phagocytosis in acanthamoeba: i. a mannose receptor is responsible for the binding and phagocytosis...
TRANSCRIPT

JOURNAL OF CELLULAR PHYSIOLOGY 145508-513 (1990)
Phagocytosis in Acanthamoeba: I. A Mannose Receptor Is Responsible for the Binding and
Phagocytosis of Yeast PHILIP C. ALLEN' AND ELIEZAR A. DAWIDOWICZ
Program in Cell and Developmental Biology, Harvard Medical School, Boston, Massachusetts 02 I 15 (P.C.A.); Department of Physiology, Tufts University School of
Medicine, Boston, Massachusetts 02 J I I ( € . A D . )
We have examined the initial events in phagocytosis by Acanthamoebe castellanii in order to understand this process at the molecular level and have determined that phagocytosis in this organism is mediated by a receptor which recognizes mannose-rich elements in the particle to be phagocytosed. We demonstrate that the binding and internalization of yeast particles can be inhibited by the sugars D(+)-mannose and D(-)-fructose in a stereospecific, concentration-dependent manner. This inhibition is specific; these sugars did not inhibi t the uptake of latex beads by this organism. Using mannosylated neoglycoproteins, which are much more potent inhibitors of particle binding as compared with the free sugar, we demonstrate the presence of a receptor on the amoeba cell surface which is necessary for the binding of yeast as the initial event of phagocytosis. The Acanthamoeba mannose receptor also appears to be able to mediate the delivery of soluble mannose-rich molecules to a degradative compartment such as the lysosome. Knowledge of this receptor will allow a better understanding of the molecular events of phagocytosis.
Phagoc tosis, a s ecialized form of endoc tosis in
extracellular milieu, is used by many organisms for immunological defense and in protozoans is the essen- tial means of food gathering. The initial event in phagocytosis can be considered to be the binding of a particle to the extracellular surface of the plasma membrane. In mammalian macrophages and neutro- phils, phagocytosis has been shown to involve s ecific
1988; Newman et al., 1985; Ukkonen et al., 1986). However, the mechanism by which the particle-re- ceptor interaction activates the internalization process is not completely understood.
In order to elucidate the initial steps of the phago- c tic pathway we have studied phagocytosis of yeast by
phagocytic protozoan is capable of the internalization of a wide varlet of particle types including latex beads (Korn, 1974), gacteria (Preston and King, 1984), eryth- rocytes (Rabinovitch and destephano, 19711, and yeast (Bowers and Olszewski, 1983). This organism also shows preferential u take of yeast over latex beads (Bowers and Olszews K i, 1983). Previous studies on the phagocytosis of latex beads have demonstrated that the availability of intracellular vacuoles ap ears to be a
1969). Acanthamoeba can be grown in large uantities
variety of biosynthetic precursors (Ulsamer et al., 19691, which have facilitated these previous biochem- ical studies.
In the studies described in this paper we have
which ce r 1s internayize insoluble particles P rom the
cell surface receptors (Lennartz et al., 1989; Me P lman,
t K e amoeba Acanthamoeba castellanii. This avidly
limiting step in the phagocytic process ( If lsamer et al.,
in fluid culture (Korn, 1974) and readily labe 7 ed with a
C 1990 WILEY-LISS, INC.
extended the observation of Brown et al. (19751, who demonstrated that phagocytosis of erythrocytes by Acantharnoeba could be inhibited by the free sugars mannose and fructose at concentrations of approxi- mately 10 mM. We have found that the neoglycopro- teins man,,-BSA and man17-BSA are more otent inhibitors by several orders of magnitude, whic R sug- gested to us that these compounds might be useful in the study of the molecular details of phagocytosis. Using these molecules we have obtained evidence for the existence of a receptor on the Acunthamoebu cell surface which can recognize mannose residues on the yeast cell wall as the initial event in phagocytosis. In the accom anying paper we present our findings which
transduction pathway: the degradation of PIPz by phospholipase C.
MATERIALS AND METHODS A. castellanii were cultured xenically in roteose-
(Mills et al., 1984; Korn, 1974). Ce ti s were harvested at 1 x 106 cellslml or less.
Quantitation of binding and phagocytosis of yeast by amoeba
The internalization of heat-killed yeast (HKY) by Acanthamoeba was quantified using an adaptation of
indicate t R a t this receptor activates a known signal
peptone, glucose media at 28°C as reviously B escribed
Received March 27, 1990; accepted September 4, 1990. *To whom reprint requestslcorrespondence should be addressed at LHRRB, Harvard Medical School, 45 Shattuck St., Boston, MA 02115.
RECEPTOR-MEDIATED PHAGOCYTOSIS IN ACANTHAMOEBA 509
the fluorescence method of Sahlin et al. (1983). In summary, we utilized the membrane impermeant dye trypan blue to quench the fluorescence of fluorescein isothiocyanate-labeled HKY (FITC-HKY) which had not been internalized by the amoeba. The remaining fluorescence was derived from particles which had been internalized.
Yeast (Saccharomyces cerevisiae, Type 11; Sigma Chemical Co., St. Louis, MO) were resuspended in PBS, H 7.2 with stirring for 1 hour. They were killed by
[eating to 90°C for 1 hour, allowed to cool to room temperature, and washed five times with PBS by centrifugation in a clinical centrifuge. Yeast were labeled at a concentration of 10’ yeast/ml with 0.05 m /ml FITC (Molecular Probes, Eugene OR) in 50 mM
temperature. After labelin the cells were washed 5 times with 5 volumes of PB f! , pH 7.2, and stored at 4°C. Aliquots were washed once in the appropriate phago- cytosis buffer before use to remove any components which had been solubilized from the cells during stor- age.
Phagocytosis assays were performed in buffers con- taining various concentrations of sugars as indicated in Figure 1 and 15 mM NaH2P04, pH 7.2. Buffers con- tainin 10 mM mannose or fructose were brought to
washed twice with the appropriate buffer. Amoeba and FITC-HKY were incubated separately at 28°C for 30 minutes. Phagoc tosis was initiated by the addition of 4 x lo’ FITC-HKY per lo6 amoeba. At various times after this addition, 1 ml aliquots were removed and diluted 10-fold into a solution containing 2 mg/ml trypan blue, 0.8% Na Azide in PBS at 2°C. Each sample was gently washed 3 times with PBS at 2°C by centrif- ugation at setting 1 on an IEC clinical tabletop centri- fuge for 5 minutes (International Equipment Corp. Needham Heights, MA) to remove excess dye and unattached east. This buffer prevented further uptake
in extracellular yeast. k v o methods were used to quantitate the uptake of
FITC-HKY. These were either b direct measurement
spectrophotometer a t excitation and emission wave- lengths of 490 and 530 nm, respectively, or by micro- scopic examination to determine the fraction of cells which had internalized yeast using a fluorescence
microscoge. The p agocytosis of latex beads by amoeba was determined under conditions described above. Amoeba were challenged with latex beads 1.1 pm in diameter (PolySciences, Warrington, PA) at a final concentra- tion of 0.1%. Cells were centrifuged, as described above, a t the appro riate time and resus ended in fresh medium at 4’5. The uptake of latex 1 eads was deter- mined, by microscopic examination with phase contrast illumination (400 x magnification), counting the num- ber of cells with, and without, associated beads.
The binding of yeast to amoeba was measured at 4°C. Amoeba (lo6 in 1 ml) were incubated with lo7 yeast cells and gently rotated on a Lab Quake shaker (Lab Industries Inc., Berkeley CA) for 30 minutes at 4°C. The binding reaction was stopped by diluting the assay
so % ium borate buffer, pH 9.0, for 1 hour a t room
100 m a with 90 mM mannitol. Prior to use cells were
of yeast an B quenched the fluorescence of any remain-
of fluorescence using a Perkin-E r mer LS5 fluorescence
30 I
0 1 0 20 30
time in minutes
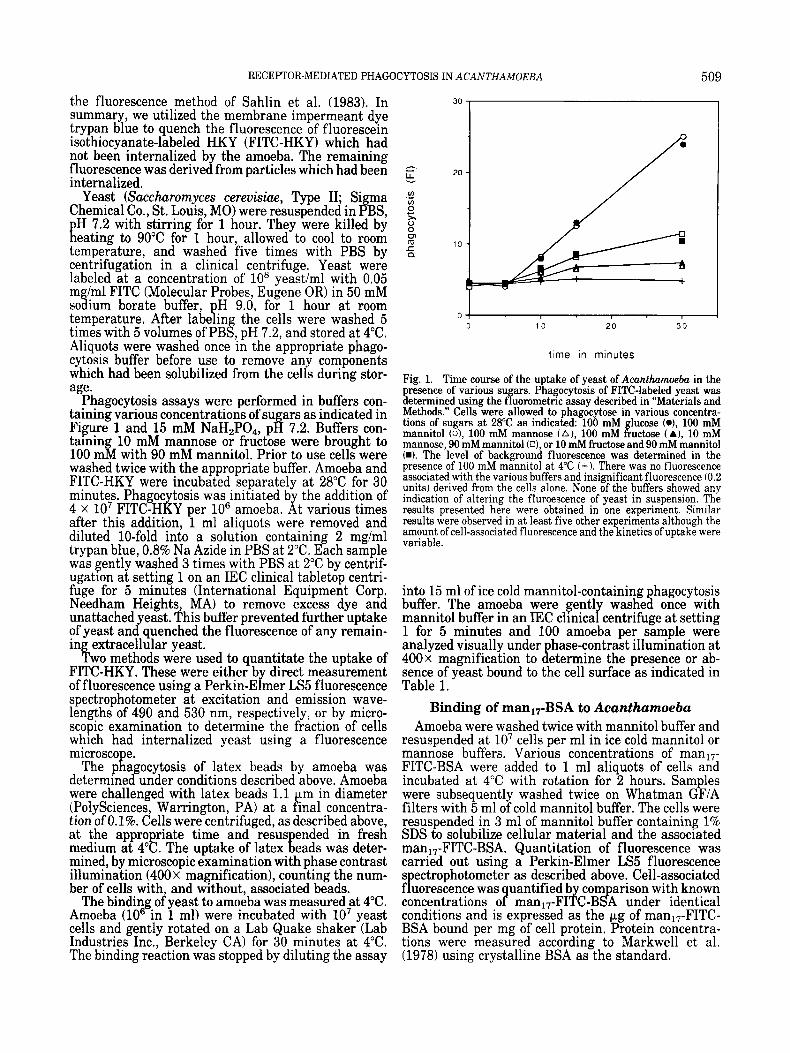
Fig. 1. Time course of the uptake of yeast of Acanthmoeba in the presence of various su ars Phagocytosis of FITC-labeled yeast was determined using the ffuoiometric assay described in “Materials and Methods.” Cells were allowed to phagocytose in various concentra- tions of sugars at 28°C as indicated: 100 mM lucose (o), 100 mM mannitol (o), 100 mM mannose ( A ) , 100 mM ffuctose (A), 10 mM mannose, 90 mM mannitol (Q, or 10 mM fructose and 90 mM mannitol W. The level of background fluorescence was determined in the presence of 100 mM mannitol at 4°C (+ 1. There was no fluorescence associated with the various buffers and insignificant fluorescence (0.2 units) derived from the cells alone. None of the buffers showed any indication of altering the fluroescence of yeast in suspension. The results presented here were obtained in one experiment. Similar results were observed in at least five other experiments although the amount of cell-associated fluorescence and the kinetics of uptake were variable.
into 15 ml of ice cold mannitol-containing phagocytosis buffer. The amoeba were gently washed once with mannitol buffer in an IEC clinical centrifuge at setting 1 for 5 minutes and 100 amoeba per sample were analyzed visually under hase-contrast illumination at
sence of yeast bound to the cell surface as indicated in Table 1.
Binding of man,,-BSA to Acanthamoeba Amoeba were washed twice with mannitol buffer and
resuspended at lo7 cells per ml in ice cold mannitol or mannose buffers. Various concentrations of manl7- FITC-BSA were added to 1 ml aliquots of cells and incubated at 4°C with rotation for 2 hours. Samples were subsequently washed twice on Whatman GF/A filters with 5 ml of cold mannitol buffer. The cells were resuspended in 3 ml of mannitol buffer containing 1% SDS to solubilize cellular material and the associated man,,-FITC-BSA. Quantitation of fluorescence was carried out using a Perkin-Elmer LS5 fluorescence s ectrophotometer as described above. Cell-associated
uorescence was uantified by comparison with known concentrations o man,,-FITC-BSA under identical conditions and is expressed as the p,g of man,,-FITC- BSA bound per mg of cell protein. Protein concentra- tions were measured according to Markwell et al. (1978) using crystalline BSA as the standard.
4 0 0 ~ magnification to b) etermine the presence or ab-
? fr

510 ALLEN AND DAWIDOWICZ
TABLE 1. Effect of sugars on the binding of yeast to Acanthamoebal
~ ~~~~
Sugar (100 mM) Glucose 24 Mannitol 25 L(-)-mannose 25 Inositol 23 D( +)-fucose 24 L( -)-fucose 22 N-acetyl glucosamine 22 D(+)-mannose 4 D(-)-fructose 5
‘56 cells with yeast bound
’Cells were incubated in 100 mM sugar at 4OC for30 minutes with gentle stirring as indicated in “Materials and Methods.” Over 100 cells from each sample were analyzed for the presence or absence of bound yeast.
Uptake and de radation of man-BSA neog !? ycoproteins
The interaction of amoeba with man-BSA neoglyco- proteins was examined to determine if Acanthamoeba could specifically internalize and degrade mannosy- lated protein. Man,,-FITC-BSA was incubated with amoeba in the presence of increasing concentrations of non-fluorescent mana8-BSA. The release of trichloro- acetic acid-soluble fluorescein was taken as an indica- tor of endocytosis and transport of the neoglycoprotein to lysosomes. TRITC-conjugated BSA was included as an indicator of uptake due to pinocytosis. The amount of TCA soluble fluorescence was determined spectro- photometrically.
Cells were washed twice with mannitol hagoc tosis buffer and resuspended at 106/ml in this i 9 uffer 28%) to ether with 20 pg/ml TRITC-BSA, 50 pglml man,,- FkC-SSA, and various concentrations of manZ8-BSA as indicated in Figure 4. At various times after the addition of the cells to the reaction mix, 1 ml aliquots were mixed with 200 ~1 of 50% TCA, vortexed, and incubated for at least 15 minutes at room temperature. The TCA-extracted suspensions were centrifuged for 5 minutes in a microfuge at 14,000 rpm. Five hundred microliters of the supernatant was collected, diluted with 2.0 ml of mannitol buffer, and adjusted to pH 9-10 with 150 p1 of 3 M NaOH. This titration was necessary to prevent the quenching of fluorescein fluorescence at acidic pH.
RESULTS We have measured the effects of various sugars on
the binding and phagocytosis of yeast by Acan- thamoeba. In order to com are these data we have
HKY as that which occurs in 100 mM glucose, the concentration of this sugar present in growth medium. The phagocytosis of yeast by Acanthumoeba at 28°C is inhibited by the free sugars D(+)-mannose and D(-1- fructose (Fig. 1) in a manner similar to the observations of Brown et al. (1975) on the effects of sugars on the phagocytosis of erythrocytes by this amoeba. We have found that this inhibition is stereospecific, as L(-1- mannose has no inhibitory effect even at a concentra- tion of 100 mM (data not shown). The effect of both mannose and fructose on phagocytosis is dose depen- dent, with an ICs0 for both sugars of approximately 10 mM. Maximal inhibition of phagocytosis occurs be-
defined the control level of p R agocytosis and binding of
tween 30 and 50 mM free sugar (Fig. 2A). This inhibi- tion appears to occur at the level of binding to the plasma membrane since the inhibition of binding of yeast to amoeba at 4°C has the same concentration dependence (Fig. 2B) as the inhibition of phagocytosis by these sugars.
Whereas the binding or unopsonized yeast to mam- malian macrophages is inhibited by mannose, fucose, and N-acetyl glucosamine (She herd et al., 1982), of
on the yeast-amoeba interaction (Table 1). The binding and hagocytosis of yeast by Acanthamoeba is also una b? ected by the related compound mannitol.
The inhibition of phagocytosis and binding of yeast to amoeba by these sugars appears to be due to a specific, competitive interaction between these sugars and the element on the cell surface responsible for binding yeast. The u take of latex beads by Acantharnoeba is
which maximally inhibit the uptake of yeast (data not shown).
The high concentration of sugar necessary to inhibit phagocytosis suggested to us that the interaction of mannose, or fructose, with the putative pha ocytosis
ing the apparent affinity of a ligand for its binding site is to make that ligand olyvalent (Hoppe and Lee, 1983;
that neoglyco roteins rich in mannose could be a more effective inhi itor of hagocytosis due to their in-
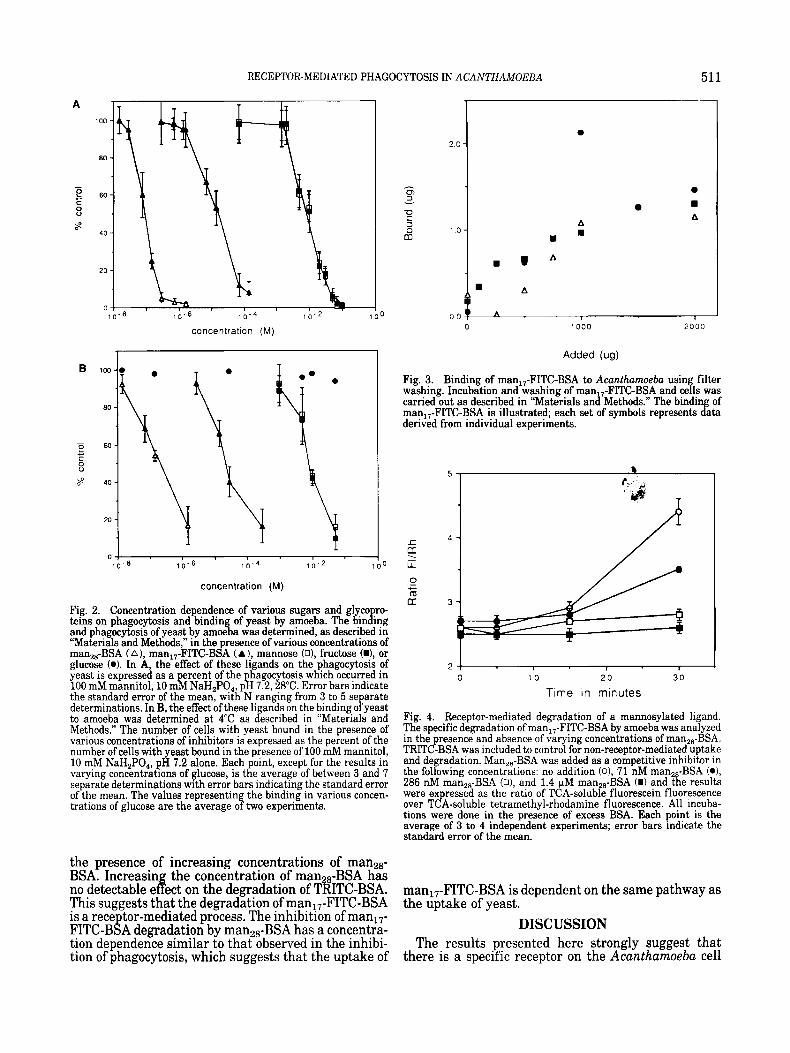
gands had been used to characterize a mannoseifucose receptor from mammalian macrophages (Shepherd et al., 1982, Hoppe and Lee, 1983; Ohsumi and Lee, 1987). Our hypothesis is validated by the data shown in Figure 2 which demonstrates that man2,-BSA and man,,-FITC-BSA are more effective inhibitors of bind- in and hagocytosis than the free sugar by several or d P ers o magnitude. The ICs0 for manz8-BSA and man17-FITC-BSA are approximately 100 nM and 10 pM, respectively.
We utilized the fluorescent moiety of the manl7- FITC-BSA to determine if this inhibitor bound to amoeba. The results illustrated in Figure 3 indicate that there is a saturable binding site for man,,-FITC- BSA on the amoeba. Bindin to this site can be
free sugars mannose or fructose. Scatchard (1949) analysis of the saturation curve su gested that the
single class of non-interacting sites (data not shown). As estimated from the saturation curve, the level of half-maximal binding is 10 pM (750 pg/ml) and the amount of ligand bound at saturation sug ests that
per cell. Acanthamoeba is capable of the receptor-mediated
degradation of the man17-FITC-BSA ligand. As indi- cated in Figure 4 we measured the production of fluorescent, TCA-soluble de adation products of
increasing concentrations of non-fluorescent man 8- BSA. There is an apparent competitive inhibition of tke degradation of the mannosylated manI7-FITC-BSA in
these compounds only mannose ‘R as a detectable effect
not inhibite CQ by concentrations of mannose or fructose
receptor was of very low affinity. One means o f increas-
Klassen et al., 1988; P ungi et al., 1988). We reasoned
creased valency. ‘ R Furt ermore, these polyvalent li-
competed by the presence of hig a concentrations of the
binding of this ligand to cells cannot % e described by a
there are approximately lo6 receptors for t f is ligand
manl7-FITC-BSA and TRITC- !iY SA in the presence of
RECEPTOR-MEDIATED PHAGOCYTOSIS IN ACANTHAMOEBA 511
A 100
80
- - 9 60 c 0
40 8
20
0 1 0 - 8
concentration (M)
B
2.0
-z ; 3
1 0
m u A
A
A
I
1000 2 0 0 0
0 I b.6 I b Z 1 b o
concentration (M)
Fig. 2. Concentration dependence of various sugars and glyco ro teins on phagocytosis and binding of yeast by amoeba. The bining and phagocytosis of yeast by amoeba was determined, as described in “Materials and Methods,” in the presence of various concentrations of man,,-BSA (A) , man,,-FITC-BSA (A ), mannose (O), fructose (W, or glucose (0). In A, the effect of these ligands on the phagocytosis of yeast is expressed as a ercent of the hagocytosis which occurred in 100 mM mannitol, 10 m b NaH,PO,, p h 7.2,28”C. Error bars indicate the standard error of the mean, with N ranging from 3 to 5 se arate determinations. In B, the effect of these ligands on the binding o f yeast to amoeba was determined at 4°C as described in “Materials and Methods.” The number of cells with yeast bound in the presence of various concentrations of inhibitors is expressed as the percent of the number of cells with yeast bound in the presence of 100 mM rnannitol, 10 rnM NaH,PO,, pH 7.2 alone. Each point, except for the results in varying concentrations of glucose, is the average of between 3 and 7 separate determinations with error bars indicating the standard error of the mean. The values representing the binding in various concen- trations of glucose are the average of two experiments.
the presence of increasing concentrations of manz8- BSA. Increasin the concentration of manz6-BSA has
This suggests that the degradation of man,,-FITC-BSA is a rece tor-mediated process. The inhibition of manl7- FITC-B8A degradation by rnana6-BSA has a concentra- tion dependence similar to that observed in the inhibi- tion of phagocytosis, which suggests that the uptake of
no detectable e d ect on the degradation of TRITC-BSA.
Added (us)
Fig. 3. Binding of man,,-FITC-BSA to Acanthumoeba usin filter washing. Incubation and washing of man,,-FITC-BSA and cefls was carried out as described in “Materials and Methods.” The binding of man,,-FITC-BSA is illustrated; each set of symbols represents data derived from individual experiments.
2 ! 1 I
0 1 0 20 30
Time in minutes
Fig. 4. Receptor-mediated degradation of a mannosylated ligand. The specific degradation of man,,-FITC-BSA by amoeba was analyzed in the presence and absence of varying concentrations of man,,-BSA. TRITC-BSA was included to control for non-receptor-mediated uptake and degradation. Man,,-BSA was added as a competitive inhibitor in the following concentrations: no addition (o), 71 nM man,,-BSA (o), 286 nM man,,-BSA (o), and 1.4 FM man,,-BSA (I) and the results were expressed as the ratio of TCA-soluble fluorescein fluorescence over TCA-soluble tetrarnethyl-rhodamine fluorescence. All incuba- tions were done in the presence of excess BSA. Each point is the average of 3 to 4 independent experiments; error bars indicate the standard error of the mean.
man17-FITC-BSA is dependent on the same pathway as the uptake of yeast.
DISCUSSION The results presented here strongly suggest that
there is a specific receptor on the Acanthumoeba cell

512 ALLEN AND DAWIDOWICZ
surface, inhibitable by millimolar concentrations of the free sugars mannose and fructose, which is responsible for the binding of mannose residues on the yeast cell wall as the initial ste in hagocytosis. This receptor is
glycoproteins man,,-BSA and man,,-FITC-BSA. We have used these polyvalent mannosylated ligands to demonstrate that this amoeba is capable of the specific binding, endocytosis, and degradation of these com- pounds. We argue that these molecules are acting as soluble analogs of the yeast cell wall and as such are useful in attempting to understand the molecular events of phagocytosis as demonstrated in the following pa er (Allen and Dawidowicz, 1990).
geveral criteria must be met in order to define an activity as a receptor-mediated process. A binding site must be specific, saturable, and have cellular function in order to be called a receptor. We have provided evidence to indicate that Acantharnoeba has a receptor for mannose-containing elements found in the yeast cell wall. The inhibition of binding and phagocytosis of yeast by mannose, or fructose, is concentration depen- dent and stereospecific. This inhibition is limited to mannose-containing particles; none of the sugars or
lyco roteins used in this work prevented the uptake of fatexieads by this amoeba. These results indicate that there is a receptor involved in the binding of yeast to amoeba which can be blocked by mannose or fructose.
Our results on the inhibition of binding and pha o-
activity mediates the initial interaction between these two organisms. However, we were not able to measure the binding of mannose or fructose, to either yeast or amoeba. The inability to measure binding is likely to be due to the ap arent low affinity of this receptor for free
in the inhibition of phagocytosis. We have used mannosylated neoglyco- proteins as tools to determine if the yeast-amoeba interaction was receptor mediated and to determine on which of the cell types used in these experiments it resided. We observed that the neoglycoproteins are much more potent inhibitors of phagocytosis than the free sugar with a concentration dependence of inhibi- tion directly related to the degree of mannosylation. Using man17-FITC-BSA we were able to demonstrate that there is a binding site on the amoeba for this molecule which can be competed with high concentra- tions of either mannose and fructose. The a parent Kd for this interaction is approximately 10 pIbf similar to the half maximal inhibitory concentration as assayed both in the binding and phagocytosis of yeast by amoeba. The apparent affinity of the receptor for manl FITC-BSA is low when compared with the re- or te l affinities of many other receptors for their
Egands, which are often in the nanomolar range (see Forgac, 1988, for a comparison of commonly studied receptor-ligand systems). However, it should be noted that the effective concentration of mannose molecules on insoluble yeast articles is like1 to be significantly higher than that ofman17-FITC-BiA. Scatchard (1949) analysis suggests that the interactions of this ligand with cells are complex and cannot be described by a model based on a single class of non-interacting sites (data not shown). After binding, the amoeba is capable
much more effective P P y b ocked by the high mannose
cytosis of yeast by amoeba suggested that a lectin-11 a e
sugars, as in B icated by their high
of endocytosis and degradation of this ligand, which suggests that the ligand has been delivered to a pro- teinase-rich compartment such as the lysosome. From these observations we conclude that Acantharnoeba have a receptor which recognizes mannose, and is capable of the internalization of molecules or particles rich in this element.
The Acanthamoeba mannose receptor has many of the characteristics of previously described phagocytic receptors. All these receptors have a higher apparent affinity for multivalent ligands (Hoppe and Lee, 1983; Jungi et al., 1988; Klaassen et al., 1988). The Fc and complement receptors of mammalian phagocytes are dependent on the serum-mediated opsonization of the particle, while the mammalian mannose/fucose recep- tor is capable of binding and initiating the internaliza- tion of many types of unopsonized particles. Both the mammalian mannoseifucose receptor and the Acan- thamoeba mannose receptor appear to be involved in the phagocytosis of micro-organisms (Speert et al., 1988; Brown et al., 1975). The mannose receptor of Acanthamoeba, like the mammalian Fc and mannose- fucose rece tors, is capable of the uptake and degrada-
al., 1983; Stahl et a f , 1980). The ability to bind and internalize elements rich in
mannose would appear to be ideal for this amoeba. The lycoproteins of many micro-organisms consist pre-
fominately of long-chain and branched polymers of mannose in various linkages. The structure of yeast cell wall mannans have been well characterized (see Naka- jima and Ballou, 1974) and consist predominately of mannose polymers. The glycoproteins of Dictyostelium discoidiurn also consist predominately of com lex poly- mers rich in mannose (Kornfeld and Kornfe 7 d, 1985). The inhibition of the phagocytosis of erythrocytes by mannose and fructose (Brown et al., 1975) would argue that this receptor may be involved in the pathogenesis of acanthamoebic infections (see Sotelo, 1987, for review). By recognizing mannose residues on glycopro- teins, Acanthamoeba is ca able of binding, and phago-
encounter in its rambles through the soil, The ability of fructose to inhibit the binding of
mannose-rich elements to the amoeba sheds some insight on the molecular nature of receptor-ligand interaction. Fructose is not a normal constituent of glycoproteins and is unlikely to be found in high concentrations in the normal environment of this or-
anism. However, the ability of fructose to block the [inding of mannose rich molecules to this receptor suggests that the side groups of carbons 2-6 of the sugar rin are important in the recognition of ligand by
positions and inhibit the binding interaction with equal effectiveness. Glucose, which is identical with mannose except for an inverted orientation of the H and OH at the number 2 carbon, has no detectable effect on the binding interaction. This indicates that the orientation of the hydrogen and hydroxy groups at the two osition
of carbons 3-6 in binding to the receptor remains to be defined.
In summary, we have the characterized the nature of
tion of solu i! le analo s of relevant particles (Mellman et
cytosing, a large group o P organisms which it should
receptor. 5 4 annose and fructose are identical at these
is critical to the binding interaction. The invo P vement

RECEPTOR-MEDIATED PHAGOCYTOSIS IN ACANTHAMOEBA 513
the molecular interaction of Acanthamoeba with one particle it is capable of phagocytosing. Using soluble neo lycoproteins we have demonstrated the specific binfing, internalization, and degradation of these ligands in a receptor-mediated process. The knowledge derived from study of this system should allow one to determine characteristics common to both the proto- zoan and mammalian phagocytic process, and therefore likely to be necessary for the phagocytic process itself. One such characteristic, the receptor-stimulated deg- radation of phosphatidylinositol 4, 5-bisphosphate, is described in the following paper (Allen and Dawido- wicz, 1990).
ACKNOWLEDGMENTS This work was supported by NSF DCB 8503856 and
NIH training grant 5 T32 GM 07226.
LITERATURE CITED Allen, P.G., and Dawidowicz, E.A. (1990) Phagocytosis in Acan-
thamoebo: 11. Soluble and insoluble mannose-rich ligands stimulate phosphoinositide metabolism. J. Cell Physiol., 145:514-521.
Bowers. B.. and Olszewski. T.E. (1983) Acanthamoeba discriminates internally between digestible and indigestible particles. J. Cell. Biol., 97317-322.
Brown, R.C., Bass, H., and Coombs, J.P. (1975) Carbohydrate binding proteins involved in phagocytosis by Acanthamoeba. Nature, 254:434435.
Forgac, M. (1988) Receptor-mediated endocytosis. In: "he Liver: Biology and Pathobiology, Second Edition. Raven Press, New York, pp. 207-227.
Hoppe, C.A., and Lee, Y.C. (1983) The binding and processing of mannose-bovine serum albumin derivatives by rabbit alveolar macrophages. J. Biol. Chem., 258 14193-14199.
Jungi, T.-W., Lerch, P.G., Cachelin, A.B., and Morell, A. (1988) Monomeric and dimeric IgGl as probes for assessing high-affinity and low-affinity receptors for IgG on human monocyte-derived macrophages and on activated macrophages. Mol. Immunol.,
Klaassen, R.-J., Goldschmeding, R., Tetteroo, P.A., and Von dem Borne, A.E. (1988) The Fc valency of an immune complex is the decisive factor for binding to low-affinity Fc gamma receptors. Eur. J. Immunol., 18:1373-1377.
Korn, E.D. (1974) The isolation of the amoeba plasma membrane and the use of latex beads for the isolation of phagocytic vacuole membranes from amoeba including the culture techniques for amoeba. Methods Enzymol., 31:686699.
Kornfeld, R., and Kornfeld, S. (1985) Assembly of asparagine-linked oligosaccharides. Annu. Rev. Biochem., 54:631-664.
25~7 19-729.
Lennartz, M.R., Cole, F.S., and Stahl, P.D. (1989) Biosynthesis and processing of the mannose receptor in human macrophages. J. Biol. Chem., 264:2385-2390.
Markwell, M.A., Hass, S.M., Bieber, L.L., and Tolbert, N.E. (1978) A modification of the Lowry procedure to simplify protein determina- tion in membranes and lipoprotein samples. Anal. Biochem., 87:206-210.
Mellman, I. (1988) Relationships between structure and function in the Fc receptor family. Cum. Top. Microbiol. Immunol., 1:1625.
Mellman, I.S., Plutner, H., Steinman, R.M., Unkeless, J.C., and Cohn, Z.A. (1983) Internalization and degradation of macrophage Fc receptors during receptor-mediated phagocytosis. J . Cell Biol.,
Mills, J.T., Furlong, S.T. and Dawidowicz, E.A. (1984) Plasma mem- brane biogenesis in eukaryotic cells: Translocation of newly syn- thesized lipid. Proc. Natl. Acad. Sci. USA. , 81:1385-1388.
Nakajima, T., and Ballou, C.E. (1974) Characterization of the carbo- hydrate fragments obtained from Saccharomyces cereuisiae mannan by alkaline degradation. J. Biol. Chem., 249:7679-7684.
Newman, S.I., Becker, S., and Halme, J . (1985) Phagocytosis by receptors for C3b(CR1), iC3b(CR3) and IgG(Fc) on human perito- neal macrophages. J. Leukocyte Biol., 38:267-278.
Ohsumi, Y., and Lee, Y.C. (1987) Mannose-receptor ligands stimulate secretion of lvsosomal enzvmes from rabbit alveolar macroohaees.
961887-895.
L -
J. Biol. C h e k , 262:7955-?962. Preston, T.M., and King, C.A. (1984) Binding sites for bacterial
flagella a t the surface of the soil amoeba Acanthamoeba. J. Gen. Microbiol., 1301449-1458.
Rabinovitch, M., and destephano, M.J. (1971) Phagocytosis of eryth- rocytes by Acanthamoeba sp. Exp. Cell Res., 64:275-284.
Sahlin, S., Hed, J., and Rundquist, I. (1983) Differentiation between attached and ingested immune complexes by a fluorescence quench- ing cytofluorometric assay. J. Immunol. Methods, 601 15124.
Scatchard, G. (1949) The attractions of proteins for small molecules and ions. Ann. N.Y. Acad. Sci., 51:660-672.
Shepherd, V.L., Campbell, E.J., Senior, R.M., and Stahl, P.D. (1982) Characterization of the mannose/fucose receptor on human mono- nuclear phagocytes. J. Reticuloendothel. SOC., 32:423-431.
Sotelo. A.C. (1987) Nwzleria and Acanthamoeba. Free-living amebas pathogenic for man. Ferspect. Pediatr. Pathol., 1051-85.-
Speert, D. P., Wright, S.D., Silverstein, S.C., and Mah, B. (1988) *Functional characterization of macrophage receptors for in vitro Dhagocvtosis of unoosonized Pseudomonas aerueinosa. J. Clin. inve"st.: 82:872-879.
- Stahl, P., Schlesinger, P.H., Sigardson, E., Rodman, J.S., and Lee, Y.C.
(1980) Receptor-mediated pinocvtosis of mannose glycoconiugates bv macroohkes: characteiization and evidence fo; receutor ;ecv- ding. Celi, 15:207-215.
Ukkonen, P., Lewis, V., Marsh, M., Helenius, A., and Mellman, I. (1986) Transoort of macroohaee Fc receptors and Fc receptor-bound ligands to lykosomes. J. Ekp. Med., 163':952-971.
Ulsamer, A.G., Smith, F.R., and Korn, E.D. (1969) Lipids of Acan- thamoeba castellanii: composition and effects of hagocytosis on incorporation of radioactive precursors. J. Cell Bio!., 43:105-114.