phagocytosis of salmonella montevideo by human neutrophils
TRANSCRIPT

200 • JID 2005:192 (15 July) • Pilsczek et al.
M A J O R A R T I C L E
Phagocytosis of Salmonella montevideo by HumanNeutrophils: Immune Adherence IncreasesPhagocytosis, whereas the Bacterial SurfaceDetermines the Route of Intracellular Processing
Florian H. Pilsczek,a Anne Nicholson-Weller, and Ionita GhiranHarvard-Thorndike Laboratory, Division of Allergy and Inflammation, and the Division of Infectious Disease, Department of Medicine, Beth IsraelDeaconess Medical Center, and Harvard Medical School, Boston, Massachusetts
Complement-opsonized particles become immune adherent to complement receptor 1 (CR1 or CD35) onhuman erythrocytes, allowing particles to be ingested by phagocytes in the liver and the spleen. We investigatedthe role that immune adherence plays in the uptake and killing of Salmonella montevideo by human neutrophils.Exposure to serum induced loss of flagella and facilitated immune adherence, which was followed by more-efficient phagocytosis and killing, compared with that after exposure to serum-opsonized, free bacteria. Onecorrelate of bacterial killing is the fusion of phagosomes with lysosomes, which can be monitored by Lyso-Tracker or lysosomal-associated membrane protein 2 colocalization with the intracellular bacteria. At 5 min,phagolysosmal fusion was significantly faster for immune-adherent bacteria than for non–immune-adherentbacteria, but, by 35 min, the difference between the 2 groups was minimal. Immune adherence also facilitatedthe ingestion of antibody complement–opsonized fluorescent microspheres, but, unlike bacteria, most inter-nalized microspheres failed to fuse with lysosomes. However, addition of lipopolysaccharide, a Toll-like receptorligand, to microspheres directed their intracellular trafficking, resulting in rapid lysosomal fusion. Thus,immune adherence facilitates phagocytosis, but the route of intracellular processing depends on the molecularnature of the target and is independent of host complement and antibody.
Complement opsonization of a particle significantly
augments its ingestion by phagocytes in tissues. In the
vascular space, where phagocytes are relatively rare,
complement-opsonized particles are immobilized to the
surface of cells for clearance by phagocytes in the liver
and the spleen. The process is known as immune-ad-
herence clearance. In humans and a few other primates,
Received 3 July 2004; accepted 16 February 2005; electronically published 15June 2005.
Financial support: National Institutes of Health (grants R01 AI42987 to A.N.-W. and R21 AI057983 to I.G.); Harvard Digestive Disease Center (Pilot FeasibilityGrant DK34854 to I.G.).
a Present affiliation: Department of Immunology and Microbiology, Albert Ein-stein College of Medicine, Yeshiva University, the Bronx, New York.
Reprints or correspondence: Dr. Ionita Ghiran, Division of Allergy and Inflam-mation, Beth Israel Deaconess Medical Center, 330 Brookline Ave., Room Dana617, Boston, MA 02215 ([email protected]).
The Journal of Infectious Diseases 2005; 192:200–9� 2005 by the Infectious Diseases Society of America. All rights reserved.0022-1899/2005/19202-0003$15.00
it is the erythrocyte using complement receptor 1 (CR1
or CD35) that ligates the particle [1, 2].
Enteric Salmonella species, in contrast to typhoidal
Salmonella species, usually cause self-limited infections
that last 3–5 days in healthy hosts. This time course of
illness suggests that innate immunity is usually able to
cure these infections, although it cannot prevent them
[3]. A high prevalence of nontyphoidal Salmonella bac-
teremia in neutropenic patients infected with HIV [4]
and in patients with chronic granulomatous disease [5]
indicates that sufficient numbers of functional neutro-
phils (polymorphonuclear leukocytes [PMNLs]) are crit-
ical for the human innate immune defense against these
organisms. Additionally, recent evidence from studies of
mice indicate that sinusoidal PMNLs in the liver and,
presumably, the spleen are responsible for clearance of
bacteria [6–8].
In the present study, we show that immune-adherent
Salmonella species are more efficiently ingested and killed
Downloaded from https://academic.oup.com/jid/article-abstract/192/2/200/855484by gueston 08 April 2018

Roles of Immune Adherence and TLRs • JID 2005:192 (15 July) • 201
Figure 1. Effect of normal human serum (NHS) on Salmonella montevideo motility. Stationary-phase bacteria were incubated with either buffer(A), heat-inactivated serum (HIS) (B), or NHS (C) for 10 min at 37�C with continuous rocking. Bacteria were washed and analyzed by time-lapse (0.75s) light microscopy, to document their movement. The tracks represent the “relative” distance, because each bacterium is moving in 3 dimensions,but only 2 dimensions can be measured by this method (see Participants, Materials, and Methods). Results are graphed as ( ).means � SD n p 100The experiment was performed 3 times, and incubation with NHS, compared with HIS, always led to decreased motility. * .P ! .01
by PMNLs than are equally opsonized non–immune-adherent
Salmonella species. Although immune adherence also enhances
phagocytosis of nonbiological particles, the ability to fuse with
lysosomal compartments is solely dependent on the molecular
composition of the particles and is independent of their im-
mune-adherence status.
PARTICIPANTS, MATERIALS, AND METHODS
Reagents. The following reagents were obtained as noted:
Hanks’ balanced salt solution (HBSS) with calcium and mag-
nesium but without phenol red (Gibco), HBSS with 0.1% bo-
vine serum albumin (BSA) (Sigma), gentamicin sulfate solution
(10 mg/mL) (United States Biological), sterile MilliQue water
(Millipore) filtered by use of a lipopolysaccharide (LPS)–free
Acrodisc filter (Pall), LysoTracker red, CellTracker green, Syto16
green, Hoescht 33342, Prolong Gold with DAPI, and Alexa 488–
BSA. A Via Gram staining kit (Molecular Probes) was also used.
Antibodies. The following antibodies were used in the pres-
ent study: mouse anti–lysosomal-associated membrane protein
(LAMP) 2 monoclonal antibody (MAb) (BD Pharmingen), rab-
bit anti-Salmonella immunogloubulin (Fitzgerald; lot P02050202),
fluorescein isothiocyanate (FITC)–labeled rabbit anti–Salmo-
nella flagella antigen (YVS 0703; Accurate Chemical and Sci-
entific), rabbit anti-BSA IgG, Alexa 488– and Alexa 594–con-
jugated goat anti-mouse IgG, Alexa 488–conjugated goat anti-
rabbit IgG (Molecular Probes), rabbit nonimmune rabbit IgG
(Jackson-ImmunoResearch), and antigen affinity-purified rab-
bit anti-CR1 [9].
Salmonella species and microspheres. S. montevideo (Amer-
ican Type Culture Collection) was grown overnight in Bacto
nutrient broth (Difco) and quantified (0.5 OD600 p 84.5 � 10
cells/mL). Bacteria were gently pelleted, washed, and resuspended
in HBSS. Wild-type (wt) S. typhimurium (SJW1103) and mutant
strains either lacking flagella (SJW1368) or possessing flagella
that could not rotate (SJW2960) were provided by May Macnab
(Yale University) [10]. Sulfate-latex microspheres (1.0 mmol/L)
(Interfacial Dynamics) were coated sequentially with Alexa 488–
BSA (Molecular Probes), rabbit anti-BSA IgG, and, in some ex-
periments, LPS (Escherichia coli; L3755; Sigma).
Human serum and cells. Normal human serum (NHS) was
obtained from healthy volunteers, and heat-inactivated serum
(HIS) was prepared by heating NHS for 50 min at 56�C. Eryth-
rocytes and PMNLs were isolated as described elsewhere [11]
and were used within 30 min of purification. Cells and serum
were obtained from autologous donors. The protocol for draw-
ing blood from healthy volunteers was approved by the Beth
Downloaded from https://academic.oup.com/jid/article-abstract/192/2/200/855484by gueston 08 April 2018

202 • JID 2005:192 (15 July) • Pilsczek et al.
Figure 2. Effect of normal human serum (NHS) on Salmonella montevideo flagella. Bacteria, stained blue with the DNA dye Hoescht 33342, wereincubated with either buffer, heat-inactivated serum (HIS), or NHS for 10 min at 37�C, washed, and treated with fluorescein isothiocyanate (FITC)–labeled rabbit anti–Salmonella flagella antigen. Images were acquired separately for Hoescht 33342 and FITC and then overlaid by use of IPLab. HISled to some clumping of flagella, likely due to antibody. NHS led to the bacteria being denuded of flagella. The flow-cytometric analysis shows rep-resentative FITC fluorescence histograms of S. montevideo prepared as above and analyzed within 15 min. As a negative control, S. montevideo wasincubated with nonimmune FITC-labeled rabbit antibody. This experiment was repeated 1 time, with similar results.
Figure 3. Effect of normal human serum (NHS) on immune adherenceof Salmonella montevideo to human erythrocytes. Shown are differen-tial interference contrast Nomarski images of 3 immune-adherent (NHS-treated) S. montevideo strains (arrows).
Israel Deaconess Medical Center Institutional Review Board,
and all participants provided informed consent.
Assay for bacterial motility. Bacteria ( ) were in-75 � 10
cubated with either buffer, 10% HIS in HBSS, or 10% NHS in
HBSS for 10 min at 37�C. Time-lapse phase-contrast images
recorded the “relative” distance traveled by individual organ-
isms during 0.75 s. Images were analyzed by use of IPLab (version
3.9; Scanalytics).
Staining for bacterial flagella. After exposure to either HIS
or NHS, bacteria were treated with FITC-labeled rabbit anti-
Salmonella flagella antigen (1:50) and 10 mg/mL Hoechst 33342
for 10 min and then were examined by fluorescence micros-
copy. In addition, bacteria opsonized under the standard con-
ditions were stained as noted and analyzed by flow cytometry
by use of a FACScan cytometer with CellQuest Pro software
(version 4.01; BD Biosciences). Fluorescence-activated cell sorter
buffer, unstained bacteria, and stained bacteria were examined
by forward side scatter and fluorescence microscopy, for iden-
tification of the bacteria and determination of the gate for fur-
ther analysis.
Immune-adherence protocol. S. montevideo bacteria were
grown for 12 h at 37�C in nutrient broth, and bacteria were
gently washed in HBSS. A total of S. montevideo bacteria75 � 10
were resuspended in 90 mL of HBSS, 10 mL of either NHS or
HIS was added, and the suspension was incubated for 10 min
at 37�C, for opsonization. Bacteria were washed once, 5 � 106
erythrocytes were added to S. montevideo, and the mix-61 � 10
Downloaded from https://academic.oup.com/jid/article-abstract/192/2/200/855484by gueston 08 April 2018

Roles of Immune Adherence and TLRs • JID 2005:192 (15 July) • 203
Figure 4. Effect of immune adherence on phagocytosis of Salmonella montevideo by human neutrophils (polymorphonuclear leukocytes [PMNLs]).A, Phagocytic assays (as described in Participants, Materials, and Methods) were set up to compare the percentage of phagocytosis (the percentageof PMNLs that had ingested intracellular S. montevideo). Buffer-treated bacteria with and without erythrocytes were inefficiently ingested (0%–1%)and were subtracted as background. B, The same experiment, analyzed for the phagocytic index (the mean no. of intracellular organisms in PMNLsthat had ingested at least 1 organism). Data are expressed as mean � SD nos. of intracellular bacteria ( ) for normal human serum (NHS)n p 30and for NHS-treated salmonellae plus erythrocytes (NHS + E) ( ). The significance of the difference between NHS samples with and withoutn p 50erythrocytes was assessed by use of Student’s t test. This experiment is representative of 4 experiments performed. HIS, heat-inactivated humanserum; HIS + E, HIS plus erythrocytes.
ture was incubated in HBSS for 10 min at 37�C. The percentage
of erythrocytes with immune-adherent S. montevideo was de-
termined by light microscopy and ranged from 0% to 5% for
HIS and from 70% to 75% for NHS.
Phagocytic assay. PMNLs ( ) were added to micro-55 � 10
fuge tubes containing 600 mL of HBSS/0.1% BSA. Serum-op-
sonized S. montevideo ( ), either free or immune adher-61 � 10
ent, were added to the PMNLs, and the mixture was incubated
Downloaded from https://academic.oup.com/jid/article-abstract/192/2/200/855484by gueston 08 April 2018

204 • JID 2005:192 (15 July) • Pilsczek et al.
Figure 5. Effect of immune adherence of Salmonella montevideo on intracellular killing in human neutrophils. Phagocytic reactions were preparedas for the phagocytic assay, and, after 5 min at 37�C, the polymorphonuclear leukocytes (PMNLs) were pelleted, washed to remove free bacteria, andincubated with gentamicin to kill PMNL-adherent, noningested bacteria. After pelleting and resuspension in Hanks’ balanced salt solution, aliquots ofPMNLs were removed from each tube at 45 and 105 min after the initiation of phagocytosis and lysed with sterile water. Four replicate samplesfrom each lysed mixture were diluted and plated for determination of colony-forming units, as described in Participants, Materials, and Methods. Dataare mean � SD colony-forming units ( ). The reduction in colony-forming units (shown as the percentage next to each pair of bars) for bacterian p 4opsonized with normal human serum (NHS) was not significant ( ) but was significant for immune-adherent bacteria (NHS-treated salmonellaeP p .09plus erythrocytes [NHS + E]) ( ), as assessed by use of Student’s t test. This experiment was repeated 3 times, with similar results. HIS,P p .018heat-inactivated human serum; HIS + E, HIS plus erythrocytes. * .P p .03
for 5 min at 37�C with end-over-end rotation at 8 rpm, as
described elsewhere [12]. PMNLs were counted by light mi-
croscopy and analyzed for the percentage of phagocytosis (the
percentage of PMNLs that had ingested intracellular S. mon-
tevideo) and the phagocytic index (the mean number of intra-
cellular organisms in PMNLs that had ingested at least 1 or-
ganism). Buffer-treated organisms had a phagocytic index of
0%–11%, which was always less than that of HIS-opsonized
organisms and was considered to be the baseline value.
Intracellular killing assay. Phagocytic mixtures were centri-
fuged to remove non–immune-adherent, noningested bacteria,
and cells were subjected to hypotonic saline to lyse erythrocytes
in the tubes that contained them. PMNLs were resuspended in
gentamicin (10 mg/mL of HBSS) for 30 min at room temper-
ature to kill extracellular, immune-adherent bacteria. Tubes were
centrifuged at 3000 g for 3 min, and the PMNL pellets were
suspended in 400 mL of HBSS. Samples were collected im-
mediately (∼45 min after the initiation of phagocytosis) and
again after an additional 60 min of incubation at 37�C. Viability
sampling consisted of 4 replicate 7-mL aliquots, each diluted
with 93 mL of sterile water and plated on separate petri dishes
made with blood agar base number 2 (Difco). The plates for
colony counts were incubated for 18 h at 37�C.
Protocol for intracellular localization. Purified PMNLs
suspended in HBSS were loaded with 50 nmol/L LysoTracker
for 30 min at 37�C, washed twice, and resuspended at a con-
centration of cells/mL in HBSS. S. Montevideo bacteria71 � 10
were loaded with 100 nmol/L Syto16 green for 30 min at 4�C
in HBSS, washed twice with HBSS, opsonized with NHS, and
resuspended at a concentration of bacteria/mL. PMNLs81 � 10
and opsonized bacteria with or without erythrocytes for im-
mune adherence were mixed as described above. Aliquots (100
mL) from each sample were cytospun, and slides were mounted
with Prolong Gold with DAPI. Cells were examined by use of
an AX-70 Provis Olympus fluorescence microscope equipped
with FITC and rhodamine filters (Olympus America) and UPLApo
40 � 1.0 Ph 3 and 100 � 1.35 Ph 3 objectives. Intracellular
bacteria not associated with the lysosomal compartment fluo-
resced in the green channel alone, whereas intralysosomal bac-
teria fluoresced in both the green and the red channels. Images
were acquired by use of a Retiga EXi digital camera (QImaging),
and the images were further processed by use of IPLab. Alter-
natively, cells were fixed with 3.7% paraformaldehyde (EM Sci-
ences) for 1 h, washed twice in HBSS, and permeabilized for
10 min in 0.2% saponin (Sigma-Aldrich). Cells were blocked
with HBSS plus 4% NHS and 4% normal goat serum for 15
min at room temperature and were incubated with mouse anti–
LAMP-2 MAb (5 mg/mL) for 20 min. Cells were washed twice
Downloaded from https://academic.oup.com/jid/article-abstract/192/2/200/855484by gueston 08 April 2018
Roles of Immune Adherence and TLRs • JID 2005:192 (15 July) • 205
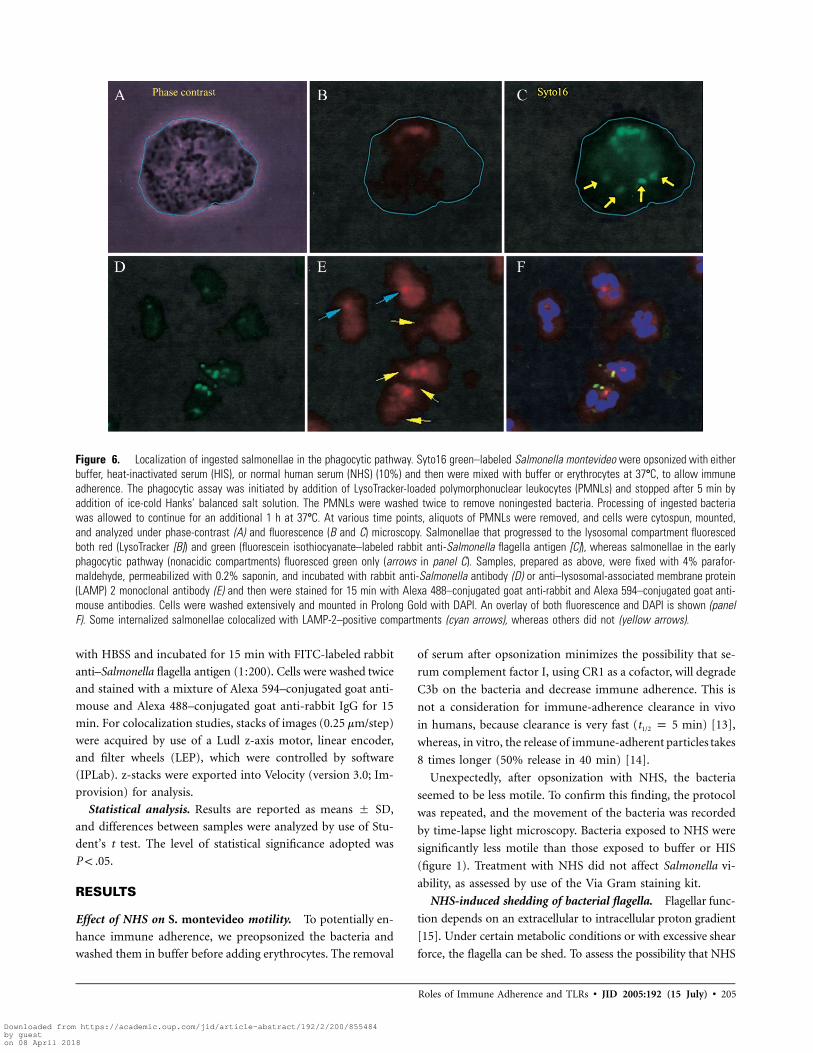
Figure 6. Localization of ingested salmonellae in the phagocytic pathway. Syto16 green–labeled Salmonella montevideo were opsonized with eitherbuffer, heat-inactivated serum (HIS), or normal human serum (NHS) (10%) and then were mixed with buffer or erythrocytes at 37�C, to allow immuneadherence. The phagocytic assay was initiated by addition of LysoTracker-loaded polymorphonuclear leukocytes (PMNLs) and stopped after 5 min byaddition of ice-cold Hanks’ balanced salt solution. The PMNLs were washed twice to remove noningested bacteria. Processing of ingested bacteriawas allowed to continue for an additional 1 h at 37�C. At various time points, aliquots of PMNLs were removed, and cells were cytospun, mounted,and analyzed under phase-contrast (A) and fluorescence (B and C) microscopy. Salmonellae that progressed to the lysosomal compartment fluorescedboth red (LysoTracker [B]) and green (fluorescein isothiocyanate–labeled rabbit anti-Salmonella flagella antigen [C]), whereas salmonellae in the earlyphagocytic pathway (nonacidic compartments) fluoresced green only (arrows in panel C). Samples, prepared as above, were fixed with 4% parafor-maldehyde, permeabilized with 0.2% saponin, and incubated with rabbit anti-Salmonella antibody (D) or anti–lysosomal-associated membrane protein(LAMP) 2 monoclonal antibody (E) and then were stained for 15 min with Alexa 488–conjugated goat anti-rabbit and Alexa 594–conjugated goat anti-mouse antibodies. Cells were washed extensively and mounted in Prolong Gold with DAPI. An overlay of both fluorescence and DAPI is shown (panelF). Some internalized salmonellae colocalized with LAMP-2–positive compartments (cyan arrows), whereas others did not (yellow arrows).
with HBSS and incubated for 15 min with FITC-labeled rabbit
anti–Salmonella flagella antigen (1:200). Cells were washed twice
and stained with a mixture of Alexa 594–conjugated goat anti-
mouse and Alexa 488–conjugated goat anti-rabbit IgG for 15
min. For colocalization studies, stacks of images (0.25 mm/step)
were acquired by use of a Ludl z-axis motor, linear encoder,
and filter wheels (LEP), which were controlled by software
(IPLab). z-stacks were exported into Velocity (version 3.0; Im-
provision) for analysis.
Statistical analysis. Results are reported as means � SD,
and differences between samples were analyzed by use of Stu-
dent’s t test. The level of statistical significance adopted was
.P ! .05
RESULTS
Effect of NHS on S. montevideo motility. To potentially en-
hance immune adherence, we preopsonized the bacteria and
washed them in buffer before adding erythrocytes. The removal
of serum after opsonization minimizes the possibility that se-
rum complement factor I, using CR1 as a cofactor, will degrade
C3b on the bacteria and decrease immune adherence. This is
not a consideration for immune-adherence clearance in vivo
in humans, because clearance is very fast (t1/2 p 5 min) [13],
whereas, in vitro, the release of immune-adherent particles takes
8 times longer (50% release in 40 min) [14].
Unexpectedly, after opsonization with NHS, the bacteria
seemed to be less motile. To confirm this finding, the protocol
was repeated, and the movement of the bacteria was recorded
by time-lapse light microscopy. Bacteria exposed to NHS were
significantly less motile than those exposed to buffer or HIS
(figure 1). Treatment with NHS did not affect Salmonella vi-
ability, as assessed by use of the Via Gram staining kit.
NHS-induced shedding of bacterial flagella. Flagellar func-
tion depends on an extracellular to intracellular proton gradient
[15]. Under certain metabolic conditions or with excessive shear
force, the flagella can be shed. To assess the possibility that NHS
Downloaded from https://academic.oup.com/jid/article-abstract/192/2/200/855484by gueston 08 April 2018

206 • JID 2005:192 (15 July) • Pilsczek et al.
Table 1. Effect of immune adherence onthe fusion between phagosomes and the ly-sosomal compartment.
Incubation time, min
Bacteriaopsonized with
NHS NHS + E
5 50a 9235 86 94
NOTE. Data are percentage of phagocytosis.NHS, normal human serum; NHS + E, NHS-treatedsalmonellae plus erythrocytes.
a (Intralysosomal bacteria/total bacteria ingested)� 100 p percentage of lysosomes.
Table 2. Effect of immune adherence on phagocytosisof opsonized microspheres.
Phagocytosis time, min IA beads Non-IA beads
5 11/14 0/035 34/42 13/19
NOTE. Data are percentage of phagocytosis/phagocytic index.Beads (microspheres) were incubated with rabbit IgG, Alexa 488–bovine serum albumin, and normal human serum. IA, immuneadherent.
might induce the shedding of flagella on S. montevideo, we used
both flow cytometry and fluorescence microscopy. Bacteria that
had been stained with a DNA dye were exposed to either buffer,
HIS, or NHS and then were stained with FITC-labeled rabbit
anti–Salmonella flagella antigen. NHS induced the shedding of
flagella, with only green flagellin–positive stubs associated with
the bacterial body, seen as a green outline in figure 2. The shed
flagella accumulated in large clumps, suggesting that they had
detached proximal to the bacterial surface. Samples that were
analyzed by microscopy were also analyzed by flow cytometry,
with similar results (figure 2). NHS opsonization rendered the
S. montevideo less motile, and the bacteria became immune ad-
herent, with consistently ∼70% of erythrocytes having �1 ba-
cillus attached (figure 3).
To assess whether NHS-induced loss of motility and becom-
ing immune adherent to erythrocytes were functionally linked
phenomena, we performed additional studies. S. typhimurium
was used because of the availability of a wt strain (SJW1103)
and 2 nonmotile mutant strains: 1 lacking flagella (SJW1368)
and 1 possessing flagella that were nonfunctional because of a
defective motor (SJW2960). With or without exposure to NHS,
the wt S. typhimurium strain remained motile, whereas the 2
mutant S. typhimurium strains were nonmotile, as determined
by microscopic observation. In a quantitative assay for im-
mune adherence, each S. typhimurium strain was exposed to
HIS or NHS and then was mixed with erythrocytes and ex-
amined for immune adherence by light microscopy. Sixty-one
percent of flagella-lacking S. typhimurium and 81% of non-
motile S. typhimurium, compared with only 33% of wt S.
typhimurium, became immune adherent. Although the wt S.
typhimurium remained motile after opsonization, there was a
marked enhancement of immune adherence in the nonmotile
mutant strains. Thus, the NHS-induced flagellar shedding of
S. montevideo may not have been essential for immune adher-
ence, but, likely, it greatly enhanced it.
Effect of immune adherence on phagocytosis of S. monte-
video. Various species of bacteria are phagocytosed more ef-
ficiently when immune adherent [1, 16]. To determine whether
immune adherence affected the uptake of S. montevideo by
human PMNLs, we compared the ingestion of opsonized im-
mune-adherent bacteria with that of opsonized non–immune-
adherent S. montevideo. Immune-adherent S. montevideo were
ingested more efficiently than were non–immune-adherent S.
montevideo, as measured by both the percentage of phagocytosis
(50% vs. 29%) (figure 4A) and the phagocytic index (3.7 vs.
2.7) (figure 4B).
Effect of of S. montevideo. The increased phagocytosis of
immune-adherent S. montevideo might favor either the host or
the pathogen. We therefore investigated the survival of S. mon-
tevideo within PMNLs after the bacteria in suspension were
washed away and the immune-adherent, noningested bacteria
were killed by gentamicin, manipulations that were completed
45 min after the initiation of phagocytosis. The viability of in-
tracellular bacteria was assessed after 45 min and again after an
additional 60 min of incubation (105 min total) at 37�C. These
times were chosen because longer incubation times (120 min)
resulted in all the bacteria being killed whether they had been
free or immune adherent.
PMNLs were lysed, and the lysate was diluted, plated in
quadruplicate, and incubated for 18 h, to allow enumeration
of viable colonies. The relative number of viable intracellular
organisms at 45 min (figure 5) reflected the efficiency of phago-
cytosis for bacteria opsonized under different conditions (figure
4A and 4B). During the subsequent 60 min of incubation, kill-
ing of previously immune-adherent S. montevideo was more
efficient (60% decrease in colony-forming units between 45
and 105 min) (figure 5) than was killing of NHS-opsonized S.
montevideo (27% decrease in colony-forming units during the
same period) (figure 5).
Effect of immune adherence on the rate of fusion of bacteria-
containing phagosomes with lysosomes. Many intracellular
bacterial pathogens, including S. typhi, delay the fusion of the
phagosome with the lysosomal compartment to subvert the host
cell’s microbicidal mechanisms (reviewed in [17, 18]). We won-
dered whether the enhanced efficiency of killing of previously
immune-adherent S. montevideo might relate to enhanced fusion
of PMNL phagosomes with lysosomes, and we devised an assay
to test this. Syto16 green was used to label S. montevideo, where-
as intracellular acidic compartments, predominately lysosomes,
Downloaded from https://academic.oup.com/jid/article-abstract/192/2/200/855484by gueston 08 April 2018

Roles of Immune Adherence and TLRs • JID 2005:192 (15 July) • 207
Figure 7. Induction of phagolysosomal fusion by lipopolysaccharide(LPS). A, Polymorphonuclear leukocytes were incubated with opsonizedAlexa 488–bovine serum albumin microspheres with and without LPSfor 5 min in suspension, washed twice, and seeded on glass. At theindicated time points, cells were fixed with 4% paraformaldehyde. Afterfixation, cells were permeabilized with 0.2% saponin and incubatedwith anti–lysosomal-associated membrane protein (LAMP) 2 monoclonalantibody. Cells were washed and incubated with Alexa 594–conjugatedgoat anti-mouse IgG for an additional 15 min, washed extensively, andmounted by use of Prolong Gold with DAPI. A, An overlaid image, withthe nucleus stained in blue, LAMP-2–positive compartments in red, andingested microspheres in green. Only LAMP-2–positive structures witha definite ring appearance around microspheres (yellow arrows) wereconsidered to be positive for colocalization. LAMP-2 structures withround-shaped distribution around beads due to a physical displacement(yellow arrowheads) were not counted. B, Distribution of internalizedmicrospheres, with respect to LAMP-2 compartments at different timepoints. At least 80 cells were counted for LAMP-2–positive intracellularmicrospheres for both the 5-min and the 45-min time points. Data aremeans � SD. This experiment was repeated twice, with similar results.* .P p .02
were labeled red with LysoTracker dye [19]. After incubation for
5 min at 37�C, to allow phagocytosis, and again after an additional
30 min of incubation (35 min total), samples were removed
immediately. A representative postphagocytic PMNL is shown in
figure 6A–6C, to indicate how phagosomes were distinguished
from phagolysosomes. The total number of intracellular bacteria
was compared with those bacteria in the lysosomal compartment.
Although the percentage of phagolysosomal fusion was higher
at 5 min, if the bacterium had been previously immune adherent,
by 35 min, the difference between free and immune-adherent
bacteria was minimal (table 1).
Although LysoTracker is well accepted in the literature as a
marker of lysosomes, there is the possibility that the dye marks
other compartments with low pH. To address this concern, we
repeated the experiment but replaced LysoTracker with anti–
LAMP-2 MAb (figure 6E) and replaced Syto16 green with rabbit
anti-Salmonella antibody (figure 6D). Because LAMP-2 has a
diffuse distribution in PMNLs, the clustering around ingested
salmonellae or the lack of clustering was relatively easy to iden-
tify and quantify (figure 6E). Although immune-adherent bac-
teria were more rapidly ingested, there seemed to be a minimal
effect on intracellular trafficking. To monitor more accurately
the intracellular movement of phagosomes, we switched to Al-
exa 488–BSA–coated latex beads (microspheres) that had been
opsonized with rabbit anti-BSA IgG and NHS. Being perfectly
spherical and birefringent, microspheres were easy to identify
by fluorescence and bright-field microscopy. To control for the
presence of erythrocytes in the non–immune-adherent samples,
we included an equivalent number of human erythrocytes that
had been pretreated with antigen affinity-purified rabbit anti-
CR1, which completely blocks immune adherence. Immune
adherence stimulated the rate of phagocytosis of the micro-
spheres (table 2). In striking contrast to the bacteria (whether
free or immune adherent), at 35 min, the majority (83%) of
the ingested microspheres remained in a LAMP-2–negative com-
partment, indicating that their phagosomes had not fused with
lysosomes. These data suggest that, independent of being im-
mune adherent and independent of antibody-complement op-
sonins, the molecular determinants of the target itself might
determine intracellular trafficking within PMNLs.
Effect of LPS, a Toll-like receptor (TLR) ligand, on phago-
somes containing latex microspheres. To test whether the
TLR ligand LPS could affect trafficking of phagosomes, PMNLs
were incubated with opsonized beads with or without LPS for
5 min at 37�C and then were washed twice to remove the
noningested microspheres. Cells were seeded on chamber slides
and fixed every 10 min for the next 35 min. Although im-
mobilized LPS had no significant effect on the percentage of
phagocytosis or the phagocytic index (LPS beads, 19%; beads,
21%), there was a striking difference (figure 7A) in the number
of microspheres with fused phagolysosomes. We considered
Downloaded from https://academic.oup.com/jid/article-abstract/192/2/200/855484by gueston 08 April 2018

208 • JID 2005:192 (15 July) • Pilsczek et al.
Figure 8. Induction of phagolysosomal fusion by lipopolysaccharide. Polymorphonuclear leukocytes were incubated for 5 min with a 1:1 mixtureof salmonellae and Alexa 488–conjugated goat anti-rabbit IgG/complement–coated microspheres, washed twice, and seeded on glass. Cells werefixed after 10 min, permeabilized, and incubated with rabbit anti-Salmonella immunoglobulin (A) and mouse anti–lysosomal-associated membraneprotein (LAMP) 2 monoclonal antibody (B) for 15 min at room temperature. Slides were washed and incubated with Alexa 488–conjugated goat anti-rabbit (A) and Alexa 594–conjugated goat anti-mouse (B) antibodies. Cells were washed repeatedly and mounted in Prolong Gold with DAPI. A mergedimage (C) shows yellow bacteria (due to colocalization) and green microspheres (due to a lack of colocalization with LAMP-2–positive compartments).
LAMP-2–positive microspheres to be only those microspheres
that had a well-defined structured ring around them (figure
7D), and not those microspheres that just displaced the LAMP-
2–positive structures around them (figure 7A).
Effect of PMNLs on the trafficking specificity of various
ingested particles. We next asked whether phagocytosis of
opsonized salmonellae would induce ingested microspheres to
fuse with lysosomes. We mixed (1:1) opsonized salmonellae and
opsonized Alexa 488 microspheres (lacking LPS) with PMNLs
for 5 min of phagocytosis. After washing, PMNLs were seeded
on glass for 15 min. Subsequently, the cells were fixed with
paraformaldehyde, permeabilized, and stained for bacteria with
anti-Salmonella MAb and anti–LAMP-2 MAb. A typical cell
(figure 8) ingested both types of particles. Salmonellae and
microspheres followed distinct pathways within the same cell:
bacteria colocalized with LAMP-2, whereas microspheres re-
mained in a LAMP-2–negative compartment.
DISCUSSION
We have investigated how immune adherence affects the killing
of S. montevideo. Because erythrocyte-dependent immune ad-
herence is unique to humans and a few other higher primates,
our in vitro studies used human reagents. Although salmonellae
can activate complement in the absence of antibody [20], it is
likely that our sources of NHS contained specific antibody, be-
cause serum samples from some donors supported immune
adherence better than serum samples from other donors.
Nontyphoidal salmonellae have diverse virulence factors [21],
and, thus, it was not surprising that exposure to NHS induced
S. montevideo, but not S. typhimurium, to shed its flagella.
Flagellar shedding not only promotes immune adherence, it
also has the potential to disseminate an inflammatory response
in vivo. Flagellin, the major flagellar protein (reviewed in [22]),
binds TLR5 [23, 24] and, in terms of induction of proinflam-
matory genes, is the most important virulence factor of S.
typhimurium [25].
Our data indicate that immune adherence accelerated uptake
and intracellular trafficking of antibody complement–opso-
nized S. montevideo and microspheres (tables 1 and 2 and fig-
ures 4A, 4B, and 5). Nelson hypothesized that enhancement of
immune adherence of phagocytosis might be due to immobili-
zation of the bacteria [1], because trapping bacteria against
surfaces had been shown to enhance the efficiency of phago-
cytosis [26] or because the erythrocyte stimulated the phago-
cyte. We observed minimal phagocytosis when HIS was used
as an opsonin, and phagocytosis was not enhanced by the pres-
ence of bystander erythrocytes (figure 4A and 4B). This finding
argues against the idea that erythrocytes provide global stim-
ulation of phagocytosis, but it does not rule out the possibility
that direct stimulation occurs when the phagocyte contacts a
particular erythrocyte to remove the immune-adherent target.
Our experiments with antibody complement–opsonized mi-
crospheres confirmed that intracellular trafficking was not affected
by immune adherence: ∼80% of ingested microspheres remained
in LAMP-2–negative compartments regardless of whether they
were free or immune adherent. However, the addition of a TLR
ligand, LPS, to the target signaled for phagolysosomal fusion.
It has been found by use of serum-opsonized green fluorescent
protein E. coli as targets and mouse macrophages as effectors
that an intact TLR signaling pathway leads to a higher rate of
phagocytosis [27]. In contrast, we did not find that the addition
of LPS to our microspheres enhanced either the percentage of
phagocytosis or the phagocytic index. These discrepant results
may relate to the fact that we used different targets and different
effector cells. As was shown for murine macrophages [27], a
single human PMNL could ingest a TLR-ligating target (bac-
Downloaded from https://academic.oup.com/jid/article-abstract/192/2/200/855484by gueston 08 April 2018

Roles of Immune Adherence and TLRs • JID 2005:192 (15 July) • 209
teria) and a non–TLR-ligating particle (microsphere), and their
trafficking was different: the bacteria stimulated rapid lysosomal
fusion, and the microsphere did not. Thus, engagement of TLRs
takes place in nascent phagosomes and signals for fusion of
that particular phagosome with the lysosomal compartment.
In summary, we have found that immune adherence facili-
tates phagocytosis of S. montevideo and that it likely is a host
defense mechanism for other nontyphoidal Salmonella strains.
During the course of these experiments, 2 novel findings were
noted: first, opsonization with NHS caused S. montevideo, but
not S. typhimurium, to shed its flagella, a finding that has im-
plications for TLR5 signaling; and, second, the enhanced killing
of immune-adherent S. montevideo was correlated with more-
rapid phagolysosomal fusion in PMNLs. Intracellular traffick-
ing, however, was not dictated either by host opsonins or by
immune adherence, but rather by the biochemical composition
of the target.
Acknowledgments
We are grateful to May Macnab (Yale University) for providing the wild-type and motility mutant strains of Salmonella typhimurium and to GregWeaver and Dr. Peter F. Weller (Beth Israel Deaconess Medical Center) fortechnical expertise and critical reading of the manuscript, respectively.
References
1. Nelson RAJ. The immune adherence phenomenon: an immunologi-cally specific reaction between microorganisms and erythrocytes lead-ing to enhanced phagocytosis. Science 1953; 118:733–7.
2. Fearon DT. Identification of the membrane glycoprotein that is theC3b receptor of the human erythrocyte, polymorphonuclear leukocyte,B lymphocyte and monocyte. J Exp Med 1980; 152:20–30.
3. Hohmann EL. Nontyphoidal salmonellosis. Clin Infect Dis 2001; 32:263–9.
4. Tumbarello M, Tacconelli E, Caponera S, Cauda R, Ortona L. Theimpact of bacteraemia on HIV infection: nine years experience in alarge Italian university hospital. J Infect 1995; 31:123–31.
5. Winkelstein JA, Marino MC, Johnston RB Jr, et al. Chronic granulo-matous disease: report on a national registry of 368 patients. Medicine(Baltimore) 2000; 79:155–69.
6. Vassiloyanakopoulos AP, Okamoto S, Fierer J. The crucial role of poly-morphonuclear leukocytes in resistance to Salmonella dublin infectionsin genetically susceptible and resistant mice. Proc Natl Acad Sci USA1998; 95:7676–81.
7. Fierer J. Polymorphonuclear leukocytes and innate immunity to Sal-monella infections in mice. Microbes Infect 2001; 3:1233–7.
8. Wing EJ, Gregory SH. Listeria monocytogenes: clinical and experi-mental update. J Infect Dis 2002; 185(Suppl 1):S18–24.
9. Fearon DT, Collins LA. Increased expression of C3b receptors on poly-morphonuclear leukocytes by chemotactic factors and by purificationprocedures. J Immunol 1983; 130:370–5.
10. Ohnishi K, Ohto Y, Aizawa S, Macnab RM, Iino T. FlgD is a scaffoldingprotein needed for flagellar hook assembly in Salmonella typhimurium.J Bacteriol 1994; 176:2272–81.
11. Tyagi SR, Nicholson-Weller A, Barbashov SF, Tas SW, Klickstein LB.Intracellular adhesion molecule 1 and b2 integrins in C1q-stimulatedsuperoxide production by human neutrophils: an example of a generalregulatory mechanism governing acute inflammation. Arthritis Rheum2000; 43:2248–59.
12. Ghiran I, Barbashov SF, Klickstein LB, Tas SW, Jensenius JC, Nicholson-Weller A. Complement receptor 1/CD35 is a receptor for mannan-binding lectin. J Exp Med 2000; 192:1797–808.
13. Schifferli JA, Ng YC, Estreicher J, Walport MJ. The clearance of tetanustoxoid-anti-tetanus toxoid immune complexes from the circulation ofhumans: complement- and erythrocyte complement receptor 1-depen-dent mechanisms. J Immunol 1988; 140:899–904.
14. Jepsen HH, Svehag SE, Jensenius JC, Sim RB. Release of immune com-plexes bound to erythrocyte complement receptor (CR1), with particularreference to the role of factor I. Scand J Immunol 1986; 24:205–13.
15. Minamino T, Imae Y, Oosawa F, Kobayashi Y, Oosawa K. Effect ofintracellular pH on rotational speed of bacterial flagellar motors. J Bac-teriol 2003; 185:1190–4.
16. Lindorfer MA, Nardin A, Foley PL, et al. Targeting of Pseudomonasaeruginosa in the bloodstream with bispecific monoclonal antibodies.J Immunol 2001; 167:2240–9.
17. Duclos S, Desjardins M. Subversion of a young phagosome: the survivalstrategies of intracellular pathogens. Cell Microbiol 2000; 2:365–77.
18. Gorvel JP, Meresse S. Maturation steps of the Salmonella-containingvacuole. Microbes Infect 2001; 3:1299–303.
19. Rajotte D, Stearns CD, Kabcenell AK. Isolation of mast cell secretorylysosomes using flow cytometry. Cytometry A 2003; 55:94–101.
20. Morrison DC, Kline LF. Activation of the classical and properdin path-ways of complement by bacterial lipopolysaccharides (LPS). J Immunol1977; 118:362–8.
21. Fierer J, Guiney DG. Diverse virulence traits underlying different clin-ical outcomes of Salmonella infection. J Clin Invest 2001; 107:775–80.
22. Underhill DM, Ozinsky A. Toll-like receptors: key mediators of microbedetection. Curr Opin Immunol 2002; 14:103–10.
23. Hayashi F, Smith KD, Ozinsky A, et al. The innate immune responseto bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001;410:1099–103.
24. Gewirtz AT, Navas TA, Lyons S, Godowski PJ, Madara JL. Cutting edge:bacterial flagellin activates basolaterally expressed TLR5 to induce epi-thelial proinflammatory gene expression. J Immunol 2001; 167:1882–5.
25. Zeng H, Carlson AQ, Guo Y, et al. Flagellin is the major proinflam-matory determinant of enteropathogenic Salmonella. J Immunol 2003;171:3668–74.
26. Wood WB Jr, Smith MR. Intercellular surface phagocytosis. Science1947; 106:86.
27. Blander JM, Medzhitov R. Regulation of phagosome maturation bysignals from Toll-like receptors. Science 2004; 304:1014–8.
Downloaded from https://academic.oup.com/jid/article-abstract/192/2/200/855484by gueston 08 April 2018