7 days of treatment with the Shh agonist, whereasvehicle-treated mice remained in telogen for the duration of the exper-iment. Intriguingly, treatment with the JAK inhibitors resulted in rapidreentry into the hair cycle, with kinetics similar to the Shh agonist (Fig.1A). To examine a direct effect on stem cell activation, we next treated
mice in telogen with tofacitinib or ruxolitinib for a short period of 4 days.Significant proliferation was noted within the hair germ compartment(P-cadherin+) of drug-treated HFs (fig. S1A), indicating activation of aprogenitor population. Quantification of the effects of drug treatmenton skin homeostasis demonstrated that drug-induced hair growth reca-pitulates normal hair growth (fig. S1B). Together, these data suggestthat local inhibition of the JAK-STAT pathway results in rapid onsetof hair growth.
Effects of JAK-STAT inhibition are dependent on theduration of telogenThe first postnatal hair cycle in C57/B6 mice follows a precise temporalprogression. Thereafter, regeneration (anagen reentry) occurs in spon-taneous, patchy waves, starting around 12 to 13 weeks of age (8, 9).Treatment of mice in telogen with JAK inhibitors consistently yieldedearly and homogeneous hair growth; however, the duration of treat-ment required to reinitiate anagen was inexplicably variable. To addressthis issue, we treated mice in early telogen (7 weeks) or mid-telogen(8.5 weeks). The treatment resulted in no growth in the 7-week-oldmice, whereas 8.5-week-old mice exhibited rapid onset of anagen (fig.S1C). To ensure that mice treated at 8.5 weeks of age were still at therefractory stage of telogen that arrests the propagation of anagen reentry(9, 10), we plucked hair from mice at 7 and 8.5 weeks and showed thathair growth after plucking followed a similar kinetics in both timepoints (fig. S1C). Notably, longer treatment of 7-week-old mice for 18to 21 days eventually induced hair growth, but only after the treatedmice had reached 8.5 weeks of age (fig. S1D). This finding implies thatJAK inhibition cannot override the quiescence-promoting micro-environment at the early stages of telogen (9, 10) but is sufficient to pro-mote hair growth at a later stage in telogen.
To demonstrate the robustness and reproducibility of hair growthresulting from treatment with JAK inhibitors at 8.5 weeks of age, we tookadvantage of skin darkening as a proxy for hair growth in C57/B6 mice(11). Indeed, ~90% of 8.5-week-old mice treated with ruxolitinib ortofacitinib for 5 days displayed skin darkening and hair growth within10 days of starting treatment, whereas no hair growth was evident incontrol-treated mice (P < 0.0001 for ruxolitinib treatment and P =0.04 for tofacitinib treatment) (Fig. 1B and fig. S1E).
1 of 12

R E S EARCH ART I C L E
on April 25, 2020
http://advances.sciencemag.org/
Dow
nloaded from
Fig. 1. Inhibition of JAK-STAT signaling restarts anagen inwild-typemice. (A) Seven-week-old wild-typemicewere shaved and treated daily with either atopical application of vehicle control, sonic hedgehog agonist (SAG), 3% ruxolitinib (JAK1/2 inhibitor), or tofacitinib (JAK3 inhibitor). Skin was harvested at the
indicated time points and stainedwith hematoxylin and eosin (H&E). Images ofmicewere taken at D21 of treatment. (B) Mice (8.5 weeks old) were treatedwithruxolitinib, tofacitinib, or vehicle control for 5 days and were monitored for the appearance of skin pigmentation, signaling the initiation of anagen. No hairgrowth (and nopigmentation)was assigned the arbitrary value of 0. Skin darkeningwas given a value from0 to 100%,with the higher number indicating darkerskin/visible hair growth. Five mice were used per condition. Nonparametric longitudinal data analysis was performed over days 8 to 18 after treatment togenerate P = 7.6 × 10−34 for control versus ruxolitinib and P = 1.5 × 10−10 for control versus tofacitinib. Double asterisks indicate high statistical significance.(C) Whole skin frommice treated with vehicle control, ruxolitinib, and tofacitinib for 4 days was analyzed bymicroarrays. Expression data were used to identifygenes thatweredifferentially expressedbetween T0 andT5 in eachof the three treatment groups. Threemicewere usedper condition andbiopsied at two timepoints. (D) IPAwas used to identify themolecular pathways andprocesses thatwereoverrepresented in our lists of differentially expressedgenes. Comparisonofthe differentially expressed gene lists revealed a subset of genes regulated by both ruxolitinib and tofacitinib. Red, genes up-regulated in drug-treated T5 versusT0; green, genes down-regulated in drug-treated T5 versus T0. (E) Mice (8.5 weeks old) were treatedwith ruxolitinib or tofacitinib on one side of their dorsal skinandwith vehicle control on the other side. Four hours after treatment, Eduwas injected into eachmouse, and skin was harvested 1 hour later. Treatments wereperformed once, twice, and thrice, and skin was analyzed for the presence of Edu+ cells. DAPI, 4′,6-diamidino-2-phenylindole.Harel et al. Sci. Adv. 2015;1:e1500973 23 October 2015 2 of 12

R E S EARCH ART I C L E
on April 25, 2020
http://advances.sciencemag.org/
Dow
nloaded from
Hair growth after JAK-STAT inhibition mimics normal anageninitiation by activating the Wnt and Shh signaling pathwaysTo examine whether anagen initiation after treatment with JAK inhibi-tors is molecularly similar to normal anagen initiation, we performedmicroarray experiments on 8.5-week-oldmice treated with vehicle con-trol, ruxolitinib, or tofacitinib for 4 days, a time point at which prolif-eration in the hair germ has begun but hair growth is not yet evident.Comparison of the differentially expressed gene lists between wholeskin harvested at day 0 (T0) and day 4 (T5) of treatment revealed a sub-set of genes regulated by both JAK inhibitors (Fig. 1C). Pathway analysisusing Ingenuity Pathway Analysis (IPA) software showed that melano-genesis and theWnt pathway were enriched in both ruxolitinib and to-facitinib treatments, but not in the vehicle treatment. Further analysisof differentially expressed genes in both drug treatments identifiedother important hair cycle regulators, such as Shh and Prom1 (2, 12–16),as significantly up-regulated in JAK inhibitor–treated skin (Fig. 1D).Differential regulation of key genes was verified by quantitative poly-merase chain reaction (qPCR) (fig. S2A). Because up-regulation ofWnt and Shhpathways is central for anagen initiation and for activationofmelanocyte stem cells (16, 17), these findings suggest that blockade ofJAK-STAT signaling allows for normal progression of the hair cycle.Further analysis of genes regulated by only one of the drug treatmentsrevealed a distinct molecular signature (fig. S2, B and C). Ruxolitinibtreatment enriched for the mTOR (mammalian target of rapamycin)and NFkB (nuclear factor kB) pathways, both previously shown to beinvolved in hair cycle regulation (12, 18–20), whereas tofacitinib treat-ment enriched for pathways involved in cell motility and migration,such as Rho and integrin signaling. Stat3−/− keratinocytes were previ-ously shown to be deficient in migration in response to stimuli (21, 22),suggesting that the JAK-STATpathway is essential for cellular motility inthe transition between telogen and anagen.
JAK-STAT inhibition causes activation of HF progenitor cellsTo investigate the cellular mechanisms responsible for HF activationafter JAK-STAT inhibition, we treated mice in telogen with ruxolitinib,tofacitinib, or vehicle control and harvested skin 5 hours after the first,second, and third treatments (Fig. 1E, schematic). Edu (5-ethynyl-2′-deoxyuridine)was injecteddaily, 1hourbeforeharvesting each timepoint,and skin samples were analyzed for the presence of Edu+ (proliferating)cells. Edu+ cells are clearly visible within the hair germ (P-cadherin+)compartment after three treatments in both ruxolitinib- and tofacitinib-treated skin but not in control-treated skin (Fig. 1E). This finding sug-gests that activation of the HF after JAK-STAT inhibition follows thenormal anagen initiation kinetics, in which hair germ proliferation pre-cedes bulge stem cell proliferation (23). Together, the data imply thatJAK-STAT signaling normally acts to prevent anagen reentry and thatJAK blockade relieves this inhibition to allow for normal hair cycleprogression.
Hair-inducing effects of JAK inhibition are not dependent onthe activity of lymphocytesThe JAK-STAT pathway is known to play a prominent role in T cellbiology (24), and theHFmicroenvironment contains a substantial pop-ulation of resident and migrating T cells. Recent studies suggested thatgd T cells secrete factors that regulate HF neogenesis in mice, as well asaspects of hair cycling (25, 26). Moreover, we previously demonstratedthat in AA, JAK-STAT inhibitors act to clear cytotoxic T cells from theHFmicroenvironment, an essential process for the onset of hair regrowth.
Harel et al. Sci. Adv. 2015;1:e1500973 23 October 2015
To evaluate whether normal hair growth after treatment with JAKinhibitors is mediated by T cells, we examined the effects of topicaldrug treatments on two different lymphocyte-deficient mouse models.Rag1−/−mice, deficient in B andT cells, andTcr b/d−/−mice, deficient inmigrating and resident T cells, were largely indistinguishable fromcontrols in terms of their ability to enter the hair cycle (fig. S3A), as wellas their response to drug treatment (Fig. 2A). This suggests that the hair-inducing effects of JAK inhibitors in normal skin are not dependenton the activity of lymphocytes. Although we have not ruled out theeffects of JAK-STAT inhibitors on skin macrophages (27) or myeloiddendritic cells (28), this result suggests that the anagen-inducing effectsof JAK-STAT inhibition likely represent a hair-intrinsic property.
The JAK-STAT pathway is dynamically regulated across thehair cycleTo examine the kinetics of the JAK-STAT pathway in HF developmentand cycling, we analyzed changes in gene expression in whole skinduring the transition fromanagen to telogen (29). Using a qPCRarray,wemeasured the expression of genes related to the JAK-STATpathway.Dynamic changes in gene expression were visualized using Gene Ex-pressionDynamics Inspector (GEDI), an algorithm that clusters expres-sion values for each gene on the qPCR array into metagene categorieson the basis of the similarities in their temporal expression profiles,placing them into 1 of 30 places on a 5 × 6 grid. Comparison betweencatagen (D17) and early anagen (D29) revealed a cluster of metagenesthat became repressed as the hair cycle progressed (boxed pixels, Fig.2B). Investigating the content of the repressed metagenes revealed thatkey genes in the JAK-STATpathway such as Stat5A/B, Stat3, Jak1, Jak3,and Socs2/3 were expressed at high levels in catagen and telogen andwere repressed in early anagen (Fig. 2, B and C, and fig. S3B).
Immunofluorescence studies of HF in anagen, catagen, and telogenconfirmed that activated (phosphorylated) Stat3 is expressed in thedermal papilla (DP), some extrafollicular cells, and the proliferatingcells of the basal epidermis (Fig. 2D and fig. S3C). In catagen and telo-gen, phospho-Stat3 can also be detected in cells of the hair germ. Acti-vated phospho-Stat5 is strongly expressed in the DP throughout thehair cycle, with expression peaking during catagen, where it can alsobe detected in the bulge (Fig. 2D). The striking expression pattern ofphospho-Stat5 in key HF stem cell compartments in telogen under-scores a potentially important role in regulation of quiescence.
Tofacitinib treatment promotes growth of human HFsWe next examined the effects of JAK inhibition on hair growth inhuman tissues. In contrast tomice, human scalpHFs grow asynchronous-ly and 90% of them are in the anagen phase of the hair cycle at anygiven time (30). Therefore, it is extremely difficult to assess the transi-tion between telogen to anagen in humans, and analysis is confined tomeasuring the rate of hair fiber elongation. Human fetal scalp skin wasgrafted onto SCID (severe combined immunodeficient) mice andallowed to recover for at least 6 weeks. Each graft was then treated withdaily topical application of vehicle control on one side and either tofa-citinib (Fig. 3A) or ruxolitinib (fig. S4A) on the other side. Small HFswere already present within the grafts, and therefore, we tested the ef-fect of JAK inhibition on the ongoing development of fetal HFs toterminal hairs andnot onHFneogenesis. Tofacitinib treatment resultedin denserHF growth as compared to controls, suggesting that tofacitinibtreatment increased the rate of hair elongation. To quantify this out-come, wemeasured the intensity of pigmentation as a proxy for density
3 of 12

R E S EARCH ART I C L E
on April 25, 2020
http://advances.sciencemag.org/
Dow
nloaded from
Fig. 2. The JAK-STAT pathway is dynamically regulated during HF cycling. (A) Eight-week-old Rag1−/− and Tcr b/d−/− mice were treated daily with JAKinhibitors.Micewere treated for aweek, and imageswere taken7days after cessationofdaily treatments. Representativepictures of threemicepergenotypeare
shown. (B)Whole skinwasharvested frommiceat postnatal day17 (D17) (catagen), day23 (telogen), day29 (early anagen), andday33 (mid-anagen). Changes ingene expression were analyzed using a JAK-STAT qPCR array that includes genes involved in the JAK-STAT pathway as well as normalizing controls (Qiagen).Three mice were used per time point, each hybridized to a separate qPCR plate. Log 2 fold changes in gene expression were used to generate GEDI self-organizing maps to visualize the dynamic changes in gene expression over the hair cycle. GEDI clusters transcripts into metagenes on the basis of their similarexpression pattern over time and placed them on a 5 × 6 grid. Metagenes repressed in experimental samples (D23, D29, and D33, respectively) versus controlsamples (D17) are in green to blue, whereas metagenes overrepresented in experimental samples are in red. The upper and lower thresholds correspond to atwofold change. Changes larger/smaller than 2× are set tomaximumcolor. (C) The gene content of the repressedmetagenes highlighted in (A) (boxed pixels) isdetailed in the table. The color of boxedpixels corresponds to the color of lines in the table. (D) Skin fromwild-typemice in anagen (day30), catagen (day42), andtelogen (D50) was harvested, fixed, and stained with anti–phospho-Stat3 (anti–p-Stat3) and anti–phospho-Stat5 (anti–p-Stat5), as well as with Krt15 (a bulgemarker) and P-cadherin (hair germ marker). Phospho-Stat3 is expressed in extrafollicular cells during anagen, as well as in DP cells (white arrows). In catagen,phospho-Stat3 is expressed in the DP (red arrow) and the hair germ (orange arrow). In early telogen, phospho-Stat3 is present in the hair germ cells that areclosest to theDP (greenarrow). Phospho-Stat5 is strongly expressed in theDP throughout thehair cycle,with expressionpeakingduring catagen (yellowarrows).Phospho-Stat5 can also be detected in the bulge of catagen follicles (magenta arrow). Images were takenwith a Zeiss confocal microscope (×40magnification).Harel et al. Sci. Adv. 2015;1:e1500973 23 October 2015 4 of 12
R E S EARCH ART I C L E
on April 25, 2020
http://advances.sciencemag.org/
Dow
nloaded from
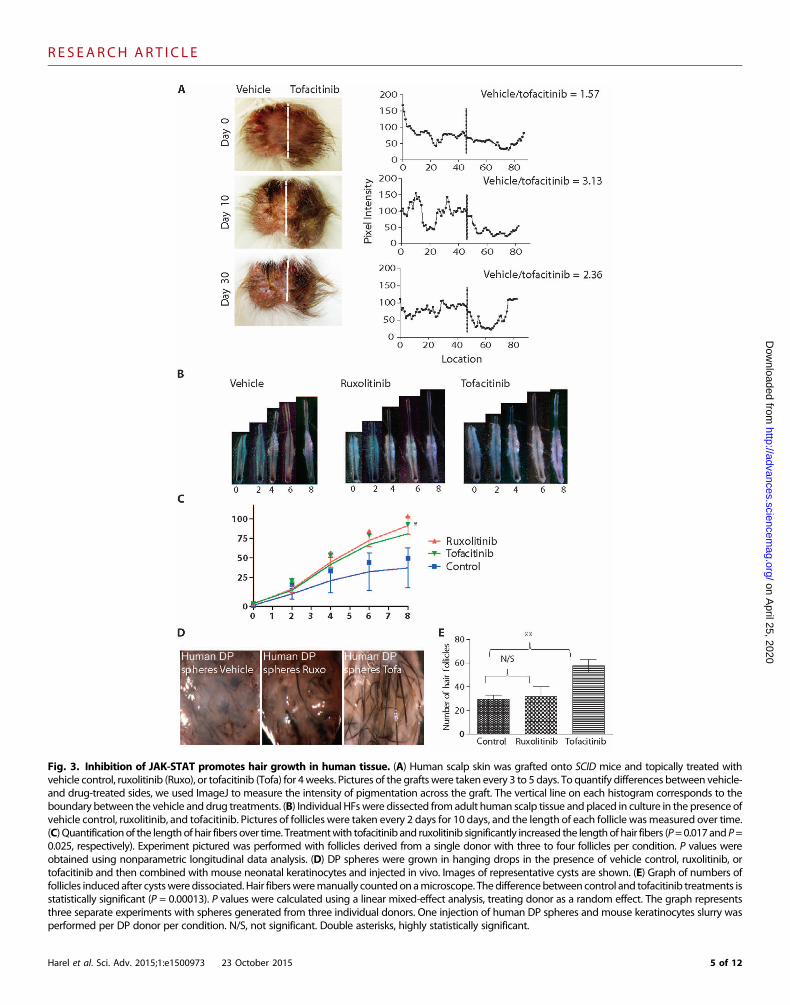
Fig. 3. Inhibition of JAK-STAT promotes hair growth in human tissue. (A) Human scalp skin was grafted onto SCID mice and topically treated withvehicle control, ruxolitinib (Ruxo), or tofacitinib (Tofa) for 4weeks. Pictures of the grafts were taken every 3 to 5 days. To quantify differences between vehicle-
and drug-treated sides, we used ImageJ to measure the intensity of pigmentation across the graft. The vertical line on each histogram corresponds to theboundary between the vehicle and drug treatments. (B) Individual HFswere dissected fromadult human scalp tissue and placed in culture in the presence ofvehicle control, ruxolitinib, and tofacitinib. Pictures of follicles were taken every 2 days for 10 days, and the length of each follicle wasmeasured over time.(C) Quantificationof the lengthof hair fibers over time. Treatmentwith tofacitinib and ruxolitinib significantly increased the lengthof hair fibers (P=0.017and P=0.025, respectively). Experiment pictured was performed with follicles derived from a single donor with three to four follicles per condition. P values wereobtained using nonparametric longitudinal data analysis. (D) DP spheres were grown in hanging drops in the presence of vehicle control, ruxolitinib, ortofacitinib and then combined with mouse neonatal keratinocytes and injected in vivo. Images of representative cysts are shown. (E) Graph of numbers offollicles induced after cystsweredissociated. Hair fibersweremanually countedonamicroscope. Thedifferencebetween control and tofacitinib treatments isstatistically significant (P = 0.00013). P values were calculated using a linear mixed-effect analysis, treating donor as a random effect. The graph representsthree separate experiments with spheres generated from three individual donors. One injection of human DP spheres and mouse keratinocytes slurry wasperformed per DP donor per condition. N/S, not significant. Double asterisks, highly statistically significant.Harel et al. Sci. Adv. 2015;1:e1500973 23 October 2015 5 of 12

R E S EARCH ART I C L E
on April 25, 2020
http://advances.sciencemag.org/
Dow
nloaded from
of dark hairs on grafted white mice and showed that the ratio of tofa-citinib/vehicle increased with days of treatment. Experiments were per-formed with skin derived from different donors with similar results(fig. S4B).
We further analyzed the effects of JAK inhibition on hair shaftelongation using the human HF organ culture model. Individual HFswere microdissected from human adult scalp tissue and cultured withvehicle control, ruxolitinib, and tofacitinib (Fig. 3B). We found thattreatment with JAK inhibitors significantly increased the length of hairshafts when treatedwith ruxolitinib and tofacitinib, indicating a positiveeffect on the rate of hair elongation (P = 0.023 and P = 0.025 for tofa-citinib and ruxolitinib, respectively). Experiments with HFs from twoadditional donors yielded a similar trend (fig. S4C). Together, the datasuggest that JAK-STAT inhibition promotes faster hair fiber growth inthe organ culture model.
Tofacitinib treatment promotes inductivity of DPBecause phospho-Stat5 is strongly expressed inmouse DP in catagenand telogen (Fig. 2D), we confirmed that phospho-STAT3 is present inthe dermal sheath andDPof humanHFs in anagen andphospho-STAT5expression is weakly present in the top portion of the DP (fig. S4D).
We recently demonstrated that growing humanDP cells in three-dimensional (3D) spheres improves their capacity to induce HF growth(31). To examine the effects of JAK inhibition on the potency of HFinduction, human DP spheres were cultured with vehicle control, rux-olitinib, or tofacitinib and then combined with neonatal mouse kerati-nocytes and injected into nude mice. This patch assay has been shownto recapitulateHFmorphogenesis and serves to evaluate trichogenic ca-pacity (32). We found that tofacitinib-treated human DP spheres in-duced larger and significantly greater numbers of HFs overall (P =0.00013) (Fig. 3D), suggesting that the inductivity of human DP isenhanced by inhibition of JAK1/3 signaling.
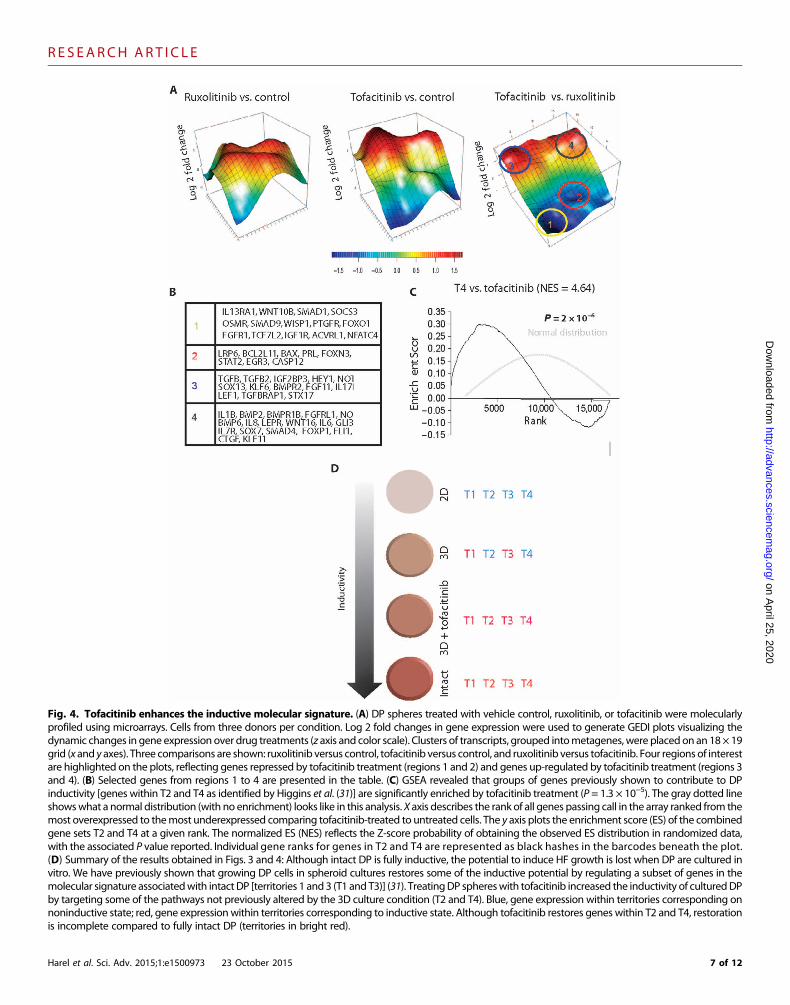
Tofacitinib treatment promotes hair growth by targetinggenes enriched in fully inductive DPTo investigate themechanisms bywhich tofacitinib treatment improvesDP inductivity, we performed microarray experiments on control-,ruxolitinib-, and tofacitinib-treated DP spheres. Log 2 fold changes ingene expression were used to generate GEDI plots. To analyze relevantchanges in gene expression, we compared ruxolitinib treatment (which didnot confer enhanced inductivity) to controls, tofacitinib treatment (whichdid enhance inductivity) to controls, and ruxolitinib and tofacitinib treat-ments to each other. This allowed us to examine gene expressionchanges resulting from JAK inhibition provided by both drugs andfocus on changes that were unique to tofacitinib treatment. The GEDIalgorithm clustered differentially expressed transcripts into metageneson the basis of their similar expression pattern across all microarrays.Data are presented in 3D form, where the z axis and colors correspondto changes in gene expression, and the x and y axes correspond tocoordinates of GEDI metapixels, plotted on an 18 × 19 grid (Fig. 4A).On the basis of the topography of the graphs, we selected four regions ofinterest (regions 1 to 4).
Among genes that were repressed in both treatments but lower intofacitinib treatment (region 1) were receptors known to be involvedin the modulation of DP inductivity, such as FGFR1, ACVRL1, IGFR1,OSMR, and PTGFR (33–38). In the region including genes thatwere up-regulated by ruxolitinib treatment but down-regulated by tofacitinibtreatment, we identified proapoptotic genes such as BAX, BCL2L11,
Harel et al. Sci. Adv. 2015;1:e1500973 23 October 2015
and CASP12 (region 2). Genes up-regulated by tofacitinib treatment(regions 3 and 4) included members of the transforming growthfactor–b (TGFb) pathway and the bone morphogenetic protein(BMP) pathway, previously shown to play a crucial role in DP induc-tivity (37–41). Key regulators of the WNT pathway, such as LEF1,known to regulate dermal-epidermal interactions (42, 43), and mem-bers of the NOTCH pathway known to control HF fate (44, 45) wereoverrepresented in tofacitinib treatment. Together, this suggests that to-facitinib treatment promotes inductivity by regulating both the induc-tive function and the survival of cultured DP.
To directly compare tofacitinib treatment with published work onDP inductivity, we turned to our previous study, which investigatedthe molecular differences between freshly isolated human DP (whichmaintain their capacity to induce hair growth), culturedDP (which losetheir inductive potential), and cultured dermal spheroids (which haverestored inductive potential), and identified unique gene signatures as-sociatedwith each state (31). These genes clustered by coexpression intofour categories that we referred to as territories (T1 to T4). We foundthat T1 and T3 contained genes whose expression was deregulated inculture and restored by growth in spheroids, whereas T2 andT4 includ-ed genes whose expression was not restored by spheroid culture (31).The genes within T2 (up-regulated in cultured cells and not restored bysphere formation) and T4 (down-regulated in cultured cells and not re-stored by sphere formation) represent molecular signatures that appearto be required for fully inductive DP.
Since tofacitinib treatment enhanced hair growth, we assayedwhether the expression of genes within T1 to T4 is enriched in a statis-tically significant manner, when comparing tofacitinib-treated to un-treated samples. Gene set enrichment analysis (GSEA) (46) on genesdifferentially expressed in these treatment arms, ranked by P valuefrom the highest to the lowest expression, revealed that territory T4 wassignificantly enriched in tofacitinib-treated spheres (P = 2 × 10−6; genelist provided in table S3) (Fig. 4, C and D). T2 was also enriched bytofacitinib treatment; however, the statistical overlap was only margin-ally significant (P = 0.03) (fig. S4E). Not all genes within T2 and T4changed in themanner predicted by our previous study, suggesting thatrestoration of inductivity by tofacitinib treatment may not be complete.Tofacitinib treatment did up-regulate the expression of genes such asLEF1, WIF1, and CD133, all known regulators of hair growth (37, 38),providing a mechanistic explanation as to why tofacitinib improved in-ductivity. Analysis of the ES of all JAK-STAT genes identified by theqPCRmicroarray used in Fig. 2B within T2 and T4 uncovered a highlystatistically significant overlap (1.45 × 10−7, gene list provided in tableS1), suggesting that there is a JAK-STAT regulatory role in the T2 andT4 inductive signature.
DISCUSSION
Our findings demonstrate that inhibition of JAK-STAT signaling pro-motes hair growth by stimulating the activation and/or proliferation ofHF stem cells, highlighting the role of this pathway in maintenance ofHF quiescence. The observation that inhibition of JAK-STAT signalscan promote activation or differentiation of stem/progenitor cells is notunique to the HF. Loss of Stat5 in hematopoietic stem cells induces exitfrom a quiescent state, leading to increased bone marrow–repopulatingcapacity after irradiation (47). Inhibitionof JAK-STATsignaling improvesskeletal muscle regeneration in aged mice by promoting symmetric
6 of 12
R E S EARCH ART I C L E
on April 25, 2020
http://advances.sciencemag.org/
Dow
nloaded from
Fig. 4. Tofacitinib enhances the inductive molecular signature. (A) DP spheres treated with vehicle control, ruxolitinib, or tofacitinib were molecularlyprofiled using microarrays. Cells from three donors per condition. Log 2 fold changes in gene expression were used to generate GEDI plots visualizing the
dynamic changes in gene expression over drug treatments (z axis and color scale). Clusters of transcripts, grouped intometagenes, were placedon an 18× 19grid (x and y axes). Three comparisons are shown: ruxolitinib versus control, tofacitinib versus control, and ruxolitinib versus tofacitinib. Four regions of interestare highlighted on the plots, reflecting genes repressed by tofacitinib treatment (regions 1 and 2) and genes up-regulated by tofacitinib treatment (regions 3and 4). (B) Selected genes from regions 1 to 4 are presented in the table. (C) GSEA revealed that groups of genes previously shown to contribute to DPinductivity [genes within T2 and T4 as identified by Higgins et al. (31)] are significantly enriched by tofacitinib treatment (P = 1.3 × 10−5). The gray dotted lineshowswhat a normal distribution (with no enrichment) looks like in this analysis. X axis describes the rank of all genes passing call in the array ranked from themost overexpressed to themost underexpressed comparing tofacitinib-treated to untreated cells. The y axis plots the enrichment score (ES) of the combinedgene sets T2 and T4 at a given rank. The normalized ES (NES) reflects the Z-score probability of obtaining the observed ES distribution in randomized data,with the associated P value reported. Individual gene ranks for genes in T2 and T4 are represented as black hashes in the barcodes beneath the plot.(D) Summary of the results obtained in Figs. 3 and 4: Although intact DP is fully inductive, the potential to induce HF growth is lost when DP are cultured invitro. We have previously shown that growing DP cells in spheroid cultures restores some of the inductive potential by regulating a subset of genes in themolecular signature associatedwith intact DP [territories 1 and3 (T1 and T3)] (31). TreatingDP sphereswith tofacitinib increased the inductivity of culturedDPby targeting some of the pathways not previously altered by the 3D culture condition (T2 and T4). Blue, gene expression within territories corresponding onnoninductive state; red, gene expressionwithin territories corresponding to inductive state. Although tofacitinib restores genes within T2 and T4, restorationis incomplete compared to fully intact DP (territories in bright red).Harel et al. Sci. Adv. 2015;1:e1500973 23 October 2015 7 of 12

R E S EARCH ART I C L E
on April 25, 2020
http://advances.sciencemag.org/
Dow
nloaded from
satellite cell expansion and reduced commitment to myogenesis (48, 49).These findings are consistent with the association of JAK-STAT withquiescence in the HF (29) and with evidence for a role of Stat3 in pro-gression of the normal hair cycle in adult mice (21, 22). Moreover, re-cent studies have shown that increased JAK-STAT signaling in agedmice inhibits HF stem cell function in vitro (50) and that Stat5 signalingcontrols HF stem cell quiescence during pregnancy and lactation (51).Therefore, the role of JAK-STAT signaling in promoting quiescencemay represent a generalized mechanism in adult stem cell populations.
Here, we observed that JAK inhibitor–mediated hair growth wasindependent of T lymphocyte function and likely represents a hair-intrinsic property. We recently demonstrated that treatment of AApatients with JAK inhibitors led to hair regrowth due in part to clear-ance of theCD8+NKG2D+ cytotoxic T cell infiltrate but did not rule outa direct effect on the HF (1). These two findings are reconciled whenconsidering hair growth in AA patients as a two-step mechanism: first,the T cell–mediated immune attack on epithelial cells must be elimi-nated, and second, anagen growthmust be reinitiated (52).Weobservedthat topical treatment with JAK inhibitors resulted in more robust hairgrowth than did systemic treatment in AA, likely because it increasesthe local concentration of drug in the HF microenvironment, allowingboth actions to occur. In unaffected individuals or in normalmice, treat-ment with JAK inhibitors may be sufficient to restart the hair cycle (inmice) or promote hair growth (in humans).
In mice, suppression of JAK signaling activates a pro-growth/anti-quiescence signal during telogen (9, 53, 54), thereby allowingreentry into anagen. We observed that activation of the hair germcompartment containing progenitor cells is an early event in JAKinhibition–mediated hair growth and noted that pathways activatedin early anagen are up-regulated after JAK inhibition. These resultssuggest that JAK inhibition–mediated hair growth follows the normalproliferative pattern of homeostatic hair cycling (16, 23).
Anagen reentry after drug treatment occurswhenmice are treated inmid-telogen but not in early telogen, suggesting that JAK inhibitioncannot override the quiescence-promoting microenvironment in earlytelogen. Several crucialmolecular events differentiate early andmid-latetelogen. BMP inhibitors and Wnt agonists rise over telogen, reducingthe threshold required for HF stem cell activation (9). Tgfb2 and Fgf7/10 up-regulation in the DP attenuates BMP signaling in the quiescence/activation step and contributes to early anagen initiation (23, 41). Astelogen progresses, the hair germ up-regulates genes involved in entryinto the cell cycle and signal transduction (23). Treatment with tofaci-tinib up-regulated the expression of TGFb2, BMP6, andLEF1 in humanDP spheres, providing a potential mechanism by which JAK inhibitionactivates theDP.Wepostulate that JAK inhibition of theDP (or the hairgerm) is buffered by opposing signals at early stages of telogen, but asthe environment becomes more permissive, the signal can induce acti-vation. Further analysis of the molecular consequences of inhibitingJAK-STAT in the DP or the hair germ will clarify the role of thispathway in regulation of HF stem cells.
Our data suggest that JAK inhibition acts at the level of hair germ–DP crosstalk, which may occur if JAK-STAT inhibition directly targetsthe hair germ or, alternatively, if it targets the DP, which then activatesthe hair germ. We observed that phospho-Stat5 is highly expressed inthe DP, suggesting potential functional importance. Moreover, by ex-amining published microarray data from mouse hair cycle (23), wenoted that Jak2, Stat5A/B, Stat3, and Socs2/3 are all up-regulated inthe DP as compared to the bulge and the hair germ. In addition, several
Harel et al. Sci. Adv. 2015;1:e1500973 23 October 2015
factors involved with inhibition of hair growth or induction are highlyexpressed during telogen. Previous studies have shown that interleukin-6(IL-6) and its familymember oncostatinM (OSM), both ofwhich signalvia the JAK-STATpathway,modulate hair growth. Inmice, overexpres-sion of IL-6 in keratinocytes causes retarded hair growth (55), andinjection of recombinant IL-6 into anagen skin caused premature onsetof catagen (56). IL-6 is also up-regulated in balding human DP versusnonbalding DP (56). IL-6 (56) and OSM (36) inhibit hair shaft elonga-tion in the human organ culture model. These findings suggest thatmultiple pathways can activate JAK-STAT signals in different compart-ments, and it is likely that the dynamic interplay of these signals driveshair growth initiation.
In human hair follicle assays, we show that JAK inhibition via tofaci-tinib treatment increases the growth rate of anagen hair shafts (skin graftsand organotypic culture assays) and enhances the inductivity of humanDP spheres (neogenesis assays). However, unlike in mice, an experimen-tal system that directly tests humanHF regeneration does not exist, so wecannot determine whether JAK inhibition drives human HF in telogeninto the anagen phase. It is surprising that ruxolitinib treatment did notimprove the inductivity of human DP spheres, despite the fact that it in-creased the rate of growth in the organ culture model. We postulate thatdown-regulation of proapoptotic signals in tofacitinib-treated spheres,which did not occur in ruxolitinib-treated spheres, may promote survivalof DP cells, leading to enhanced hair growth in this assay. Investigation ofthe molecular effects of tofacitinib treatment revealed that the treatmentcauses a molecular restoration of a subset of genes that are disrupted inculture but are present in fully inductive DP cells (Fig. 4, C and D). Thisfinding suggests that JAK inhibitionmay enhance applications such as au-tologous cell transplantation approaches for the treatment of hair loss.
JAK inhibitors are currently approved by the U.S. Food and DrugAdministration for the treatment of myeloproliferative diseases (ruxoli-tinib) (57, 58) and rheumatoid arthritis (tofacitinib) (59), and there areongoing clinical trials demonstrating the efficacy of these drugs in treat-ment of psoriasis (60–62), Vitiligo (63), andAA (1, 64, 65). Our studyraises the possibility that in addition to immune-driven conditions,JAK inhibitionmay prove useful in directly targeting tissue stem cellsand their respective niches, opening new avenues for exploration ofJAK-STAT inhibition for promoting hair growth in humans.
MATERIALS AND METHODS
Study designWe hypothesized that inhibition of JAK-STAT signaling promotesentry into the hair cycle in mice. To determine the onset of anagen,back-skin hairs were trimmed with an electric shaver, and entry ofHF into anagen was observed by the appearance of darkening skinand by hair regrowth. For the hair cycle experiments described inFig. 1A, biopsies were taken from two mice per time point, and the ex-periment was replicated on three sets of littermates. Data presented inthe graph in Fig. 1B were generated by observing skin darkening overtime after treatment, as shown in fig. S1E. For the experiments describedin fig. S1 (C and D), three to four mice were used per group (7-weekversus 8.5-week), and the experiment was replicated three times. Forthe experiment described in Fig. 1B and fig. S1E, four mice were treatedwith control (half of the back skin) and ruxolitinib (half of the back skin)and four mice were treated with control (half of the back skin) andtofacitinib (half of the back skin). The experiment presented in Fig. 1E
8 of 12

R E S EARCH ART I C L E
on April 25, 2020
http://advances.sciencemag.org/
Dow
nloaded from
was independently replicated once, with three mice per condition. Inthe experiments described in Fig. 2A, three mice per genotype wereused, and the experiment was replicated in two sets of littermates.
In Fig. 1 (C and D), gene expression profiling was performed onwhole-skin biopsies from nine B6/C57 female mice (8.5 weeks of age).Biopsies were taken from dorsal skin at day 0 (T0) of the experiment.Mice were treated daily with dimethyl sulfoxide (DMSO), ruxolitinib,and tofacitinib on days 1 to 4, at which point a second biopsy was takenfrom treatedmice at day 5 (T5). Quality control was performed using theaffy analysisQC package from http://arrayanalysis.org/. Two samples,DMSO1 T5 and RUXO3 T5, were removed from further downstreamanalysis because they failed quality control. Data were normalized usingthe GCRMA (GC Robust Multi-array Average) method as implemen-ted in affy analysisQC.
For Fig. 2B, skin was harvested from 3mice per time point, and fourtime points were tested (12 individual mice were used for the analysis).For the humanorgan culture assay in Fig. 3B, follicleswere derived froma single donor, and three to four follicles were used per condition. Theexperiment was replicated twice with follicles derived from two addi-tional donors. For the DP sphere microarray analysis in Fig. 4A, cellswere prepared from three different donors, with vehicle, ruxolitinib, andtofacitinib treatments applied to cells from each individual.
MiceAll wild-type mice in this study (adult and neonatal) are of C57/B6background, either bred in our laboratory or purchased from theJackson Laboratory. ICR-SCIDmice (IcrTac:ICR-Prkdcscid) for graftingexperiments were purchased from Taconic. Athymic nude mice forthe hair patch assay were purchased from Charles River. Rag1−/−
and Tcr b/d−/− mice were purchased from the Jackson Laboratory(stock nos. 002216 and 002122, respectively). All animals were main-tained at the AAALAC (Accreditation of Laboratory Animal Care In-ternational) Institute for Comparative Medicine at Columbia University.Procedures were performed using Institutional Animal Care and UseCommittee–approved protocols.
Human specimensScalp skin for grafting experiments were obtained from AdvancedBioscience Resources Inc. Occipital scalp follicles were from discardedtissue obtained during hair transplantation surgery, in accordance withthe Declaration of Helsinki, after we received an exemption under 45CFR (Code of Federal Regulations) 46 by the Institutional ReviewBoardexemption at Columbia University.
Pharmacological inhibitors of JAK-STAT and other drugsused in this studyRuxolitinib was purchased fromChemieTek (catalog no. CT-INCB).Tofacitinib was purchased from AbMole BioScience (catalog no.477600-75-2). Hedgehog agonist (SAG) was purchased from EMDMillipore (catalog no. 566660). JAK-STAT inhibitors were dissolvedin DMSO and used at 2 to 3% for in vivo experiments, as indicated,and 400 nM for in vitro experiments. SAG was used at 120 mM, asdescribed by Paladini et al. (2).
Antibodies and immunofluorescenceImmunofluorescence on fresh-frozen sections of mouse skin was per-formed as previously described (1). All fluorescence images were takenwith the Zeiss LSR Excited confocal microscope. All bright-field images
Harel et al. Sci. Adv. 2015;1:e1500973 23 October 2015
were taken with Zeiss Axioplan 2 system. Primary antibodies used anddilutions can be found in the Supplementary Materials. Nuclei werestained using DAPI. List of antibodies is available in table S1.
Analysis of differential gene expression by qPCR arrayTotal RNA was isolated from mouse dorsal skin at indicated timepoints by using the RNeasy Mini Kit (Qiagen) in accordance with themanufacturer’s instructions. Total RNA (2 mg) was reverse-transcribedwith oligo(dT) primers and SuperScript III (Invitrogen). Resultingcomplementary DNA (cDNA) from each sample was aliquoted intoa single JAK-STAT signaling qPCR array (Qiagen/SABiosciences cat-alog no. PAMM-039, gene list provided online). The array includes84 genes related to the JAK-STAT pathway, plus five housekeepinggenes and quality controls. Real-time PCR was performed on an ABI7300 (Applied Biosystems). Data analysis was performed using RT2
Profiler PCR Array data analysis software, provided by SABiosciences.Fold change in expression was determined using the DDCt method, andvalues used in downstream analysis were derived by taking themeans offold changes in three biological replicates per time point.
Gene Expression Dynamics InspectorValues for average log 2 fold changewere calculated as−DDCt relative topostnatal day 17 (early telogen) and were used to perform GEDI anal-ysis to visualize how “metagenes” identified with a self-organizing mapalgorithm vary across samples. Metagenes are clusters of genes that showsimilar temporal expression patterns across samples (66) and that areassigned to a single pixel in a 2D grid. Neighboring pixels demonstratesimilar expression patterns to one another. The self-organizing mapswere then rendered using the level plot function in the lattice packagein R.
Proliferation experimentsC57/B6 mice at 8.5 weeks of age were treated on half of the dorsalback with either topical vehicle control (left side) or JAK inhibitors(right side). Four hours after treatment, each mouse received a singleinjection of Edu (20mg/kg) (Invitrogen). One hour after injection, skinwas harvested, fixed and stained using the Click-iT EdU Alexa Fluor488 Imaging Kit (Life Technologies) per the manufacturer’s instruc-tions, and costained for P-cadherin. Experiment was replicated once,with one mouse per time point.
Microarray analysisTotal RNA was extracted using the Qiagen RNeasy Micro Kit. TheOvationRNAAmplification kit (Nugen)was used to generate amplifiedcDNA formicroarray analysis. Arrays used were Affymetrixmouse 4302.0, hybridized at the Columbia University Medical Center microarrayfacility. qPCR analysis was performed on the ABI 7300 cycler. Primersequences are available in table S2. LIMMA (Linear Models for Micro-arrayData) was usedwith amultilevelmodel to identify genes that weredifferentially expressed between samples at T5 and T0 for each treat-ment group (DMSO, RUXO, and TOFA) using a threshold of foldchange of 1.5 andP< 0.05. Results from the differential expression anal-ysis were uploaded to IPA to identify the molecular pathways that wereoverrepresented in each of the lists of differentially expressed genes foreach of our treatment groups.
For the DP sphere microarray analysis, total RNA was extractedfromDP spheres cultured in DMSO, ruxolitinib, or tofacitinib from eachof the three donors using the Qiagen RNeasy Micro Kit. The Ovation
9 of 12

R E S EARCH ART I C L E
on April 25, 2020
http://advances.sciencemag.org/
Dow
nloaded from
RNA Amplification kit (Nugen) was used to generate amplified cDNAfor microarray analysis. LIMMA was used with a linear model treatingboth treatment and the blocking factor donor as fixed effects. Contrastsof interest between treatments were used to identify genes that were dif-ferentially expressed between pairs of treatments for the treatmentsDMSO, RUXO, and TOFA. A threshold of absolute fold change of1.5 andP<0.05was used. ForGSEA, theHG_U133_Plus_2.0 chip annota-tion file downloaded from ftp://ftp.broad.mit.edu//pub/gsea/annotations/was used to annotate the Affymetrix probe sets and to remove dupli-cate probe sets for a given gene symbol. Duplicates were removed onthe basis of the maximum absolute fold change observed for eachgene symbol. Preranked gene lists were generated for each treatmentcontrast of interest using themoderated t-statistics returned by LIMMA.GSEA was performed on these lists to identify KEGG (Kyoto Encyclo-pedia of Genes and Genomes) pathways overrepresented in each list(ES, P = 0.01).
Gene set enrichment analysisThe GSEA comparing tofacitinib-treated and control cells was per-formed by generating a list of all detected transcripts ranked by dif-ferential expression (P value) and direction of fold change (up or down).This ranked list was tested for the enrichment of the union of territories2 and 4 (gene list provided in table S3) as previously described (31). Anull distribution for computing the NES and its associated P value wasobtained by label shuffling to randomize the gene rankings. These ran-domized sets were then used to compute null ESs over 5000 iterations togenerate a null distribution. The observed leading-edge ES was normal-ized to this distribution, and a two-tailed P value was generated for thisNES.
Human HF organ culture assayAdult human follicles were microdissected from occipital skin understerile conditions as previously described. Individual follicles wereplaced in a 24-well tissue culture plate in Williams E medium sup-plemented with hydrocortisone, insulin, and glutamine (67) in thepresence of DMSO, ruxolitinib, or tofacitinib (400 nM). Medium wasreplaced every 2 days, and images of individual follicleswere taken every2 days. Analysis of the growth rate of each follicle was performedusing ImageJ.
Patch assay with DP spheresDP cells were grown in spheres as previously described (31). DMSO,ruxolitinib, or tofacitinib was added to the medium (400 nM). Eachhanging drop contained 1000 cells, which aggregated to form DPspheroids after 24 hours. Forty-eight hours after plating, 500 spheresin each condition were collected for use in the patch assay. Each experi-ment was performed with cells derived from a single individual, andfour separate experiments were conducted. Keratinocytes were isolatedfrom newborn mice by following the protocol outlined by Lichti et al.(68). Cells were cultured in defined keratinocyte serum-freemedium for2 to 4 days before being harvested for the patch assay. Newbornmurinekeratinocytes (1 x 106) were then mixed together with 500 human DPspheres in each condition and subdermally injected into dorsal skin of anude mouse. Twelve days after performing the injections, cysts haddeveloped in the dermis, some of which contained HF and hair fibers.These cysts were collected, photographed, and then digested in 0.35%collagenase. The digested slurries were spread onto microscope slides,and hair fiber countsweremanually performed under a stereomicroscope.
Harel et al. Sci. Adv. 2015;1:e1500973 23 October 2015
Human skin grafting assayHuman scalp skin (16 weeks old), about 2 × 2 cm in diameter, wasgrafted onto the back of SCID mice. The mice were bandaged, andthe grafts were left to recover for 6 weeks. After 6 weeks, small hairscould be observed on the grafts. Grafts were treated daily with topicalapplication of vehicle control, ruxolitinib, or tofacitinib. Treatmentcontinued for 4weeks, and pictureswere taken every 3 to 5 days. Experi-ments were performed with skin from three separate donors.
Quantification of hair growth was performed using ImageJ. Takingadvantage of the fact that the donor hairwas dark and the graftedmousewas white, we measured the intensity of pigmentation as a proxy fordensity of HF. Because the control and experimental treatments wereperformed on the same graft, each image allows for direct comparisonbetween the vehicle- and drug-treated skin. Intensity was scored acrossthree lines per image and averaged to generate the histograms shown inFig. 3A and fig. S4A. Darker regions are given a lower value than brightregions. Ratio was calculated by averaging the intensity values for thecontrol-treated side by the intensity values for the drug-treated side,accounting for pretreatment differences in hair growth on the graftand for changes in hair density occurring over time.
Statistical analysisA significance level ofP= 0.05was used for all tests. For both the organculture longitudinal study (Fig. 3, B and C) and the skin pigmentationstudy (Fig. 1B), nonparametric longitudinal data analysis was per-formed using the R package nparLD to test the hypothesis that atreatment-by-time interaction exists, that is, time profiles are not parallel,for comparisons of pairs of treatments. In the organ culture study, anF1-LD-F1 design was used. In the skin pigmentation study, an LD-F2model was performed for the ruxolitinib versus control and tofacitinibversus control comparisons to account for the paired design, and an F1-LD-F1model was used for the tofacitinib versus ruxolitinib comparisonbecause the data were unpaired. The analysis of variance (ANOVA)–type statistic was tested for significance at a = 0.05 (a randomizedcomplete block design was used to analyze the human DP sphere patchassay) (Fig. 3E). Three treatments eachwere applied toDP spheres fromeach of three donors. Donor was treated as a fixed blocking factor. Theunit of statistical analysis is the number of observed HFs. The data wereanalyzed using a linear mixed-effects model treating the blocking factordonor as a random effect. The R packages lme4 and lmerTest were usedto testwhether the fixed factor treatment significantly contributed to thenumber of follicles observed and to perform post hoc comparisons oftreatmentmeans. The package lmerTest used the Satterthwaite approx-imation to obtain P values and denominator degrees of freedom.
SUPPLEMENTARY MATERIALSSupplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/1/9/e1500973/DC1Fig. S1. JAK inhibition–mediated hair growth recapitulates endogenous hair growth and isdependent on the duration of telogen.Fig. S2. Molecular pathways enriched in genes differentially regulated by each JAK-STATinhibitor.Fig. S3. Effects of JAK inhibition are independent of T cells and instead represent cell-intrinsicproperties.Fig. S4. Inhibition of JAK-STAT promotes human HF growth.Table S1. List of antibodies and dilutions used in this study.Table S2. List of qPCR primers used in this study.Table S3. List of genes within territories T2 and T4 [as adapted from Higgins et al. (31)].
10 of 12

R E S EARCH ART I C L E
on April 25, 2020
http://advances.sciencemag.org/
Dow
nloaded from
REFERENCES AND NOTES1. L. Xing, Z. Dai, A. Jabbari, J. E. Cerise, C. A. Higgins, W. Gong, A. de Jong, S. Harel, G. M. DeStefano,
L. Rothman, P. Singh, L. Petukhova, J. Mackay-Wiggan, A. M. Christiano, R. Clynes, Alopecia areatais driven by cytotoxic T lymphocytes and is reversed by JAK inhibition. Nat. Med. 20, 1043–1049(2014).
2. R. D. Paladini, J. Saleh, C. Qian, G.-X. Xu, L. L. Rubin, Modulation of hair growth with small mol-ecule agonists of the hedgehog signaling pathway. J. Invest. Dermatol. 125, 638–646 (2005).
3. M. Cutolo, The kinase inhibitor tofacitinib in patients with rheumatoid arthritis: Latest findingsand clinical potential. Ther. Adv. Musculoskelet. Dis. 5, 3–11 (2013).
4. T Zhou, S. Georgeon, R. Moser, D. J. Moore, A. Caflisch, O. Hantschel, Specificity and mechanism-of-action of the JAK2 tyrosine kinase inhibitors ruxolitinib and SAR302503 (TG101348).Leukemia 28, 404–407 (2014).
5. R. A. Mesa, U. Yasothan, P. Kirkpatrick, Ruxolitinib. Nat. Rev. Drug Discov. 11, 103–104 (2012).
6. D. C. Borie, J. J. O’Shea, P. S. Changelian, JAK3 inhibition, a viable new modality of immu-nosuppression for solid organ transplants. Trends Mol. Med. 10, 532–541 (2004).
7. S. Verstovsek, R. A. Mesa, J. Gotlib, R. S. Levy, V. Gupta, J. F. DiPersio, J. V. Catalano, M. Deininger,C. Miller, R. T. Silver, M. Talpaz, E. F. Winton, J. H. Harvey Jr., M. O. Arcasoy, E. Hexner, R. M. Lyons,R. Paquette, A. Raza, K. Vaddi, S. Erickson-Viitanen, I. L. Koumenis, W. Sun, V. Sandor,H. M. Kantarjian, A double-blind, placebo-controlled trial of ruxolitinib for myelofibrosis.N. Engl. J. Med. 366, 799–807 (2012).
8. S. Müller-Röver, B. Handjiski, C. van der Veen, S. Eichmüller, K. Foitzik, I. A. McKay, K. S. Stenn,R. Paus, A comprehensive guide for the accurate classification of murine hair follicles indistinct hair cycle stages. J. Invest. Dermatol. 117, 3–15 (2001).
9. M. V. Plikus, J. A. Mayer, D. de la Cruz, R. E. Baker, P. K. Maini, R. Maxson, C.-M. Chuong,Cyclic dermal BMP signalling regulates stem cell activation during hair regeneration. Nature451, 340–344 (2008).
10. M. V. Plikus, R. E. Baker, C.-C. Chen, C. Fare, D. de la Cruz, T. Andl, P. K. Maini, S. E. Millar,R. Widelitz, C.-M. Chuong, Self-organizing and stochastic behaviors during the regenerationof hair stem cells. Science 332, 586–589 (2011).
11. L. Alonso, E. Fuchs, The hair cycle. J. Cell Sci. 119, 391–393 (2006).
12. R. Schmidt-Ullrich, D. J. Tobin, D. Lenhard, P. Schneider, R. Paus, C. Scheidereit, NF-kBtransmits Eda A1/EdaR signalling to activate Shh and cyclin D1 expression, and controlspost-initiation hair placode down growth. Development 133, 1045–1057 (2006).
13. I. Brownell, E. Guevara, C. B. Bai, C. A. Loomis, A. L. Joyner, Nerve-derived Sonic hedgehogdefines a niche for hair follicle stem cells capable of becoming epidermal stem cells. Cell StemCell 8, 552–565 (2011).
14. W.-M. Woo, H. H. Zhen, A. E. Oro, Shh maintains dermal papilla identity and hair morpho-genesis via a Noggin–Shh regulatory loop. Genes Dev. 26, 1235–1246 (2012).
15. Y. Ito, T. S. Hamazaki, K. Ohnuma, K. Tamaki, M. Asashima, H. Okochi, Isolation of murinehair-inducing cells using the cell surface marker prominin-1/CD133. J. Invest. Dermatol.127, 1052–1060 (2007).
16. Y.-C. Hsu, L. Li, E. Fuchs, Transit-amplifying cells orchestrate stem cell activity and tissueregeneration. Cell 157, 935–949 (2014).
17. P. Rabbani, M. Takeo, W. Chou, P. Myung, M. Bosenberg, L. Chin, M. M. Taketo, M. Ito,Coordinated activation of Wnt in epithelial and melanocyte stem cells initiates pigmentedhair regeneration. Cell 145, 941–955 (2011).
18. R. M. Castilho, C. H. Squarize, L. A. Chodosh, B. O. Williams, J. S. Gutkind, mTOR mediatesWnt-induced epidermal stem cell exhaustion and aging. Cell Stem Cell 5, 279–289(2009).
19. A. J. Kellenberger, M. Tauchi, Mammalian target of rapamycin complex 1 (mTORC1) maymodulate the timing of anagen entry in mouse hair follicles. Exp. Dermatol. 22, 77–80(2013).
20. R. Schmidt-Ullrich, T. Aebischer, J. Hülsken, W. Birchmeier, U. Klemm, C. Scheidereit, Re-quirement of NF-kB/Rel for the development of hair follicles and other epidermalappendices. Development 128, 3843–3853 (2001).
21. S. Sano, S. Itami, K. Takeda, M. Tarutani, Y. Yamaguchi, H. Miura, K. Yoshikawa, S. Akira,J. Takeda, Keratinocyte‐specific ablation of Stat3 exhibits impaired skin remodeling, butdoes not affect skin morphogenesis. EMBO J. 18, 4657–4668 (1999).
22. S. Sano, M. Kira, S. Takagi, K. Yoshikawa, J. Takeda, S. Itami, Two distinct signaling pathwaysin hair cycle induction: Stat3-dependent and -independent pathways. Proc. Natl. Acad. Sci.U.S.A. 97, 13824–13829 (2000).
23. V. Greco, T. Chen, M. Rendl, M. Schober, H. A. Pasolli, N. Stokes, J. dela Cruz-Racelis, E. Fuchs,A two-step mechanism for stem cell activation during hair regeneration. Cell Stem Cell 4,155–169 (2009).
24. A. Ferrajoli, S. Faderl, F. Ravandi, Z. Estrov, The JAK-STAT pathway: A therapeutic target inhematological malignancies. Curr. Cancer Drug Targets 6, 671–679 (2006).
25. D. Gay, O. Kwon, Z. Zhang,M. Spata, M. V. Plikus, P. D. Holler, M. Ito, Z. Yang, E. Treffeisen, C. D. Kim,A. Nace, X. Zhang, S. Baratono, F.Wang,D.M.Ornitz, S. E.Millar, G. Cotsarelis, Fgf9 fromdermal gdT cells induces hair follicle neogenesis after wounding. Nat. Med. 19, 916–923 (2013).
Harel et al. Sci. Adv. 2015;1:e1500973 23 October 2015
26. J. E. Kloepper, K. Kawai, M. Bertolini, T. Kanekura, R. Paus, Loss of gd T cells results in haircycling defects. J. Invest. Dermatol. 133, 1666–1669 (2013).
27. D. Castellana, R. Paus, M. Perez-Moreno, Macrophages contribute to the cyclic activation ofadult hair follicle stem cells. PLOS Biol. 12, e1002002 (2014).
28. A. Heine, S. A. E. Held, S. Nora Daecke, S. Wallner, S. Parampalli Yajnanarayana, C. Kurts, D. Wolf,P. Brossart, The JAK-inhibitor ruxolitinib impairs dendritic cell function in vitro and in vivo.Blood 122, 1192–1202 (2013).
29. K. K. Lin, D. Chudova, G. W. Hatfield, P. Smyth, B. Andersen, Identification of hair cycle-associated genes from time-course gene expression profile data by using replicate vari-ance. Proc. Natl. Acad. Sci. U.S.A. 101, 15955–15960 (2004).
30. K. S. Stenn, R. Paus, Control of hair follicle cycling. Physiol. Rev. 81, 449–494 (2001).31. C. A. Higgins, J. C. Chen, J. E. Cerise, C. A. Jahoda, A. M. Christiano, Microenvironmental repro-
gramming by three-dimensional culture enables dermal papilla cells to induce de novo hu-man hair-follicle growth. Proc. Natl. Acad. Sci. U.S.A. 110, 19679–19688 (2013).
32. Y. Zheng, X. Du, W. Wang, M. Boucher, S. Parimoo, K. Stenn, Organogenesis fromdissociated cells: Generation of mature cycling hair follicles from skin-derived cells. J. Invest.Dermatol. 124, 867–876 (2005).
33. R. R. Driskell, A. Giangreco, K. B. Jensen, K. W. Mulder, F. M. Watt, Sox2-positive dermal papillacells specify hair follicle type in mammalian epidermis. Development 136, 2815–2823 (2009).
34. M. Kawano, A. Komi-Kuramochi, M. Asada, M. Suzuki, J. Oki, J. Jiang, T. Imamura, Comprehen-sive analysis of FGF and FGFR expression in skin: FGF18 is highly expressed in hair follicles andcapable of inducing anagen from telogen stage hair follicles. J. Invest. Dermatol. 124, 877–885(2005).
35. H. Kamp, C. C. Geilen, C. Sommer, U. Blume-Peytavi, Regulation of PDGF and PDGF receptorin cultured dermal papilla cells and follicular keratinocytes of the human hair follicle. Exp.Dermatol. 12, 662–672 (2003).
36. M. Yu, S. Kissling, P. Freyschmidt-Paul, R. Hoffmann, J. Shapiro, Interleukin-6 cytokine familymember oncostatin M is a hair-follicle-expressed factor with hair growth inhibitory properties.K. J. McElwee, Exp. Dermatol. 17, 12–19 (2008).
37. M. Rendl, L. Lewis, E. Fuchs, Molecular dissection of mesenchymal–epithelial interactions inthe hair follicle. PLOS Biol. 3, e331 (2005).
38. M. Rendl, L. Polak, E. Fuchs, BMP signaling in dermal papilla cells is required for their hairfollicle-inductive properties. Genes Dev. 22, 543–557 (2008).
39. R. Paus, K. Foitzik, P. Welker, S. Bulfone-Paus, S. Eichmüller, Transforming growth factor-breceptor type I and type II expression during murine hair follicle development and cycling.J. Invest. Dermatol. 109, 518–526 (1997).
40. K. Foitzik, R. Paus, T. Doetschman, G. P. Dotto, The TGF-b2 isoform is both a requiredand sufficient inducer of murine hair follicle morphogenesis. Dev. Biol. 212, 278–289(1999).
41. N. Oshimori, E. Fuchs, Paracrine TGF-b signaling counterbalances BMP-mediated repression inhair follicle stem cell activation. Cell Stem Cell 10, 63–75 (2012).
42. P. Zhou, C. Byrne, J. Jacobs, E. Fuchs, Lymphoid enhancer factor 1 directs hair folliclepatterning and epithelial cell fate. Genes Dev. 9, 700–713 (1995).
43. K. Kratochwil, M. Dull, I. Farinas, J. Galceran, R. Grosschedl, Lef1 expression is activated byBMP-4 and regulates inductive tissue interactions in tooth and hair development. GenesDev. 10, 1382–1394 (1996).
44. B. Hu, K. Lefort, W. Qui, B.-C. Nguyen, R. D. Rajaram, E. Castillo, F. He, Y. Chen, P. Angel, C. Brisken,G. P. Dotto, Control of hair follicle cell fate by underlying mesenchyme through a CSL–Wnt5a–FoxN1 regulatory axis. Genes Dev. 24, 1519–1532 (2010).
45. C. A. Ambler, F. M. Watt, Expression of Notch pathway genes in mammalian epidermis andmodulation by b-catenin. Dev. Dyn. 236, 1595–1601 (2007).
46. A. Subramanian, P. Tamayo, V. K. Mootha, S. Mukherjee, B. L. Ebert, M. A. Gillette, A. Paulovich,S. L. Pomeroy, T. R. Golub, E. S. Lander, J. P. Mesirov, Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. U.S.A.102, 15545–15550 (2005).
47. Z. Wang, G. Li, W. Tse, K. D. Bunting, Conditional deletion of STAT5 in adult mouse hema-topoietic stem cells causes loss of quiescence and permits efficient nonablative stem cellreplacement. Blood 113, 4856–4865 (2009).
48. F. D. Price, J. von Maltzahn, C. F. Bentzinger, N. A. Dumont, H. Yin, N. C. Chang, D. H. Wilson,J. Frenette, M. A. Rudnicki, Inhibition of JAK-STAT signaling stimulates adult satellite cellfunction. Nat. Med. 20, 1174–1181 (2014).
49. M. T. Tierney, T. Aydogdu, D. Sala, B. Malecova, S. Gatto, P. L. Puri, L. Latella, A. Sacco, STAT3signaling controls satellite cell expansion and skeletal muscle repair. Nat. Med. 20, 1182–1186(2014).
50. J. Doles, M. Storer, L. Cozzuto, G. Roma, W. M. Keyes, Age-associated inflammation inhibitsepidermal stem cell function. Genes Dev. 26, 2144–2153 (2012).
51. J. Goldstein, S. Fletcher, E. Roth, C. Wu, A. Chun, V. Horsley, Calcineurin/Nfatc1 signalinglinks skin stem cell quiescence to hormonal signaling during pregnancy and lactation.Genes Dev. 28, 983–994 (2014).
52. S. J. Divito, T. S. Kupper, Inhibiting Janus kinases to treat alopecia areata. Nat. Med. 20,989–990 (2014).
11 of 12

R E S EARCH ART I C L E
http://advances.scienD
ownloaded from
53. E. Festa, J. Fretz, R. Berry, B. Schmidt, M. Rodeheffer, M. Horowitz, V. Horsley, Adipocyte lineagecells contribute to the skin stem cell niche to drive hair cycling. Cell 146, 761–771 2011).
54. C. A. B. Jahoda, A. M. Christiano, Niche crosstalk: Intercellular signals at the hair follicle. Cell146, 678–681 (2011).
55. K. Turksen, T. Kupper, L. Degenstein, I. Williams, E. Fuchs, Interleukin 6: Insights to its functionin skin by overexpression in transgenic mice. Proc. Natl. Acad. Sci. U.S.A. 89, 5068–5072 (1992).
56. M. H. Kwack, J. S. Ahn, M. K. Kim, J. C. Kim, Y. K. Sung, Dihydrotestosterone-inducible IL-6inhibits elongation of human hair shafts by suppressing matrix cell proliferation and pro-motes regression of hair follicles in mice. J. Invest. Dermatol. 132, 43–49 (2012).
57. A. Quintás-Cardama, K. Vaddi, P. Liu, T. Manshouri, J. Li, P. A. Scherle, E. Caulder, X. Wen, Y. Li,P. Waeltz, M. Rupar, T. Burn, Y. Lo, J. Kelley, M. Covington, S. Shepard, J. D. Rodgers, P. Haley,H. Kantarjian, J. S. Fridman, S. Verstovsek, Preclinical characterization of the selective JAK1/2inhibitor INCB018424: Therapeutic implications for the treatment of myeloproliferative neo-plasms. Blood 115, 3109–3117 (2010).
58. H. L. Geyer, R. A. Mesa, Therapy for myeloproliferative neoplasms: When, which agent, andhow? Blood 124, 3529–3537 (2014).
59. K. Ghoreschi, M. I. Jesson, X. LI, J. L. Lee, S. Ghosh, J. W. Alsup, J. D. Warner, M. Tanaka,S. M. Steward-Tharp, M. Gadina, C. J. Thomas, J. C. Minnerly, C. E. Storer, T. P. LaBranche,Z. A. Radi, M. E. Dowty, R. D. Head, D. M.Meyer, N. Kishore, J. J. O’Shea, Modulation of innate andadaptive immune responses by tofacitinib (CP-690,550). J. Immunol. 186, 4234–4243 (2011).
60. H. Bachelez, P. C. van de Kerkhof, R. Strohal, A. Kubanov, F. Valenzuela, J. H. Lee, V. Yakusevich,S Chimenti, J. Papacharalambous, J. Proulx, P. Gupta, H. Tan, M. Tawadrous, H. Valdez, R. Wolk,Tofacitinib versus etanercept or placebo in moderate-to-severe chronic plaque psoriasis: Aphase 3 randomised non-inferiority trial. Lancet 386, 552–561 (2015).
61. N. Punwani, T. Burn, P. Scherle, R. Flores, J. Shi, P. Collier, D. Hertel, P. Haley, Y. Lo, P. Waeltz,J. Rodgers, S. Shepard, K. Vaddi, S. Yeleswaram, R. Levy, W. Williams, A. B. Gottlieb, Down-regulation of key inflammatory cell markers with topical Janus kinase 1/2 inhibitor. Br. J. Dermatol.,10.1111/bjd.13994 (2015).
62. L. L. Levy, J. Urban, B. A. King, Treatment of reclacitrant atopic dermatitis with the oralJanus kinase inhibitor tofacitinib citrate. J. Am. Acad. Dermatol. 73, 395–399 (2015).
63. B. G. Craiglow, B. A. King, Tofacitinib citrate for the treatment of vitiligo: A pathogenesis-directed therapy. JAMA Dermatol. 10.1001/jamadermatol.2015.1520 (2015).
64. B. G. Craiglow, B. A. King, Killing two birds with one stone: Oral tofacitinib reverses alopeciauniversalis in a patient with plaque psoriasis. J. Invest. Dermatol. 134, 2988–2990 (2014).
65. A. Jabbari, Z. Dai, L. Xing, J. E. Cerise, Y. Ramot, Y. Berkun, G. A. Sanchez, R. Goldbach-Mansky,A. M. Christiano, R. Clynes, A. Zlotogorski, Reversal of alopecia areata following treatment withthe JAK1/2 inhibitor baricitinib. EBioMedicine 4, 351–355 (2015).
Harel et al. Sci. Adv. 2015;1:e1500973 23 October 2015
66. G. S. Eichler, S. Huang, D. E. Ingber, Gene Expression Dynamics Inspector (GEDI): For integrativeanalysis of expression profiles. Bioinformatics 19, 2321–2322 (2003).
67. D. J. Tobin, Ex Vivo Organ Culture of Human Hair Follicles: A Model Epithelial–Neuroectodermal–Mesenchymal Interaction System (Methods in Molecular Biology Series, Humana Press, NewYork, 2011), vol. 695, pp. 213–227.
68. U. Lichti, J. Anders, S. H. Yuspa, Isolation and short-term culture of primary keratinocytes,hair follicle populations and dermal cells from newborn mice and keratinocytes from adultmice for in vitro analysis and for grafting to immunodeficient mice. Nat. Protoc. 3, 799–810(2008).
Acknowledgments: We thank D. Owens for advice and technical assistance, A. De Jong andE. Wang for their valuable comments on the manuscript, and E. Chang and M. Zhang fortheir help with animal experiments. Funding: This work was supported in part by U.S. PublicHealth Service NIH National Institute of Arthritis and Musculoskeletal and Skin Diseasesgrants R01AR056016 (to A.M.C.), R21AR061881 (to A.M.C. and R.C.), and P30AR044535 (theColumbia University Skin Disease Research Center), as well as the Locks of Love Foundationand the Alopecia Areata Initiative. S.H. was funded with the Dermatology Foundation CareerDevelopment Award. J.E.C. and J.C.C. were supported by the Kirschstein National ResearchService Award postdoctoral training grant in Medical Genetics [T32GM082271 (to A.M.C.)].Author contributions: S.H., C.A.H., Z.D., R.C., and A.M.C. contributed to the conception anddesign of the study. S.H., C.A.H., J.E.C., J.C.C., and Z.D. contributed to data collection, analysis,and interpretation. S.H. and A.M.C. drafted the report. All authors approved the final versionof the report. Competing interests: Columbia University has filed patent applications relat-ing to the discoveries reported in this paper, which are being commercialized through VixenPharmaceuticals Inc., of which A.M.C. is a founder. The other authors declare that they haveno competing interests. Data and materials availability: Microarray data were deposited inGene Expression Omnibus and will be available upon publication of the article.
Submitted 22 July 2015Accepted 30 July 2015Published 23 October 201510.1126/sciadv.1500973
Citation: S. Harel, C. A. Higgins, J. E. Cerise, Z. Dai, J. C. Chen, R. Clynes, A. M. Christiano,Pharmacologic inhibition of JAK-STAT signaling promotes hair growth. Sci. Adv. 1, e1500973(2015).
cem
12 of 12
on April 25, 2020
ag.org/

Pharmacologic inhibition of JAK-STAT signaling promotes hair growthSivan Harel, Claire A. Higgins, Jane E. Cerise, Zhenpeng Dai, James C. Chen, Raphael Clynes and Angela M. Christiano
DOI: 10.1126/sciadv.1500973 (9), e1500973.1Sci Adv
ARTICLE TOOLS http://advances.sciencemag.org/content/1/9/e1500973
MATERIALSSUPPLEMENTARY http://advances.sciencemag.org/content/suppl/2015/10/20/1.9.e1500973.DC1
REFERENCES
http://advances.sciencemag.org/content/1/9/e1500973#BIBLThis article cites 65 articles, 23 of which you can access for free
PERMISSIONS http://www.sciencemag.org/help/reprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAAS.Science AdvancesYork Avenue NW, Washington, DC 20005. The title (ISSN 2375-2548) is published by the American Association for the Advancement of Science, 1200 NewScience Advances
Copyright © 2015, The Authors
on April 25, 2020
http://advances.sciencemag.org/
Dow
nloaded from