pharmacokinetics and pharmacodynamics of single-chain recombinant human follicle-stimulating hormone...
TRANSCRIPT

Pharmacokinetics and pharmacodynamicsof single-chain recombinant human follicle-stimulating hormone containing the humanchorionic gonadotropin carboxyterminalpeptide in the rhesus monkey
Jeffrey Klein, M.D.,a Leslie Lobel, M.D., Ph.D,a,b Susan Pollak, B.A.,a
Michel Ferin, M.D.,a,b Ennian Xiao, M.D.,a Mark Sauer, M.D.,a,b andJoyce W. Lustbader, Ph.D.a,b
Columbia University, College of Physicians and Surgeons, New York, New York
Objective: To compare the pharmacokinetics of a long-acting FSH analog containing the hCG-� carboxy-terminal peptide (recombinant hFSH–CTP) with native recombinant hFSH and describe the pharmacodynam-ics of recombinant hFSH–CTP after SC injection in female rhesus monkeys.
Design: Rhesus monkey study.
Setting: Academic research environment.
Animal(s): Ten female rhesus monkeys.
Intervention(s): Recombinant hFSH and recombinant hFSH–CTP were administered via a single SC or IVdose to rhesus monkeys, and serial phlebotomy was performed (n� 2 and n� 4 for SC recombinant hFSHand recombinant hFSH–CTP, respectively; for IV dosing, n� 1 in each group). An additional two monkeyswere pretreated with SC ganirelix and received SC recombinant hFSH–CTP after confirmation of pituitarysuppression.
Main Outcome Measure(s): Plasma disappearance rate of recombinant hFSH and recombinant hFSH–CTPand serum estradiol levels.
Result(s): The elimination half-life of recombinant hFSH–CTP was twofold and fourfold longer than that forrecombinant hFSH after SC and IV dosing, respectively. The absorption half-life was approximately threefoldlonger for recombinant hFSH–CTP than for recombinant hFSH after SC administration. RecombinanthFSH–CTP stimulates estradiol secretion for 5–7 days after an isolated SC dose.
Conclusion(s): Addition of the hCG-� carboxyterminal peptide to hFSH-� results in an FSH analog withlonger absorption and elimination half-lives compared with native hormone. This analog is capable ofprolonged ovarian stimulation in rhesus monkeys after an isolated SC injection. (Fertil Steril� 2002;77:1248–55. ©2002 by American Society for Reproductive Medicine.)
Key Words: Human follicle stimulating hormone, controlled ovarian hyperstimulation, carboxyterminalpeptide, FSH analog, glycoprotein hormones
Human FSH is a pituitary glycoprotein hor-mone responsible for ovarian follicular devel-opment in women and induction of spermato-genesis in men. It is a heterodimer comprisedof two dissimilar, noncovalently bound sub-units, � and �. The �-subunit is common toother pituitary glycoprotein hormones, LH andTSH, and to placenta-derived hCG, whereasthe �-subunit is unique and confers biologicspecificity.
Pharmacologic formulations include puri-fied urinary derivatives and, more recently, re-combinant hFSH. Because of its relativelyshort half-life (24 hours after repeated SC dos-ing) (1), hFSH must be administered as a dailyintramuscular or subcutaneous injection, oftenfor 8 days to 12 days when used for ovulationinduction. These regimens of controlled ovar-ian hyperstimulation are associated with anumber of side effects, including local irrita-
Received September 17,2001; revised andaccepted December 6,2001.Supported by a grant fromthe National Institute ofDiabetes/Digestive/KidneyDiseases, NIDDK-51266,Bethesda, Maryland 20892.Presented at the 48thAnnual Meeting of theSociety for GynecologicInvestigation, Toronto,Ontario, Canada, March14–17, 2001.Reprint requests: Joyce W.Lustbader, Ph.D.,Department of Obstetricsand Gynecology, ColumbiaUniversity, 630 W 168thStreet, New York, NewYork 10032 (FAX: 212-305-5140; E-mail: [email protected]).a Department of Obstetricsand Gynecology, ColumbiaUniversity.b Center for ReproductiveSciences, ColumbiaUniversity.
REPRODUCTIVE BIOLOGY FERTILITY AND STERILITY �VOL. 77, NO. 6, JUNE 2002
Copyright ©2002 American Society for Reproductive MedicinePublished by Elsevier Science Inc.
Printed on acid-free paper in U.S.A.
0015-0282/02/$22.00PII S0015-0282(02)03113-8
1248

tion and discomfort, which may result in poor compliance.Development of a long-acting FSH formulation requiringless frequent administration should make this treatment moretolerable.
Among the glycoprotein hormones, hCG is known tohave the longest circulating half-life. This has been attrib-uted to the four O-linked glycosylation sites present on thecarboxyterminal peptide (CTP) sequence on the �-subunit ofhCG (2). Human chorionic gonadotropin shares a commonreceptor with hLH, and both hormones elicit similar biologicactivity after receptor binding. Relative to hLH, however, theserum half-life of hCG is more than twofold greater (31hours as compared with 11 hours) (3, 4). Except for the CTPon the �-subunit of hCG (amino acids 113–145), the twohormones share �85% sequence homology (through the first113 amino acids) (5). The CTP thus appears to alter themetabolism of hCG without interfering with receptor bind-ing and signal transduction.
A long-acting FSH agonist developed by fusing the CTPfrom the �-subunit of hCG to the carboxyterminus of�-hFSH has been reported (6). The intact fusion protein hadsimilar in vitro bioactivity compared with native hFSH andincreased bioactivity in vivo in rats as assessed by ovarianweight gain (6). This latter observation was attributed toslower clearance of the molecule, as increased serum levelswere observed 24 hours after IV injection in rats comparedwith control animals receiving native hFSH (6). Pharmaco-kinetic parameter estimates, however, were not describeduntil recently, when this FSH analog was administered to 13hypogonadotropic men and shown to have an eliminationhalf-life two to three times longer than native recombinantFSH after SC injection (7). In vivo bioactivity and pharma-cokinetics in females, however, have not been described.
We expressed an alternative construct, a single-chainfusion protein containing hFSH�, CTP, and the common�-subunit in a Chinese hamster ovary cell line and examinedthe pharmacokinetics and pharmacodynamics of this productafter IV and SC administration to female rhesus monkeys.The in vivo bioactivity of the molecule was investigated innormally cycling, pituitary-suppressed animals.
MATERIALS AND METHODS
Approval was obtained from the Institutional AnimalCare and Use Committee of Columbia University in accor-dance with the National Institutes of Health guide for thecare and use of laboratory animals. Female rhesus monkeys(macaca mulatta) weighing 4 kg to 8 kg were used in thisstudy.
Bacterial StrainsAll molecular biology techniques and large-scale prepa-
ration of plasmid DNA were performed with Escherichiacoli strain DH5�. Clones were grown in standard Luria–
Bertani medium for purification of recombinant DNA con-structs.
Molecular BiologyAll enzymes for recombinant DNA were purchased from
New England Biolabs (Beverly, MA). The Columbia Uni-versity Core Laboratory synthesized DNA primers for poly-merase chain reaction (PCR) CDNAs for �-hFSH, �-hCG,and the common �-subunit were generously obtained fromDr. William Moyle (Robert Wood Johnson Medical School,Piscataway, NJ). Polymerase chain reactions were per-formed with Vent DNA polymerase (New England Biolabs),and all products of the reactions were sequenced to ensurethat no mutations were introduced during the amplificationprocess. All primer sequences are available upon request.Ligation reactions were transformed into DH5�, and DNAclones were grown in DH5� for large-scale plasmid prepa-ration. Transformation of DH5� was performed according tostandard techniques with calcium chloride.
Construction of a �-hFSH–CTP-� FusionClone
A 5� primer introduced a XhoI site in the same frame andadjacent to the 5� ATG of the �-hFSH complementary DNAsequence, whereas the 3� primer introduced an in-frameBamHI site adjacent to the codon for the last residue of themature �-FSH, which eliminated the terminator codon. In asimilar fashion, a complementary DNA encoding the car-boxy terminal peptide of hCG (residues 113–145 of the hCG�-subunit sequence) was amplified with an in-frame BamHIsite adjacent to the codon for residue 113 and an in-frameXbaI site adjacent to the codon for residue 145. These twofragments were ligated to form a contiguous XhoI–BamHI–XbaI �-hFSH–CTP fusion without a terminator codon at the3� end. This fusion was then ligated to a cDNA encoding themature (lacking the amino-terminal signal peptide) �-sub-unit, including the terminator codon, flanked by in-frame 5�SpeI and 3� SacI sites. The final construct encodes a fusionof the �-hFSH and �-subunit with the hCG �-subunit CTPsequence as the linker sequence (Fig. 1). This final fusionsequence was then inserted into an SV40 expression vector.
Expression and Purification of r-hFSH-CTPAn SV40 expression clone containing the fusion construct
was cotransfected into Chinese hamster ovary–K1 cellsalong with an SV2neo clone encoding resistance to G418.The Chinese hamster ovary cell transformation was per-formed using a standard calcium phosphate precipitate tech-nique. Selectable media containing G418 (Gemini Bioprod-ucts, Woodland, CA) were used to isolate colonies that werepooled and maintained in Ham’s F-12 medium containing500 �g/mL G418, 10% fetal bovine serum, 100 units/mLpenicillin, 100 �g/mL streptomycin, and 4-mM glutamine.Pooled colonies were subcloned in 96-well microtiter dishes,and a clone was isolated that secreted 3 pmol/mL hFSH-CTP. To obtain higher yields, the cells were grown in
FERTILITY & STERILITY � 1249

suspension cultures that produced 9–14 pmol/mL. Spinnerbottles were seeded at 105 cells/mL in Chinese hamsterovary S-SFM (Life Technologies, Rockville, MD) contain-ing 400 �g/mL G418. Cultures generally reached a densityof 2 � 106 cells/mL on day 6 or 7, and the cell supernatantwas harvested on day 7 or 8. Phenylmethylsulfonyl fluoridewas added to the supernatant at a concentration of 0.2 mMand then filtered through a 0.2-�m membrane and kept at4°C until additional purification was performed.
Affinity purification of recombinant hFSH–CTP was ac-complished using an A201 (�-subunit–specific) antibodycolumn (8). The column was prepared by coupling purifiedA201 immunoglobulins to CNBr-Sepharose-4B according tothe manufacturer’s instructions (Amersham Pharmacia Bio-tech, Piscataway, NJ) at a concentration of 5 mg of antibodyper milliliter of Sepharose. After applying the cell superna-tant, the column was washed with 50 bed volumes of PBS,followed by 2 bed volumes of distilled water. The recombi-nant hFSH–CTP was eluted with 3 bed volumes to 4 bedvolumes of 1 M acetic acid and immediately dried on aSpeed-Vac concentrator (Savant Instruments, Holbrook,NY).
In Vitro BioactivityReceptor-binding activity of recombinant hFSH–CTP
was performed using Y-1 cells transfected with the FSHreceptor. Y-1 cell cultures were mixed with recombinanthFSH–CTP and native pituitary hFSH (control) at varyingconcentrations; media were assayed for cyclic adenosine3�:5� monophosphate activity as previously described (9).
PharmacokineticsRecombinant hFSH (Follistim; Organon Inc, West Or-
ange, NJ) was used as a control. The Immulite assay (Diag-nostic Products Corporation, Los Angeles, CA) was used tomeasure recombinant hFSH–CTP and recombinant hFSH.This assay was able to detect recombinant hFSH–CTP invitro and in vivo and did not cross-react with rhesus FSH.
Subcutaneous Protocol
Rhesus monkeys were injected SC with recombinantFSH–CTP (n � 4) or recombinant hFSH (Follistim; n � 2)at a dose of 10 IU/kg. All of the monkeys had been previ-ously ovariectomized except for one monkey in each of the
two treatment groups. Serum recombinant hFSH-CTP orrecombinant hFSH was assayed before drug administrationand at the following intervals after injection: 12 hours 16hours, 20 hours, 24 hours, 36 hours, 48 hours, 60 hours, andevery 24 hours thereafter until levels reached baseline (9days for control animals, 19–22 days for treatment animals).
Intravenous Protocol
One rhesus monkey was given an IV bolus of recombi-nant hFSH–CTP (10 IU/kg). A second animal was given anIV bolus of recombinant hFSH at the same dose. Serum wasassayed for recombinant hFSH–CTP and recombinant hFSHbefore drug administration at the following intervals afterinjection: .5 hours, 1 hour, 2 hours, 4 hours, 8 hours, 12hours, 24 hours, 36 hours, 48 hours, 72 hours, 96 hours, 120hours, 144 hours, and 168 hours.
Pharmacokinetic Analysis
Each individual data set was evaluated by the pharmaco-kinetic data analysis program PKAnalyst (Micromath, Inc.,Salt Lake City, UT). For the IV dosing study, the followingbiexponential equation was fitted to the data:
C�t� � Ae�at � Be�bt
where C(t) is the plasma concentration at time t, and A andB are the multiexponential coefficients. Values of a and brepresent the initial-phase disposition rate constant and theterminal-phase disposition rate constant, respectively. PKA-nalyst was used to generate the best-fit critical pharmacoki-netic parameters, including elimination rate constant, half-life of initial (distribution) phase (t1/2a), half-life of terminal(elimination) phase (t1/2b), and total area under the bloodconcentration–time curve (AUC).
For the SC dosing studies, the blood concentration–timedata were represented by the following biexponential equa-tion:
C�t� � A(e�Ket � e�Kat)
where C(t) is the blood concentration at time t, and A, themultiexponential coefficient. Ke and Ka represent the elim-
F I G U R E 1
Schematic of recombinant hFSH–CTP construct with locations of restriction sites.
Klein. r-hFSH-CTP in rhesus monkeys. Fertil Steril 2002.
1250 Klein et al. r-hFSH-CTP in rhesus monkeys Vol. 77, No. 6, June 2002
ination rate constant and absorption rate constant, respec-tively. All parameter estimates were computed by PKAna-lyst. Bioavailability of recombinant hFSH–CTP andrecombinant hFSH were estimated from the ratio of AUC(SC)–AUC (IV), at a constant dose (10 IU/kg).
In Vivo BioactivityGanirelix acetate (250 �g) was administered by SC in-
jection for 10 consecutive days to two normally cyclingrhesus monkeys beginning menstrual cycle day 4. Recom-binant hFSH–CTP was administered as a single SC dose (10IU/kg) on cycle day 6. Venipuncture was performed daily,and serum was assayed for estradiol levels from cycle day 2through cycle day 14. Serum estradiol was measured usingan automated Immulite assay (Diagnostic Products Corpo-ration).
RESULTS
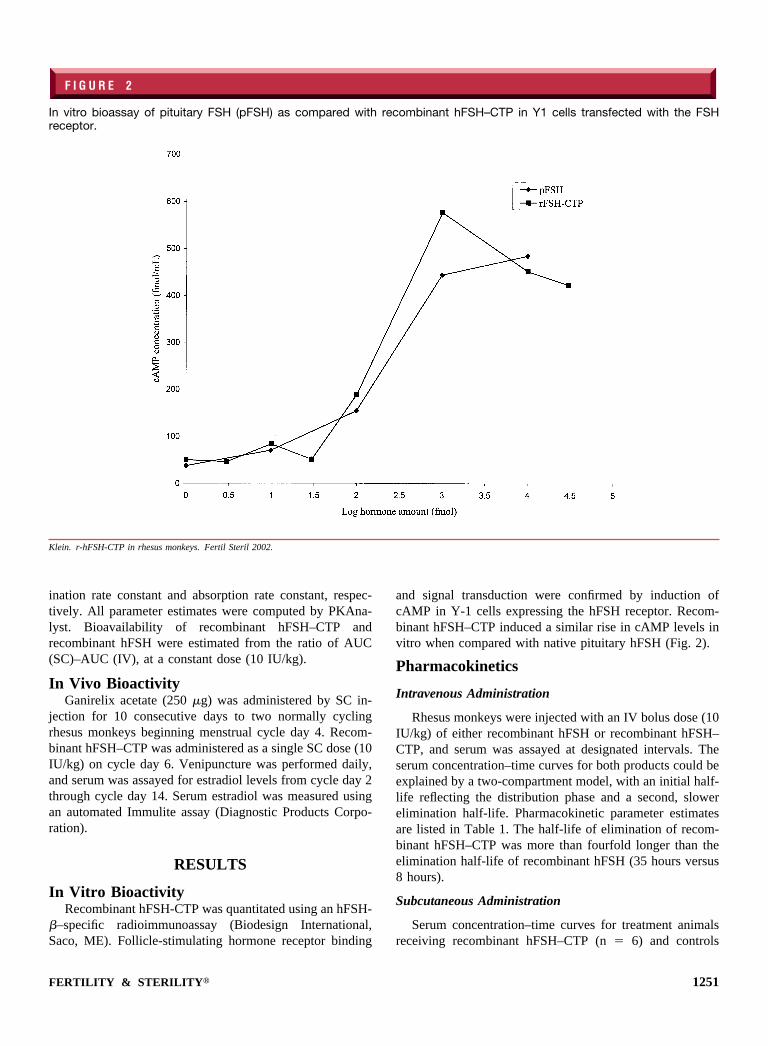
In Vitro BioactivityRecombinant hFSH-CTP was quantitated using an hFSH-
�–specific radioimmunoassay (Biodesign International,Saco, ME). Follicle-stimulating hormone receptor binding
and signal transduction were confirmed by induction ofcAMP in Y-1 cells expressing the hFSH receptor. Recom-binant hFSH–CTP induced a similar rise in cAMP levels invitro when compared with native pituitary hFSH (Fig. 2).
Pharmacokinetics
Intravenous Administration
Rhesus monkeys were injected with an IV bolus dose (10IU/kg) of either recombinant hFSH or recombinant hFSH–CTP, and serum was assayed at designated intervals. Theserum concentration–time curves for both products could beexplained by a two-compartment model, with an initial half-life reflecting the distribution phase and a second, slowerelimination half-life. Pharmacokinetic parameter estimatesare listed in Table 1. The half-life of elimination of recom-binant hFSH–CTP was more than fourfold longer than theelimination half-life of recombinant hFSH (35 hours versus8 hours).
Subcutaneous Administration
Serum concentration–time curves for treatment animalsreceiving recombinant hFSH–CTP (n � 6) and controls
F I G U R E 2
In vitro bioassay of pituitary FSH (pFSH) as compared with recombinant hFSH–CTP in Y1 cells transfected with the FSHreceptor.
Klein. r-hFSH-CTP in rhesus monkeys. Fertil Steril 2002.
FERTILITY & STERILITY� 1251

receiving recombinant hFSH (n � 2) are illustrated in Fig-ures 3 and 4, respectively. Serum levels of recombinanthFSH approached baseline by day 4 after injection, whereaselevated (�2 mIU/mL) levels of recombinant hFSH–CTPwere maintained for approximately 10 days (range, 9.6–10.6days).
The data for both groups could be explained by a one-compartment pharmacokinetic model. Pharmacokinetic pa-rameter estimates for all monkeys are summarized in Table2. The half-life of elimination for both products correlateswell with the IV data and confirms the slower metabolismand clearance of the fusion hormone. The half-life of ab-sorption was approximately threefold longer for recombinanthFSH–CTP compared with recombinant hFSH. Addition ofthe CTP moiety to hFSH thus induced a depot effect, retard-ing the absorption of the product after SC administration.
This explains the slower time to reach peak concentration(tmax) for animals receiving recombinant hFSH–CTP. Bothdrugs were highly bioavailable after SC administration (Ta-ble 2).
In Vivo BioactivityTo establish the bioactivity of recombinant hFSH–CTP in
vivo, two normally cycling monkeys were injected with asingle dose of recombinant hFSH–CTP, and serum wasassayed serially for estradiol. Both monkeys were given aGnRH antagonist (Antagon; Organon) for the duration of thestudy, eliminating any effect on ovarian estrogen productionfrom endogenous rhesus FSH. Estradiol increased in both
T A B L E 1
Pharmacokinetic parameter estimates after IV bolusinjection of 10 IU/kg of recombinant hFSH–CTPrecombinant hFSH.
ParameterRecombinanthFSH–CTP
RecombinanthFSH
T1/2a (distribution) (h) 3.16 1.39T1/2b (elimination) (h) 35.29 8.25AUC (mIU/mL � d) 278 38.8Clearance (1/kg � h) 1.50 10.74
Klein. r-hFSH-CTP in rhesus monkeys. Fertil Steril 2002.
T A B L E 2
Mean pharmacokinetic parameter estimates after SCinjection of recombinant hFSH-CTP and recombinant hFSHat a dose of 10 IU/kg.
Parameter
RecombinanthFSH–CTP
(n � 4)
RecombinanthFSH
(n � 2)
T1/2 (elimination) (h) 35.23 15.74T1/2 (absorption) (h) 5.04 1.75Cmax (mIU/mL) 101.26 25.77Tmax (h) 16.39 5.95AUC (mIU/mL � d) 275.31 30.96Bioavailability (%) 99 80(AUCSC/AUCIV)
Klein. r-hFSH-CTP in rhesus monkeys. Fertil Steril 2002.
F I G U R E 3
Serial recombinant hFSH–CTP levels in four rhesus monkeysafter a single SC injection at a dose of 10 IU/kg. Each symbolis representative of a different monkey.
Klein. r-hFSH-CTP in rhesus monkeys. Fertil Steril 2002.
F I G U R E 4
Serial r-hFSH levels in two rhesus monkeys after a single SCinjection at a dose of 10 IU/kg. Each symbol is representativeof a different monkey.
Klein. r-hFSH-CTP in rhesus monkeys. Fertil Steril 2002.
1252 Klein et al. r-hFSH-CTP in rhesus monkeys Vol. 77, No. 6, June 2002

animals, with peak levels achieved 3 days and 5 days aftergonadotropin injection. One monkey attained supraphysi-ologic levels of estradiol (peak, 773 pg/mL) on day 4 afterrecombinant hFSH–CTP injection, suggesting early recruit-ment of multiple follicles (Fig. 5).
DISCUSSION
We have demonstrated that addition of CTP to the car-boxy terminus of FSH had no adverse impact on folding ofthe molecule, receptor binding, or in vitro signal transduc-tion. The fusion protein was metabolized more slowly thanthe native hormone, as circulating levels remained elevatedfor an extended period. The half-life of elimination after SCadministration was two to three times longer for recombinanthFSH–CTP compared with native recombinant hFSH. Thisdifference corresponds well with the only previous report onpharmacokinetics in human male subjects, in which thehalf-life of elimination after SC administration was pro-longed by a similar magnitude compared with the case ofhistoric controls receiving native hormone.
We were also able to confirm the accuracy of our param-eter estimate for elimination half-life by assessing pharma-cokinetics after IV administration. Surprisingly, however,absorption of recombinant hFSH–CTP was delayed by ap-proximately threefold after SC administration. Absorptionkinetics of recombinant hFSH–CTP have not been reportedpreviously, and it would be interesting to see whether this
phenomenon is reproduced in humans. The long-circulatingpresence of recombinant hFSH–CTP after SC administrationis thus explained by not only an altered metabolism of theproduct but by a depot effect resulting in slower absorption.
An unexpected finding in our study was the observationof extremely high peak concentration (Cmax) and AUC afteradministration of recombinant hFSH–CTP. These parame-ters were four to five times greater than corresponding pa-rameter estimates for recombinant hFSH, suggesting muchhigher bioavailability of the fusion protein. RecombinanthFSH, however, is known to have excellent bioavailability inhumans (1). A comparison of the AUC after SC and IVinjection of recombinant hFSH in our study confirmed thehigh bioavailability of the native recombinant product (80%)in rhesus monkeys. Although bioavailability of recombinanthFSH–CTP was approximately 20% higher (99%), this dif-ference cannot explain the disparity in the AUC (total drugburden) between our fusion hormone and native recombinanthFSH. This must therefore be related to a technical problemin assessing the administered dose of medication or quanti-tating serum levels of hormone.
We specifically chose the same assay system (Immulite)both to calibrate the dose administered and to assess serumlevels after injection to avoid an assay-related disparity be-tween hormone levels in vitro and in vivo. Nonetheless, wesuspect that the Immulite system underestimates the admin-istered dose of recombinant hFSH–CTP without signifi-
F I G U R E 5
Change in estradiol level in two normally cycling monkeys after a single SC injection of recombinant hFSH–CTP. Both monkeyswere given the GnRH antagonist ganirelix acetate for the duration of the study.
Klein. r-hFSH-CTP in rhesus monkeys. Fertil Steril 2002.
FERTILITY & STERILITY� 1253

cantly deviating from the true serum level of the product.Total drug exposure, as assessed by the serum concentra-tion–time curve (i.e., AUC), would thus erroneously exceedtotal drug administered, explaining the discrepancy in ourresults.
To confirm this hypothesis, we compared a sample ofrecombinant hFSH–CTP quantitated by an RIA using adifferent FSH-�–specific antibody (Biodesign International,Saco, ME) with the Immulite assay. As expected, the Im-mulite assay system consistently yielded a result four to fivetimes lower than the Biodesign FSH-�–specific RIA. Anamino acid analysis confirmed that the Immulite assay un-derestimates the actual concentration of the fusion protein(whereas the results of the Biodesign FSH-�–specific RIAcorrelated well with those of the amino acid analysis).
Levels of recombinant-hFSH, in contrast, were consistentbetween assays. The CTP thus appears to partially obscurecertain epitopes on hFSH necessary for recognition by theImmulite assay. Incubation of recombinant hFSH–CTP infetal bovine serum for 6 hours resulted in a greater thantwofold increase in the measured quantity of product by theImmulite assay. This suggests that over time, these epitopesbecome progressively more exposed. This seems to occurrather quickly in vivo, as we assayed a random sample ofcryopreserved rhesus serum (which was available for some,but not all experimental time points) using the BiodesignFSH-�–specific RIA and confirmed that by 12 hours afterSC injection, the Immulite assay becomes accurate (thisrepresents the earliest bleed after SC injection of hormone).We speculate that a conformational change occurs in therecombinant hFSH–CTP molecule in vivo, perhaps inducedby enzymatic cleavage (neuraminidase, for instance, isknown to progressively desialylate native hFSH in vivo andalter its biologic and immunologic activity) (1, 10).
Significantly, a number of pharmacokinetic parameterestimates such as absorption half-life, elimination half-life,and tmax are dose independent. Our results for these param-eter estimates remain valid despite the inability of the Im-mulite assay system to accurately quantitate the level ofrecombinant hFSH–CTP in vitro. Repeating this study usingan assay for recombinant hFSH–CTP that yields consistentin vitro and in vivo measurements would be necessary toobtain an accurate assessment of clearance and volume ofdistribution (which are both related to the dose per AUC). Alarger number of monkeys would be required to repeat thesestudies. Unfortunately we could not repeat these experimentswith our monkey colony, as monkeys develop antibodies tohuman FSH and therefore can only be enrolled in the studyfor a single injection of human hormone.
The pharmacodynamics of recombinant hFSH–CTP in aprimate model are described for the first time. Administra-tion of recombinant hFSH–CTP to two monkeys given aGnRH antagonist (to suppress endogenous FSH activity)elicited a dramatic rise in serum estradiol. A single SC dose
resulted in elevated estradiol levels for 5–7 days, with onemonkey achieving a peak estradiol level greater than threetimes that seen during a normal endogenous rhesus cycle.This supraphysiologic response is indicative of multifollicu-lar recruitment, although sonographic confirmation was notperformed. Such prolonged elevations in estradiol are notnormally seen after isolated SC injections of native recom-binant hFSH (11).
For the first time, we have confirmed the feasibility ofachieving prolonged ovarian stimulation after an isolatedinjection of gonadotropin. Although it is impossible to pre-dict an appropriate dosing schedule based on these prelimi-nary results, it is likely that such a drug will require lessfrequent administration than will native recombinant hFSH.Fewer injections should result in less patient discomfort,improved compliance, and possibly a reduction in the num-ber of local side effects.
Additional data will confirm the safety and efficacy of thisnovel gonadotropin product. Given the potential for ovarianhyperstimulation syndrome with excessive gonadotropinstimulation, initial trials in female human subjects shouldperhaps focus on patients at low risk for ovarian hyperstimulation syndrome (e.g., advanced age, diminished ovar-ian reserve, no history of polycystic ovaries, etc.). Combi-nation therapy using both long and short-acting FSH formu-lations, either together or sequentially during a stimulationcycle, should also be considered. In these cases, the short-acting (native) formulation may be used to fine-tune the FSHdose after an initial bolus of a long-acting analog.
Infertile males with hypogonadotropic hypogonadismtypically require prolonged courses of gonadotropin and mayalso be ideal candidates for future investigation using long-acting FSH agonists. Care must be taken to ensure a highdegree of precision when measuring recombinant hFSH–CTP using hFSH assays, as important epitopes on nativeFSH may be obscured by addition of the carbohydrate-richCTP. Ultimately, a more patient-friendly dosing scheduleand perhaps improved ovarian stimulation make this FSHanalog an exciting product for future investigative work.
Acknowledgments: The authors thank James A. Dias, M.D., New York StateDepartment of Health, Albany, NY, for performing the in vitro bioactivityassays and Wei Zheng, M.D., Columbia University, New York, NY, forperforming the pharmacokinetic analysis and a critical review of this work.The authors also thank Keith Gordon, Ph.D., Organon, Inc., West Orange,NJ, for providing the Follistim and Antagon used in these studies. Theauthors also are grateful to Alinda Barth, B.A., Organon, Inc., West Orange,NJ, for technical support.
References1. le Contonnec JY, Porchet HC, Beltrami V, Khan A, Toon S, Rowland
M. Clinical pharmacology of recombinant human follicle-stimulatinghormone. II. Single doses and steady state pharmacokinetics. FertilSteril 1994;61:679–86.
1254 Klein et al. r-hFSH-CTP in rhesus monkeys Vol. 77, No. 6, June 2002

2. Matzuk MM, Hsueh AJ, Lapolt P, Tsafriri A, Keene JL, Boime I. Thebiological role of the carboxyl-terminal extension of human chorionicgonadotropin beta-subunit. Endocrinology 1990;126:376–83.
3. Porchet HC, LeContonnec JY, Neuteboom B, Canali S, Zanolo G.Pharmacokinetics of recombinant human luteinizing hormone afterintravenous, intramuscular, and subcutaneous administration in mon-keys and comparison with intravenous administration of pituitary hu-man luteinizing hormone. J Clin Endocrinol Metab 1995;80:667–73.
4. Saal W, Glowania HJ, Happ J. Pharmacodynamics and pharmacokinet-ics after subcutaneous and intramuscular injection of human chorionicgonadotropin. Fertil Steril 1991;56:225–8.
5. Pierce JG, Parsons TF. Glycoprotein hormones: structure and function.Annu Rev Biochem 1981;50:465–95.
6. Fares FA, Suganuma N, Nishimori K, Lapolt PS, Hsueh AJW, BoimeI. Design of a long-acting follitropin agonist by fusing the C-terminalsequence of the chorionic gonadotropin beta subunit to the follitropinbeta subunit. Proc Natl Acad Sci USA 1992;89:4304–8.
7. Bouloux PMG, Handelsman DJ, Jockenhovel F, Nieschlag E, Rabi-novici J, Frasa WLH, et al. First human exposure to FSH-CTP inhypogonadotropic hypogonadal males. Hum Reprod 2001;16:1592–7.
8. Krichevsky A, Birken S, O’Connor JF, Bikel K, Schlatterer JP, CanfieldRE. The development of a panel of monoclonal antibodies to humanluteinizing hormone and its application to immunological mapping andtwo-site assays. Endocrine 1994;2:511–20.
9. Lindau-Shepard BA, Brumberg HA, Peterson AJ, Dias JA. Reversibleimmunoneutralization of human follitropin receptor. J Reprod Immunol2001;49:1–19.
10. Blum WFP, Gyupta D. Heterogeneity of rat FSH by chromatofocusing:studies on in-vitro bioactivity of pituitary FSH forms and effect ofneuraminidase treatment. J Endocrinol 1985;105:17–27.
11. Anderson RE, Cragun JM, Chang RJ, Stanczyk FZ, Lobo RA. Apharmacodynamic comparison of human urinary follicle-stimulatinghormone and human menopausal gonadotropin in normal women andpolycystic ovary syndrome. Fertil Steril 1989;52:216–20.
FERTILITY & STERILITY� 1255