pasteurella multocida biofilm formation, and the ... · glycogen exopolysaccharide (eps)....
TRANSCRIPT

Pasteurella multocida biofilm formation, and the interrelationship of P. multocida with
Histophilus somni in a polymicrobial biofilm during bovine respiratory disease
Briana Lynn Petruzzi
Dissertation submitted to the faculty of the Virginia Polytechnic Institute and State University in
partial fulfillment of the requirements for the degree of
Doctor of Philosophy
In
Biomedical and Veterinary Sciences
Thomas J. Inzana
Clayton Caswell
Kevin Edgar
F. William Pierson
December 11, 2017
Blacksburg, Virginia
Keywords: Biofilm, Pasteurella multocida, Histophilus somni, bovine respiratory disease
Copyright (2017)

Pasteurella multocida biofilm formation, and its importance in polymicrobial biofilms with
Histophilus somni during bovine respiratory disease
Briana Lynn Petruzzi
ABSTRACT
Pasteurella multocida is an important multi-host animal and zoonotic pathogen that is
capable of causing respiratory and multi-systemic diseases, bacteremia, and infections resulting
from bite wounds. The glycosaminoglycan capsule (CPS) of P. multocida is an essential
virulence factor, protecting the bacterium from host defenses. However, chronic infections such
as bovine respiratory disease (BRD) and avian cholera may be associated with biofilm formation.
Biofilm formation was inversely related to capsule production (determined by uronic acid
and N-acetylglucosamine assays), and was confirmed with capsule-deficient mutants of mucoid
strains. Capsule-deficient mutants formed biofilms with a larger biomass that was much thicker
and smoother than encapsulated strains.
Gas chromatography-mass spectrometry, nuclear magnetic resonance, and enzymatic
digestion demonstrated that the matrix material of the biofilm was composed predominately of a
glycogen exopolysaccharide (EPS). Therefore, CPS may interfere with biofilm formation by
blocking adherence to a surface or by preventing the EPS matrix to encase large numbers of
bacterial cells.
Chemical mutagenesis was performed on P. multocida strain P1059, resulting in isolation
of an acapsular mutant designated as P1059-R8. A uridyltransferase encoded by gene
P1059_01979 was mutated in such a way that a polar amino acid was changed to a non-polar
amino acid near the active site. The protein product of P1059_01979 is important for the

biosynthesis of the CPS subunit N-acetylglucosamine. CPS quantification revealed that the
subunit glucuronic acid was produced in equal concentrations to the parent, but the CPS subunit
N-acetylglucosamine was not detected in the chemical mutant. Biofilm formation in the chemical
mutant was significantly higher than in WT P1059 and the capsule-deficient mutant. We
hypothesize that P1059_01979 is essential for CPS production in P. multocida serogroup A.
Histophilus somni and Pasteurella multocida cause bovine respiratory disease (BRD) and
systemic infections in cattle. Following respiratory infection of calves with H. somni, P.
multocida is also often isolated from the lower respiratory tract. Because H. somni normally
forms a biofilm during BRD, we suspected that P. multocida may co-exist with H. somni in a
polymicrobial biofilm. Interactions between the two species in the biofilm were characterized and
quantified by fluorescence in situ hybridization (FISH), and the biofilm matrix of each species
examined by fluorescently-tagged lectins (FTL), confocal scanning laser microscopy of in vitro
biofilms and bovine pulmonary tissue following dual H. somni and P. multocida infection. FISH
and FTL were used to show that P. multocida and H. somni were evenly distributed in the in
vitro biofilm, and both species contributed to the polymicrobial biofilm matrix. COMSTAT z-
stack image analysis revealed that the average biomass and biofilm thickness of the individual
and polymicrobial biofilms were greatest when both species were present. Encapsulated P.
multocida isolates not capable of forming a biofilm still formed a polymicrobial biofilm with H.
somni, but only the EPS of H. somni could be detected by FTL staining of bovine tissues from
which both species were isolated. Bacteria within a biofilm are more quiescent than during
planktonic growth and induce less of an inflammatory response, indicating encapsulated P.
multocida may take advantage of the H. somni biofilm to persist in the host during less severe,
but more chronic, BRD. These results may have important implications for the management of
BRD.

Acute avian cholera is associated with encapsulated P. multocida, while chronic and
asymptomatic cases of avian cholera are associated with acapsular P. multocida isolates. We
hypothesize that biofilm formation is present and an important factor for chronic and
asymptomatic avian cholera. Experimental infections of chickens with biofilm deficient P.
multocida strain WT X73, proficient biofilm forming P. multocida strain X73ΔhyaD, and
proficient biofilm forming clinical isolates 775 and 756 showed that virulence inversely
correlated with biofilm formation. Histopathological analysis showed that biofilm forming
isolates induced little inflammation in the lungs, heart, and liver, while biofilm deficient isolates
induced greater inflammation. Biofilm material was located in pulmonary tissues of chickens
diagnosed with chronic avian cholera using FTL staining.. Quantitative real-time PCR for
expression of cytokine genes in the spleens of infected chickens indicated that P. multocida
induced Th1 and Th17 immune responses during acute and chronic avian cholera. Chickens that
succumbed to acute avian cholera after experimental challenge with WT X73 had high levels of
INF-ƴ, IL-1β, IL-6, IL-12A, IL-22, IL-17A, and IL-17RA expression in the spleen compared to
all other experimental groups. Antibody titers were low, indicating that antibodies may be less
important in managing and clearing P. multocida infections.

Pasteurella multocida biofilm formation, and its importance in polymicrobial biofilms with
Histophilus somni during bovine respiratory disease
Briana Lynn Petruzzi
GENERAL AUDIENCE ABSTRACT
Pasteurella multocida is a zoonotic pathogen, which means it can be transferred from
animals to humans as part of the normal flora of many animals including household pets such as
cats and dogs, and agriculture species such as cattle. P. multocida is responsible for infected
animal bites, especially those resulting from household and large cats. Additionally, P. multocida
is responsible for several diseases of veterinary importance, including avian cholera and bovine
respiratory disease (BRD).
Capsule, composed of capsular polysaccharide (CPS), is an essential virulence factor for
P. multocida. Virulence factors are genetically encoded attributes that aid the bacteria in causing
an infection. Capsule covers the surface of bacterial cells, which allows P. multocida to survive
within the host and avoid detection by the immune system. The P. multocida capsular serogroup
A is composed of hyaluronic acid.
Biofilms are communities of bacteria that survive within a hydrated matrix composed of
polysaccharides, proteins, enzymes, antimicrobial compounds, extracellular DNA, and other
bacterial and host components. Biofilms can be compared to multicellular organs of eukaryotes.
While less complex, biofilms similarly regulate nutrients, water, composition, remove waste, and
perform other processes such as DNA transfer. Biofilms protect bacterial communities by
shielding them from the host immune response. Bacteria living in biofilms also grow slowly, and
as a result are protected from many antibiotic treatments. While biofilm formation has been

suggested for P. multocida, the biofilm has not yet been characterized. The work reported here
characterizes biofilm formation by P. multocida isolates of capsular serogroup A. Biofilms
formed by P. multocida were stained with fluorescently-tagged lectins, DNA stain, and other
fluorescent dyes, as well as crystal violet stain. Biofilms were imaged using several microscopy
techniques. Biofilm formation was prominent for serogroup A strains of P. multocida that were
acapsular. However, in the presence of CPS, biofilm formation was inhibited.
H. somni forms a biofilm during BRD that allows the bacterium to survive within the
heart and lungs of the bovine host. BRD is often caused by several different bacterial, viral, and
even parasitic microbes – resulting in a polymicrobial disease. Polymicrobial diseases are more
difficult to diagnose and treat, which is a challenge when trying to control this economically
important disease. Experimental infections of bovines with H. somni have resulted in
polymicrobial infections with P. multocida. We hypothesize that these two bacterial species may
form a mutualistic or commensalistic interaction together during BRD to improve the survival of
one or both species within the host. The polymicrobial biofilm was observed using fluorescent
microscopy techniques. We confirmed that H. somni and P. multocida form a polymicrobial
biofilm.
Avian cholera can be an acute, chronic, or asymptomatic disease that affects poultry
farms and migratory flocks around the world. The spread of P. multocida and avian cholera is
thought to occur through infected water, infected insects, and through other infected animals
surrounding water supplies such as deer, raccoon, and even fish. We hypothesize that P.
multocida can produce a biofilm and survive within the respiratory tract of birds for extended
periods of time, that biofilm formation is important for the establishment of chronic and
asymptomatic avian cholera, and that a biofilm assists in the spread of disease between flocks of
birds. Chickens were challenged in the respiratory tract with a highly encapsulated, poor biofilm
forming strain, or a prominent biofilm forming strain. After 7, 14, and 28 days chicken lungs

were examined to identify bacteria, biofilm material, and inflammation. Biofilm-forming P.
multocida strains were less virulent and caused less inflammation than non-biofilm forming P.
multocida strains. Biofilms were visible in the airways of pulmonary tissue by scanning electron
microscopy. Biofilm formation by P. multocida was observed within the pulmonary tissue of
chickens with chronic and acute avian cholera.

viii
To my parents Peter and Karen Esposito,
& my husband Dominic-
Who have always encouraged me to follow my dreams-
Wherever they may take me.

ACKNOWLEDGEMENTS
I would like to recognize and give special thanks to:
My advisor Thomas Inzana for his guidance and mentorship.
My graduate committee, including Clayton Caswell, Kevin Edgar, and F. William Pierson for
their guidance and support.
My lab mates, both past and present, for their daily guidance, technical support, and company.
The Biomedical and Veterinary Sciences graduate department- specifically the former graduate
coordinator, Becky Jones, for her invaluable support, and former Dean of research & graduate
studies Roger Avery.
Kathy Lowe for her assistance with electron microscopy
Kristi DeCourcy for her assistance with fluorescence and confocal scanning laser microscopy
Edward Swords for his assistance with biofilm growth and COMSTAT analysis
Virginia Buechner-Maxwell for her assistance with bovine experiments

TABLE OF CONTENTS
Abstract……………………………………………………………………………………ii
General Audience Abstract………………………………………………………………..v
Dedication……………………………………………………………………………….viii
Acknowledgements……………………………………………………………………….ix
Table of Contents………………………………………………………………………….x
List of Figures……………………………………………………………...…………...xviii
List of Tables……………………………………………………………..………………xx
Chapter 1: Introduction & Literature Review……………………………….…………1
1.1 Introduction to Pasteurella multocida……………………………………..……...2
1.2 Typing systems used to organize Pasteurella multocida…………………….........2
1.2.1 Serogroups based on capsular polysaccharides…………………………………...2
1.2.2 Serotypes based on lipopolysaccharides…………………………………………5
1.3 Virulence factors important for P. multocida pathogenesis……………………….5
1.3.1 Capsular polysaccharide…………………………………………………………..7
1.3.2 Lipopolysaccharide………………………………………………………………..7
1.3.3 Outer membrane and secreted proteins……………………………………………8
1.3.4 Invasion of host cells……………………………………………………………..12
1.3.5 Possible hemolytic and proteolytic secretions…………………………………....12
1.3.6 Biofilm formation………………………………………………………………...13

1.3.7 Regulation of virulence genes……………………………………………………14
1.4 Epidemiology…………………………………………………………………….14
1.4.1 Avian Cholera……………………………………………………………………15
1.4.2 Bovine respiratory disease complex……………………………………………..17
1.4.3 Hemorrhagic Septicemia………………………………………………………...18
1.4.4 Porcine Respiratory Disease……………………………………………………..19
1.4.5 Atrophic Rhinitis………………………………………………………………....20
1.4.6 Snuffles in Rabbits……………………………………………………………….21
1.4.7 Other P. multocida diseases……………………………………………………...21
1.5 Concluding Summary……………………………………………………………21
Chapter 2: Exopolysaccharide Production and Biofilm Formation by Histophilus
somni………………………………………………………………………………….….23
2.1 Histophilus somni biofilm formation and its relevance during infection………..24
2.1.1 An introduction to biofilm formation……………………………………………24
2.1.2 Polymicrobial relationships are common within biofilm………………………..24
2.1.3 Biofilm formation during respiratory infection and septicemia…………………25
2.2. Differences in biofilm structure between pathogenic and commensal isolates….25
2.2.1 Methods of studying biofilm formation reviewed……………………………….25
2.2.2 The H. somni biofilm life cycle………………………………………………….26
2.2.3 Comparison of biofilm formation between strains 2336 and 129Pt……...27

2.2.4 Gene products essential for biofilm formation…………………………………..28
2.3. Histophilus somni exopolysaccharide identification and production……………29
2.3.1 H. somni exopolysaccharide production…………………………………………29
2.3.2 Genes responsible for exopolysaccharide formation………………………….....31
2.3.3 Sialylation of the exopolysaccharide…………………………………………….32
2.3.4 Diagnostic application of the exopolysaccharide………………………....32
2.4 Concluding Summary…………………………………………………………….33
Chapter 3: Capsular Polysaccharide Interferes with Biofilm Formation by Pasteurella
multocida serogroup A…………………………………………………………….…….35
3.1 Abstract…………………………………………………………………………..36
3.2 Importance……………………………………………………………………….36
3.3 Introduction………………………………………………………………………37
3.4 Results……………………………………………………………………………39
3.4.1 Relationship between CPS production and biofilm formation by P. multocida....39
3.4.2 Chemical and genomic analysis of the matrix exopolysaccharide……………….46
3.4.3 Enzymatic treatment of biofilms…………………………………………………48
3.4.4 Scanning Electron Microscopy (SEM)…………………………………………..50
3.4.5 Confocal Laser Scanning Microscopy (CLSM)…………………………………50
3.4.6 qRT-PCR of putative EPS matrix genes…………………………………………52
3.5 Discussion…..……………………………………………………………………53

3.6 Materials and methods…………………………………………………………...58
3.6.1 Isolates and growth conditions…………………………………………………...58
3.6.2 Isolation of a biofilm-proficient P. multocida variant…………………...……….59
3.6.3 Construction of acapsular P. multocida mutants………………………………....59
3.6.4 RNA extraction, PCR, and qRT-PCR…………………………………………….61
3.6.5 Biofilm quantification…………………………………………………………….62
3.6.6 Capsule quantification…………………………………………………………....63
3.6.7 Purification of EPS from the biofilm……………………………………………..63
3.6.8 Chemical analysis of EPS………………………………………………………...64
3.6.9 LOS purification………………………………………………………………….64
3.6.10 Treatment of growth medium with hyaluronidase, α-amylase, proteinase K, or hyaluronic
acid……………………………………………………………………………...………...65
3.6.11 Bacterial hydrophobicity and auto-aggregation………………………………….65
3.6.12 SEM……………………………………………………………………………...65
3.6.13 CLSM…………………………………………………………………….……...66
3.6.14 Statistical Analysis………………………………………………………………66
3.7 Acknowledgements………...…………………………………………………....66
Chapter 4: Chemical mutagenesis of Pasteurella multocida P1059 reveals the importance of
P1059_01979 in capsular polysaccharide production and biofilm
formation………………………………………………………………………………..68

4.1 Abstract…………………………………………………………………………..69
4.2 Introduction……………………………………………………………………....69
4.3 Materials & methods…..….………………………………………...……………71
4.3.1 P. multocida and growth conditions………………………………………….….71
4.3.2 Isolation of a chemically-modified P. multocida mutant………………………...71
4.3.3 Genome sequencing and analysis of P1059-R8………………………………….71
4.3.4 Biofilm quantification……………………………………………………………72
4.3.5 Capsule quantification using uronic acid and N-acetyl-glucosamine chemical
assays……………………………………………………………………………….…….72
4.3.6 Bacterial hydrophobicity and auto-aggregation……………………………….....72
4.3.7 Scanning Electron Microscopy…………………………………………………..73
4.3.8 Confocal laser scanning microscopy…………………………………………….73
4.3.10 Statistical analysis………………………………………………………………..73
4.4 Results…..………………………………………………………………………..74
4.4.1 Isolation of a CPS-deficient, biofilm-proficient mutant …………………….…..74
4.4.2 Characterization of chemical mutant P1059-R8 through genome sequencing and
analysis………………………………………………………………………………...…74
4.4.3 Biofilm Quantification by crystal violet staining……………………………......78
4.4.4 Capsule quantification using uronic acid and N-acetyl-glucosamine chemical
assays……………………………………………………………………………………..79

4.4.5 Confocal Scanning Laser Microscopy of live/dead stained biofilms……………80
4.4.6 Scanning Electron Microscopy (SEM) of biofilms on glass coverslips…………82
4.4.7 Determination of hydrophobicity and auto-aggregation……………………........83
4.5 Discussion…..……………………………………………………………………83
Chapter 5: Polymicobial Interaction between Histophilus somni and Pasteurella multocida
during Biofilm Formation………………………………………………………...……..87
5.1 Abstract…...……………………………………………………………………...88
5.2 Introduction……...……………………………………………………………….88
5.3 Materials and methods……...……………………………………………………90
5.3.1 Bacterial growth………………………………………………………………….90
5.3.2 Fluorescent in situ hybridization…………………………………………………90
5.3.3 Fluorescently-tagged lectin staining of exopolysaccharide material…………….91
5.3.4 Polymerase Chain Reaction……………………………………………………...92
5.3.5 Enzyme-linked immunosorbent assay…………………………………………...92
5.3.6 Biofilm protein and carbohydrate concentrations………………………………..92
5.3.7 Auto-aggregation of single species and polymicrobial suspensions……………..93
5.3.8 Determination of polymicrobial biofilm formation in vivo……………………...93
5.3.9 Statistical Analysis……………………………………………………………....94
5.4 Results…..……………………………………………………………………….94
5.4.1 Fluorescent in situ hybridization (F.I.S.H.) using DNA-specific probes………..94

5.4.2 Fluorescently-tagged lectin staining of biofilm EPS……………………….……98
5.4.3 Concentrations of Protein and Carbohydrate in polymicrobial biofilms……….101
5.4.4 Auto-aggregation of polymicrobial cultures……………………………………103
5.4.5 Determination of polymicrobial biofilm formation in vivo……………………..104
5.4.6 Histological analysis of polymicrobial respiratory disease……………………..105
5.4.7 Enzyme-linked immunosorbent assay…………………………………………..105
5.5 DISCUSSION…………………………………………………………………..106
Chapter 6: Avian biofilm formation and immune response following experimental acute
and chronic avian cholera due to Pasteurella multocida……………………………..109
6.1 Abstract………………………………………………………………………....109
6.2 Introduction……………………………………………………………………..110
6.3 Methods…………………………………………………………………………112
6.3.1 Bacteria used and growth conditions……………...……………………………112
6.3.2 P. multocida-chicken challenge experiments …………..……………………...113
6.3.3 In vivo Histopathology (H&E stain)…………………………………………....114
6.3.4 Scanning Electron Microscopy of In vivo and biofilms………………………...115
6.3.5 Fluorescently-tagged lectin staining for biofilm exopolysaccharide…………...115
6.3.6 Enzyme-Linked Immunosorbent Assay (ELISA)………………………………115
6.3.7 Bacterial numbers in pulmonary tissue…..…………………………………......116
6.3.8 RNA extraction and qRT-PCR….……………………………………………...116

6.3.9 Statistical analyses…………………………………………………………...…117
6.4 Results…..………………………………………………………………………117
6.4.1 Respiratory infection: Pilot study………………………………………………117
6.4.2 Histopathology of chicken lungs, heart, and trachea...…………………………119
6.4.3 Scanning Electron Microscopy (SEM)…………………………………………121
6.4.4 Fluorescently-tagged lectin staining of biofilm exopolysaccharide……………122
6.4.5 Follow-up experimental challenge.…………………………………..…………123
6.4.6 Antibody response…………………...…………………………………………124
6.4.7 Detection of cytokines using qRT-PCR of spleen cells following challenge…..126
6.5 Discussion…..…………………………………………………………………..131
6.6 Conclusion……………………………………………………………………...134
6.7 Acknowledgements………………………………………………………….....135
References…………………...…………………………………………………………136

LIST OF FIGURES
Figure 1.1 Carbohydrate sequences of glycosaminoglycan chains using monosaccharide
symbols…………………………………………………………………………………..….3
Figure 1.2 Hyaluronic acid composition……………………………………………....4
Figure 1.3 Genetic organization of region 2 of the CPS biosynthetic loci……………5
Figure 1.4 Outer membrane and outer membrane-associated proteins of P.
multocida……………………………………………………………………………………9
Figure 1.5 Functional domains of dermonecrosis-inducing toxins………………….12
Figure 1.6 Pathology of atrophic rhinitis…………………………………………….20
Figure 2.1 Structure of the EPS of Histophilus somni strain 2336…………………..30
Figure 3.1 Correlation between CPS production and biofilm formation by P. multocida
clinical isolates and laboratory strains……………………………...…...……………...….45
Figure 3.2 Effect off hyaluronidase enzyme on biofilm formation during growth….46
Figure 3.3 Structural analysis of the P. multocida serogroup A EPS……………….47
Figure 3.4 Enzyme digestion of biofilm matrix……………………………………..49
Figure 3.5 SEM images of P. multocida biofilms after 48 hours of incubation on glass
coverslips...…………………………………………………………………………...…...50
Figure 3.6 CSLM z-stack showing live/dead staining of WT P1059 during biofilm
formation………………………………………………………………………………..…51
Figure 3.7 Cross section of the biofilm by CSLM…………………………………..52

Figure 3.8 Normalized fold-increase of genes significantly upregulated during biofilm
formation…………………………………………………………………………………..53
Figure 4.1 Quantification of biofilm matrix with crystal violet stain……………….79
Figure 4.2 Quantification of capsular glucuronic acid and N-acetyl-glucosamine….80
Figure 4.3 CSLM of mutant P1059-R8 biofilm……………………………………..81
Figure 4.4 SEM of biofilm matrix formed on glass coverslips……………………...83
Figure 5.1 Fluorescent in situ hybridization of polymicrobial and single species
biofilms………………………………………………………………………………..…..96
Figure 5.2 Increased biomass and thickness within the polymicrobial biofilm……..97
Figure 5.3 Fluorescently-tagged lectin staining of in vitro biofilm EPS……………99
Figure 5.4 Changes in carbohydrate concentration during polymicrobial biofilm
growth………………………………………………………………………………….....102
Figure 5.5 Changes in protein concentration during polymicrobial biofilm growth.102
Figure 5.6 Auto-aggregation of polymicrobial suspensions over 24 hours………...104
Figure 5.7 Fluorescently-tagged lectin staining of in vivo biofilm EPS…………....105
Figure 6.1 Pilot study survival curve……………………………………………….119
Figure 6.2 Gram stained tissue sections from pilot study…………………………..120
Figure 6.3 SEM of chicken lung after experimental infection……………………...122
Figure 6.4 Fluorescently-tagged lectin staining of in vivo biofilm EPS…………....123
Figure 6.5 Antibody response to experimental infection…………………………...126

LIST OF TABLES
Table 3.1 Laboratory strains and clinical isolates used…………………………….39
Table 3.2 Pasteurella multocida strain WT P1059 genes with amino acid identity to
glycogen synthesis proteins in related bacteria…………………………………………...48
Table 3.3 COMSTAT analysis results obtained from CSLM z-stack images……...51
Table 3.4 Primers used for qRT-PCR analysis……………………………………..62
Table 4.1 Genes within the P1059-R8 genome with a SNP resulting from chemical
mutagenesis…………………………………………………………………………….…74
Table 4.2 Putative malfunctioning proteins due to SNPs……………………...…..78
Table 4.3 COMSTAT analysis of z-stack images presented in Figure 4.2…….......82
Table 5.1 COMSTAT analysis of Fluorescent in situ hybridization polymicrobial and
single species biofilms…………………………………………………………….………95
Table 5.2 COMSTAT analysis of fluorescently-tagged lectin staining of in vitro biofilm
EPS…………………………………………………………………………………...…..101
Table 6.1 P. multocida strains and bacterial isolates used………………………...113
Table 6.2 Histopathology scoring of lung, heart, trachea of experimentally infected
chickens…………………………………………………………………………….…….121
Table 6.3 Percent bacterial recovery………………………………………………124
Table 6.4 Cytokine genes tested by qRT-PCR on infected spleen samples…….…128

Table 6.5 Relative fold increase of cytokine gene expression in spleens of chickens
experimentally infected with P. multocida……………………………………………..136

CHAPTER 1
INTRODUCTION & LITERATURE REVIEW

1.1 Introduction to Pasteurella multocida
Pasteurella multocida is a gram negative bacterium in the pasteurellaceae family. It is a potential
pathogen of most – if not all- animals, but is most commonly associated with domestic and agricultural mammals
and avian species 1. P. multocida is often found as part of the upper respiratory or gastrointestinal normal flora of
mammals 2-6 but not avian species 7, 8. There are 4 subspecies of P. multocida: multocida, gallicida, septica, and
tigris 9-11. The research reported here focuses primarily on P. multocida subspecies multocida, as it is the most
common subspecies responsible for serious infections.
Pasteurella multocida is zoonotic, and has been isolated from human cases of meningitis 12-14, systemic
infections 15, 16, and other infections including abscesses 17 of immunocompromised individuals 18, infants 19, and
healthy adults, most of which had been in contact with or bitten by cats or dogs 20. P. multocida is the primary
species isolated from infected cat bites (75% of reported cases) and dog bites (50% of reported cases) 2, 21, 22, and
is assumed to be present in most wild animal bite infections such as those from large, wild cats 23-25. P. multocida
has also been isolated from infected pig 26 and horse 27 bites.
Recently, P. multocida has been associated with nosocomial infections 28-30. In human cases, transmission
of the bacteria occurs through contact with animals. However, one recent case study reported post-operative
pasteurellosis in the sternum of a woman who had no previous contact with animals 31. The source of P.
multocida, in that particular case is unknown.
1.2 Typing systems used to organize Pasteurella multocida
1.2.1 Serogroups based on capsular polysaccharides
Each capsule type, known as serogroups A, B, D, E, and F, is relatively host and disease specific, for
reasons still unclear. The 5 capsular polysaccharides (CPS) vary in composition, but are all glycosaminoglycans
(GAGs), which are long, unbranched polysaccharides composed of a repeating unit. The repeating unit consists of

an amino sugar and uronic acid. However, there are exceptions. The type B capsule contains mostly mannose,
arabinose, and galactose, but is one of the least characterized capsules. Serogroup A capsule is composed of
hyaluronic acid, making it a very poor antigen 32. Hyaluronic acid has a repeating unit disaccharide of N-
acetylglucosamine and glucuronic acid. Xylan may also be present in the capsule of some strains 33. However, our
work suggests xylan may be present during biofilm formation. The serogroup F capsule is composed of
chondroitin, and is thought to be similar to serogroup A 34. Serogroup D contains mostly heparin, and is also
thought to be related to type A 32, 35. Serogroup D is almost exclusively associated with cases of atrophic rhinitis in
swine and snuffles in rabbits 36.
Figure 1.1: Carbohydrate sequences of glycosaminoglycan chains using monosaccharide symbols. (A)
Hyaluronan, (B) Chondroitin, (C) Dermatan, (D) Heparin and (E) Keratan. Possible sulfation presence and
location (2S, 4S or 6S) is indicated. Image used from MiliporeSIgma Glycobiology Analysis Manual, 2nd Edition,
Glycosaminoglycans and Proteoglycans. (http://www.sigmaaldrich.com/technical-
documents/articles/biology/glycobiology/glycosaminoglycans-and-proteoglycans.html)

Serogroup E is the least studied, due to its infrequency and isolation to a smaller geographical area;
serogroup E has only been associated with isolates from cases of hemorrhagic septicemia in African cattle 37.
Many of the functions of the genes in the serogroup E capsule locus remain unknown 38.
Figure 1.2: Hyaluronic acid composition. Alternating monosaccharide residues B-D-(1→3) glucuronic acid and
B-D-(1→4)-N-acetylglucosamine. Image used from MiliporeSIgma Glycobiology Analysis Manual, 2nd Edition,
Glycosaminoglycans and Proteoglycans. (http://www.sigmaaldrich.com/technical-
documents/articles/biology/glycobiology/glycosaminoglycans-and-proteoglycans.html)
The capsule loci for all 5 serogroups have been characterized. Genes found in the loci can be placed into 3
conserved regions. Region 1 contains four genes that encode for an ABC (ATP Binding Cassette) transporter,
designated either hexABCD or cexABCD 38, 39. Region 2 consists of synthesis genes specific for each capsule type.
In addition, a cryptic heparin synthase gene was identified outside of the capsule locus in serogroups A, D, and F
40. This may represent a possible modification system for the capsule composition but has not been investigated
further. Region 3 contains 2 genes, phyAB or lipAB 41. The fis regulatory protein affects all 3 regions of the type A

capsule loci and also regulates approximately 42 other genes, of which 16 are involved in cell surface expression
or synthesis of cell surface components 42.
Figure 1.3: Genetic organization of region 2 of the CPS biosynthetic loci. Serogroups represented: A,B,D,E,F.
CPS loci are shown from the following strains: P. multocida X73 (GenBank accession number AF067175),
M1404 (GenBank accession number AF169324), P934 (GenBank accession number AF302465), P1234
(GenBank accession number AF302466), and strain P4218 (GenBank accession number AF3302467). Numbers
above the boxes indicate the distance (in base pairs) between the last base of the preceding gene and the first base
of the next gene. Genes depicted by boxes above the line are transcribed in the left-to-right direction, while those
beneath the link are transcribed in the right-to-left direction. †, percent identity at amino acid level to A:1 CPS
biosynthetic locus; ‡, percent identity at amino acid level to B:2 CPS biosynthetic locus 43.
1.2.2. Serotypes based on lipopolysaccharides
Variations in lipopolysaccharide (LPS) are used to describe the P. multocida serotype, separating isolates
further into serotypes 1 – 16, as designed by Heddleston using a gel immunodiffusion test 44. The LPS serotypes

are based on outer core variations and two inner core structures 45. The P. multocida LPS molecule lacks an o-
antigen, resulting in what is more commonly referred to as a lipooligosaccharide (LOS). However, this term is
misleading and does not represent the high number of saccharide repeating units present. As a result, LOS is
typically not used to describe P. multocida LPS 45. Structures of most known P. multocida LPS serotypes (1, 2, 3,
5, 8, 9, 13, and 14) have been established 46-49.
Serotypes 1 and 3 are associated with avian cholera, while serotypes 2 and 5 are associated with
hemorrhagic septicemia in bovines. Serotypes 1 and 14 have a genetically similar LPS outer core locus. However,
serotype 14 LPS is truncated due to a genetic mutation in the phosphocholine biosynthesis gene pcgA 46.
Serotypes 2 and 5 also share an almost identical genetic locus. However, a phosphoethanolamine residue is absent
in serotype 2 due to a point mutation in the phosphoethanolamine transferase gene lpt_3 48. Variations between
LPSs of the same serotype is apparently common 38, and it is not always possible to differentiate serotypes 46.
1.3 Virulence factors important for P. multocida pathogenesis
Many of the virulence factors described in the following sections are poorly understood. Approximately
70 genes of unknown function were found to be upregulated during an infection 50. Strain Pm70 is estimated to
harbor 104 putative virulence genes, which accounts for about 7% of the transcribed genome 51.
Typing systems help to organize virulence factors 52, as many are serogroup or serotype specific. For
example, PMT (Pasteurella multocida toxin) is frequently expressed by serogroup D isolates, less commonly by
serogroup A isolates, and rarely by serogroups B, E, or F 53-55. Cases of avian cholera are associated with P.
multocida type A:1 and A:3, while cases of hemorrhagic septicemia are most commonly associated with P.
multocida types B:2, B:5, and E:5 56. However, virulence similarities based on current typing systems are
generalizations and not always observed 52.

1.3.1 Capsular polysaccharide
One of the most thoroughly studied virulence factors of P. multocida is its CPS. CPS is essential for
attachment and invasion of host tissues 57-59. However, the importance of CPS is controversial in the literature 38.
Discrepancies in the reported importance of CPS for virulence may be explained by the combination of strain,
experimental model, and time points used for each experimental observation.
P. multocida may downregulate capsule to present adhesins to attach to host tissues, then upregulate the
capsule again once in the blood stream to prevent phagocytosis 39. Capsule prevents phagocytosis of serogroup A
strains by bovine neutrophils; eleven percent of a capsulated serogroup A strain was phagocytized after 30
minutes. In comparison, 100% of non-encapsulated serogroup A isolates were phagocytized after 15 minutes,
62% of a non-encapsulated serogroup B isolate was phagocytized after 30 minutes 60. Despite its apparent
importance to colonization of the host, a vaccine against serogroup A CPS poorly protects against challenge 61, 62.
1.3.2 Lipopolysaccharide
Signs of disease noted during P. multocida infection have been attributed primarily to endotoxin, or the
lipid A component of LPS, which is the predominant fatty acid in the outer membrane 63. An injection of purified
LPS was enough to elicit signs of disease in buffalo identical to that of natural infection 64. Even small volumes of
LPS are able to elicit an innate immune response in the host, leading to a cytokine storm that results in immune
cell activation and often death of the host 65.
The LPS displayed by serotype 3 strain Pm70 may contain sialic acid and is structurally similar to an
antigen displayed on mammalian cells, which may contribute to its pathogenicity. However, not all serotype 3
strains contain sialic acid 45, 66. Vaccination with serotypes 1, 2, or 3 poorly protected against P. multocida
infections of other serotypes, and protection varied greatly among animal species and disease type 45, 67, 68.

1.3.3 Outer membrane and secreted proteins
The outer membrane (OM) is a structure only found on gram-negative bacteria. The OM acts as a
selective barrier, and is responsible for monitoring and limiting nutrient uptake, molecular transport, and is
responsible for interacting with the extracellular environment and host cells 56. Approximately 20-30% of
bacterial genomes encode membrane proteins, and 50% of the OM mass is protein 69. P. multocida OM proteins
can be categorized by their function: structural proteins, transport proteins, binding proteins, protein assembly
machinery, and membrane-associated enzymes. The functions of the OM proteins described here assist in the
survival of P. multocida in the host, and as a result contribute to its virulence 56.
Structural outer membrane proteins include OmpA, a small B-barrel membrane anchor that serves as the
physical link between the OM and peptidoglycan layer 70. In Escherichia coli, OmpA is reported to play a role in
adhesion and invasion of host cells, and is also involved in biofilm formation 71. P. multocida OmpA has been
demonstrated to act as an adhesion protein by bridging fibronectin and heparin on the surface of host cells 72.
OmpH, or Protein H, is an OM transporter protein that is considered a major protein of the OM envelope
73. OmpH is a channel forming transmembrane porin that is highly conserved among serotypes 74 and following
immunization protective against homologous and heterologous challenge 74-77, but may result in overstimulation
of the immune system, which can lead to adverse side effects 78.

Figure 1.4: Outer membrane and outer membrane-associated proteins of P. multocida. The outer membrane
is represented in green, the inner membrane is represented in blue, periplasm represented as the white space in
between. Structures are shown for proteins, if known, and computer generated models are representative of
proteins in which the structure is still unknown 56. Proteins represented in the figure that are discussed in this
introduction include Tad and associated proteins, FhaB, NanB, NanH, OmpA, HasR, TonB, ExbB, ExbD, and
PlpE. Figure originally published elsewhere: Hatfaludi T, Al-Hasani K, Boyce JD, Adler B: Outer membrane
proteins of Pasteurella multocida. Veterinary microbiology 2010, 144:1-17.
Structural protein PCP-Lpp (peptidoglycan-associated lipoprotein cross-reacting protein) shares 80%
similaritly with nontypable Haemophilus influenzae PCP, which is a target for host serum bactericidal activity 79.
P. multocida PCP-Lpp was expressed during an in vivo experimental infection, but was not protective 80. Omp16
is also highly similar to a nontypable H. influenzae surface protein, P6. Vaccination with P6 was protective in a
chinchilla otitis media model 81. In P. multocida, Omp16 elicited a high antibody response in turkeys, but was not
protective 82.

Lipoprotein-binding OM proteins PlpB and PlpE are periplasmic binding components of uptake systems
83-85. PlpB mutants are attenuated, but not protective 56, 86. PlpE mutants were attenuated, and chickens vaccinated
with recombinant PlpE were protected from heterologous challenge 86.
OM proteins important for binding include a type IV pili, PtfA, found only in serogroups A, B, and D 87.
P. multocida PtfA comprises 12 amino acids, which is an uncharacteristically long sequence for type IV pili. This
kind of type IV pili sequence has only been noted in one other bacterial species – H influenzae 88. Adhesion
ComE1 is a fibronectin binding protein, which binds soluble and immobilized forms of fibronectin and type 1
collagen 89. Adhesion proteins FhaB1 and FhaB2 are filamentous haemagglutinins. FhaB2 is essential for
virulence – mutants are fully attenuated in heterologous mouse 61 and turkey 90, 91 challenge models. The Tad
(tight adherence macromolecular transport system) locus is a subtype of the type II secretion system, and a
putative adhesion in P. multocida. The Tad locus encodes genes for the putative assembly of an Flp pili, which is
important for biofilm formation and host colonization 92, 93.
Iron acquisition is important for the survival of P. multocida and other pathogens within the host. Iron
uptake systems involve an outer membrane receptor, a periplasmic binding protein, and an inner membrane ABC
transporter. Energy needed to drive these systems is provided by TonB 94. The TonB complex also comprises the
proteins ExbB and ExbD, which are found on the inner membrane. The TonB complex works closely with iron
transport receptors in the outer membrane 95.
It is estimated that more than 2.5% of the P. multocida genome encodes for iron acquisition and
regulation, and many genes exist in duplicate or triplicate. Several iron related proteins are unique to Pasteurella,
and are not found in genera such as Escherichia and Haemophilus, which are thought to be most similar based on
evolutionary relatedness 51. Known P. multocida iron acquisition genes include HgbA (hemoglobin-binding
protein) 95, HgbB which is constitutively expressed 50, 96, the siderophore multocidin 97, bovine transferrin binding
protein TbpA 98, and HasR, which binds hemophores 99, 100. Other putative iron acquisition proteins exist but have
not been described 101.

The uptake of sialic acid from host tissues by OM-associated enzymes is important for virulence in
serogroups A and D. Sialic acid is incorporated into some LPS types, which aids in immune evasion 102. The
uptake system for sialic acid in P. multocida consists of two outer membrane-associated enzymes, NanB and
NanH 103; which are similar to sialic acid uptake proteins found on the surface of H. influenzae 102. Many putative
sialometabolism genes exist in the P. multocida type A genome, suggesting that this process is important for its
survival within the host 102.
One study determined outer membrane proteins to be poor vaccine candidates because the expression of
major and minor OMPs between P. multocida isolates varies greatly 104. However, 3 proteins have proven to be
protective against heterologous challenge: PlpE, OmpH, and FhaB2. Despite these findings, no commercial
vaccine is available 56.
While not considered an outer membrane protein, serogroup D and occasionally serotype A isolates
produce an exotoxin known as PMT (Pasteurella multocida toxin) 53, 55. PMT is only produced by isolates
carrying a lysogenic bacteriophage pathogenicity island, and has to date only been associated with cases of
atrophic rhinitis in swine 105-107. However, vaccination of rabbits with vaccines composed of PMT were protective
108-110. PMT is a dermonecrotic toxin, and functions similarly to the dermonecrotic toxin (DNT) produced by
Bordetella parapertussis, Bordetella bronchiseptica, and Bordetella pertussis 111, 112.

Figure 1.5: Functional domains of dermonecrosis-inducing toxins. Toxins represented: PMT, Escherichia coli
cytotoxic necrotizing factor 1 (CNF), and Bordetella pertussis dermonecrotic toxin (DNT). Toxins carry the N-
terminal receptor-binding domain (grey box) and the C-terminal active domain (white box) 113, 114.
1.3.4 Invasion of host cells
P. multocida is also able to invade host cells by an unknown mechanism. A serogroup A bovine
pneumonia isolate was able to invade non-phagocytic bovine aortic endothelial cells. P. multocida survived inside
the cells within tightly formed vacuoles, but did not multiply. Some bacteria were able to leave the vacuole and
reenter the extracellular environment 58. Another study found similar results with a serogroup A strain isolated
from avian cholera. The strain adhered to and invaded chicken embryo fibroblasts in the presence of a capsular
material. A non-encapsulated serogroup B strain showed decreased invasion compared to the encapsulated
serogroup A strain 39. P. multocida also binds both immobilized and soluble forms of fibronectin with the
assistance of OM proteins, which have been described in detail above 59.
1.3.5 Possible hemolytic and proteolytic secretions

Although P. multocida is characterized as being non-hemolytic and non-proteolytic, there are a few
studies that suggest otherwise. Under detergent conditions in vitro (0.5% Tween20), a heat and pH resistant
hemolytic extract was detected in all avian isolates observed 115. This extract was difficult to obtain in vitro, but
may be important during in vivo conditions. Avian isolates are also able to survive, and in many cases were able
to proliferate, in activated chicken serum 116. In some but not all strains observed, the presence of capsule was
correlated with increased serum resistance. Strains displayed different degrees of resistance in sera from different
animal species, including turkey, sheep, goat, pig, horse, cattle, guinea pig, and rabbit 116. The degree of resistance
varied greatly, indicating a species-specific complement-degrading enzyme may be present.
Metalloprotease extracts have been identified in the growth media, and are thought to be secreted by P.
multocida. These proteases degraded host IgG, and are highly similar to Actinobacillus pleuropneumoniae
proteases 117.
1.3.6 Biofilm formation
Interestingly, P. multocida isolates from bovine pneumonia and avian cholera have been shown to form a
biofilm in vitro, although this has not yet been observed in vivo 118. The same has been suspected for isolates of
swine pneumonia 119, suggesting that biofilm formation is not limited to serotype A. It is well known that biofilms
are important to other veterinary pathogens such as H. somni 120 in cases of bovine respiratory disease. Biofilm
formation may contribute to P. multocida survival within its host, potentially as a chronic infection.
Infectious biofilms are a complex matrix composed of bacterial cells, host cells, exopolysaccharide,
nucleic acid, trapped nutrients and water, enzymes, and protein. These communities of cells are comparable to
multicellular tissues. Bacterial cells show cooperation, circulation of fluids and nutrients, and are protected from
changes in the external environment 121. Within a biofilm, bacterial communities are protected by the matrix,
which is tightly adhered to a surface and provided with a constant source of nutrients 122. Outside of the host,
bacteria are likely part of a biofilm 123-125, which is considered a universal mode for survival in harsh

environments. While undergoing stress, bacteria will often mutate due to DNA uptake or phase variation to better
survive 126-131. These genetic mutations may contribute to the persistence of P. multocida in environmental and
host reservoirs essential to the spread of infections such as avian cholera.
1.3.7 Regulation of virulence genes
Small RNA molecules (sRNA) regulate bacterial protein expression by interacting with, and sometimes
modifying mRNA. These interactions and modifications can inhibit translation and alter the mRNA lifespan 132-
134. The actions of sRNAs are facilitated by the chaperone Hfq 135. Hfq monomers form a ring that binds to target
mRNA sequences and assists sRNAs in posttranscriptional regulation 136. In some cases, Hfq can interact directly
with mRNA in the absence of sRNA 137, 138. Recent research has identified Hfq as a global regulator for many
gram-negative pathogens 139, including P. multocida 140. A P. multocida hfq mutant expressed 128 genes
differently from the WT, which resulted in reduced CPS production and reduced virulence. Genes differentially
regulated included those for molecular transport, adhesion, and LPS biosynthesis 140.
Another regulator of P. multocida CPS production is Fis. Fis is important in the regulation of virulence
factors in other pathogenic bacteria, including pathogenic E. coli 141-145. P. multocida Fis shares 80% similarity
with E. coli Fis, which is a nucleoid-associated transcriptional regulator. In P. multocida, spontaneous acapsular
mutants had downregulated expression of capsular genes due to Fis. Thirty one genes were down-regulated in the
spontaneous acapsular strain including the gene that encodes OM protein PlpE, while 11 were upregulated. This
differential expression can be associated with the global transcriptional regulator, Fis 146.
Two additional putative transcriptional regulators- Pm0762 and Pm1231, have been identified in P.
multocida, but have not been characterized 147.
1.4 Epidemiology

Infections caused by P. multocida can be placed into one of two categories- the first describes P.
multocida as the primary causative agent, such as in avian cholera. The second describes cases in which P.
multocida is an opportunist; P. multocida takes advantage of a preexisting situation that is often poorly
understood or otherwise difficult to detect 148. An example of the latter is pasteurellosis in the lower respiratory
tract of immunocompromised bovines, as either a single species or polymicrobial infection, known as bovine
respiratory disease (BRD) complex.
1.4.1 Avian cholera
All avian species are considered to be susceptible to avian cholera 8, 149. Some species appear to be more
susceptible to disease than others, such as turkeys and waterfowl 148, 150. A study in Mallard ducks showed that the
fatal infectious dose for these birds is 12 P. multocida bacterial cells 151. Avian cholera is caused primarily by
serotype A strains of P. multocida, but type F strains are also commonly diagnosed, especially in turkeys 148.
Neither serotype commonly produces PMT in association with avian cholera. Avian cholera is a worldwide
concern, causing significant economic and ecological losses.
Avian cholera can be an asymptomatic, acute, peracute, or chronic infection 7, 152. Transmission from one
migratory flock to another causes globally widespread infections, which are damaging to ecological niches 153, 154
as well as poultry farms. It is common for thousands of birds to die off during a single outbreak, with the highest
reported death count close to 20,000 birds 150. P. multocida is transmitted from infected to healthy birds 155 in
water supplies such as troughs and ponds that infected and healthy birds share 156-160, and is also suspected to be
spread through rodent infestations 161 , arthropods 162, 163, contaminated soil 164, 165, non-fatal animal bites 166, 167,
and by a fecal-oral route 168. In addition, some healthy birds carry potentially pathogenic strains of P. multocida
asymptomatically 149, 169-171. This is especially common for flocks of geese, which are hypothesized to be the
primary carrier 149. Some of the largest reported outbreaks of avian cholera have happened in locations where
flocks of geese migrate. In a study performed during winter, researchers isolated pathogenic strains of P.

multocida A:1 from nasal, oral, and cloacal samples of otherwise healthy snow and Ross’s geese. Recovered
isolates varied in virulence when used to experimentally infect different species of birds 150.
During an active acute infection, P. multocida has been isolated from the lungs and air cavities, heart,
spleen, bone marrow, and waddle of turkeys and chicken, as well as from the small intestine, meningeal vesicles,
air spaces within the skull, kidneys, eyes, and synovial cavities 151, 172.
Acute avian cholera starts as an upper respiratory tract disease. Experimental challenges show that
invasion of the lung, trachea, and air sac tissue occurs before 1hr post intratracheal inoculation. After 1 hour,
invasion of the spleen and liver occurs. After 3 hours most other organs (listed above) harbor P. multocida, with
the infection lasting up to 14 days or possibly more 168. Pathological signs of the disease include caseous lung
lesions, necrosis (of the spleen, liver, and lungs), inflammation of affected organs, granulomas blocking air
cavities within the lungs, and hemorrhagic foci on various surfaces such as: epicardium, serosal surfaces, liver,
spleen, and gastrointestinal tract. Other uncommon signs of diseases are ocular lesions, arthritis, and synovitis168.
Acute avian cholera often results in death. Evidence of recovery is seen in fibrous lesions within the lungs
and other affected organs 168. Due to the rapid proliferation of the bacteria, antibody production does not aid in
controlling the infection. During the early stages of infection, the phagocytic response is either efficient in
controlling the infection, or not 151. However, this may be dependent on the virulence of the strain- since
infections often present differently 50. Interestingly, immunocompromised chickens suffered less severe symptoms
and a shorter span of infection (7 days on average) than immunocompetent chickens (infection lasting 10 days on
average) 168, suggesting that the cellular heterophilic response of avian species maximizes the pathogenicity of P.
multocida 173, 174. Most research to date has focused on highly virulent strains of P. multocida and acute avian
cholera outbreaks, and has largely underestimated the importance of chronic infections and asymptomatic
carriage. During chronic infections, P. multocida is likely surviving as part of a biofilm 123-125. Chronically
infected birds show few signs of disease, if any 149, 151, 168, 175.

Vaccines against avian cholera have been attempted since Pasteur, who after many passages was able to
obtain an attenuated strain. He used this strain to successfully provoke a protective immune response in birds 176,
likely by inducing a chronic, asymptomatic infection. More recent attempts at a commercial vaccine include
subunit vaccines of CPS, LPS, and/or OM protein candidates (refer to section 1.3).
1.4.2 Bovine respiratory disease complex
Bovine respiratory disease (BRD) complex, known as Shipping fever or bovine pneumonia, is common in
bovines that have been put under stress, such as being transported 177, 178. This disease is considered multifactorial
– predisposing factors include bacterial and/or viral and environmental factors 179, 180. The period of fasting
induced during travel reduces the cattle’s ability to respond to an invading pathogen. Transportation also exposes
cattle to exhaust fumes and to animals raised in different herds, which may also predispose cattle to contaminated
aerosols 181. Stresses such as weaning and viral infection commonly predispose young calves, which are more
susceptible to the disease 182. It is estimated that BRD costs the cattle industry more than $500 million a year,
making it one of the most important diseases to the industry. However, this estimation is subject to 5-year
fluctuations that can result in greater or lesser values 183. Losses come from increased preventative measures and
treatment costs, morbidity, mortality, and reduced carcass value due to treatment of disease 184, 185.
Signs of disease include depression, inappetence, cough, fever, and nasal discharge 87. Signs of BRD are
difficult to detect, and often not realized until postmortem observation of the animal 180. Histological signs of
disease include a fibrinous pleuritis, edema and abscesses of the lungs, necrosis of respiratory epithelium, and the
presence of exudate within the alveoli and bronchioles. The presence of neutrophils and macrophages within
alveolar discharge is evidence of P. multocida as the cause of the infection 87. Invasion of macrophages by P.
multocida may also be observed, probably in chronic cases 60.
Pasteurella multocida A:3 and D:3 are common causative agents of BRD 87. BRD has also been
associated with Mannheimia haemolytica, Mycoplasma spp., H. somni, and viral agents such as adenovirus,

parainfuenza-3 virus, bovine respiratory syncytial virus, and others 87, 120, 186, 187. P. multocida is more prevalent in
respiratory diseases of dairy cattle than beef cattle 188-190, while M. haemolytica is more prevalent in respiratory
disease of beef cattle 191. Respiratory pathogens, including P. multocida can be found in the upper respiratory tract
of both healthy and diseased bovines 186, suggesting that BRD is an opportunistic infection 180. BRD is often
described as polymicrobial; it is common to recover more than one causative agent from infected cattle 192-195.
This further complicates diagnosis and treatment of the disease 196-198.
All bacterial agents of BRD have been demonstrated to produce biofilms, which may be associated with
their pathogenic role during BRD 120, 199-201 (unpublished data). An experimental infection demonstrated the
potential importance of biofilms formed by H. somni during BRD within the cattle lung. After challenge with H.
somni, P. multocida was recovered from pharyngeal-nasal swabs, as well as from the lungs 120. Pre-challenge
screening did not detect P. multocida or other respiratory pathogens. Since H. somni biofilms were observed in
the lungs of experimentally infected calves from which P. multocida was also cultured, we hypothesize that P.
multocida and H. somni can occur in a polymicrobial biofilm together during BRD infection. It has been
suggested that biofilms actively attempt to capture other species in order to become polymicrobial 121, 202.
Polymicrobial biofilms are increased in genetic diversity, which can result in a greater advantage for survival to
both species; diverse bacterial populations are more likely resistant to a broader range of antibiotics, are more
metabolically diverse, and present a broader range of virulence factors 202-205.
Vaccines against BRD have largely focused on viral causative agents 206, 207. Commercial vaccines against
bacterial causative agents are lacking.
1.4.3 Hemorrhagic Septicemia
In bovines and ovines, P. multocida B:2 and E:2 are the primary causative agents of hemorrhagic
septicemia (HS). However, other serotypes have been associated with disease, including A:1, A:3, A:4, B:1, and

F3 1, 208-210. HS is cited as the most economically important disease in Africa and Asia, resulting in 100% mortality
in endemic locations 211, 212. HS has also been reported in deer populations in Denmark 213.
HS can be an acute or subacute septicemia resulting in elevated body temperature and ultimately death.
Death is usually sudden, and occurs within 24 hours in acute forms, after 2-3 days of incubation in sub-acute
forms, and in both cases usually with no observable clinical signs. However, visible signs can include a mild
bronchopneumonia, shallow respiration, and cyanosis of visible mucous membranes 1. Because of rapid disease
onset and high mortality rate, treatment of HS is usually not performed 214. Antibiotic treatments can be effective
at early stages of disease. However, it has become clear that providing antimicrobial treatments has led to multi-
drug resistant strains in endemic areas 208, 209. Vaccines are available in most countries but are ineffective-
outbreaks of HS still occur despite rigorous vaccination efforts, due to the short-lived protection offered by
current vaccines 208, 211.
1.4.4 Porcine Respiratory Disease
In swine, P. multocida is one of the causative agents of the most important disease affecting pork
production - porcine respiratory disease (PRD) complex. PRD is similar to BRD- it is a multifactorial disease
involving many primary and opportunistic causative agents, environmental factors, and potentially genetic factors
215, 216. It is not uncommon to isolate 3 or more causative agents from an outbreak, which together lead to a
chronic, difficult to treat respiratory disease. Causative agents include P. multocida, Mycoplasma hyopneumoniae,
Actinobacillus pleuropneumoniae, Haemophilus parasuis, Haemophilus suis, Streptococcus suis, Porcine
circovirus type 2, Pseudorabies virus, Porcine Reproductive and Respiratory Syndrome Virus (PRRSV), swine
influenza virus, and potentially others 215, 216. Because of the polymicrobial nature of PRD, control and treatment
of disease is problematic 217. Disease pathogenesis remains largely unknown, but is thought to be a combination of
pathogenic features provided by many bacterial and viral agents aiding in the breakdown of host immunity. Due

to the multifactorial nature of the disease, it has been difficult to recreate experimentally, leading to a severe lack
in knowledge about the disease 216.
1.4.5 Atrophic Rhinitis
Figure 1.6: Pathology of atriphic rhinitis. Sections of left (A) and right (B) ventral turbinate of pig, 13 days
after inoculation. Right turbinate reduced <75% the size of the left. Bar = 1mm218.
Atrophic rhinitis (AR) is an upper respiratory disease in swine, characterized by atrophy of the nasal
turbonate bones, which can result in a complete loss of turbonate structure and shortened, twisted snouts in
extreme cases. Signs of disease are attributed to PMT production by toxigenic isolates of P. mutocida 219 and
dermonecrotic toxin (DNT) produced by Bordetella bronchiseptica. Purified toxins from both bacterial species
were able to reproduce bone lesions similar to those reported in AR 220-222. Progressive forms of AR start as a mild
upper respiratory infection with B. bronchiseptica, followed by a secondary infection with P. multocida, but
progressive AR also describes a single agent infection with P. multocida. Non-progressive AR is an upper

respiratory infection with mild turbonate lesions and is due to B. bronchiseptica 114, 223-226. Vaccination of pigs
with a PMT subunit vaccine showed promising results 227, 228.
1.4.6 Snuffles in Rabbits
In rabbits, P. multocida causes the upper respiratory tract disease snuffles, characterized by acute or
chronic rhinitis and suppurative pneumonia in extreme cases. Signs of disease include rhinitis that may increase,
coughing, fever, mild or severe respiratory difficulty, and eventually death if left untreated 148. Transmission is
thought to occur through aerosols and contact. Genitalia may become infected, and venereal transmission can
occur 229. Snuffles is primarily associated with P. multocida serogroups A and D, but other bacterial species such
as B. bronchiseptica and Pseudomonas ssp. may be present in a polymicrobial infection with P. multocida 230, 231.
1.4.7 Other P. multocida diseases
Outbreaks of pneumonia caused by P. multocida have been recorded in the western United States, and
these have been attributed as the cause of the decline in numbers of wild bighorn sheep populations 232-234. In
addition, septicemia by P. multocida has been reported as the cause of death for elk populations in Wyoming 235.
P. multocida has also been isolated from individuals who suffered unusual deaths in black bears, seals, sea lions,
and bison across the United States 236-239.
1.5 Concluding Summary
This detailed introduction aims to summarize current knowledge about P. multocida and its role in
pathogenesis. The two different typing systems used to organize strains are through CPS and LPS. The best
classification includes both the serogroup (CPS) and serotype (LPS) separated by a colon. Since serogroups and

serotypes can be disease and/or location specific, this information can infer more information about the genetic
makeup and pathogenicity of a particular strain or isolate.
Known and putative virulence factors such as CPS, LPS, and OM proteins have been detailed in the
context of their importance to disease progression and current vaccine development efforts. Other putative
virulence factors such as biofilm formation, hemolysis, and proteolysis have yet to be fully characterized.
Diseases described in this introduction include those where P. multocida is the primary causative agent:
avian cholera, AR, snuffles, and also where P. multocida is an opportunistic pathogen: BRD, PRD. The diversity
of strains, diseases, and degree of virulence make commercial vaccines against pasteurellosis difficult to develop.
While some outer membrane proteins have shown to be protective, more research is needed. However, some
vaccines are available commercially with varying degrees of success.

CHAPTER 2
Exopolysaccharide Production and Biofilm Formation by Histophilus somni
This chapter was adapted from the published work:
Petruzzi, B., & Inzana, T. J. (2015). Exopolysaccharide Production and Biofilm Formation by Histophilus somni.
In Histophilus somni (pp. 149-160). Springer International Publishing.

2.1. Histophilus somni biofilm formation and its relevance during infection
2.1.1 An introduction to biofilm formation
Biofilms formed following an infection within the host are a complex matrix of bacterial cells, host cells,
exopolysaccharide, nucleic acid, trapped nutrients and water, enzymes, and protein. Due to advances in
microbiological research, it is now clear that most bacterial species have the capacity to form a biofilm.
Planktonically grown bacteria in rich media are poor models for studying disease, since this does not represent the
way bacteria most commonly interact within the host. During infection, bacteria are exposed to the host
environment, which harbors antibodies and phagocytic cells. Bacterial populations living within a biofilm are
tightly adhered to a surface, where they are provided with a constant source of nutrients and are protected by the
matrix. Once planktonic bacteria are shed from the biofilm, they can travel through the host to find a new surface
to colonize 122. Biofilm formation is an essential survival mechanism utilized by bacteria associated with chronic
or otherwise persistent infections 240, 241. Therefore, it is not surprising that H. somni is capable of forming a
biofilm and can do so in its only habitat: mucosal surfaces and systemic sites of cattle and sheep. In vitro, H.
somni always grows as a biofilm in stationary or in slowly rotating broth cultures with little or no headspace 242.
Biofilm formation by other bacterial pathogens of bovine respiratory disease (BRD), such as Mannheimia
haemolytica 199 and Pasteurella multocida 201, have also been described. However, H. somni biofilm formation is
more prominent in myocardial tissue than in pulmonary tissue, suggesting that biofilm formation can be
associated with any systemic infection as well as with BRD 120.
2.1.2 Polymicrobial relationships are common within biofilm
Biofilms in nature are often polymicrobial, and there may be advantages to bacteria in polymicrobial
relationships 202. Within a polymicrobial biofilm, the genetic diversity of the population is increased. In the same
way that planktonic growth does not often represent bacteria in their natural environment, single species biofilms
may also not be representative. Genetically diverse populations are more likely to be resistant to a broader range

of antibiotics, they may be more metabolically diverse, and be better protected from host defenses or the external
environment 202-205. BRD is often described as being a polymicrobial infection (Gagea et al., 2006), which further
suggests the potential importance of biofilm formation to disease progression 243. Isolation of more than one
causative bacterium or virus from a BRD infection is common, which may be evidence of a polymicrobial
etiology. For example, Pasteurella multocida and other BRD pathogens have been isolated from H. somni
pneumonia 244, 245 in calves that tested negative for respiratory pathogens prior to experimental challenge. Other
reports have indicated relationships between H. somni and Bovine Respiratory Syncytial virus (BRSV) during
BRD, and that this relationship correlates with disease severity 246-248.
2.1.3 Biofilm formation during respiratory infection and septicemia
Infections involving biofilms can occur throughout host tissues and have been identified in dental caries 249,
250, osteomyelitis 251, endocarditis 252, and otitis media 253-255, to name a few. Subclinical symptoms allow
infections to go undetected, which is common in bovine chronic pneumonia. Calves with subclinical infections
can be difficult to identify in herds, and can contribute to disease transmission 256, 257. Experimental infection of
calves with H. somni via the respiratory tract can result in biofilm formation within the myocardium and the
pulmonary tissues. The biofilm found in the myocardium is more prominent than the biofilm observed within the
lungs, which would correlate with the more prominent formation of biofilm during growth in tissues with reduced
levels of oxygen 120. Of significance is that P. multocida is often isolated with H. somni from BRD infections in
which biofilm is present 120, 245.
2.2. Differences in biofilm structure between pathogenic and commensal isolates
2.2.1 Methods of studying biofilm formation reviewed
H. somni appears to prefer the biofilm lifestyle, and forms a biofilm in vitro when grown under environmental
conditions with low oxygen availability (non-shaken cultures or in flasks filled with medium) or NaCl

concentrations above that of saline 258. Thus, stressful, poor growth conditions under which relatively few cells are
present results in the greatest amount of biofilm. For example, low oxygen conditions result in increased biofilm
growth, while aerobic conditions result in little to no biofilm formation. Although H. somni is a facultative
anaerobe, growth is poor anaerobically, resulting in EPS formation and a biofilm. Low oxygen conditions can be
simulated experimentally using a flask sealed and filled with broth medium, with minimal shaking (50 or less
rotations per minute) in order to reduce aeration and distribution of nutrients. H. somni biofilm growth is most
pronounced during the late stationary phase of growth, which represents a period of decreased availability of
nutrients. The addition of sodium chloride has also been shown to increase biofilm formation in H. somni.
Continuous flow cell systems provide an almost natural environment for biofilm formation to occur. In
such a flow system, there is a continuous supply of nutrients across the developing biofilm, allowing biofilms to
be sustained for longer periods of time. The resulting biofilms are most commonly analyzed by microscopy:
confocal scanning laser microscopy (CLSM), scanning electron microscopy (SEM) and transmission electron
microscopy (TEM). These microscopy techniques can be used to analyze the thickness, biomass, substratum
coverage, surface-area-to-volume ratio, architecture, viability, over-all morphology and other characteristics of
the biofilm that may be essential to their function. These techniques were used to show that the biofilm of strain
2336 was far more robust than the biofilm of strain 129PT under in vitro growth conditions.
2.2.2 The H. somni biofilm life cycle
Four distinct stages of H. somni biofilm growth occur in vitro over seven days when grown under continuous
flow conditions. Stage one involves attachment of the cells to a surface. During stage two growth/multiplication
occurs after approximately three-days. The third stage corresponds to maturation, which occurs in five-day old
biofilms. Stage four is the detachment of some planktonic cells, and occurs by the time biofilms are seven days
old. 259. Details of the stages of biofilm development in H. somni are described below:

Stage 1. Attachment: Sparse cell aggregates adhering to the abiotic or biotic surface are
typical during the attachment stage. Adhesion proteins (such as Fha and type IV pili) are
essential at this stage in the biofilm life cycle. The cell aggregates are composed of
predominantly live cells, as determined by CSLM live/dead staining and are not yet
surrounded by a substantial extracellular matrix.
Stage 2. Growth: Cell clusters of predominately live cells increase in size during this stage.
Biofilms are still relatively sparse. Thickness, biomass, and substratum coverage are still
increasing in size. The surface-area-to-volume ratio is high, as is to be expected in early
biofilms.
Stage 3. Maturation: During maturation, the biofilms reach their maximum thickness and
most of the cells within the biofilm are still living. The surface-area-to-volume ratio is low
and the architecture of the biofilm is most complex. In H. somni, maturation of biofilms for
all strains tested is approximately five days in vitro.
Stage 4. Detachment: At this time, large microcolonies start to disperse individual
planktonic cells in order to colonize a new location. The biofilm is composed primarily of
dead cells and there is an overall decrease in substratum coverage, mean thickness, and
biomass.
2.2.3 Comparison of biofilm formation between strains 2336 and 129Pt
During attachment, microcolonies of strain 2336 are present in greater quantity than strain 129Pt. The early
architecture of strain 2336 biofilm forms a structure of large microcolonies interconnected by an extracellular
matrix (ECM), which is not present in the biofilm of strain 129Pt. Strain 129Pt microcolonies are smaller and
more elongated, with little connective extracellular matrix (ECM) visible. The fully mature biofilms of strains

2336 and 129Pt display the most distinct differences. Strain 2336 has a mean thickness one hundred times greater
than the mean thickness of strain 129Pt. The mature biofilm of strain 2336 is a complex structure of
microcolonies with visible water channels seen in the ECM. The substratum coverage is decreased while the
surface-area-to-volume ratio, mean thickness, and mean biomass are increased. Mature biofilms formed by strain
129Pt display almost the opposite features: increased substratum coverage and a decrease in surface-area-to-
volume ratio. The biofilm architecture of 129Pt is composed of tower-shaped microcolonies intertwined with
strands of EPS. The roughness coefficient ‘r’ is significantly different between strains 2336 (r = 0.1) and 129Pt (r
= 2). However, during the detachment stage, both strains contain primarily dead cells in their matrices.
EPS production is significantly greater in pathogenic strain 2336 than commensal strain 129Pt, particularly if
sialic acid is added to the medium. This is expected, since the average biofilm thickness of strain 2336 is
approximately 100 times greater than that of strain 129Pt. Assays using crystal violet staining to measure the
amount of biofilm formed by various strains indicated that most isolates from systemic sites formed more biofilm
than isolates from genital sites, suggesting that biofilm formation is important in resistance to systemic host
defenses and correlated inversely with the roughness coefficient. Biofilms that have a roughness coefficient closer
to 0 are considered to be smoother. Smooth biofilms have fewer towers and gaps between microcolonies than
biofilms with a roughness coefficient closer to 2. Biofilms that are rougher have higher towers and spaces that are
devoid of biofilm matrix. The biofilm of strain 2336 is significantly smoother than the biofilm of strain 129Pt,
which may be related to the amount of EPS produced, as EPS functions to coat and connect the components of the
biofilm matrix. The bacterial populations within biofilms of both strains grow at an equal pace, indicating they are
both able to survive and thrive in this lifestyle 259.
2.2.4 Gene products essential for biofilm formation
Random mutagenesis of strain 2336 with the EZ::Tn5TM(KAN-2)Tnp TransposomeTM (Epicentre) has
identified genes whose products are important for biofilm formation, in addition to those already described for

EPS production and export 120. Mutations in IbpA, which has homology to the filamentous haemagglutinin (Fha)
of Bordetella pertussis 260, Haemophilus ducreyi 261, and others results in mutants that are deficient in biofilm
formation, indicating that Fha plays a role in biofilm formation. Since Fha is an important adhesin in those
bacteria in which it has been studied, it likely contributes to attachment during the initial stage of biofilm
formation, resulting in less biofilm or one that takes much longer to form. Furthermore, expression of fha in H.
somni is increased four-fold during biofilm formation in comparison to planktonic growth120. However, further
research is needed to confirm the role of Fha in H. somni virulence. A transposon mutation in the Major
Facilitator Superfamily 1 (MFS_1) protein increased biofilm formation by an unknown mechanism that warrants
further study 120.
2.3. Histophilus somni exopolysaccharide identification and production
2.3.1 H. somni exopolysaccharide production
Unlike many of the Pasteurellaceae, H. somni does not produce a capsular polysaccharide (CP). However, H.
somni does produce an exopolysaccharide (EPS), which is a major component of the biofilm matrix. A CP
remains closely associated with the individual bacterium and remains adherent to the cell surface through an outer
membrane-associated lipid anchor 262. In contrast, EPS is released from the bacterial cell, does not maintain close
contact with the cell surface, and therefore can envelop a large community of bacteria that share in the EPS, as
well as proteins and extracellular DNA, which together make up the architecture of the biofilm matrix material.
The exopolysaccharide produced by H. somni has been characterized as a highly branched α-(1 6)-pD-
mannan polymer. The comb-like branches are composed of mannopyranose units attached to the backbone at C-2.
C-3 branches extend from the C-2 substituted mannose residues, creating trisaccharide lateral chains. Occasional
galactose residues are also found at the non-reducing ends of the mannose side chains creating tetrasaccharide
branches Figure 2.1). As described for the lipooligosaccharide (LOS), the terminal galactose residues can be
sialylated in the presence of N-acetyl-5-neuraminic acid, making the biofilm thicker and more prominent 258.

Although the structure of the EPS is similar to yeast mannan, the H. somni EPS can be isolated from growth
medium lacking any yeast extract and the biofilm matrix specifically reacts with fluorescein-conjugated Moringa
M lectin (specific for mannose). Therefore, the EPS is not derived from the bacterial growth medium 258.
[→6-α-D-Manp-(1→6)-α-D-Manp-(1→6)-α-D-Manp-(1→6)-α-D-Manp-(1→]n
2 2 2
↑ ↑ ↑
1 1 1
α-D-Manp α-D-Manp α-D-Manp
2 2
↑ ↑
1 1
α-D-Manp α-D-Manp
3
↑
1
α-D-Manp
(β-D-Galp
Figure 2.1: Structure of the EPS of H. somni strain 2336 242. The H. somni EPS backbone is composed of pD-
mannan residues, with branching pD-mannose and pD-galactose residues.
Proteins are also present in the H. somni biofilm matrix, and may comprise approximately 50% of its dry
weight (unpublished data). One protein that has been identified in the biofilm matrix is Hsp60, and antibodies to
this protein are effective in inhibiting biofilm formation in vitro 263. The activity of Hsp60 in the biofilm may have
little to do with protein folding, but may be important for ligand binding 264.

2.3.2 Genes responsible for exopolysaccharide formation
The expression levels of putative EPS-associated genes during H. somni biofilm formation and planktonic
growth have been determined using quantitative real-time PCR (qRT-PCR). Genes with significant increases in
expression have been identified that encode for periplasmic substrate binding proteins and transmembrane
constituents of the ribose ABC transporter. These gene products represent domain II of the carbohydrate uptake
proteins responsible for transport of monosaccharides and forming the transport system responsible for high-
affinity ribose transport. Expression of another putative ABC transporter, rbs2a, is eight times greater during
biofilm growth than during planktonic growth 258.
Other genes that may be responsible for synthesis of the D-mannan polymer have been annotated as manB,
galU, and csrA. The galU gene is upregulated 7-fold during biofilm growth in comparison to planktonic growth.
GalU is a glucose-1-phosphate uridylyltransferase responsible for catalyzing the reversible production of UDP-
glucose. GalU is important for the synthesis of many carbohydrate components necessary for glycolipids and
proteoglycans, and has been associated with biofilm formation in Haemophilus parasuis 265. CsrA encodes a
putative carbon storage regulator 266, while manB encodes for a phosphomannomutase homolog responsible for
the conversion of mannose-6-phosphate to mannose-1-phosphate 258.
Another gene cluster of approximately 19 kb is predicted to encode for transport proteins,
glycosyltransferases, and proteins involved in polysaccharide biosynthesis that are also predicted to be important
for EPS and biofilm formation. This cluster includes pldB, ybhA, araD, sgbU, rmpA, xylB, rbs1C, rbs1A, rbs1B,
glsS, rbs2A, dctP, dctM and dctQ. Between the two loci mentioned, 14 genes are expressed at higher levels during
biofilm formation in pathogenic strain 2336. The genes encoding proteins important for the production of EPS are
also present in the genome of commensal strain 129Pt. However, only 5 genes are significantly expressed at
higher levels during biofilm formation in strain 129Pt 258. The biofilms formed by pathogenic strain 2336 are
thicker and display a different architecture than those formed by commensal strain 129Pt. The differences in gene
expression between strains 2336 and 129Pt during biofilm formation may account for their differing biofilm

phenotypes (described later in this chapter), but may be well suited for their respective ecological niches (strain
129Pt is restricted to the genital tract, whereas strain 2336 can disseminate and persist in systemic sites).
2.3.3 Sialylation of the exopolysaccharide
Terminal galactose residues of H. somni LOS, like the LOS of H. influenzae 267 are sialylated when the
bacteria are grown in the presence of N-acetyl-5-neuraminic acid (Neu5Ac) 268, 269. A small amount of galactose is
also present in the EPS, which can also be sialylated when sialic acid is present in the growth medium. When H.
somni is grown as a biofilm in the presence of Neu5Ac the density of the EPS increases, sialic acid can be
extracted from the EPS, and two amino sugars (N-acetylglucosamine and N-acetylgalactosamine) are detected in
the biofilm of strain 2336 258. These amino sugars are absent when sialic acid is not present. Strain 129Pt lacks
functional sialyltransferases and Neu5Ac-synthetase, and therefore even when grown in the presence of Neu5Ac
cannot sialylate its LOS 270 or EPS, and the amount of biofilm formed is not altered. In addition, expression of the
siaB gene in strain 2336, which encodes for an α-2, 3-sialyltransferase, is increased 15-fold during biofilm growth
in comparison to growth under planktonic conditions, further supporting a role for sialylation in development of a
mature biofilm 258. Sialylation also contributes to biofilm formation in other bacteria. Sialic acid, apparently on
the LOS, contributes to the integrity of biofilms of non-typable Haemophilus influenzae (NTHI), as NTHI is
unable to produce a typical EPS 271, 272.
2.3.4 Diagnostic application of the exopolysaccharide
The galactomannan polymer of H. somni EPS may not be an obvious antigenic marker as it is composed
entirely of simple sugar monomers. However, detection of anti-EPS antibodies by enzyme-linked immunosorbent
assay (ELISA) has been shown to be a useful diagnostic assay to differentiate infected from colonized animals 273.
A hindrance of current serological assays is the inability to distinguish between colonization with commensal

isolates, previously infected, or currently infected cattle. Current diagnostic procedures utilize isolation of H.
somni from a normally sterile site as indication of infection. The EPS diagnostic test is based on the principle that
pathogenic isolates form biofilms in vivo, but not when growing planktonically, which is how they are normally
grown in the laboratory. Furthermore, antibodies are more likely to be produced to foreign antigens in systemic
sites than to antigens in sites commonly colonized by many commensal bacteria, such as mucosal sites. Since
EPS is a major component of the biofilm, detection of antibodies to the EPS should indicate the presence of
infection rather than colonization. In addition, there is no cross-reactivity between polysaccharides of other
common bovine respiratory pathogens and the H. somni EPS. In a preliminary study, the EPS diagnostic test had a
specificity of 92.5% and a sensitivity of 90.5% 273. In cases where isolation of the bacteria is difficult or is too
time-consuming, the EPS ELISA may help to diagnose and quickly initiate corrective measures and prevent
future herd infections by H. somni.
2.4 Concluding Summary
The biofilm matrix of H. somni is a complex architecture that differs significantly between a pathogenic and a
commensal isolate, with the pathogenic isolate producing more EPS than the commensal isolate. However, there
is correlation between the amount of biofilm formed and the virulence of the strain. EPS produced during biofilm
formation is essential in bridging microcolonies within the biofilm and is responsible for the smooth biofilm
formation observed in pathogenic isolates. The EPS is composed of a D-mannan polymer, with occasional
galactose residues present on side chains, which is similar to the composition of yeast mannan. The EPS is also
the target of a recently developed diagnostic test that is capable of distinguishing between healthy and currently
infected bovines with promising results. This test is based on the production of antibodies to EPS in animals
during infection, but not during colonization. Two putative genetic loci that are likely responsible for EPS
production have been identified. However, whether both loci are necessary for production of the EPS has yet to be
determined. The expression of most of the genes in this locus is upregulated during biofilm growth in comparison

to planktonic growth. Differences in gene expression are evident during biofilm formation in strains 2336 and
129Pt, which explains the differences in EPS production and biofilm architecture observed in these two strains.
When grown in the presence of sialic acid, the EPS of strain 2336, but not the EPS of strain 129Pt, becomes
sialylated and the amino sugars N-acetylglucosamine and N-acetylgalactosamine can be detected within the EPS.
In vitro biofilm formation follows a typical 4-stage life cycle (attachment, growth, maturation and detachment) for
both strains studied.

CHAPTER 3
Capsular Polysaccharide Interferes with Biofilm Formation by Pasteurella multocida serogroup A
This chapter was adapted from the published work:
mBio
Briana Petruzzi1, Robert E. Briggs2, W. Edward Swords3,¶, Cristina De Castro4, Antonio Molinaro5, and Thomas J.
Inzana1,7*

3.1 ABSTRACT
Pasteurella multocida is an important multi-host animal and zoonotic pathogen that is capable of causing
respiratory and multi-systemic diseases, bacteremia, and infections resulting from bite wounds. The
glycosaminoglycan capsule of P. multocida is an essential virulence factor, protecting the bacterium from host
defenses. However, chronic infections (such as swine atrophic rhinitis and the carrier state in birds and other
animals) may be associated with biofilm formation, which has not been adequately investigated or characterized.
Biofilm formation (determined by crystal violet staining, and electron and confocal microscopy) by clinical
isolates was inversely related to capsule production (determined by uronic acid assay Congo Red uptake), and was
confirmed with capsule-deficient mutants of mucoid (highly encapsulated) strains. Capsule-deficient mutants
formed biofilms with more biomass that were much thicker and smoother than encapsulated strains. Passage of a
highly encapsulated, poor biofilm-forming strain under conditions that favored biofilm formation resulted in the
production of less CPS and a more robust biofilm, as did addition of hyaluronidase to the growth medium of all
strains tested. Gas chromatography-mass spectrometry, nuclear magnetic resonance, and enzymatic digestion
demonstrated that the matrix material of the biofilm was composed predominately of a glycogen
exopolysaccharide (EPS). However, a putative glycogen synthesis locus was not differentially regulated during
biofilm formation compared to when the bacteria were growing planktonically, as determined by quantitative
reverse transcriptase-polymerase chain reaction. Therefore, the negatively-charged capsule may interfere with
biofilm formation by blocking adherence to a surface or by preventing the EPS matrix from encasing large
numbers of bacterial cells. This is the first detailed description of biofilm formation and a glycogen EPS by P.
multocida.
3.2 IMPORTANCE
Pasteurella multocida is an important pathogen responsible for severe infections in food animals,
domestic and wild birds, pet animals, and humans. P. multocida was first isolated by Louis Pasteur in 1880, and

has been studied for over 130 years. However, aspects of its lifecycle have remained unknown. Although
formation of a biofilm by P. multocida has been proposed, this report is the first to characterize biofilm formation
by P. multocida. Of particular interest is that the biofilm matrix material contained a newly reported amylose-like
glycogen as the exopolysaccharide component, and that production of capsular polysaccharide (CPS) was
inversely related to biofilm formation. However, even highly mucoid, poor biofilm forming strains could form
abundant biofilms by loss of CPS or following in vitro passage under biofilm growth conditions. Therefore, the
carrier state or subclinical, chronic infections by P. multocida may result from down-regulation of CPS with
concomitant enhanced biofilm formation.
3.3 INTRODUCTION
Pasteurella multocida is a zoonotic 274, gram-negative bacterium in the family Pasteurellaceae. P. multocida
is part of the normal microbial flora of the upper respiratory tract of many animal species, but is also a potential
pathogen of many domestic and agriculturally important animals, such as dogs, cats, cattle, pigs, and avian
species 275. P. multocida is also an important human pathogen following direct inoculation into subcutaneous
tissues (e.g. bite wounds) 276. In hosts in which the innate immune response is compromised (such as by prior viral
infection, immunosuppression, or stress) P. multocida is able to gain access to the lower respiratory tract and
cause respiratory disease and systemic infection. In swine, P. multocida can cause a chronic polymicrobial
infection (usually with Bordetella bronchiseptica) called atrophic rhinitis 148, 277, 278. However, P. multocida is not
considered part of the normal flora of birds, in which it can be a highly invasive primary pathogen 8. Nonetheless,
birds that recover from infection and obtain specific immunity can remain colonized by P. multocida, resulting in
asymptomatic carriage and spread of the organism to non-immune birds 150, 279, 280. Furthermore, birds can also
become colonized with P. multocida strains of low virulence 175, 281. An important question is whether low
virulence strains can revert to a highly virulent phenotype if they infect naive animals. An essential virulence
factor of P. multocida is a glycosaminoglycan capsular polysaccharide (CPS), which helps shield other surface

antigens from the host immune system 282 and prevents phagocytosis and bactericidal activity, among other roles
32. There are five P. multocida CPS serogroups based on capsular antigens of distinct structural and antigenic
specificity, designated A 41, B, D, E 283, and F 34. CPS serogroup A is composed of hyaluronic acid, serogroup D is
a polysaccharide susceptible to enzymes that degrade chondroitin sulfate A and C, and heparinase, and serogroup
F is a polysaccharide similar to chondroitin 284. The serogroup B CPS is composed predominately of mannose, but
also contains arabinose and galactose, while the composition of the CPS of serogroup E strains has not been
determined 32.
One of the most economically important diseases of cattle in the U.S. beef and dairy industries is bovine
respiratory disease (BRD) complex 285. The cost of BRD to the cattle industry has been estimated at more than
$500 million per year 183. The most common bacterial agents responsible for BRD include Mannheimia
haemolytica, Pasteurella multocida (CPS serogroup A), Histophilus somni, and Mycoplasma spp. Isolation of
more than one causative agent from a BRD infection is common 243. For example, P. multocida has been isolated
from calves with lower respiratory disease following challenge with H. somni 120, 192. Stresses such as crowding
(feedlots), shipping, weaning, and viral infection further predispose the animals to infection 182. Transmission of
BRD disease agents likely occurs by aerosol or physical contact between animals. Another common disease
associated with P. multocida, as described above, is avian cholera, which can affect most avian species and occurs
worldwide. However, some birds, such as turkeys and waterfowl 148, 150, are more susceptible to serious disease.
The fatal infectious dose of P. multocida for Mallard ducks is as few as 12 cells 151. P. multocida can be
transmitted through watering systems 152 (such as troughs and ponds that are shared by infected and healthy
birds), rodent infestations 161, and by the fecal-oral route 168 resulting in widespread infection and mortality. The
most devastating outbreaks of avian cholera occur in locations where flocks of geese tend to migrate. For
example, a single outbreak affected close to 20,000 birds and was associated with healthy migrating geese
carrying P. multocida A:1 in nasal, oral, and cloacal samples 150. In humans, about 300,000 visits to emergency
rooms are due to animal bite or scratch wounds 286, and Pasteurella spp. are isolated from about 50% of bites

from dogs and about 75% of bites from cats 287. However, bacteremia and systemic diseases without invasive
animal contact have also been reported in humans 288.
Pasteurella multocida isolates from BRD and avian cholera (both serogroup A), have been reported to form a
biofilm in vitro 201, and it has been proposed that swine atrophic rhinitis (serogroup D isolates) is a biofilm
infection 119. However, characterization and careful analysis of a P. multocida biofilm and the biofilm matrix has
not been reported. Biofilm infections within the host are a complex mixture of bacterial and host cells,
exopolysaccharide (EPS), extracellular nucleic acids, trapped nutrients in water, and proteins. These bacterial
communities are comparable to tissues formed by multicellular eukaryotes - the bacterial cells show cooperation,
fluids and nutrients are circulated, and the bacteria are protected from unfavorable conditions in the external
environment 121.
In this report the formation of biofilm in vitro and the biofilm extracellular matrix (ECM) of P. multocida
serogroup A laboratory strains and clinical isolates were thoroughly characterized. Of significance was
determining that the amount of CPS produced by P. multocida was inversely proportional to the amount of
biofilm formed, and that highly encapsulated, poor biofilm forming strains could be converted to robust biofilm
formers following loss or reduction of CPS by mutagenesis or in vitro passage. The biofilm matrix consisted of at
least protein and a newly identified glycogen EPS. A putative polysaccharide synthesis and export locus was also
identified, but appeared to be constitutively expressed during biofilm formation or during planktonic growth.
3.4 RESULTS
3.4.1 Relationship between CPS production and biofilm formation by P. multocida.
Strain Common
name
Serotypea/
phenotype
Source

P. multocida subsp. gallicida P1059 WT P1059 A:3/
encapsulated
National Animal
Disease Center
(NADC)
P. multocida subsp. gallicida P1059ΔhyaE P1059ΔhyaE Non-
encapsulated
This study
P. multocida subsp. multocida P1062 WT P1062 A:3/
encapsulated
NADC
P. multocida subsp. multocida P1062ΔhyaE P1062ΔhyaE Non-
encapsulated
This study
P. multocida X73 WT X73 A:1/
encapsulated
NADC
P. multocida X73ΔhyaD X73ΔhyaD Non-
encapsulated
This study
Pasteurella multocida C0513 WT C0513 A/
encapsulated
Experimental calf
infection3
Pasteurella multocida C0513-P5 C0513-P5 A (capsule-
deficient)
This study
Clinical
Isolates
Location of Origin Host species and infection site Serogroup
or Serotype
756 Virginia Department of Agriculture,
Division of Animal and Food
Industry Services
Chicken heart, spleen, bone
marrow
F:9
765 Virginia Department of Agriculture,
Division of Animal and Food
Industry Services
Turkey liver, spleen, lung A
704 Virginia Department of Agriculture,
Division of Animal and Food
Industry Services
Chicken wattle A:8
801 Virginia Department of Agriculture,
Division of Animal and Food
Industry Services
Chicken heart, spleen, bone
marrow
A
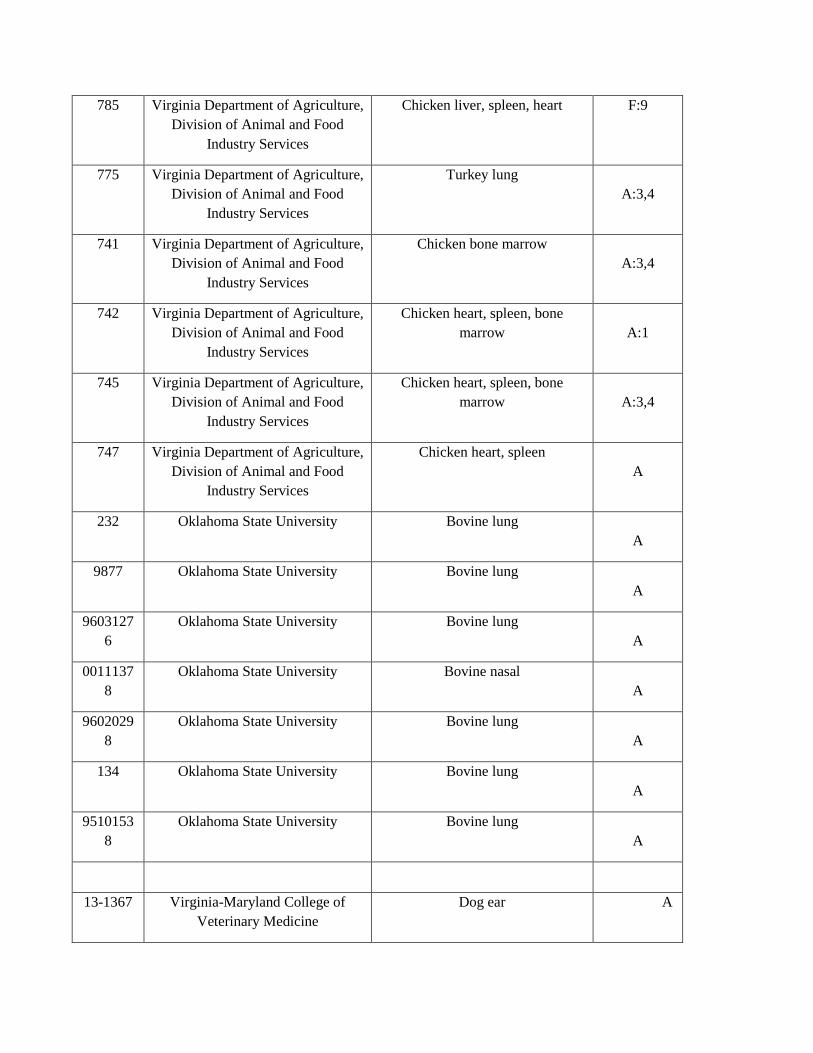
785 Virginia Department of Agriculture,
Division of Animal and Food
Industry Services
Chicken liver, spleen, heart F:9
775 Virginia Department of Agriculture,
Division of Animal and Food
Industry Services
Turkey lung
A:3,4
741 Virginia Department of Agriculture,
Division of Animal and Food
Industry Services
Chicken bone marrow
A:3,4
742 Virginia Department of Agriculture,
Division of Animal and Food
Industry Services
Chicken heart, spleen, bone
marrow
A:1
745 Virginia Department of Agriculture,
Division of Animal and Food
Industry Services
Chicken heart, spleen, bone
marrow
A:3,4
747 Virginia Department of Agriculture,
Division of Animal and Food
Industry Services
Chicken heart, spleen
A
232 Oklahoma State University Bovine lung
A
9877 Oklahoma State University Bovine lung
A
9603127
6
Oklahoma State University Bovine lung
A
0011137
8
Oklahoma State University Bovine nasal
A
9602029
8
Oklahoma State University Bovine lung
A
134 Oklahoma State University Bovine lung
A
9510153
8
Oklahoma State University Bovine lung
A
13-1367 Virginia-Maryland College of
Veterinary Medicine
Dog ear A

H1 Virginia-Maryland College of
Veterinary Medicine
Cat mouth A
12B-
21481
University of Wyoming Bovine liver, lung, small intestine A
Virginia-Maryland College of
Veterinary Medicine
Porcine lung
D
Pm70 ATCC (BAA-1909) Chicken F
Pm70 Dr. Kapur, Pennsylvania State
University
Chicken
F
989 Virginia-Maryland College of
Veterinary Medicine
Turkey
Table 3.1: Laboratory strains and clinical isolates used. aA standard system for designating serotypes of P.
multocida was established using a combination of capsular serogroup typing and LPS serovar typing systems 289.
A collection of clinical isolates and laboratory strains (Table 3.1) of P. multocida was screened for their
capability to form a biofilm by crystal violet assay. Upon initial screening, it was noted that there was an inverse
correlation between the mucoid appearance of the colonies (an indication of the degree of encapsulation) and the
amount of biofilm formed (Figure 3.1A). To confirm the association between encapsulation and biofilm
formation, isogenic mutants of strains WT P1059 and WT P1062 were made by mutating the hyaE gene, and in
strain X73 by mutating hyaD. Recent clinical isolate C0153 was subcultured in vitro daily for 5 passages, by
which time it was able to form a prominent biofilm. The P. multocida serogroup A capsule is composed of
hyaluronic acid, which is not immunogenic due to its presence in host connective, epithelial, and neural tissues,
thus negating the use of assays that utilize antibodies for antigen quantification. Therefore, to quantify CPS on the
mutants mid-log phase cultures (OD562 = 0.6) were treated with hyaluronidase to release from the CPS the uronic
acid, which was quantified by chemical assay. We confirmed the phenotypic observations (colony iridescence
under incandescent lighting) that mutants P1062ΔhyaE and X73ΔhyaD, and the passaged variant of C0153, all
made significantly less CPS than the wildtype (WT) strains. WT P1062 produced 67.01 μg/ml of uronic acid

compared to 22.62 μg/ml of uronic acid for P1062ΔhyaE; WT X73 produced 83.02 μg/ml of uronic acid, whereas
X73ΔhyaD produced 15.90 μg/ml of uronic acid (p < 0.0001 for both strains). P. multocida P1059ΔhyaE also
made less uronic acid than WT P1059, but the difference was not significant (p = 0.1147) because WT P1059
produced relatively little uronic acid (CPS) compared to other strains (20.39 μg/ml of uronic acid for the parent
compared to 13.22 μg/ml of uronic acid by P1059ΔhyaE (Figure 3.1B). Thus, all mutants with the same mutation
in hyaE and the passaged strain were capsule-deficient. To further confirm that a deficiency in CPS production
was responsible for enhanced biofilm formation, the mutation in strains P1059ΔhyaE and P1062ΔhyaE was
complemented in trans. Uronic acid production was restored and enhanced in complemented mutant
P1059ΔhyaE[hyaE], and biofilm formation was reduced. Biofilm formation by complemented mutant
P1062ΔhyaE[hyaE] was also reduced to the same amount as the parent (Figure 3.1B). P. multocida clinical
isolate, WT C0513, was a mucoid, poor biofilm forming strain that was isolated from a calf experimentally
challenged with Histophilus somni 120. WT C0513 was subcultured under biofilm-favoring growth conditions (5
subcultures in RPMI medium every 48 hrs) until the strain could form a significant biofilm compared to the
parent (p < 0.0001). WT C0513 produced 89.38 μg/ml of uronic acid, while the subcultured variant (C0513-P5)
produced 13.87 μg/ml of uronic acid (p < 0.0001) (Figure 3.1B), further supporting that CPS production was
inversely correlated with biofilm formation.
We also observed that when P. multocida was grown on Dextrose Starch agar (DSA) supplemented with
Congo Red (CR) the presence of CPS inhibited CR uptake, enabling us to develop an assay to semi-quantify CPS.
Strains deficient in biofilm formation had low CR absorption indexes, while proficient biofilm forming strains
had significantly higher CR absorption indexes (P1059ΔhyaE, p ≤ 0.0001; P1062ΔhyaE, p ≤ 0.05; X73ΔhyaD, p
≤ 0.001) compared to their respective parent strains. Differences in CPS quantity between the parent and mutant
strains were greater for the CR assay than the uronic acid assay. A correlation graph was generated to evaluate the
effectiveness of this assay in predicting biofilm forming potential (Figure 3.1C). However, the correlation
between CR absorption and CV biofilm quantification for P1059ΔhyaE was an outlier on the correlation graph.
Upon further investigation, we determined that there was a modification in the lipooligosaccharide (LOS)

electrophoretic profile of P1059ΔhyaE that was distinct from that of the parent and not present in the other mutant
strains (data not shown). The LOS modification may have contributed to enhanced CR uptake, particularly if
more neutral sugars were expressed 290. If P1059ΔhyaE was excluded from statistical analysis of the correlation
graph, the Pearson r-value was 0.9635, further supporting a strong inverse correlation between CPS and biofilm
formation. The CR assay was repeated on each group of clinical isolates (Figure 3.1A) and CR uptake correlated
inversely with colony morphology (iridescence): group 1 (rough, capsule-deficient strains), group 2 (smooth,
moderately encapsulated strains), and group 3 (mucoid, highly encapsulated strains). Biofilms were quantified by
CV uptake, and CPS by inhibition of CR uptake. Comparative measurements between the amount of biofilm
formed with the amount of CPS produced consistently indicated that when more CPS was present less biofilm
was formed. Group 1 isolates overall produced a more robust biofilm than either group 2 or group 3 isolates (p <
0.0001 for both groups) based on CV quantification. Group 2 isolates produced some biofilm, which was more
than that produced by the mucoid strains, but the differences were not significant (p > 0.05).
To further investigate the inverse relationship between CPS production and biofilm formation, hyaluronidase
was added to the biofilm growth medium to degrade the CPS, as this process has been shown to at least partially
eliminate the hyaluronic acid CPS from the surface of P. multocida serogroup A isolates 291, 292. When 300 U of
hyaluronidase was added to encapsulated cultures of strains WT P1059, WT P1062, WT X73, and WT C0513, all
of which were poor biofilm formers, biofilm formation by each strain increased significantly (p < 0.0001 for each
strain), as determined by quantitative CV staining (Figure 3.2). These collective results indicated that serogroup A
CPS interfered with biofilm formation.

Figure 3.1: Correlation between CPS production and biofilm formation by P. multocida clinical isolates and
laboratory strains. A) Clinical and laboratory isolates were grouped based on their colonial morphology (amount
or lack of mucoid appearance and iridescence) on blood agar in relation to biofilm formation. The Y-axis
represents the A562 after staining the biofilm with crystal violet and solubilizing the residue with 95% ethanol.
Groups: (1) rough colonies/unencapsulated, (2) smooth colonies/intermediate encapsulation, (3) mucoid
colonies/highly encapsulated. The amount of biofilm formed (as indicated by CV assay) by isolates from groups 2
and 3 was significantly lower than the amount of biofilm formed by group 1 isolates (****, p ≤ 0.0001). B)
Comparison of biofilm formation by wild type (WT) strains and their respective isogenic capsule-deficient
mutants or an in vitro-passaged variant of WT C0153 (C0153-P5). The amount of biofilm and CPS was
determined by crystal violet assay and uronic acid assay, respectively. The WT strains and their respective
capsule-deficient mutants are listed on the X-axis. The left Y-axis represents μg/ml uronic acid removed from the
cell surface. The right Y-axis is the absorbance of solubilized crystal violet after staining. White bars indicate the
absorbance value from crystal violet staining; black bars indicate uronic acid content. Biofilm formation was
significantly higher in isolates producing less CPS. Significant differences between parent and mutant strain for
the CV assay are indicated by asterisks, and for the uronic acid assay by number symbol, respectively: *, p ≤ 0.05,
**, p ≤ 0.01, ***, p ≤ 0.001, **** or ####, p ≤ 0.0001. C) Correlation plot of P. multocida Congo Red uptake
absorbance values (y-axis) and CV absorbance values for biofilms (x-axis). The Pearson coefficient is 0.7324 for
all values and 0.9635 if P1059ΔhyaE is excluded. Encapsulated isolates are represented by solid dots, while
acapsular isolates are represented by hollow dots.

Figure 3.2: Effect of hyaluronidase on biofilm formation during growth. Either 0 or 300 μg of hyaluronidase
was added to the culture medium of P. multocida strains under growth conditions favoring biofilm formation.
After two days the biofilms were rinsed, solubilized with 95% ethanol and the absorbances determined at A562.
Significance values are based on comparison to the sample with no enzyme added. *, p ≤ 0.05; **, p ≤ 0.01; ***,
p ≤ 0.001; ****, p ≤ 0.0001. White bars, strain WT C0513; dark grey bars, strain WT X73; black bars, strain WT
P1059; light grey bars, strain WT P1062.
3.4.2 Chemical and genomic analysis of the matrix exopolysaccharide.
Gas chromatography-mass spectroscopy (GC-MS) indicated that the EPS extracted from the biofilm
matrix was a polymer composed of glucose that was either terminal 4-linked, or 4,6-linked (Figure 3.3A). The
nuclear magnetic resonance (NMR) proton spectrum displayed one main anomeric signal at 5.3 ppm (Figure
3.3B) that is distinct for a glucose with α configuration at the anomeric center. This information along with
chemical data, suggested that the EPS had a glycogen-like structure, later confirmed by digesting the
polysaccharide with pullulanase, which depolymerized the polymer, as determined by NMR measurement (Figure
3.3C).
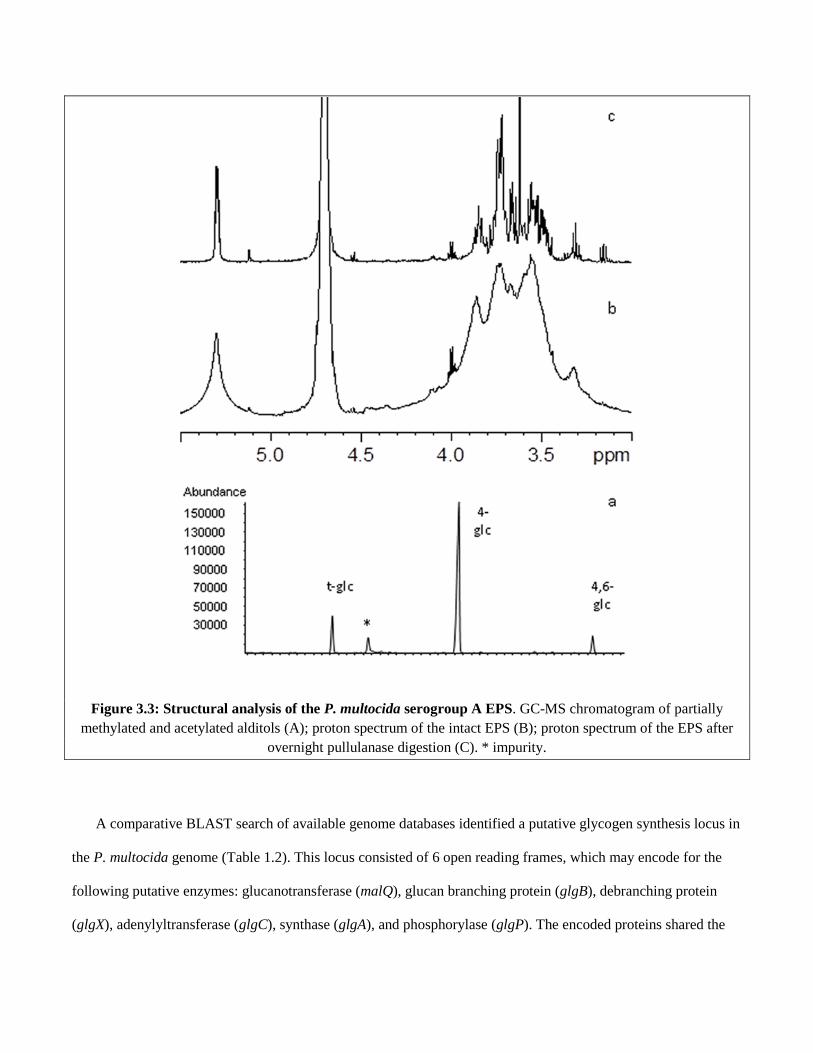
Figure 3.3: Structural analysis of the P. multocida serogroup A EPS. GC-MS chromatogram of partially
methylated and acetylated alditols (A); proton spectrum of the intact EPS (B); proton spectrum of the EPS after
overnight pullulanase digestion (C). * impurity.
A comparative BLAST search of available genome databases identified a putative glycogen synthesis locus in
the P. multocida genome (Table 1.2). This locus consisted of 6 open reading frames, which may encode for the
following putative enzymes: glucanotransferase (malQ), glucan branching protein (glgB), debranching protein
(glgX), adenylyltransferase (glgC), synthase (glgA), and phosphorylase (glgP). The encoded proteins shared the

greatest identity with other members of the Pasteurellaceae, most notably Aggregatibacter spp., Necropsobacter
rosorum, and Haemophilus influenzae (Table 1.2).
Table 3.2: Pasteurella multocida strain WT P1059 genes with amino acid identity to glycogen synthesis
proteins in related bacteria.
3.4.3 Enzymatic treatment of biofilms.
Developing biofilms were treated with proteinase K and α-amylase to determine their effect on biofilm
development, and to evaluate matrix composition (Figure 3.4). The WT P1059 biofilm retained 91% and 93% of
the CV stain after exposure to proteinase K (p = 0.9957) and α-amylase (p = 0.9977), respectfully, when
compared to biofilm growth in the absence of these enzymes. These results reflect the lack of biofilm formed by
strain WT P1059. In the more biofilm robust, CPS-deficient strain P1059ΔhyaE, there was 61% and 57% less
biofilm after growth with proteinase K and α-amylase, respectfully (p = 0.0065 and p = 0.0118, respectively).
Therefore, while more than half of the biofilm matrix was removed after enzymatic treatment of P1059ΔhyaE,
less than 10% of the WT P1059 matrix was removed after treatment with either enzyme. Similar results were

obtained for the highly encapsulated, biofilm-poor strain WT C0513 and the in vitro-passaged variant C0513-P5.
In WT C0513, 79% of biofilm CV stain was retained after proteinase K treatment (p=0.9123), and a small, but
insignificant, increase in CV staining occurred after treatment with α-amylase (p=0.2249). In subcultured variant
C0513-P5, 44% of the biofilm CV stain was lost after proteinase K treatment (p < 0.0001), and 37% of the
biofilm CV stain was lost after α-amylase treatment (p < 0.0001). The H. somni 2336 biofilm was not affected by
treatment with α-amylase, but was diminished to a similar extent after treatment with proteinase K (data not
shown).
Figure 3.4: Enzyme digestion of biofilm matrix. Prior to inoculation of the culture media with P. multocida, 0.1
mg/ml of α-amylase or Proteinase K was added, followed by incubation for 48 hrs. Biofilms were rinsed, stained
with CV, solubilized with ethanol and the A562 determined. White bars, no treatment; Light grey bars, treated
with Proteinase K; Dark grey bars, treated with α-amylase. Significance values are based on comparison to the
sample with no enzyme added. *, p ≤ 0.05; **, p ≤ 0.01; ***, p ≤ 0.001; ****, p ≤ 0.0001; ns, not significant.

3.4.4 Scanning Electron Microscopy (SEM).
Strains WT P1059 and P1059ΔhyaE biofilms were grown on abiotic cover slips and examined for
formation and structure by SEM. WT P1059 formed micro-colonies surrounded by small amounts of biofilm
matrix. Individual bacterial cells or small clusters of 2-6 bacteria adhered to the glass coverslip, and were
dispersed between micro-colonies (Figure 3.5A). The P1059ΔhyaE biofilm matrix was clearly visible, included
peaks and valleys characteristic of biofilm matrices, and was more extensive than the biofilm of the parent. Some
individual bacterial cells adhered to the glass coverslip below the matrix, similar to that of the WT (Figure 3.5B).
Figure 3.5: SEM images of P. multocida biofilms after 48 hours incubation on glass coverslips. A) Strain WT
P1059; B) P1059ΔhyaE biofilm formation.
3.4.5 Confocal Laser Scanning Microscopy (CLSM).
Two-day-old biofilms of strains WT P1059 and P1059ΔhyaE were imaged using CLSM, converted into
z-stacks, and analyzed for structural characteristics (Table 1.3). Biofilms of WT P1059 (n=10) were best
characterized as a cell monolayer, with an average thickness of 0.001719 µm3/µm2, with a roughness coefficient
of 2, which indicated maximum roughness. Of interest, live/dead staining of the WT P1059 monolayer identified
living cells adherent to the glass coverslip, while dead cell debris was present on the biofilm exterior (Figure

3.6A, Figure 3.7A). The P1059ΔhyaE biofilm had an average biomass volume of 21 µm3/µm2, and live/dead
staining indicated the biofilm was composed primarily of living cells. The topmost layer appeared to have small
clusters of dead cellular debris (Figure 3.6B, Figure 3.7B).
Straina Biomass
(µm3/µm2)
Thickness
(µm)
Maximum
Thickness
(µm)
Roughness
Coefficient (0-
2)
Surface area to
Bio-volume
ratio
WT P1059 0.0449 ±
0.0032
0.001719 ±
0.0003
4.5 2 9.92
P1059ΔhyaE 21 ± 3.23 29.79 ± 4.73 34.31 2 5.05
Table 3.3: COMSTAT analysis results obtained from CSLM Z-stack images. a N=10 for all strains.
Figure 3.6: CSLM z-stack showing live/dead staining of WT P1059 during biofilm formation. A) WT P1059;
B) P1059ΔhyaE. Live stain (left column), dead stain (center column), live-dead overlay (right column).

Figure 3.7: Cross section of the biofilm by CSLM. This is a cross-sectional view through the center of the WT
P1059 biofilm (A) and P1059ΔhyaE biofilm (B) by CSLM. The left column and top row show the height and
thickness of each biofilm, which is larger and thicker for capsule-deficient mutant P1059ΔhyaE than for the
parent strain.
3.4.6 qRT-PCR of putative EPS matrix genes.
The expression of genes proposed to encode for the glucan EPS of subcultured variant C0513-P5 and
parent WT C0513 during planktonic growth were similar, with only minor differences noted based on qRT-PCR.
Gene expression of both WT C0513 and C0513-P5 during planktonic growth was also compared with gene
expression of the same strain during biofilm formation (Figure 3..8). Constitutively expressed gyrB was used as a
control to normalize the fold-increases between strains. Most genes in the CPS locus, putative glycogen synthesis
locus, genes encoding putative glycosyltransferases 293, 294, LPS synthesis (e.g. waaA 295), and genes that encode
other putative EPS biosynthesis proteins (e.g. opsX 296), were not differentially expressed between cells grown
planktonically or as a biofilm for either strain (not shown). Therefore, differences in gene expression that would
distinguish a proficient from a deficient biofilm former were not identified in those genes. However, CsrA
(carbon storage regulator A), HexD (CPS export gene), and XylB (xylulose kinase) were upregulated in both

parent and proficient biofilm former C0513-P5. However, of interest was that expression of SgbU (L-xyulose-5-
phosphate 3-epimerase) was greatly enhanced in only parent WT C0513.
Figure 3.8: Normalized fold-increase of genes significantly upregulated during biofilm formation. Strain
WT C0513 was grown for 48 hrs under stationary conditions to promote biofilm growth, and subcultured 5
consecutive times under the same growth conditions to obtain strain C0153-P5. Grey bars, strain WT C0513
(biofilm-deficient strain); white bars, variant C0513-P5 (biofilm-proficient strain). *, p ≤ 0.05; **, p ≤ 0.01; ***,
p ≤ 0.001; ****, p ≤ 0.0001.
3.5 DISCUSSION
Pasteurella multocida has the capacity to cause acute or chronic infections in many animal species and
humans. Chronic infections include atrophic rhinitis in swine or snuffles in rabbits, and birds and cattle may carry
P. multocida asymptomatically and act as a host carrier. Avian P. multocida carrier animals demonstrate few or
no outward signs of infection 7. Carrier migratory birds, such as geese 280, may act as persistent reservoirs of
infection that can spread virulent P. multocida as they migrate between water supplies 297, 298. Samuel et al. 281
reported that parenteral challenge of Mallard ducks with “relatively low virulence” P. multocida resulted in
establishment of a carrier state in both challenged birds and birds in contact with the infected birds. Of interest is

that Pritchett et al. 299 described endemic fowl cholera in flocks of Leghorn chickens from which isolates that
formed “blue” colonies (acapsular phenotype) were associated with a relatively high incidence of the carrier state
in birds, relatively little disease, and a high incidence of contact carrier birds. Subsequently, Prichett et al. 175
reported that intranasal challenge of Leghorn chickens with “fluorescent colonies” (presumably encapsulated) of
an epidemic strain resulted in high mortality, but low incidence of carrier animals and little spread to contact
birds. In chronic or asymptomatic infections by “blue” colony types, the bacteria may be present as a biofilm, in
which the bacteria are relatively innocuous and there is little inflammatory response 8, 151, 300. However, bacteria
shed from the biofilm may be in a planktonic state, which may be more pathogenic and contagious to susceptible
animals. In this transmission model, biofilm formation is essential to the spread of potential pathogens. Although
biofilm formation has been proposed as a component of some P. multocida infections 119, 201, this is the first study
to more thoroughly characterize the composition and formation of P. multocida biofilms.
Examination of biofilms made by a wide variety of clinical and laboratory isolates indicated that all strains of
P. multocida could form some biofilm, but that there was wide variation in the amount of biofilm that individual
strains could form. Only isolates of non-toxigenic CPS serogroups A and F of avian or bovine origin were
reported. However, biofilm formation by serogroup D isolates of porcine origin was also confirmed in our assays
(not shown), as proposed previously 119. When the extent of biofilm formation was compared with colony
morphology under iridescent light it was noted that there was an inverse relationship between the iridescence and
how mucoid the colonies were, and the amount of biofilm formed by that strain. It has been clearly established
that cells with the most CPS on their surface form the most mucoid, iridescent colonies, with smooth colonies
containing less CPS, and smaller, blue colonies (rough) being CPS-deficient 301, 302. Inverse correlations between
CPS content and biofilm formation that are similar to the results described here for P. multocida, have also been
reported for Neisseria meningitidis 303, Escherichia coli 304, 305, and other bacterial species 303, 306.
In order to confirm the inverse relationship between CPS content and biofilm content an accurate assay to
quantify CPS was necessary. Immunoassays are unavailable because antibodies against the type A CPS
hyaluronic acid are difficult to obtain because this CPS is recognized as “self” by the immune system. An

alternative method to quantify P. multocida type A CPS production is by observing the colony morphology on
solid agar medium 301. This method has been used to distinguish encapsulated, virulent P. multocida from less
virulent or avirulent decapsulated P. multocida for almost a century 171. However, this method is not quantitative
and we determined that P. multocida was capable of producing more than one polysaccharide, prompting the need
for a quantification method that can differentiate hyaluronic acid from glycogen and potentially xylan 307. Several
methods that have been used for quantifying hyaluronic acid were not useful in distinguishing hyaluronic acid
from other or similar polysaccharides due to a lack of test specificity; these assays use reagents that react with
carbohydrates, but not specifically hyaluronic acid 282, 308, 309. Therefore, enzymatic removal of the CPS from the
cell surface was used to quantify the uronic acid present using a carbazole assay 310. In addition, we determined
that the CPS inhibits CR uptake by the bacteria because CR preferentially binds to neutral carbohydrates 311.
Therefore, the amount of CPS produced was inversely proportional to CR uptake by P. multocida isolates. This
assay correlated with biofilm formation based on larger index values of isolates that produced a proficient biofilm,
and a smaller index value that correlated with isolates that were deficient in biofilm formation.
Further evidence that CPS inhibited biofilm formation was provided through the use of three isogenic CPS-
deficient mutants with mutations in hyaE or hyaD. Subculture of P. multocida in vitro has long been known to
result in spontaneous loss of CPS (48). We confirmed this observation by growing an encapsulated isolate under
biofilm-favoring growth conditions for 5 passages, resulting in reduced CPS production, while simultaneously
increasing biofilm formation (strain C0513-P5). Thus, the removal of CPS (based on uronic acid assay of
enzymatically treated cells and the CR uptake assay) enhanced proficient biofilm formation in a previously poor
biofilm forming strain. Therefore, CPS may inhibit biofilm formation by blocking surface proteins essential for
adherence; the first step involved in biofilm formation 312. CPS has also been shown to physically block adhesion
factors essential to biofilm formation in E. coli 313, 314. Complementation of mutants P1059ΔhyaE and
P1062ΔhyaE with hyaE in trans reduced biofilm formation and enhanced uronic acid content in both
complemented recombinant strains, further supporting that the presence and the amount of CPS interfered with
biofilm formation. Finally, hyaluronidase treatment to remove most of the CPS from the cell surface of each

parent strain significantly enhanced biofilm formation, but had little to no effect on rough, good biofilm forming
strains or CPS-deficient strains (data not shown for the latter). However, hyaluronidase treatment was less
effective, though still significant, at enhancing biofilm formation by WT X73 than the other 3 strains. Why
hyaluronidase was not as effective in enhancing biofilm formation in strain X73 could be explained if this strain
also produces the β-(1,4)-D-xylan polysaccharide that has been reported for at least one other strain 307. A D-xylan
polysaccharide would not be susceptible to removal by hyaluronidase.
Biofilms are composed primarily of cells encased in a matrix material that is predominately EPS, but also
proteins and extracellular nucleic acids 121. Once it was established that some strains of P. multocida could form a
substantial biofilm, we sought to identify whether the matrix material consisted of a polysaccharide distinct from
the CPS. Following procedures used previously to isolate EPS from H. somni 258, an α-(1,4)-linked polymer of
glucose (glycogen) was isolated. This is the first description of a glycogen polymer produced by P. multocida.
However, qRT-PCR indicated that the putative genes responsible for glycogen synthesis were not upregulated
during biofilm formation, indicating that some glycogen may always be produced 315, thus accounting for small
amounts of biofilm in all strains tested. It is possible that the large amount of CPS produced by mucoid strains
masked the presence of the glycogen. Surface-expression of CPS may also inhibit biofilm formation by blocking
cell surface adhesins 312. The addition of hyaluronic acid, the component of serogroup A CPS, to the biofilm
growth medium had no effect on biofilm formation, likely because the added polysaccharide was not associated
with the cell surface, as it is when produced by the cell. However, as mentioned above, the addition of
hyaluronidase to hydrolyze the serogroup A CPS present did enhance biofilm formation by highly encapsulated,
poor biofilm formers. Furthermore, at least one gene responsible for CPS export was upregulated during
proficient biofilm formation, indicating the EPS and CPD may be exported by a common pathway.
Enzymatic digestions of developing biofilms were used to determine the relative amounts of protein and
carbohydrate within the biofilm. Proteinase K and α-amylase enzymes supplemented in the biofilm growth
medium resulted in partial digestion of the biofilm material, but had little effect on isolates deficient in biofilm
formation. These results supported that the biofilm matrix consisted of glycogen, the substrate for α-amylase, and

that the biofilm matrix was also composed of protein or that the matrix structure was dependent on a protein
scaffolding.
Previous studies have shown that P. multocida can display large quantities of lipids on its surface, causing the
bacteria to become hydrophobic 316, 317, and increasing the adherence of the bacteria to surfaces 318; an essential
early step in biofilm formation. However, in studies not shown there was no difference between the parent and
acapsular mutant strains in hydrophobicity (as determined by the bacteria moving from an aqueous phase to an
organic phase), indicating that hydrophobic interactions did not contribute to biofilm formation in P. multocida.
Auto-aggregation can also contribute to biofilm formation, and is often facilitated by EPS and/or bacterial
adhesins 319, 320. However, no differences in auto-aggregation were noted between proficient and deficient biofilm
forming strains (p = 0.227) (data not shown), indicating that cell-to-cell interactions were not inhibited by CPS,
and auto-aggregation was not a good predictor of biofilm formation by P. multocida.
Changing growth conditions from those that favor planktonic growth (approximately 6 hours with rapid
shaking) to those that favor biofilm growth (>6 hours under stationary conditions) was sufficient to initiate
changes in gene expression. H. somni and P. multocida are genetically related. Therefore, homologs of genes
shown to be upregulated during H. somni biofilm formation 259 were included in our qRT-PCR assays during
biofilm formation for P. multocida. As expected, some of the genes upregulated during H. somni biofilm
formation were also upregulated during P. multocida biofilm formation. For example, the carbon storage regulator
CsrA was upregulated during biofilm formation by both H. somni 258 and P. multocida. However, CsrA is a RNA-
binding protein that represses biofilm formation in some Gram-negative bacteria, such as E. coli 321 and
Campylobacter jejuni 322. In E. coli, CsrA influences biofilm suppression and dispersal primarily through its
regulatory effect on glycogen synthesis and catabolism 321. Although the effect of CsrA on biofilm formation is
reversed between E. coli and P. multocida, upregulation of CsrA enhances glycogen synthesis, which in P.
multocida enhances biofilm formation. Xylulose kinase (XylB) and L-xyulose-5-phosphate 3-epimerase (SgbU)
were also upregulated during P. multocida biofilm formation (though in the latter only in the parent strain). A β(1-
4)-D-xylan polysaccharide has been identified as an additional polysaccharide of at least one other P. multocida

serogroup A strain 307. Therefore, xylan could be another EPS that forms the biofilm matrix of some strains, such
as C0153, which was used for qRT-PCR, but not for EPS purification. Expression of SgbU was examined because
this gene was also upregulated during H. somni biofilm formation259. Of interest was that SgbU was only
upregulated during biofilm formation by the parent and not by the mutant. Therefore, we suspect that SgbU was
not directly related to biofilm formation, but may be affected by a global regulator, such as Fis, which also down-
regulates CPS expression following subculture42.
A gene in the P. multocida CPS locus, hexD, encodes for a protein required for polysaccharide export 32, 41
and was up-regulated during biofilm formation. Therefore, one or more genes responsible for CPS export may
also contribute to EPS export. Genes involved in hyaluronic acid biosynthesis are in region 2, and were neither
up-regulated or down-regulated. Genes in the putative glycogen synthesis locus were not differentially regulated
during proficient biofilm formation when compared to isolates deficient in biofilm formation. Therefore, glycogen
may be constitutively expressed, but can only contribute to matrix formation when CPS is absent and adherence
and biofilm formation can be initiated.
In summary, we have demonstrated that biofilm formation by P. multocida in vitro is inversely related to CPS
production. The biofilm was composed of protein and a newly identified glycogen amylose EPS (and likely
extracellular DNA). Since isolates that produce less CPS are less virulent and proficient biofilm formers, we
propose that down-regulation of CPS 146 may result in chronic pasteurellosis and avian carriers due to enhanced
biofilm formation.
3.6 MATERIALS AND METHODS
3.6.1 Isolates and growth conditions.
Laboratory and clinical isolates of P. multocida used in this study, and their sources, are described in
Table 1. Strain C0513 was isolated from the trans-tracheal wash of a calf following experimental challenge 120

with Histophilus somni. Prior to challenge naso-pharyngeal cultures from the calf were negative for P. multocida
and other respiratory pathogens. All P. multocida strains were cultured on Brain Heart Infusion (BHI) or
Columbia blood agar (BD; Franklin Lakes, NJ) supplemented with 5% defibrinated sheep blood (HemoStat
Laboratories Inc.; Dixon, CA), or DSA supplemented with or without 0.005% CR. Agar-grown cultures were
grown at 37oC with 6% CO2. Broth cultures were grown in BHI or RPMI-1640 medium without glutamine or
phenol red (Lonza; Walkersville, MD). For biofilm formation, 50 μl of mid-log phase P. multocida was
inoculated into 5 ml of RPMI-1640 in a 50-ml polyethylene tube, and incubated without shaking at 37oC in 6%
CO2 for at least 48 hours.
All P. multocida isolates were confirmed and serogrouped by multiplex PCR, as described elsewhere 43. All
isolates used in this study were nontoxigenic, and either serogroup A, F, or untypable (neither A, B, D, E, or F).
3.6.2 Isolation of a biofilm-proficient P. multocida variant.
Isolate WT C0513 was subcultured every 48 hours under conditions favoring proficient biofilm growth
(described above) until the isolate was capable of producing a proficient biofilm, which occurred after 5
subcultures. This variant is referred to as C0513-P5.
3.6.3 Construction of acapsular P. multocida mutants.
Gene-replacement mutants of P. multocida isolates WT P1059 (avian, serogroup A:3), WT P1062 (bovine,
serogroup A:3), and WT X73 (avian, serogroup A:1) were generated using previously described techniques with
minor modifications 90. Briefly, DNA fragments containing the hyaE gene of P1059 and P1062 were amplified
with forward primer 5'-ATGAAAAAGGTTAATATCATTGG-3' and reverse primer 5'-
TTAACCTTGCTTGAATCGTTTACC-3’ to produce an approximately 1870 bp fragment containing the hyaE
coding region of each isolate. These PCR products were cloned into pCR2.1 (Invitrogen Inc., LaJolla, Calif.), and

amplified with deletion primers 5'-AAAGATATCTTGGTTTACTTCAATAATTTC-3' and 5'-
AAAGATATCACTGCATCTGTTCAATCAACGAGC-3', which produced a linear product with pCR2.1 flanked
by the upstream and downstream arms of the replacement plasmid. Digestion with EcoRV (sites contained in the
primers) and ligation re-circularized the plasmid, deleting amino acids 239 through 359 of the encoded protein
and substituting isoleucine for the leucine residue at former position 360. An attempt was made to frame-shift the
deleted gene in WT P1062 by inserting a SmaI linker (5’-CCCCGGGG-3’) into the EcoRV site, but the product
produced was found to contain a series of three of the linkers, which restored the reading frame to encode an
additional 8 amino acids (Pro Arg Gly Pro Gly Ala Pro Gly) in the deletion site.
A DNA fragment containing a portion of hyaE and hyaD coding regions of X73 was amplified with forward
primer 5’-GAAGATGCGCATGAAGCCAATCGCATT-3’ and reverse primer 5’-
GCCATTTGGTTTAGACATGATG-3’ to produce an approximately 1680 bp product. This PCR product was
cloned into pCR2.1, digested with BglII restriction endonuclease, and ligated to introduce a 225-bp deletion in the
hyaD gene in-frame to the coding region.
Each of the above DNA segments from strains WT P1059, WT P1062, and WT X73 replacement DNA
segments was excised from pCR2.1 and cloned into pGA301ori, and then introduced by electroporation into the
respective strains and handled as previously described 90. PCR was used to confirm that kanamycin-sensitive
colonies lacked plasmid pGA301ori. Mutations in the CPS biosynthetic locus were confirmed by PCR using the
primers described above. Strains WT P1059 and P1062ΔhyaE yielded a PCR product approximately 360 bp
smaller than their wild-type counterparts, and the X-73ΔhyaD PCR product was approximately 220 bp smaller
than its wild-type counterpart. Each mutant appeared non-iridescent when grown on DSA under oblique
illumination, whereas each parent was clearly iridescent due to capsule formation.
To complement the mutation in P1062ΔhyaE a forward primer (5’-
AAAAAGGATCCCAAGCGTTGGGTAAAAAAACCGCTTA-3’) and reverse primer (5’-
AAAAGGATCCTCCATAGATTCCGCCGACTTTTCA-3’) were used to amplify the promoter and coding

region of hyaE using the WT P1062 genomic template. The BamHI-digested product was ligated into pBC SK-
(Stratagene) containing a kanamycin marker (pUC4K) in the SalI site. The backbone of the resulting plasmid was
replaced by digestion with BssHII and ligated to a DNA fragment containing the Mannheimia haemolytica pD80
(a 4.2 kb ampicillin-resistance plasmid) origin-of-replication. The resultant product was electroporated into
competent WT P1062 as described 323. P. multocida was made competent by growth to mid-log and 37 μl of
hyaluronidase solution (10 mg/ml) was added to 10 ml of culture and incubated for 30 minutes without shaking.
The bacteria were washed 3 times with ice cold 10% glycerol, and the bacteria allowed to resuspend in any
remaining solution after the final wash. About 50 μl of cell suspension was transferred to cold electroporation
cuvettes for electroporation into the mutant strains 90.
3.6.4 RNA extraction, PCR, and qRT-PCR.
Primer set 1 specific for toxA was used as described by Kamp et al. 324. All PCR reactions were performed in
an Eppendorf Mastercycler® pro PCR system with vapo.protect technology (Eppendorf; Germany) using OneTaq
reagents per the manufacturer’s instructions (New England Biolabs; Ipswich, MA).
At least three technical replicates of WT C0513 and C0513-P5 biofilm and planktonic cultures were pooled to
obtain RNA from each growth phase. RNA was isolated using Qiagen RNA Protect bacterial reagent, Qiagen
QiaShredder, and Qiagen RNeasy kits (Qiagen; Hilden, Germany) according to manufacturer’s instructions for
prokaryotic RNA. RNA was transcribed into cDNA using the Quanta qScript kit (Quanta Biosciences;
Gaithersburg, MD) following the manufacturer’s instructions. Quantitative real-time PCR (qRT-PCR) was
performed on an Applied Biosciences 7300 Real-Time PCR System (Applied Biosystems, Foster City, CA) using
the Quanta SYBR FastMix kit (Quanta Biosciences; Gaithersburg, MD) following the manufacturer’s
instructions. Quantitative real-time PCR was performed in triplicate for each primer set on two biological
replicates for each strain. The primers used are described in Table 3.4. A primer set identifying gyrB was used as
a control. The 2-ΔΔCT method 325 was used to calculate the normalized fold-differences in gene expression. Genes

analyzed by qRT-PCR were selected based on their relevance to biofilm formation in the related species H. somni
258, homology to genes involved in glycogen synthesis (Table 3.2), or P. multocida CPS production 41. Homology
to annotated genes was determined using BLASTP 326. The E. coli glycogen synthase (GlgA) gene sequence was
used to determine that a glgA homolog exists in the P. multocida genome 327.
Gene description Gene Forward primer sequence Reverse primer sequence
Control gyrB 5’-TATGAGTGGCTTGGCGTTAG-3’ 5’-ACACCGCTTCTGGATAATAGC-
3’
Glycogen synthesis malQ 5’-TGAACAGGCGACGAAGTATG-3’
5’-
CCGACGAGAAGATGAGCTATAAG
-3’
Glycogen synthesis glgB 5’-
CAGTAAAGACGGAGAGCGAATTA-3’
5’-CCACATAGTTACCCGCCATATT-
3’
Glycogen synthesis glgX 5’-AGCGATGTTTGCGCTAGAA-3’ 5’-
TCGATACCCGCTTGATGTAATG-3’
Glycogen synthesis glgC 5’-GGGAACCCAGTACAAACCTATC-
3’ 5’-TAATCACGCAACCACCAGAA-3’
Glycogen synthesis glgA 5’-ACATGCTTGCTCGGAGTTATAC-3’
5’-
GGATACGCCGGTAAGACAATAC-
3’
Glycogen synthesis glgP 5’-CGGAATTTATCCGTCGTGTCT-3’ 5’-GCCACGCCATTCACTTTATTC-3’
Table 3.4 Primers used for qRT-PCR analysis. Primers for qRT-PCR of putative glycogen synthesis
genes.
3.6.5 Biofilm quantification.
A modification of the CV staining method by Sandal et al. 259 was used to quantify biofilm formation. Two
hundred microliters of 0.1% crystal violet were gently added to a tube containing 5 ml of bacterial culture and
incubated at room temperature for 10-15 minutes. Media and CV were removed by pipetting and the tube gently
washed with phosphate buffered saline, pH 7.2 (PBS). To quantify biofilm formation, the CV was solubilized

with 500 μl of 95% ethanol, 200 μl was transferred to a 96-well microtiter plate, and the OD562 was determined
using a Molecular Devices Vmax Kinetic Plate Reader (Molecular Devices; Sunnyvale, CA). At least three
biofilms were tested for each biological replicate.
3.6.6 Capsule quantification.
Isolates and strains were grown in BHI broth to mid-log phase, washed with saline, and resuspended to an
OD562 of 0.7 in 10 ml saline. Bacterial suspensions were incubated at 37oC with 200 μl of 5 mg/ml hyaluronidase
for 30 minutes, then harvested at 10,000 x g for 10 minutes. A uronic acid assay was performed on the
supernatant 310, and quantified in comparison to purified uronic acid control standards. The OD562 was recorded
using a Molecular Devices Vmax Kinetic Plate Reader (Molecular Devices). In addition, isolates were grown
overnight on BHI agar supplemented with 0.005% CR. Bacterial colonies were removed from the agar surface
and suspended in a pre-weighed 1.5-ml tube containing 1 ml of PBS. The tubes were centrifuged at 10,000 x g for
5 min and the PBS discarded. The wet weight of each sample was determined and the bacteria were resuspended
in 1 ml of 1% SDS in PBS, releasing the CR-bound material from the bacterial lysate. Two hundred microliters of
each sample were transferred to a 96-well microtiter plate. The OD490 was determined using a Molecular Devices
Vmax Kinetic Plate Reader (Molecular Devices). The CR absorption index (CRA) was calculated as follows: A490
/ (W2 – W1) = CRA, where W2 refers to the weight of the tube and PBS buffer after the addition of bacteria, and
W1 refers to the weight of the tube and PBS buffer before the addition of bacteria.
3.6.7 Purification of EPS from the biofilm.
Biofilm EPS was extracted from P. multocida as previously described 258 using RPMI-1640 without
phenol red or glutamine (Lonza; Walkersville, MD) as the growth medium. After the biofilm matured, the growth
medium was carefully removed, leaving the biofilm and sediment intact. The purified EPS was lyophilized.

3.6.8 Chemical analysis of EPS.
EPS composition and substitution patterns were determined by analyzing monosaccharides as acetylated
methylglycoside derivatives of the partially methylated and acetylated alditols 328. GC-MS conditions were the
same for each derivative, and were performed on an Agilent Gas Chromatograph 6850A equipped with a SPB-5
capillary column (Supelco, 30 m x 0.25 mm i.d., flow rate 0.8 mL min-1 using He as the gas carrier), interfaced
with a Mass Detector 5973N. Electron impact mass spectra were recorded with an ionization energy of 70 eV and
an ionizing current of 0.2 mA. The temperature program used was set at 150°C for 5 min, 3°C/min up to 280°C,
and 300°C for 5 min. EPS glycogen was quantified using the Anthrone reagent method 329.
Pullulanase (Sigma-Aldrich, P5420, Milan, Italy) digestion prior to NMR spectroscopy was carried out by
adding 20 μl (2 U) of the suspension to the sample, followed by incubation at 37°C overnight. Proton spectra were
recorded in D2O at 298 K with a Bruker 600 MHz equipped with a cryogenic probe; acetone was used as an
internal standard (δH=2.225 ppm; δC=31.45 ppm). For the proton spectrum recorded with deuterated water,
residual solvent peak was reduced by presaturation sequence. For the proton spectrum of the pullulanase digest,
the water peak was suppressed by applying excitation sculpting sequence. All spectra were recorded with 16K
points of resolution, FID were zero-filled to 32 K points and an exponential window function (LB = 0.5) was
applied to enhance signal to noise ratio. The spectra were processed and analyzed using Bruker TopSpin 3.1
software.
3.6.9 LOS purification.
LOS was extracted as described 330, and was examined by sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and silver staining, as previously described 331.

3.6.10 Treatment of growth medium with hyaluronidase, α-amylase, proteinase K, or hyaluronic acid.
To remove capsular material during biofilm formation, RPMI-1640 was supplemented with 0 or 300 units
of hyaluronidase (Sigma-Aldrich; Raleigh, NC) prior to inoculation. In another experiment, hyaluronic acid
(Sigma Aldrich) was added to RPMI-1640 at concentrations of 0, 10, and 100 μg/ml prior to bacterial inoculation
to determine the effect of extracellular CPS on biofilm formation. To assess the presence of carbohydrate or
protein in the biofilm matrix, the growth medium was supplemented with 0.1 mg/ml of α-amylase or Proteinase K
(both from Sigma Aldrich) prior to inoculation. Biofilms were grown and analyzed as described above.
3.6.11 Bacterial hydrophobicity and auto-aggregation.
Bacterial hydrophobicity was determined as described 332, as was auto-aggregation 320 with the following
modifications: P. multocida was suspended in PBS at an OD of 1.0 at the start of the experiment. After 24 and 48
hours, the OD562 of the top 200 μl was recorded.
3.6.12 SEM.
P. multocida strains were grown on glass cover slips in RPMI-1640 medium without phenol red or
glutamine (Lonza; Walkersville, MD) and incubated at 37oC under stationary conditions for 48 hours. The cover
slips were gently washed and fixed in a solution of 5% glutaraldehyde, 4.4% formaldehyde, and 2.75% picric acid
in 0.05% sodium cacodylate buffer for at least 1 hour. Sequential dehydration of the sample was carried out using
25%, 50%, 70%, 80%, and 95% ethanol. SEM was performed as previously described 258 on a Carl Zeiss EVO40
scanning electron microscope.
3.6.13 CLSM.

Biofilms were grown on LabTek II 8 chamber coverglass slides (Thermo Fisher Scientific; Rockford, IL) for
48 hours. Biofilms were gently washed, resuspended in sterile PBS, stained with 1 μl of SYBR live stain,1 μl of
propidium iodide dead stain (Life Technologies; Frederick, MD), and incubated for approximately one hour at
room temperature. CSLM was performed using a ZEISS 880 Laser Scanning Microscope (ZEISS; Germany).
3.6.14 Statistical Analysis.
Determination of median, standard deviation, and p values of comparative values were determined by
unpaired student t test using Excel (Microsoft) or InStat (GraphPad Software, Inc., La Jolla, CA) software. One-
way and 2-way ANOVA tests were performed in Prism GraphPad software version 6.01 (Graphpad Software,
Inc.). Multiple comparisons were performed using the Sidak multiple comparisons test. Correlation of data was
also performed using GraphPad software version 6.01. Significance was considered p ≤ 0.05. Biofilms examined
by CLSM were analyzed using COMSTAT1 software 333.
3.7 ACKNOWLEDGEMENTS
We thank Kathy Lowe for assistance with electron microscopy, Kristi DeCourcy for assistance with CLSM,
and the Genomics Research Laboratory at Virginia Biocomplexity Institute, Virginia Tech, for genome
sequencing.
This work was supported by grant 2013-67015-21314 from the U.S. Department of Agriculture-National
Institute of Food and Agriculture to TJI and funds from the Virginia-Maryland College of Veterinary Medicine.
TJI and BP conceived and designed the experiments. BP carried out most of the experimental work. CD and
AM performed the carbohydrate chemistry. AD contributed to bioinformatic analysis. REB performed the allelic
exchange mutagenesis experiments. BP and WES carried out the confocal microscopy. BP, TJI, CD, AD, and
REB contributed to writing the manuscript. TJI and BP reviewed and performed final edits of the manuscript.

The funders had no role in study design, data collection and interpretation, or the decision to submit the work
for publication.

CHAPTER 4
Chemical mutagenesis of Pasteurella multocida P1059 reveals the importance of P1059_01979 in capsular
polysaccharide production and biofilm formation.
Petruzzi, B., Dickerman, A., Inzana, T.

4.1 ABSTRACT
Pasteurella multocida is a gram-negative, zoonotic pathogen of veterinary importance. Capsular
polysaccharide (CPS) production by P. multocida is an important virulence factor that inhibits phagocytosis by
the host. Additionally, biofilm formation is a virulence factor that is inhibited by CPS production. Chemical
mutagenesis was performed on P. multocida strain P1059, and an acapsular mutant designated as P1059-R8 was
identified by its acapsular appearance on dextrose starch agar supplemented with congo red. Genome sequencing
revealed that genes in the CPS biosynthesis locus and known CPS regulators were not affected by the chemical
exposure. However, a uridyltransferase encoded by gene P1059_01979 was mutated in such a way that a polar
amino acid was changed to a non-polar amino acid near the active site. The protein product of P1059_01979 is
important for the biosynthesis of the CPS subunit N-acetylglucosamine. CPS quantification revealed that the
subunit glucuronic acid was produced in equal concentrations to the parent. However, N-acetylglucosamine was
detected in the parent but not detected in P1059-R8. Biofilm formation by P1059-R8 was significantly higher than
the parent strain P1059. We hypothesize that P1059_01979 is essential for CPS production in P. multocida
serogroup A, and supports that CPS production inhibits biofilm formation in P. multocida.
4.2 INTRODUCTION
Pasteurella multocida is a gram-negative zoonotic pathogen of the Pasteurellaceae family. P. multocida
can be highly virulent, and is most commonly associated with diseases of veterinary importance such as avian
cholera, bovine respiratory disease, porcine respiratory disease, and hemorrhagic septicemia. Capsular
polysaccharide (CPS) is considered the most important virulence factor for Pasteurella multocida, and has been
used as a vaccine target 334. However, vaccines against CPS poorly protect against challenge 61, 62. The CPS is
non-immunogenic, and camouflages P. multocida from the host immune response. Acapsular P. multocida are
readily phagocytized by macrophages in vitro 32, 41, 282. CPS has specifically been shown to inhibit phagocytosis
of serogroup A strains by neutrophils 60.

Bacterial CPSs are an extracellular component essential for survival within the host. CPSs are composed of
long chain polysaccharides that are usually negatively charged and hydrated. It is common for bacterial species to
produce several different CPS structures, which are often reflected in their serotyping scheme 335. P. mutocida has
5 distinct CPS serogroups, designated as A, B, D, E, and F 32, 34, 284. While each serogroup has a distinct CPS
composition, all 5 CPSs are composed of glycosaminoglycans (GAGs). GAGs are long, unbranched
polysaccharides composed of a dimer repeating unit usually consisting of an amino sugar and uronic sugar. CPS
serogroup A is composed of hyaluronic acid (HA). HA is the simplest GAG, and is composed of repeating N-
acetylglucosamine (GlcNAc) and glucuronic acid (GlcA) dimers. Some GAGs, such as chondroitin and heparin,
are sulfated 32.
Biofilm formation is another virulence factor of P. multocida and other veterinary pathogens such as
Histophilus somni 258, 259, 336, 337. Like CPS, the biofilm matrix is composed of an exopolysaccharide (EPS) that
protects bacteria from the host immune response. The resulting protection allows P. multocida to persist in the
host for weeks at a time without detection. Biofilm formation in P. multocida has been demonstrated for bovine
pneumonia and avian cholera isolates201, and characterized elsewhere (Petruzzi et al., 2017, in press). In P.
multocida, biofilm formation is inversely related to CPS production. While CPS is important for evading the host
immune responses and is associated with high virulence, biofilm production is associated with chronic infections
that may be asymptomatic 32, 140, 282.
Pasteurella multocida P1059 was isolated from a turkey that succumbed to acute avian cholera. An acapsular
mutant (described in chapter 3) of P. multocida P1059 showed reduced CPS and increased biofilm production.
Similarly, this report details a chemical mutant of P. multocida P1059 that displays a reduced CPS phenotype and
an increased biofilm phenotype. In this mutant, P1059-R8, no genes in the CPS locus or known regulators of CPS
production were mutated. Alternatively, a gene important for enzymatically altering a CPS precursor
monosaccharide putatively lost its function, resulting in an acapsular phenotype.

4.3 MATERIALS & METHODS
4.3.1 P. multocida and growth conditions. P. multocida strain P1059 was cultured on Brain Heart
Infusion (BHI) or Columbia blood agar (BD; Franklin Lakes, NJ) supplemented with 5% defibrinated sheep blood
(HemoStat Laboratories Inc.; Dixon, CA), or Dextrose Starch agar (DSA) supplemented with or without 0.005%
Congo Red (CR). Agar-grown cultures were grown at 37oC with 6% CO2. Broth cultures were grown in BHI or
RPMI-1640 medium without glutamine or phenol red (Lonza; Walkersville, MD). For biofilm formation, 50 μl of
mid-log phase P. multocida was inoculated into 5 ml of RPMI-1640 in a 50-ml polyethylene tube, and incubated
stationary at 37oC in 6% CO2 for at least 48 hours.
4.3.2 Isolation of a chemically-modified P. multocida mutant. Chemical mutagenesis of strain P1059
was performed using the method from Adelberg et al. 338. After mutagenesis, the cells were suspended in cold
RPMI-1640 medium lacking phenol red or glutamine (Lonza; Walkersville, MD), and grown on DSA containing
0.005% CR. A CPS-deficient mutant was selected based on the uptake of CR and colony morphology; this isolate
was designated P1059-R8. Comparative growth curves of the parent and mutant were similar, indicating that any
mutations present did not affect growth rate.
4.3.3 Genome sequencing and analysis of mutant P1059-R8.
P1059-R8 genomic DNA was isolated using the Qiagen DNeasy kit (Germantown, MD) according to the
manufacturer’s instructions. DNA sequencing of P1059-R8 was carried out at the Biocomplexity Institute at
Virginia Tech (Blacksburg, VA) using an Illumina MiSeq that generated >15 million paired-end 250 bp quality-
filtered reads. The sequence data was assembled into a single contig of 2307905 base pairs using the iterative
method of Tae 339. To identify any mutations generated by the mutagenesis, the genome sequence of P. multocida
subsp. gallicida P1059 (RefSeq NZ_CM001581.1) was aligned to the assembled mutant genome using Mauve 340,

revealing 2014 single nucleotide polymorphisms (SNPs) and 117 indels. These mutations impinged on 51 distinct
protein coding regions, one tRNA (Glu) and two pseudogenes, altering the protein translation of 45 proteins
(Table 4.2).
4.3.4 Biofilm quantification.
Crystal violet (CV) staining was used to quantify biofilm formation, modified from Sandal et al. 259. Two
hundred microliters of 0.1% crystal violet were gently added to 5 ml of biofilm culture in a 50-ml conical tube
and incubated at room temperature for 10-15 minutes. Media and CV were removed by pipetting and the tube
gently washed with phosphate buffered saline, pH 7.2 (PBS). To quantify biofilm formation, 500 μl of 95%
ethanol was added to solubilize the CV, 200 μl was transferred to a 96-well microtiter plate, and the OD562 was
determined using a Molecular Devices Vmax Kinetic Plate Reader (Molecular Devices; Sunnyvale, CA). Biofilms
were grown at least in triplicate for each biological replicate.
4.3.5 CPS quantification using uronic acid and N-acetylglucosamine chemical assays.
Isolates and strains were grown in BHI broth to mid-log phase, washed with saline, and resuspended to an
OD562 of 0.7 in 10 ml saline. Bacterial suspensions were incubated at 37oC with 200 μl of 5 mg/ml hyaluronidase
for 30 minutes, then harvested at 10,000 x g for 10 minutes. A uronic acid assay 310 and N-acetylglucosamine
assay 341 was performed on the resulting supernatant, including purified uronic acid control standards. The OD562
was recorded using a Molecular Devices Vmax Kinetic Plate Reader (Molecular Devices; Sunnyvale, CA).
4.3.6 Bacterial hydrophobicity and auto-aggregation.

Bacterial hydrophobicity was determined as described 332, as was auto-aggregation 320 with the following
modifications: P. multocida was suspended in PBS at an OD of 1.0 at the start of the experiment. After 24 and 48
hours, the OD562 of the top 200 μl was recorded.
4.3.7 Scanning Electron Microscopy.
P. multocida strains were grown on glass cover slips in RPMI-1640 medium without phenol red or glutamine
(Lonza; Walkersville, MD) and incubated at 37oC under stationary conditions for 48 hours. The cover slips were
gently washed and fixed in a solution of 5% glutaraldehyde, 4.4% formaldehyde, and 2.75% picric acid in 0.05%
sodium cacodylate buffer for at least 1 hour. Step-wise dehydrations were carried out using 25%, 50%, 70%,
80%, and 95% ethanol. SEM was performed as previously described 258 on a Carl Zeiss EVO40 scanning electron
microscope.
4.3.8 Confocal laser scanning microscopy (CLSM).
Biofilms were grown on LabTek II 8 chamber coverglass slides (Thermo Fisher Scientific; Rockford, IL) for
48 hours. Biofilms were gently washed, resuspended in sterile PBS, stained with 1 μl of SYBR live stain,1 μl of
propidium iodide dead stain (Life Technologies; Frederick, MD), and incubated for approximately one hour at
room temperature. CSLM was performed using a ZEISS 880 Laser Scanning Microscope (ZEISS; Germany).
4.3.10 Statistical analysis.
One-way and 2-way ANOVA tests were performed using Prism GraphPad software version 6.01
(Graphpad Software, Inc.; La Jolla, CA USA). Multiple comparisons were performed using the Sidak multiple
comparisons test. Correlation of data were also performed using GraphPad software version 6.01. Significance

was considered p ≤ 0.05. The software tool Protein Variation Effect Analyzer (PROVEAN; J. Craig Venter
Institute, USA) (http://provean.jcvi.org/index.php), was used to predict whether an amino acid substitution or
deletion would deleteriously affect protein function. A significant deletion or substitution is a PROVEAN score
less than -2.5. Biofilms were analyzed using COMSTAT1 software 333.
4.4 RESULTS
4.4.1 Isolation of a CPS-deficient, biofilm-proficient mutant.
Dextrose agar plates containing congo red were examined for colonies displaying an acapsular, or rough,
morphology after chemical mutagenesis. One colony appeared dark red in color and had a rough morphology.
This colony was isolated and named P1059-R8. The growth rate of P1059-R8 was similar to parent strain P1059.
4.4.2 Characterization of P1059-R8 by genome sequencing and analysis.
DNA sequencing of P1059-R8 identified 45 mutations that resulted in a change, deletion, or insertion of
an amino acid, or a basepair change into a termination mutation throughout the P1059 genome (Table 4.1).
PROVEAN statistical analysis revealed 20 mutations resulting in an amino acid change potentially deleterious to
protein function in 19 genes (two mutations occurred in the multimodular transpeptidase-transglycosylase). None
of the mutations occurred in the CPS locus or within known regulatory genes. The proteins affected are listed with
their putative function, the mutation, and PROVEAN score in Table 4.2. Of interest was gene P1059_01979,
which encodes an N-acetylglucosamine-1-phosphate uridyltransferase. This enzyme is responsible for the last
phosphorylation step in the production of UDP-N-acetylglucosamine, a monosaccharide subunit of HA.
RefSeqID Description

P1059_00035 L-serine dehydratase
P1059_00067 protein YaaA
P1059_00126 Aspartokinase
P1059_00127 Homoserine kinase
P1059_00130 HflK protein
P1059_00137 Arginine ABC transporter, periplasmic arginine-binding protein ArtI
P1059_00257 Ribonuclease
P1059_00296 L,D-transpeptidase YcbB
P1059_00704 hypothetical protein
P1059_00771 Riboflavin synthase eubacterial/eukaryotic
P1059_00819 Molybdopterin biosynthesis protein MoeA
P1059_00839 Autotransporter adhesin
P1059_00877 NAD(P) transhydrogenase subunit beta
P1059_00886 Translation initiation factor 2
P1059_00896 HrpA-like helicase
P1059_00918 Selenide,water dikinase
P1059_00932 putative zinc protease pqqL
P1059_00949 Hypothetical protein
P1059_00970 DNA gyrase subunit A
P1059_00999 Fructose repressor FruR, LacI family
P1059_01105 18K peptidoglycan-associated outer

P1059_01264 Polyribonucleotide nucleotidyltransferase
P1059_01271 Mannosyltransferase OCH1 like enzyme
P1059_01295 Hypothetical protein
P1059_01320 Membrane protein, putative
P1059_01332 Hypothetical protein
P1059_01330 Glycerol-3-phosphate acyltransferase
P1059_01380 Multimodular transpeptidase-transglycosylase
P1059_01482 Na(+)-translocating NADH-quinone reductase subunit B
P1059_01553 Biosynthetic arginine decarboxylase
P1059_01586 SSU ribosomal protein S10p (S20e)
P1059_01589 tRNA dihydrouridine synthase A
P1059_1599 ATP-dependent DNA helicase RecQ
P1059_01663 ATP synthase delta chain
P1059_01696 Transcriptional activator of 4-hydroxyphenylacetate 3-monooxygenase
operon, XylS/AraC family
P1059_01707 2,4-dihydroxyhept-2-ene-1,7-dioic acid aldolase
P1059_01708 Succinate-semialdehyde dehydrogenase [NADP+]
P1059_01714 33 kDa chaperonin, Heat shock protein 33
P1059_01746 Transcriptional regulator, RpiR family protein
P1059_01965 Trimethylamine-N-oxide reductase
P1059_01979 N-acetylglucosamine-1-phosphate uridyltransferase
P1059_02008 Coproporphyrinogen III oxidase, oxygen-independent

P1059_02016 ATP-dependent RNA helicase SrmB
P1059_02034 Phosphoglycerate kinase
P1059_021100 Excinuclease ABC subunit A
Table 4.1: Genes within the P1059-R8 genome with a SNP resulting from chemical mutagenesis.
Determined by genome sequencing of P1059-R8 in comparison to the P1059 genome.

Table 4.2: Putative malfunctioning proteins due to SNPs. PROVEAN statistical analysis was used to
determine the effect of a SNP on protein function. PROVEAN scores of -2.5 or less were considered
significant.
4.4.3 Biofilm Quantification by crystal violet staining.

The amount of biofilm formed was determined with crystal violet to stain the biofilm matrix. Strain
P1059 made a poor biofilm, which is consistent with the data presented for P1059 in Chapter 3, whereas mutant
P1059-R8 made significantly more biofilm material (Figure 4.1).
Figure 4.1: Quantification of biofilm matrix by crystal violet staining. The biofilm matrix was stained with
crystal violet, suspended with ethanol, and the optical density determined.
4.4.4 CPS quantification using uronic acid and N-acetylglucosamine chemical assays.
CPS was hydrolyzed using hyaluronidase, which removed the CPS subunits from the cell surface. The
individual HA subunits, GlcNAc and GlcA were quantified using chemical assays. The amount of GlcA produced
by P1059 and P1059-R8 were not significantly different (Figure 4.2 A). However, the amount of GlcNAc
produced by P1059-R8 was significantly less than that produced by P1059 (p < 0.0001) (Figure 4.2 B).

Figure 4.2: Quantification of capsular Glucuronic acid and N-acetyl-glucosamine. CPS serogroup A is
composed of hyaluronic acid (HA). HA is a dimer of glucuronic acid and N-Acetylglucosamine. The subunits
were separated by enzymatic digestion and quantified individually: Glucuronic acid (GlcA) (A); N-
Acetylglucosamine (GlcNAc) (B). P1059 and P1059-R8 were compared to CPS-deficient P1059ΔhyaE as a
control.
4.4.5 Confocal Scanning Laser Microscopy of live/dead stained biofilms.
Biofilms of P1059, P1059ΔhyaE, and P1059-R8 were grown and dyed with SYBR stain and propidium
iodide to determine live and dead cells, respectively, within the biofilm matrix. SYBR stained confocal z-stacks
were analyzed using COMSTAT programming 333. Mutant P1059-R8 biofilm had significantly more biomass than

P1059 and P1059ΔhyaE CPS-deficient mutant, and was also significantly thicker. P1059 did not appear to make a
biofilm, but rather a monolayer of cells adhered to the glass coverslip (Figure 4.2). The R8 mutant also had a
significantly smaller surface area to biovolume ratio. The roughness coefficient indicated that the P1059-R8
biofilm was more uniform in thickness and topography than the P1059ΔhyaE biofilm, which varied in thickness
(Table 4.3).
Figure 4.3: CSLM of P1059 and mutant biofilms: Z-stacks of biofilms grown on glass cover slips, and dyed
with SYBR (live-determining stain) and propidium iodide (dead-determining stain). SYBR stain: P1059 (A),
P1059ΔhyaE (D), P1059-R8 (G). Propidium iodide stain: P1059 (B), P1059ΔhyaE (E), P1059-R8 (H), Merge:
P1059 (C), P1059ΔhyaE (F), P1059-R8 (I).
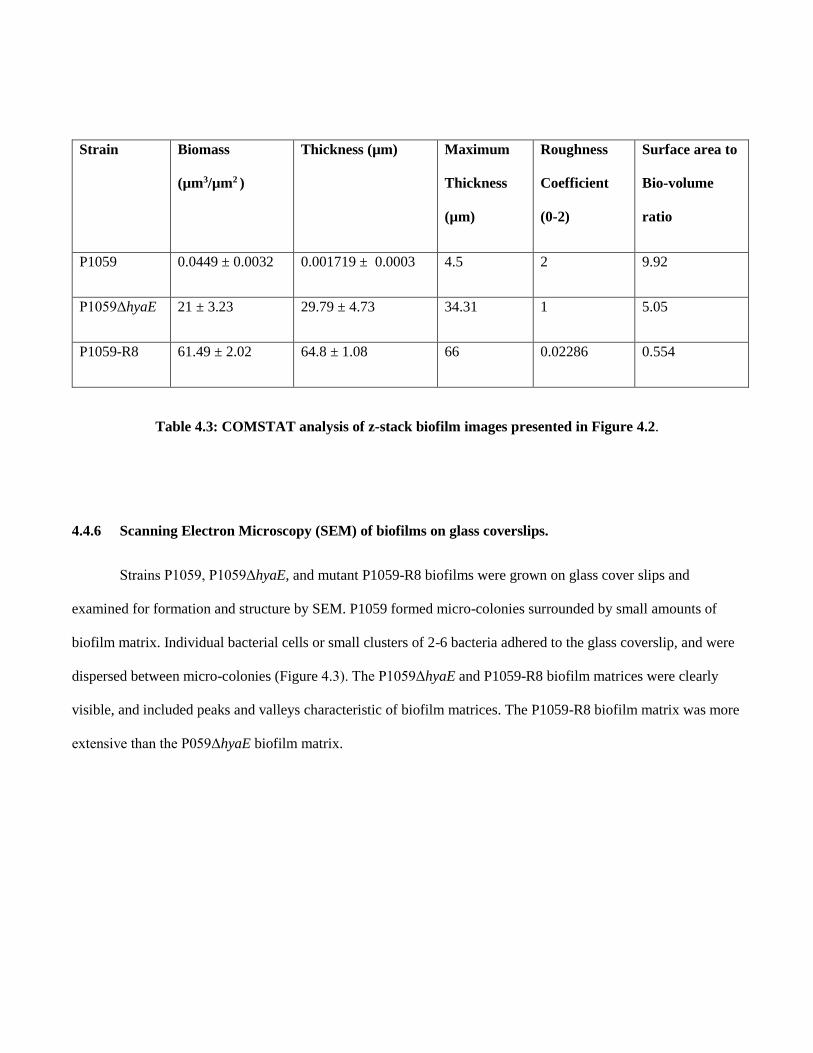
Strain Biomass
(µm3/µm2 )
Thickness (µm) Maximum
Thickness
(µm)
Roughness
Coefficient
(0-2)
Surface area to
Bio-volume
ratio
P1059 0.0449 ± 0.0032 0.001719 ± 0.0003 4.5 2 9.92
P1059ΔhyaE 21 ± 3.23 29.79 ± 4.73 34.31 1 5.05
P1059-R8 61.49 ± 2.02 64.8 ± 1.08 66 0.02286 0.554
Table 4.3: COMSTAT analysis of z-stack biofilm images presented in Figure 4.2.
4.4.6 Scanning Electron Microscopy (SEM) of biofilms on glass coverslips.
Strains P1059, P1059ΔhyaE, and mutant P1059-R8 biofilms were grown on glass cover slips and
examined for formation and structure by SEM. P1059 formed micro-colonies surrounded by small amounts of
biofilm matrix. Individual bacterial cells or small clusters of 2-6 bacteria adhered to the glass coverslip, and were
dispersed between micro-colonies (Figure 4.3). The P1059ΔhyaE and P1059-R8 biofilm matrices were clearly
visible, and included peaks and valleys characteristic of biofilm matrices. The P1059-R8 biofilm matrix was more
extensive than the P059ΔhyaE biofilm matrix.

Figure 4.4: SEM of biofilm matrix formed on glass coverslips. Biofilms were grown on glass coverslips for 48
hours, then imaged using scanning electron microscopy (SEM); A, P1059; B, P1059ΔhyaE (control); C, P1059-
R8.
4.4.7 Determination of hydrophobicity and auto-aggregation. Previous studies have shown that P.
multocida can display large quantities of lipids on its surface, causing the bacteria to become hydrophobic 316, 317,
and increasing the adherence of the bacteria to surfaces 318; an essential early step in biofilm formation. Strain
P1059 and P1059ΔhyaE did not enter the organic phase during the hydrophobicity assay, but 76% of P1059-R8
cells did move to the organic phase. Hydrophobicity of the cellular surface did not appear to contribute to biofilm
formation in P. multocida P1059ΔhyaE. However, mutant P1059-R8 exhibited strong surface hydrophobicity.
Auto-aggregation can also contribute to biofilm formation, and is often facilitated by EPS and/or bacterial
adhesins 319, 320. After 24 hours, 20% of the P1059 and P1059ΔhyaE cell suspensions had aggregated. No
differences in auto-aggregation were noted between the isogenic mutant and parent, indicating that cell-to-cell
interactions were not inhibited by CPS, and auto-aggregation was not a good predictor of biofilm formation.
However, suspensions of strain P1059-R8 cells aggregated 50% after 24 hours. No further aggregation was
observed beyond 24 hours for any strain. In contrast, suspensions of the H. somni biofilm control strains 2336 and
129Pt (which do not form a CPS), aggregated greater than 75%.
4.5 DISCUSSION

The CPS synthesis locus of P. multocida has been well characterized 41. Here, a chemical mutant of strain
P1059, named P1059-R8, has been described. Chemical mutagenesis is a useful tool for discovering genes
associated with a particular phenotype, particularly when allelic exchange or transposon mutagenesis is difficult.
In this report, chemical mutagenesis was performed to assess the relationship between CPS and biofilm formation.
Mutant P1059-R8 was deficient in CPS production, but produced more biofilm than P1059 or CPS-deficient
P1059ΔhyaE. P1059-R8 was identified by its acapsular appearance on agar, and its ability to take up larger
amounts of congo red in comparison to the P1059 and CPS-deficient P1059ΔhyaE. Congo red can be used to
identify CPS deficiency because it binds to hydrophobic substances or neutral glycoses (311). Genome sequencing
of P1059-R8 revealed 45 SNPs, but none of the SNPs were in the CPS synthesis locus or any known regulators.
SNPs that result in amino acid changes deleterious to protein function were identified using statistical models.
P1059_01979 encodes a bifunctional enzyme, GlcNAc-1-P uridyltransferase, which converts GlcNAc-1-P to
UDP-GlcNAc. The product is a substrate for hyaluronic acid synthase 342, 343. P1059_01979 has a SNP which
caused an isoleucine to replace a threonine at position 116. Since threonine is a polar amino acid and isoleucine is
a non-polar amino acid, it is likely that the protein structure, and consequently its function, is modified by this
SNP. While the PROVEAN cut-off for SNPs with significant modifications affecting protein function is -2.5, the
SNP in P1059_01979 had a PROVEAN statistical score of -2.483. Because this value can be rounded to -2.5, and
may explain the acapsular phenotype of this mutant, it has been included in the table of statistically relevant
SNPs.
P. multocida GlcNAc-1-P Uridyltransferase (product of P1059_01979) shares 69% identity with Escherichia
coli GlmU 344, 345. E. coli GlmU is a bifunctional enzyme that catalyzes the two biosynthetic steps essential for
UDP-N-acetyl-glucosamine synthesis. Since this enzyme is bifunctional, it has two active sites. The first active
site is in the C-terminal domain, and has acetyltransferase activity. The second active site is in the N-terminal
domain, and catalyzes the final phosphorylation step in UDP-N-acetylglucosamine synthesis. The second active
site is formed by a pocket bound by two lobes from Asn3 – Val111, and His216 – Asn227 344, 346. Essential to
protein function is a hydrogen bond between Asp114, Ser112, and Lys22. Since amino acid 116 has been changed

from polar threonine to non-polar isoleucine, we suspect that the phosphorylase function of the protein encoded
by P1059_01979 is inhibited. These residues are highly conserved among GlmU sequences in other bacterial
species, including E. coli, Mycobacterium tuberculosis, Haemophilus influenzae 347, and Streptococcus
pneumoniae 346, 348.
UDP-N-acetylglucosamine is essential for hyaluronic acid synthesis, and we hypothesize that the
loss of phosphorylase function explains the acapsular phenotype of P1059-R8. To date, no other proteins
encoded in the P1059 genome perform functions redundant to GlcNAc-1-P uridyltransferase (data not
shown).
CPS was quantified by measuring the glucuronic acid and N-Acetylglucosamine concentrations in the
supernatant of a hyaluronidase-treated bacterial suspensions. Hyaluronic acid biosynthesis is catalyzed from
UDP-glucuronic acid and UDP-N-acetylglucosamine subunits 343. P1059-R8 made glucuronic acid in similar
quantities as P1059. However, N-acetylglucosamine was not present in CPS digests of chemical mutant P1059-
R8. We can conclude that N-acetylglucosamine biosynthesis is inhibited by the SNP in P1059_01979. Glucuronic
acid was detected in enzymatic digests of the CPS, indicating that glucuronic acid biosynthesis is not inhibited.
P1059-R8 produces sufficient amounts of UDP-glucuronic acid, but does not have the ability to create UDP-N-
acetylglucosamine to complete hyaluronic acid synthesis.
However, it remains undetermined whether other proteins containing statistically relevant SNPs were
contributing to this phenotype. The lack of CPS on the surface of the bacteria due to the invert P1059_01979
knocked out phenotype likely explains increased hydrophobicity and autoaggregation. The parent and CPS-
deficient mutant P1059ΔhyaE were not hydrophobic, and did not auto-aggregate to the extent of the chemical
mutant. However, P1059ΔhyaE still produces decreased amount of CPS compared to P1059, and is not truly
acapsular. Isogenic mutagenesis will need to be performed in order to determine the phenotype of a P1059_01979
knock out P. multocida mutant.

The CPS biosynthesis deficiency in P1059-R8 resulted in increased biofilm formation. Biofilm formation was
observed through crystal violet staining, as well as several microscopy techniques. Increased biofilm formation in
comparison to the parent and CPS-deficient mutant P1059ΔhyaE could be observed by SEM, and quantified using
CSLM z-stack images.
In conclusion, we suspect that the protein product of gene P1059_01979 is essential for CPS production in P.
multocida. The absence of CPS in mutant P1059-R8 further supports that CPS inhibits biofilm formation in P.
multocida.

CHAPTER 5
Polymicobial Interaction between Histophilus somni and Pasteurella multocida during Biofilm Formation
Petruzzi, B.; Lahmers, K.; Huang, S.; Scarratt, W. K.; Inzana, T.

5.1 ABSTRACT
Histophilus somni and Pasteurella multocida cause bovine respiratory disease (BRD) and systemic
infections in cattle. Following respiratory infection of calves with H. somni, P. multocida may also be isolated
from the lower respiratory tract. Because H. somni normally forms a biofilm during BRD, P. multocida may co-
exist with H. somni in a polymicrobial biofilm. We sought to examine the interactive nature of the two species
during biofilm formation in vitro and in vivo. Interactions between the two species in the biofilm were
characterized and quantified by fluorescence in situ hybridization (FISH), and the biofilm matrix of each species
examined by fluorescently-tagged lectins (FTL), confocal scanning laser microscopy of in vitro biofilms and
bovine pulmonary tissue following dual H. somni and P. multocida infection. Bacterial interactions were
determined by auto-aggregation and biofilm morphology. FISH and FTL were used to show that P.
multocida and H. somni were evenly distributed in the in vitro biofilm, and both species contributed to the
polymicrobial biofilm matrix. COMSTAT z-stack image analysis revealed that the average biomass and biofilm
thickness, and the total carbohydrate and protein content of the biofilm, were greatest when both species were
present. Polymicrobial bacterial suspensions auto-aggregated faster than single species suspensions, suggesting
physical interactions between the two species. Encapsulated P. multocida isolates not capable of forming a
biofilm still formed a polymicrobial biofilm with H. somni, but only the EPS of H. somni could be detected by
FTL staining of bovine tissues from which both species were isolated. Bacteria within a biofilm are more
quiescent than during planktonic growth and induce less of an inflammatory response, indicating encapsulated P.
multocida may take advantage of the H. somni biofilm to persist in the host during less severe, but more chronic,
BRD. These results may have important implications for the management of BRD.
5.2 INTRODUCTION
Bovine respiratory disease (BRD) complex causes significant economic losses to the beef industry through
increased prevention and treatment costs, reduced carcass value due to treatment and prevention measures,

morbidity, and mortality 184, 185. BRD is a collective term describing respiratory infections from causative agents
including the bacterial species Histophilus somni, Pasteurella multocida, Mannheimia haemolytica, and
Mycoplasma bovis 191. It is common to detect more than one causative agent during an outbreak 194, 195, 248,
suggesting BRD is often polymicrobial.
A definitive diagnosis can only be made after post-mortem examination, resulting in broad, poorly defined
diagnostic criteria 349. Signs of disease can go undetected, and include a lack of appetite, nasal discharge,
coughing, rapid breathing, fever, and diarrhea. Polymicrobial infections can display a more complex pathology
than single species infections 196-198, and are more resistant to antibiotics 350. Cattle are predisposed to disease after
experiencing stress, including situations such as weaning, excessive handling, a change in diet, and transportation
or exposure to new locations and/or herds 177, 178. Genetic factors may also predispose cattle to disease 178. It is
generally accepted that transmission occurs via contaminated aerosols in locations with limited ventilation such as
during transportation, in auction houses or crowded barns, or after a change in climate or location.
The most common bacterial agents responsible for BRD have been demonstrated to produce biofilms,
which may be associated with their pathogenic role during BRD 120, 199, 200. H. somni biofilm formation has been
directly associated with BRD 120. Bacterial populations within a biofilm actively pursue other bacterial species,
leading us to hypothesize that polymicrobial bacterial infections during BRD exist in a biofilm 202. An
experimental bovine respiratory infection performed by our lab indicated that biofilms of H. somni in the
cardiopulmonary tissue of infected calves also contained small numbers of P. multocida. Prior health screenings
indicated the calves had not been exposed to P. multocida 120. In addition, a recent report of BRD described
polymicrobial infections with H. somni and P. multocida 193. Research in this field over the past 4 decades has
aided in the reduction of disease incidence and has promoted the implementation of preventative techniques by
veterinarians. Despite progress, BRD is still the largest cause of morbidity and mortality in the beef industry,
indicating a need for further research 184, 349, 351.

The work described in this report aims to characterize the coaggregative polymicrobial relationship
between the BRD pathogens H. somni and P. multocida during biofilm formation. Novel microscopy techniques
have allowed each species to be observed simultaneously within the biofilm matrix, providing a detailed
understanding of their polymicrobial relationship. Additionally, data obtained from an experimental infection in
calves supported the occurrence of H. somni and P. multocida polymicrobial biofilms in vivo. The information
presented here further supports the role of biofilm during BRD, provides evidence that BRD is a complex
polymicrobial disease, and supports the need for better treatment and preventative measures.
5.3 MATERIALS & METHODS
5.3.1 Bacterial growth.
Histophilus somni 2336 and Pasteurella multocida strain C0513 and passed variant C0513-P5 were used.
Bacterial strains were grown in BHI Broth (BD Scientific; Irvine, CA) containing 0.5% Yeast extract (BD
Scientific; Irvine, CA), 0.1% Trizma base, 0.01% thiamine monophosphate (TMP) (Sigma; St. Louis, MO), and
1% bovine serum at 37oC with rapid shaking until H. somni reached 135 Klett units, P. multocida C0513 reached
99 Klett units, and P. multocida C0513-P5 reached 120 Klett units (mid-log phase, 109 CFU). Biofilms were
grown in 50mL conical tubes using the same media at 37oC with shaking at 50 rpm or less. H. somni biofilms
were grown to maturity, approximately 3 days 258 before the addition of P. multocida for polymicrobial biofilm
growth. After the addition of P. multocida, biofilms continued to grow for 2 days, until P. multocida biofilm
maturity. Single species biofilms were grown for 5 days for H. somni, and 2 days for P. multocida.
5.3.2 Fluorescent in situ hybridization.
Fluorescent in situ hybridization (F.I.S.H.) was performed using the following probes: ‘/5FluorT/TT
AAG AGA TTA ATT GAT TGA’ to detect H. somni, and ‘/5Cy5/CT ATT TAA CAA CAT CCC TTC’ to detect

P. multocida. DNA-specific probes were purchased from Integrated DNA Technologies Inc. (Coralville, IA).
Probes were determined to be species-specific using a BLAST search and through experimental hybridization.
F.I.S.H. was carried out as described 352, with some modifications. The hybridization buffer determined to provide
the best attachment for both probes contained 25% formamide. Biofilms were grown on glass coverslips and fixed
with 4% paraformaldehyde for at least 60 minutes. Biofilms were incubated with hybridization buffer for at least
15 minutes at 37oC. After pre-incubation, 0.2 μg of each probe was added to the fixed biofilm, and incubated
overnight at 37oC. Biofilms were then washed 1x with wash buffer, then rinsed thoroughly with water. Glass
coverslips and slides were embedded biofilm-side down with 20 μl [poly]vinyl alcohol (Sigma-Aldrich; St. Louis,
MO) onto glass slides, and stored in the dark. Confocal scanning laser microscopy was performed on a Zeiss 880
Confocal microscope (Zeiss; Oberkochen, Germany) at 40x magnification using Cy5 (deep red, 660-710nm) and
fluorescein (green, 500-540 nm) filters.
5.3.3 Fluorescent lectin staining of exopolysaccharide material.
FITC-conjugated Griffonia simplicifolia lectin (GS-II) (EY laboratories; San Mateo, CA) was used to
detect glycogen EPS produced by P. multocida 353, 354. TRITC-conjugated Moringa M Lectin (MNA-M) (EY
laboratories) was used to detect the H. somni galactomannan EPS 120. Lectins were suspended in a 10 mM
phosphate buffer, pH 7.5, at a concentration of 10 μg/ml. Fifty microliters of lectin solution was applied to
biofilms and incubated 30-60 minutes in the dark at room temperature, then washed with phosphate buffer 4
times. Coverslips were fixed onto slides with 20 μl of [poly]vinyl alcohol (Sigma-Aldrich; St. Louis, MO) and
stored in the dark for at least 6 hours before imaging. Confocal scanning laser microscopy was performed on a
Zeiss 880 Confocal microscope (Zeiss,) at 40x magnification using TRITC (red, 532 nm) and fluorescein (green,
500-540nm) filters.
5.3.4 Polymerase Chain Reaction.

The primers specific to P. multocida and H. somni that were used in this study are described elsewhere 43,
355. Isolated colonies were suspended in sterile water and boiled for 10 minutes. Suspensions were centrifuged at
12000 rpm for 5 minutes. The supernatant was used as a DNA template for PCR reactions. DNA was extracted
from sections of lung tissue using the Blood and Tissue DNeasy Kit (Qiagen, Germantown, MD) according to
manufacturer’s instructions. PCR was performed on extracted DNA material using OneTaq Kit (NEB, Ipswich,
MA), according to manufacturer’s instructions.
5.3.5 Enzyme-linked immunosorbent assay.
Enzyme-linked immunosorbent assays (ELISA) were performed to determine the antibody response to H.
somni whole cells, P. multocida whole cells, or crude single species biofilm material. Twenty micrograms of
biofilm material per well was used to coat the surface of Immulon 4HBX plates (ThermoFisher, Waltham, MA).
Bacteria were grown to 109 CFU/ml (mid-log phase), then resuspended in carbonate coating buffer, pH 9.6 (15
mM Na2CO3, 35 mM NaHCO3, 3 mM NaN3) to the same concentration. Wells were coated with 108 CFU of
whole cells. Phosphate buffered saline (PBS) supplemented with 0.05% Tween 20 (Sigma,St. Louis, MO) and 2%
non-fat dry milk (Kroger,) was used as a blocking agent. Primary antibody used was calf serum separated from
whole blood at the time of euthanasia. The secondary antibody used was rabbit IgG to bovine heavy & light chain
conjugated to horseradish peroxidase (Jackson laboratory; Bar Harbor, ME). Immunosorbent reactions were
developed using 3,3’,5,5’-Tetramethylbenzidine (TMB) substrate (Pierce Biotechnology; Rockford, IL).
5.3.6 Biofilm protein and carbohydrate concentrations.
Biofilm protein concentration was determined using a BCA assay (ThermoFisher, Waltham, MA).
Carbohydrate concentrations were determined using the Anthrone assay 356, 357. Biofilm material was suspended in

1 ml PBS, and vortexed rapidly for several seconds to break up the biofilm matrix. Suspensions were then used in
BCA and Anthrone assays.
5.3.7 Auto-aggregation of single species and polymicrobial suspensions.
Auto-aggregation was performed as previously described 320 with modifications. Bacterial cells from P.
multocida C0513, C0513-P5, H. somni 2336, or in combination were suspended in sterile PBS with or without
supplementation with 100 μg/mL H. somni crude biofilm material. OD562 measurements were taken prior to the
start of the experiment, after 1 hour, and after 24 hours. Starting OD562 was between 0.8-1. Aggregation was
determined by the percentage of cells still in suspension after a certain period of time.
5.3.8 Determination of polymicrobial biofilm formation in vivo.
Two 8-week old male Holstein calves were challenged with 109 CFU of H. somni strain H.S.25 in 10 ml
sterile phosphate buffered saline containing calcium and magnesium intratracheally using a syringe (6-12 cc) and
a Tom Cat/Small Animal Catheter (3.5 Fr, 5.5 inches long). Calves were monitored daily for signs of respiratory
infection. Twenty-two days after challenge, calves were euthanized, and lungs were harvested for further analysis.
Bacterial load within the pulmonary tissue was determined by manually breaking down lung tissue in sterile PCM
buffer. The resulting digest was serially diluted. Dilutions were spread onto Columbia blood agar and incubated
for 72 hours at 37oC in 6% CO2. The presence of H. somni and P. multocida was determined by colony
morphology upon isolation and subsequent PCR of isolated colonies and lung tissue. Tissue sections were
embedded in paraffin and cut into 5 μm sections on glass slides. The slides were deparaffinated and stained by
F.I.S.H. and fluorescently-tagged lectins as described above. Sections of parafin-embedded, formalin-fixed tissue
were stained with hematoxylin & eosin, and observed for histological signs of bovine respiratory disease.

5.3.9 Statistical Analysis.
Analysis of polymicrobial biofilms was performed on Z-stack images obtained by confocal scanning laser
microscopy. Z-stacks were analyzed using COMSTAT coded files 358 through Matlab software (MathWorks;
Natick, MA). Statistical analysis was performed using GraphPad Prism software version 6.01 (GraphPad; La
Jolla, CA). Statistical significance was considered if P values were ≤ 0.05.
5.4 RESULTS
5.4.1 Fluorescent in situ hybridization (F.I.S.H.) using DNA-specific probes.
Fluorescently-tagged DNA probes specific for H. somni and P. multocida were used to determine the
spatial arrangement of P. multocida within an established H. somni biofilm. After 48 hours, P. multocida
integrated evenly throughout the H. somni biofilm (Figure 5.1A, B, C). Large microcolonies of P. multocida were
not visible, which has been reported for other polymicrobial bacterial biofilms 359, 360. Additionally, the H. somni
biofilm continued to mature after the addition of P. multocida over the monitored 48-hour period, indicated by
increased green fluorescence between time points. COMSTAT analysis was used to determine the three-
dimensional spatial occupation (biomass) of DNA from each species, as well as the two dimensional thickness
DNA that each species occupies (Table 5.1). Thickness was used as a determination of the depth to which P.
multocida had integrated into the established biofilm matrix.
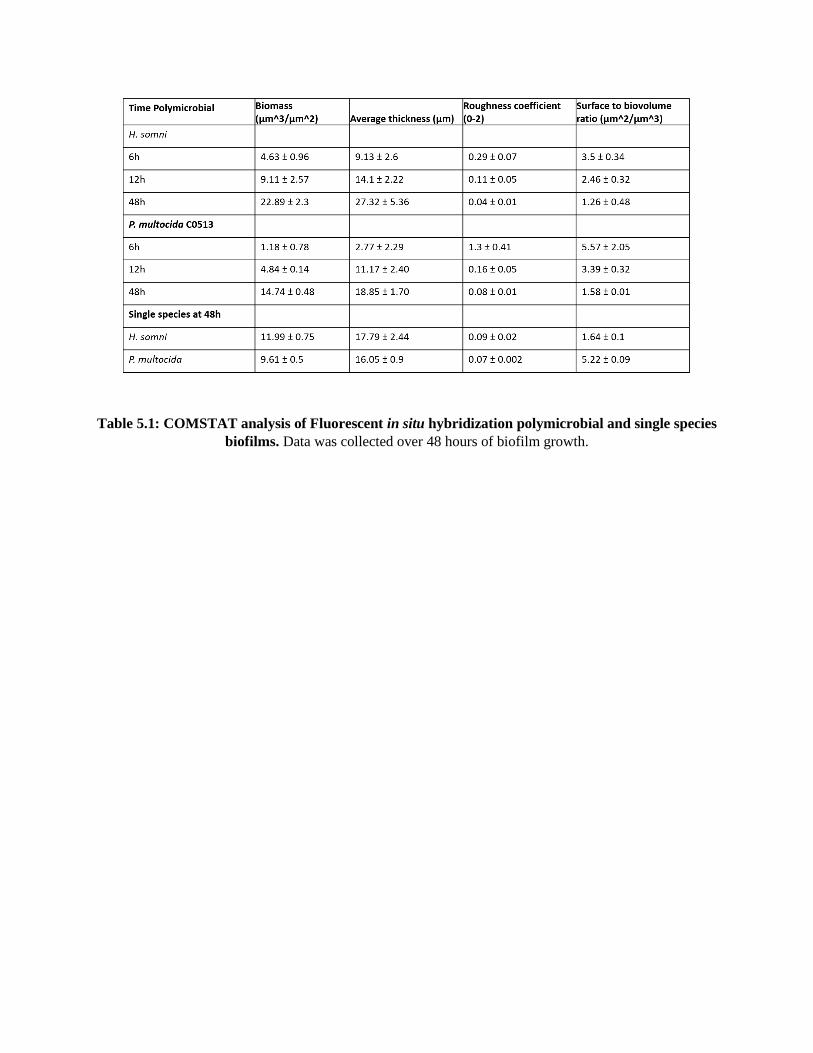
Table 5.1: COMSTAT analysis of Fluorescent in situ hybridization polymicrobial and single species
biofilms. Data was collected over 48 hours of biofilm growth.
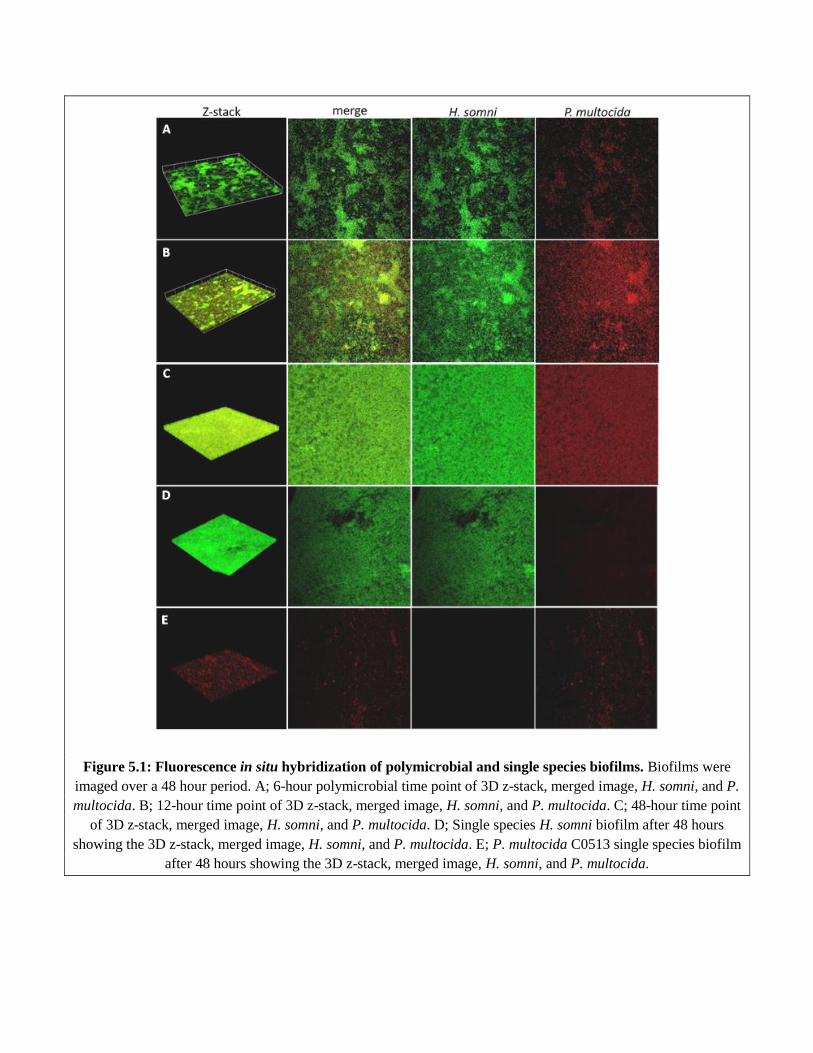
Figure 5.1: Fluorescence in situ hybridization of polymicrobial and single species biofilms. Biofilms were
imaged over a 48 hour period. A; 6-hour polymicrobial time point of 3D z-stack, merged image, H. somni, and P.
multocida. B; 12-hour time point of 3D z-stack, merged image, H. somni, and P. multocida. C; 48-hour time point
of 3D z-stack, merged image, H. somni, and P. multocida. D; Single species H. somni biofilm after 48 hours
showing the 3D z-stack, merged image, H. somni, and P. multocida. E; P. multocida C0513 single species biofilm
after 48 hours showing the 3D z-stack, merged image, H. somni, and P. multocida.

The H. somni biomass increased from 4 ± 1 μm3/μm2 at 6 hours to 22 ± 2 μm3/μm2 at 48 hours (Figure
5.2A). The average thickness similarly increased over the 48 hour period. The roughness coefficient is an
arbitrary value used to determine the topography of the biofilm surface, which remained relatively constant
throughout the experiment, indicating that the addition of P. multocida did not significantly alter biofilm
architecture. The surface to biovolume ratio decreased over the 48 hour period, indicating biofilm growth. The
values provided by the H. somni DNA probe have been used to determine the total thickness of the biofilm, while
the total biomass has been determined by the addition of the H. somni and P. multocida biomass values.
Figure 5.2: Increase in biomass and thickness within the polymicrobial biofilm. A; H. somni biomass and
thickness within the polymicrobial biofilm. B; P. multocida biomass and thickness within the polymicrobial
biofilm.
Six hours after addition to the established H. somni biofilm, P. multocida had attached itself to the H.
somni biofilm likely as a monolayer (Figure 5.1A). This is indicated by a P. multocida thickness measuring 2.77 ±
2.29 μm. The P. multocida biomass was determined to be 1.18 ± 0.78 μm3/μm2 at 6 hours. However, at the 48
hours polymicrobial time point, P. multocida had expanded to a thickness of 18.85 ± 1.7μm and a biomass of
14.76 ± 0.48 μm3/μm2 (Figure 5.2B, Figure 5.1C). This indicated that P. multocida had incorporated into 69% of
the total H. somni biofilm matrix and contributed 39% of the total biomass. Over the 48 hour period, the
roughness coefficient of the P. multocida biofilm contribution decreased to values similar to the H. somni
roughness coefficient, indicating that P. multocida conformed to the H. somni biofilm structure. Despite their

apparent cooperation, the polymicrobial biofilm did not appear to provide a growth advantage- the biomass and
thickness values for the polymicrobial biofilm (indicated by the values provided for H. somni within the
polymicrobial biofilm) were not significantly different than the addition of the values for each single species
biofilm (Figure 5.1D, E).
5.4.2 Fluorescently-tagged lectin staining of biofilm EPS.
Fluorescent lectins were used to determine the presence of EPS from each species to the in vitro
polymicrobial biofilm. Galactomannan EPS and glycogen EPS from H. somni and P. multocida, respectively, was
detected in the polymicrobial biofilm with FITC-conjugated GS-II and TRITC-conjugated MNA-M, respectively.
Glycogen EPS was detected in the polymicrobial biofilm for both P. multocida strains tested: biofilm-deficient
strain C0513 and biofilm-proficient passed variant C0513-P5 (Figure 5.3C, D).

Figure 5.3: Fluorescently-tagged lectin staining of in vitro biofilm EPS. A; P. multocida C0513-P5 single
species biofilm Z-stack, merged image, H. somni EPS, P. multocida EPS. B; H. somni single species biofilm Z-
stack, merged image, H. somni EPS, P. multocida EPS. C; Polymicrobial biofilm with P. multocida C0513-P5 Z-
stack, merged image, H. somni EPS, P. multocida EPS. D; Polymicrobial biofilm with P. multocida C0513 Z-
stack, merged image, H. somni EPS, P. multocida EPS.
Single species C0513 biofilm did not fluoresce, indicating little or no EPS was produced or was
accessible to the lectin (data not shown). COMSTAT analysis of polymicrobial biofilms indicated that 48 hours
after P. multocida C0513 incorporated into an established H. somni biofilm, the average thickness of EPS

produced by C0513 in the polymicrobial biofilm was 20.73 ± 17.03 μm (Table 5.2). The average thickness of EPS
produced by H. somni in the polymicrobial biofilm was 20.58 ± 28.07 μm, indicating variability between the
spatial arrangement of EPS within the biofilm. The average biomass and roughness coefficient displayed similar
variability. The surface to biovolume ratios for EPS produced by C0513 and H. somni in the polymicrobial
biofilm were 5.47 ± 3.16 and 7.21 ± 1.92, respectively.
Polymicrobial biofilms in which passed variant C0513-P5 integrated into an established H. somni biofilm
contained more EPS overall than what was observed for polymicrobial biofilms grown with P. multocida C0513
(Table 5.2). COMSTAT analysis 48 hours after P. multocida C0513-P5 incorporated into the H. somni biofilm
indicated an average thickness of 31.98 ± 19.84 μm for C0513-P5 EPS, and an average thickness of 25.06 ± 18.54
μm for H. somni EPS. The average biomass values were also larger when C0513-P5 was integrated into the
biofilm, and the roughness coefficient was closer to 0 than compared to polymicrobial biofilms incorporating
C0513.
Single species biofilms of C0513 and C0513-P5 contained less EPS than when incorporated into
polymicrobial biofilms. The average EPS thickness and biomass for a single species C0513 biofilm was 2.31 ±
0.12 μm and 1.65 ± 0.09 μm3/μm2 respectively. The average EPS thickness and biomass for a single species
C0513-P5 biofilm were 24.12 ± 0.14 μm and 19.23 ± 0.68 μm3/μm2, respectively (Figure 5.3A). However, the H.
somni single species biofilm contained more EPS than polymicrobial biofilms (Figure 3B). The average EPS
thickness and biomass for the H. somni single species biofilm was 26.2 ± 1.99 μm and 19.69 ± 1.16 μm3/μm2,
respectively.

Table 5.2: COMSTAT analysis of fluorescently-tagged lectin staining of in vitro biofilm EPS. Data was
recorded for biofilms grown 2 days as a single species H. somni biofilm, and then for an additional 3 days as a
polymicrobial biofilm.
5.4.3 Concentrations of Protein and Carbohydrate in polymicrobial biofilms.
Carbohydrate concentrations within the polymicrobial biofilm were determined daily over a period of 6
days (Figure 5.4). The carbohydrate concentration of the established single species H. somni biofilm increased
between days 3 and 4. The average carbohydrate concentration of the P. multocida single species biofilm
remained relatively constant throughout the duration of biofilm growth. The carbohydrate concentration of the
polymicrobial biofilm remained constant until day 3, when P. multocida was added to the established H. somni
biofilm. At this point an increase in biofilm carbohydrate content occurred, and this increase continued until the
end of the experiment on day 6.

Figure 5.4: Changes in carbohydrate concentration during polymicrobial biofilm growth. A; Single species
H. somni biofilm over 5 days of growth. B; Single species P. multocida biofilm over 5 days of growth. C;
Polymicrobial biofilm growth over 6 days, showing H. somni biofilm maturation from days 1-3, with the addition
of P. multocida on day 3.
Figure 5.5: Changes in protein concentration during polymicrobial biofilm growth. A; Single species H.
somni biofilm over 5 days of growth. B; Single species P. multocida biofilm over 5 days of growth. C;
Polymicrobial biofilm growth over 6 days, showing H. somni biofilm maturation from days 1-3, with the addition
of P. multocida on day 3.
Protein concentration was measured daily throughout the biofilm growth period (Figure 5.5). During
single species biofilm growth of H. somni, protein concentration increased over time until the end of the study at
day 5. Protein concentration did not increase over time during single species P. multocida biofilm growth, and
remained at concentrations seen during early stages of biofilm maturation. Similar to the carbohydrate
concentration of the polymicrobial biofilm, the protein concentration of the polymicrobial biofilm increased over

time, with the most dramatic increase occurring between days 3 and 4, which coincided with the addition of P.
multocida to the established H. somni biofilm on day 3.
5.4.4 Auto-aggregation of polymicrobial cultures.
Adhesion is the first stage of biofilm formation. Auto-aggregation is used as a method to determine
cellular adhesion 306, 320, 361. In this study, auto-aggregation of polymicrobial bacterial suspensions was used to
determine whether interactions between the species occurred. H. somni auto-aggregates 80% within 24 hours in a
single species suspension, while P. multocida auto-aggregates significantly less, 20% within a 24 hour period
(Figure 5.6). No additional auto-aggregation was observed for P. multocida or H. somni after a 24 hour period.
Polymicrobial bacterial suspensions autoaggregated 50% when either the encapsulated or unencapsulated P.
multocida variants are used, indicating that interaction may occur between H. somni and P. multocida when in a
polymicrobial environment. Capsular polysaccharide on the surface of P. multocida did not inhibit these putative
interactions. Polymicrobial auto-aggregation experiments carried out in the presence of crudely extracted H.
somni biofilm material did not display aggregation (data not shown), indicating that H. somni cells must be
present in order to induce P. multocida aggregation.

Figure 5.6: Auto-aggregation of polymicrobial suspensions over 24 hours. A; H. somni single species auto-
aggregation. B; Polymicrobial auto-aggregation between H. somni and P. multocida C0513 or C0513-P5. C; P.
multocida C0513-P5 auto-aggregation; D: P. multocida C0513 auto-aggregation.
5.4.5 Polymicrobial biofilm formation in vivo.
Lung tissue from calves experimentally infected with H. somni was analyzed for polymicrobial biofilm
formation by histopathology. Previous experimental infections have shown that P. multocida can establish itself
in calves experimentally infected with H. somni 120. Both H. somni and P. multocida were isolated from calf 65,
but not calf 66. However, PCR revealed P. multocida and H. somni DNA present in the lungs of both calves 65
and 66. H. somni and P. multocida could not be detected by FISH in vivo. However, fluorescent lectin staining
was successfully used to detect the presence of EPS produced by H. somni in calf 65, but not in calf 66 (Figure
5.7).

Figure 5.7: Fluorescently-tagged lectin staining of in vivo biofilm EPS. Lung tissue sections from claf 65
stained with fluorescently-tagged lectins. A,B: merged image, EPS contributed by H. somni, EPS provided by P.
multocida. Scale bar is equal to 20um.
5.4.6 Histological analysis of polymicrobial respiratory disease.
Tissue sections from calf 65 and 66 were observed for histological changes. Lungs appeared slightly
inflamed and partially collapsed. Sections of lung contained pleural fibrosis. Neutrophils and macrophages were
seen in lung airways, and lymphocytes were present at the lung periphery. Acute pulmonary edema was present in
the interlobular septa, and bronchioles displayed minor hemorrhaging. Visible lesions were only detected in one
lobe.
5.4.7 Enzyme-linked immunosorbent assay.

Calves 65 and 66 produced an antibody response to H. somni whole cells at a titer of 1:20. However, both
calves produced an antibody response to H. somni biofilm material at a 1:800 titer. Both calves produced an
antibody response to P. multocida whole cells at a titer of 1:1600, and a response to P. multocida crude biofilm
material at a titer of 1:3200. No response to P. multocida was detected prior to challenge with H. somni.
5.5 DISCUSSION
Despite being intensely investigated for many years, BRD is still the leading cause of morbidity and mortality
in the US beef and dairy industries. Research over the past 40 years has revealed much about the transmission of
the causative agents of BRD, but signs of disease, vaccines, and treatments are still under investigation 184, 185.
BRD is a multifactorial disease, and diagnosis is often complicated by the presence of several causative agents.
Recently, all 4 common bacterial causative agents have been shown to form biofilms in vitro120, 199, 200. Some, such
as the H. somni biofilm, have been demonstrated to be important during BRD and shown to survive in vivo within
the cardiopulmonary tissue 120. In order to further understand the potential role of biofilms in BRD, we have
characterized the polymicrobial relationship between BRD causative agents H. somni and P. multocida.
The adherence of bacteria to a framework or scaffolding, such as a biofilm matrix, that is assembled by
different bacterial species or by the host is known as coadhesion 362, 363. Preliminary studies in our lab indicated
that the best conditions for polymicrobial biofilm formation between H. somni and P. multocida were to allow H.
somni to establish a preliminary biofilm before the addition of the secondary colonizer, P. multocida. This is
consistent with other coadhesive polymicrobial biofilms and was confirmed by experimentation 362-365.
Labelling the polymicrobial and single species biofilms with DNA probes allowed for observation of the
spatial arrangement of bacterial species within the biofilm, and allowed the monitoring of bacterial growth over
time. Antibodies raised against H. somni and P. multocida surface antigens displayed cross reactivity, limiting the
microscopy and fluorescent techniques available for scientific observation 366. The use of DNA probes allowed for
the specific detection of DNA from either species simultaneously 352. However, DNA probes were unable to

distinguish between cellular-associated DNA and extracellular DNA that could potentially be part of the biofilm
matrix 367. Over a 48-hour polymicrobial growth period, the DNA present from both species increased and did not
appear to be separated into large microcolonies or separate biofilms, indicating an integrated polymicrobial
biofilm had formed.
The polymicrobial relationship was further confirmed using fluorescently-tagged lectins specific for EPS
produced by H. somni and P. multocida. Interestingly, the use of lectins provided insight into the glycogen
synthesis of a capsule-producing bovine-associated strain, C0513. This strain did not produce a single species
biofilm. However, once incorporated into the H. somni biofilm matrix, it began producing EPS and contributing
to the overall biofilm matrix. The passed variant of this strain, C0513-P5 did produce a single species biofilm, and
produced visibly more EPS when incorporated into the H. somni biofilm matrix than in a single species biofilm.
The amount of EPS produced by H. somni in the polymicrobial biofilm was lower than the EPS produced during a
single species H. somni biofilm. The reason for this was not determined, but it may be due to a synergistic
relationship with P. multocida, resulting in a decreased need for H. somni EPS 365.
Biofilm matrix carbohydrate and protein content was determined to increase over time, further supporting the
polymicrobial incorporation of P. multocida into the H. somni biofilm matrix. Aggregation experiments were
performed to elucidate any interactions between H. somni and P. multocida cell surfaces. While polymicrobial
suspensions did show an increase in aggregation of P. multocida, no difference was observed in the presence of
H. somni biofilm material. Further experimentation on species interactions was not performed.
Calves challenged with H. somni formed a polymicrobial respiratory infection with both H. somni and P.
multocida, an effect reported in a previous experimental infection 120. Calves presented with few signs of disease
before euthanasia. Upon necropsy, histopathology revealed that both calves had a respiratory infection consistent
with experimental BRD. The calves lungs appeared inflamed and slightly collapsed, and contained pleural
fibrosis. However, lesions were only visible in one lobe of each calf.

Lung sections taken from visible lesions were microscopically observed using the same techniques for in
vitro polymicrobial biofilms. DNA from neither H. somni nor P. multocida were detected in the lungs of calves
using FISH, but DNA from both species was detected in the lungs of calves by PCR. H. somni biofilm EPS was
detected in the lungs of calf 65, but EPS produced by P. multocida was not detected in the lungs of either calf. P.
multocida EPS may have been masked due to the presence of capsular polysaccharide, which is required for full
virulence in vivo. However, both calves produced an antibody response to crude P. multocida biofilm material,
indicating the calves were exposed to biofilm matrix material produced by P. multocida during infection. Because
of this, it is undetermined whether P. multocida contributed EPS to the in vivo biofilm matrix detected.
Additionally, calves produced an antibody responses to P. multocida whole cells, H. somni whole cells, and H.a
somni biofilm matrix.
Biofilms in animal tissues are a significant problem that have been poorly studied 336. However, the relevance
of polymicrobial biofilms to human medicine has become common knowledge 368, 369. This is the first report of a
coadhesive polymicrobial biofilm associated with BRD, leading us to suspect polymicrobial interactions between
other bacterial agents involved in BRD occur. Polymicrobial BRD has been reported, but the presence of biofilm
had not been considered.

CHAPTER 6
Avian biofilm formation and immune response following experimental acute and chronic avian cholera due
to Pasteurella multocida
Briana Petruzzia, Rami A. Dalloulb, Tanya LaRoitha, Nicholas P. Evansc, F. William Piersonc, Thomas J.
Inzanaa,d,*
aVirginia-Maryland College of Veterinary Medicine, Department of Biomedical Sciences & Pathobiology, Virginia Tech,
Blacksburg, VA 24061
bCollege of Agriculture and Life Sciences, Department of Animal and Poultry Sciences, Virginia Tech, Blacksburg, VA 24061
cVirginia-Maryland College of Veterinary Medicine, Department of Population Health Sciences, Virginia Tech, Blacksburg,
VA 24061
dVirginia Tech Carilion School of Medicine, Virginia Tech, Blacksburg, VA 24061

6.1 ABSTRACT
Pasteurella multocida is the causative agent of avian cholera, an important economic and ecological disease that
can present as a peracute, acute, chronic, or asymptomatic infection. Acute avian cholera is associated with
encapsulated P. multocida, while chronic and asymptomatic cases of avian cholera may be associated with
capsule-deficient P. multocida isolates. We hypothesize that biofilm formation is important for chronic and
asymptomatic avian cholera. Experimental infections of chickens with encapsulated, biofilm-deficient P.
multocida strain X73, proficient biofilm forming P. multocida strain X73ΔhyaD, and proficient biofilm forming
clinical strains 775 and 756 showed that virulence was inversely correlated with biofilm formation. Biofilm
forming isolates induced chronic avian cholera in the chicken host. Histopathological analysis was used to show
that biofilm forming isolates induced little inflammation in the lungs, heart, and liver, while biofilm-deficient
isolates induced greater inflammation and induced the recruitment of heterophil granulocytes. Biofilm material
was located in the lungs of chickens diagnosed with chronic avian cholera. Biofilm exopolysaccharide was
located in pulmonary tissue using a fluorescently-tagged lectin, further suggesting the role of biofilm in chronic
avian cholera. P. multocida induced Th1 and Th17 immune responses during acute and chronic avian cholera, as
determined by quantitative real-time PCR of splenic cytokine genes. Chickens that succumbed to acute avian
cholera after experimental challenge with strain X73 had high levels of INF-ƴ, IL-1β, IL-6, IL-12A, IL-22, IL-
17A, and IL-17RA expressed in the spleen compared to all other experimental groups. Birds infected with
capsule-deficient strains had chronic infections lasting 7 days or longer, and had increased levels of IL-17RA,
CCR6, and IL-16 compared to non-infected control chickens. However, specific antibody titers were low,
indicating that antibodies are less important in managing and clearing P. multocida infections.
6.2 INTRODUCTION
Pasteurella multocida is known as the causative agent of avian cholera and other diseases such as bovine
respiratory disease and atrophic rhinitis in swine. Avian cholera is most commonly caused by P. multocida

capsular polysaccharide (CPS) serogroups A and F 148. P. multocida is transmitted from infected or carrier birds to
healthy, non-carrier birds in shared water supplies such as troughs and ponds 156, through rodent infestations 161,
non-fatal animal bites 166, and by a fecal-oral route 168. A common preventative measure used on poultry farms is
to include antibiotics, such as tetracycline, in the drinking water and/or feed. However, this practice may
contribute to enhanced bacterial antibiotic resistance.
Infection with P. multocida can be asymptomatic 7, or can present as avian cholera, an acute, peracute, or
chronic infection. The globalization of avian cholera is the direct result of infected migratory flocks, and has had
profound damaging effects on ecological niches such as the arctic 154, and on poultry farms around the world;
thousands of birds may die during a single outbreak.
During acute infection, P. multocida has been isolated from the air cavities, heart, spleen, bone marrow, and
wattle of turkeys and chickens, as well as from the small intestine, meningeal vessels, air spaces within the skull,
kidneys, eyes, and occasionally synovial cavities 370. Chronic avian cholera begins as an upper respiratory tract
infection. Invasion of the lung, trachea, and air sac tissue occurs less than 1hr post intratracheal inoculation (P.I.).
After 1 hour, invasion of the spleen and liver can occur. By three hours P.I. most other organs have been
colonized by P. multocida. Chronic infections can last 14 days or more and may present with few signs of disease
175.
Interestingly, P. multocida isolates from bovine pneumonia and avian cholera can form a biofilm in vitro,
although such biofilm formation has not yet been observed in vivo 118, 371.118, 371. Biofilm formation has also been
proposed to be involved in chronic swine pneumonia 119, suggesting that the correlation between chronic P.
multocida infections and biofilm formation is not limited to a specific serogroup or host. Biofilm formation has
also been shown to be an important contributing factor to other veterinary diseases 337.
Biofilms are communities of slow growing, often antibiotic-resistant bacteria surrounded by an extracellular
matrix composed of proteins, extracellular nucleic acids, and exopolysaccharide (EPS). Because of these
characteristics, biofilms are often resilient to host defenses and are difficult to eradicate 372. Biofilm formation by

P. multocida may contribute to chronic infection and asymptomatic carriage. While P. multocida has been
demonstrated to make a biofilm, little is known about the role of biofilm during infection, and if biofilm
formation contributes to asymptomatic carriage in some animals. For this study, we sought to identify any
correlation between biofilm formation, strain encapsulation, and chronic avian cholera. Additionally, the cellular
and humoral immune response of the chicken host to chronic and acute avian cholera was examined.
6.3 METHODS
6.3.1 Bacteria and growth conditions
P. multocida strains and clinical isolates used are listed in Table 6.1. Bacteria were cultured on brain heart
infusion (BHI) agar (BD, Franklin Lakes NJ) or Columbia blood agar (BD) supplemented with 5% defibrinated
sheep blood, or Dextrose Starch agar (DSA). Inoculated agar plates were incubated at 37oC with 6% CO2 for 18-
24 hours. Broth cultures were grown at 37°C with shaking at 150 rpm in BHI broth or RPMI-1640 medium
without glutamine or phenol red. For biofilm formation, 50 μl of mid-log phase P. multocida were inoculated into
5 ml of RPMI-1640 in a 50-ml polyethylene tube or onto a glass slide in a Petri dish. Biofilms were incubated
without shaking at 37oC in 6% CO2 for at least 48 hours. Biofilm formation was confirmed by staining with 200
μl of crystal violet, followed by incubation at room temperature for 10 minutes. Biofilms were then gently washed
with sterile PBS, and the crystal violet dissolved in 500 μl 95% ethanol. The OD560 of the biofilm-crystal violet
solution was recorded using a Molecular Devices Vmax Kinetic Plate Reader (Molecular Devices; Sunnyvale,
CA). CPS content was quantified as described 371.

_____________________________________________________________________________
Strain Serogroup Source or reference Disease origin
______________________________________________________________________________
X73 A Dr. R. Briggsb Avian cholera
X73ΔhyaDa nontypable 371 NAc
775 A Virginia Dept. Ag. Turkey lung
756 F Virginia Dept. Ag. Chicken heart, spleen
______________________________________________________________________________
Table 6.1. P. multocida strains used. aCapsule-deficient mutant of strain X73. bNational Animal Disease Center,
Agricultural Research Service, U.S. Department of Agriculture, Ames, Iowa, USA. cNA-not applicable
6.3.2 P. multocida-chicken challenge experiments.
A pilot study was performed to determine the appropriate challenge dose needed to induce chronic and acute
avian cholera with biofilm-proficient and biofilm-deficient isolates. Chicks were hatched from SPF eggs (Charles
River, Wilmington, MA) and raised until 8 weeks old. Three infection groups (inoculated with strains 775, 756, or
X73) contained 3 subgroups each (with the exception of birds inoculated with strain 756, which contained only 2
subgroups) of 5 birds each. Each subgroup was inoculated with either 104, 106, or 108 colony forming units (CFU)
of each P. multocida strain administered in 500 μl of sterile saline. One hundred microliters was administered
intranasally: 50 μl given in each nostril, and the remaining 400 μl intratracheally. A control group was
administered saline only using the same intranasal and intratracheal routes. Birds were euthanized if they became
moribund. All birds were euthanized by 3 weeks post-inoculation. Lung, trachea, and heart samples were
collected at the time of euthanasia and stored in 10% buffered formalin for microscopic observation.

A second challenge experiment was performed with chickens of the same age (8 weeks). Three
animal infection groups (with strains X73, X73ΔhyaE, and 775) contained 21 birds each, and a control
group contained 7 birds. Birds inoculated by the intranasal and intratracheal routes with virulent strain
X73 were given a dose of 104 CFU. Birds inoculated with capsule-deficient strains 775 and X73ΔhyaD
were given a dose of 106 CFU. Control birds were inoculated with sterile saline. P. multocida was
administered in 500 μL of sterile saline as described above. Birds were euthanized if they became
moribund. Seven birds from each group were euthanized 1 week post-inoculation and another 7 birds
were euthanized 2 weeks post-inoculation. On day 21, surviving birds were re-inoculated with 108 CFU
of the same strain of P. multocida by the same route. All remaining birds were euthanized 3 days after
the second inoculation. Blood for antibody determination was collected from the heart at the time of
euthanasia due to the difficulty in obtaining blood from the wings of live birds. Lung, spleen, and liver
samples were collected at the time of euthanasia and stored in RNAlater RNA storage reagent (Qiagen;
Venlo, Limburg). Lung and heart samples were collected at the time of euthanasia, and stored in both
liquid nitrogen and 10% buffered formalin for further observation.
Symptoms of chronic avian cholera included reduced activity (secluded to the back of the cage), reduced feed
intake, reduced water intake, and ruffled feathers. Symptoms of acute avian cholera were sudden reduction in
activity, reduced feed intake, ruffled feathers, rapid breathing, mucous discharge from mouth and/or nostrils, and
diarrhea. Sudden death was sometimes the first sign of disease.
All animal studies were performed using protocols approved in accordance with the Institutional Animal Care
and Use Committee (IACUC) at the Virginia Polytechnic Institute and State University
(https://www.researchcompliance.vt.edu/iacuc/).
6.3.3 In vivo histopathology (H&E stain).

Lung, heart, and trachea tissues were obtained from experimentally challenged chickens and non-infected
controls at the time of euthanasia. Tissue samples were embedded in paraffin wax, cut onto slides, and stained
with hematoxylin and eosin (H&E) stains, or a tissue Gram stain. Lung, heart, and trachea samples were scored
using parameters described elsewhere 373. Scoring was based on thrombosis, necrosis, neutrophils, heterophils,
and perivascular edema. The presence of intracellular or extracellular P. multocida or biofilm matrix were noted.
6.3.4 Scanning electron microscopy of tissues for biofilms.
Tissue sections were cut into approximately 1 cm x 1 cm x 1 cm cubes, washed, and fixed in a solution of 5%
glutaraldehyde, 4.4% formaldehyde, and 2.75% picric acid in 0.05% sodium cacodylate buffer for at least 1 hour.
Specimens were gradually dehydrated using 25%, 50%, 70%, 80%, and 95% ethanol. SEM was performed on a
Carl Zeiss EVO40 scanning electron microscope.
6.3.5 Fluorescent-lectin staining of the exopolysaccharide biofilm.
Fluorescein isothiocyanate (FITC)-conjugated Griffonia simplicifolia lectin (GS-II) (EY laboratories; San
Mateo, CA) was used to detect the glycogen EPS that makes up the P. multocida biofilm. FITC-GS-II was
suspended in a 10 mM phosphate buffer, pH 7.5, at a concentration of 10 μg/ml. Fifty microliters of lectin
solution were applied to specimens and incubated 30 to 60 minutes in the dark at room temperature, then washed
four times with phosphate buffer. Coverslips were fixed onto slides with 20 μl of polyvinyl alcohol (Sigma-
Aldrich; St. Louis, MO) and stored in the dark for at least 6 hours before imaging. Confocal scanning laser
microscopy was performed on a Zeiss 880 Confocal microscope (Zeiss, Germany) at 40x magnification using a
fluorescein (green, 500-540 nm) filter.
6.3.6 Enzyme-linked immunosorbent assay (ELISA).

ELISAs were performed by modification of a previous protocol 374. Bacterial cells (109 CFU/ml) were washed
and suspended in carbonate coating buffer pH 9.6 (15 mM Na2CO3, 35 mM NaHCO3, 3 mM NaN3), or were lysed
using a Fisher Scientific Sonic Dismembraner Model 500, at 60% amplitude, in carbonate coating buffer. One
hundred μl of whole cell suspension or lysate were added to wells of a flat bottom 96-well microtiter plate
(Immulon 4HBX, ThermoFisher Scientific; Waltham, MA). Nonspecific binding was blocked by addition of 2%
non-fat milk (Kroger; Cincinnati, OH) supplemented with 0.5% tween 20 (Sigma Aldrich; Raleigh, NC). Chicken
serum was added at a starting dilution of 1:10 in the same blocking buffer. Goat anti-chicken IgY antibody
conjugated to horseradish peroxidase (Pierce Biotechnology; Rockford, IL) was used at a concentration of 1:5000
in PBS supplemented with 0.05% Tween-20 (Sigma Aldrich; Raleigh, NC). The bound conjugate was detected by
addition of TMB reagent (Pierce Biotechnology; Rockford, IL), and the reaction was stopped with 1M sulfuric
acid. Optical densities were determined at 560 nm using a Molecular Devices Vmax Kinetic Plate Reader
(Molecular Devices). Endpoint antibody titers were considered the dilution of serum with A560 values that were
two times the control values of wells lacking only specific antigen.
6.3.7 Bacterial numbers in pulmonary tissue.
Lung tissues that were frozen at -80°C at the time of necropsy were weighed and homogenized in 1 ml of
sterile saline. Homogenates were diluted, cultured onto Columbia Blood agar, and incubated overnight at 37oC
with 6% CO2. Bacterial colonies were counted, and based on the initial tissue weight, the CFUs/gram tissue were
determined.
6.3.8 RNA extraction and qRT-PCR.
RNA was extracted directly from spleen samples stored in RNAlater (Qiagen; Hilden, Germany). Twenty 50-
mg sections of spleen were homogenized in 900 μl TriZol (ThermoFisher; Waltham, MA), and stored at room

temperature for several minutes. Chloroform (180 μl) was then added to the homogenate, and vortexed
vigorously. After 3 minutes at room temperature, the homogenate solutions were centrifuged at 12,000 x g for 15
minutes at 4oC. The aqueous phase was transferred to a clean tube containing an equal volume of 70% ethanol.
The Qiagen RNeasy kit (Qiagen; Hilden, Germany) was used to extract RNA from the resulting solution. The
purity of the RNA was determined by the A260/280 and A260/230 ratios with a nanodrop (NanoDrop; Wilmington,
DE). Fifty ng of pure RNA was transcribed into cDNA using the Quanta Biosciences qScriptTM XLT cDNA
Supermix kit (Quanta Biosciences; Gaithersburg, MD) according to the manufacturer’s instructions. Quantitative
real-time polymerase chain reaction (qRT-PCR) was performed using the Quanta Biosciences PerfeCta® SYBR®
Green FastMix, ROXtm (Quanta Biosciences; Gaithersburg, MD) according to the manufacturer’s instructions on
an Applied Biosciences 7300 Real-Time PCR system (Applied Biosciences; Foster City, CA). Quantitative RT-
PCR was performed in 20 μl reactions using 50 ng cDNA template and 300 mM of each primer. The genes
targeted for RT-PCR are listed in Table 6.4.
6.3.9 Statistical analyses
Determination of the median, standard deviation, and p values of comparative values were determined by
unpaired student t test. Virulence was evaluated by comparing survival of birds in each inoculation group during
the pilot study using a 2 x 2 contingency table with Fisher’s exact test. All statistical calculations were performed
using GraphPad Prism version 6.01 software (GraphPad Software, Inc., La Jolla, CA). Statistical significance was
determined at p ≤ 0.05.
6.4 RESULTS
6.4.1 Respiratory infection: Pilot study.

All birds given intranasal and intratracheal challenge with 108 CFU of highly virulent, biofilm-deficient strain
X73 succumbed to acute avian cholera within the first 24 hours of the study and were euthanized. Four birds
challenged with 106 CFU, and 2 birds challenged with 104 CFU of strain X73 also needed to be euthanized within
1 day of challenge. The surviving four birds inoculated with strain X73 continued to show signs of avian cholera
until the end of the study on day 15 P.I. (Fig. 6.1A).
Chronic infections were induced in chickens inoculated with biofilm-proficient, CPS-deficient strains 756 and
775 at 104, 106, or 108 CFU. Strains 775 and 756 made less CPS (67.5 ± 16.86 μg/ml and 66.67 ± 10.48 μg/ml
uronic acid, respectively) than strain X73 (92.97 ± 36.82 μg/ml uronic acid). However, this difference was not
significant. Fourteen birds challenged with 108, 106, or 104 CFU of strain 775 developed symptoms of chronic
avian cholera, and such symptoms remained until the end of the study (Fig. 6.1B). One bird in the 108 CFU
challenge group died due to avian cholera, but strain X73 was isolated from this bird due to an apparent mis-
challenge. All birds challenged with strain 756 also displayed signs of chronic avian cholera until the end of the
study (Fig. 6.1C). None of the birds challenged with strains 775 or 756 succumbed to acute avian cholera.
Strain X73 inoculated intranasally and intratracheally into birds (108 CFU) was significantly more virulent
than the same dose of strain 755 in chickens (p = 0.0079). Birds challenged with strain X73 at 108 CFU or 106
CFU were also significantly more virulent than strain 756 inoculated into birds at 106 CFU (p = 0.0079). The
difference in virulence of strain 775 inoculated at 108 CFU and 106 CFU (p = 1.0000) compared to strain 756
challenged at 106 CFU was not significantly different. Virulence was determined by mortality using 2x2
contingency tables with Fisher’s exact test comparing chickens that succumbed to acute avian cholera and/or were
euthanized during the experiment to those with a chronic avian cholera. Upon necropsy, pure cultures of P.
multocida were recovered from the nostrils, trachea, lungs, and air sacs of all chickens challenged with P.
multocida at all CFU concentrations, and appeared phenotypically similar to the parent.

Figure 6.1: Pilot Study Survival Curve. P. multocida X73 (A), Clinical isolate 775 (B), clinical isolate 756 (C).
6.4.2 Histopathology of chicken lungs, heart, and trachea.
Inflammation of the lungs, heart, and trachea was analyzed in chickens experimentally challenged during the
pilot study. Organs were scored from 0-4 based on thrombosis, necrosis, heterophils, and perivascular edema.
Inflammation present in the lungs, heart, and trachea was heterophilic, which is a common pathological sign of
avian cholera 375. A score of 0 represented normal tissue pathology, while a score of 1 represented minimal
inflammation, a score of 2 represented mild or focal inflammation, a score of 3 represented moderate or
multifocal inflammation, and a score of 4 represented severe or widespread inflammation (Fig. 2 A-E).
Chickens inoculated with strains 756 and 775 had similar pathologies among challenge groups (104, 106, or
108 CFU). Inflammation scores ranged between 1 and 2 (minimal to mild or focal inflammation) for lungs, 0-1 for
heart (normal to minimal inflammation), and 0-2 for the trachea (normal to mild/focal inflammation). However,
chickens challenged with 106 CFU of strains 775 and 756 had minimal/mild signs of inflammation in the lungs
and trachea compared to higher (108 CFU) and lower (104 CFU) challenge doses, which were scored normal or
with minimal inflammation.
Chickens that survived challenge with strain X73 appeared to recover from infection, and showed few signs
of inflammation (minimal in the lung, mild or focal inflammation in the heart, and no inflammation in the

trachea). Chickens that succumbed to an acute infection after inoculation with strain X73 at 104, 106, and 108 CFU
had inflammation of the lung ranging from moderate/multifocal to severe, but had minimal to mild/focal
inflammation in the trachea and heart (Table 6.2). Clusters of bacteria were observed in the lumen of the trachea
of birds that succumbed to acute avian cholera after challenge with strain X73. Bacteria, either intracellular or
extracellular, were observed in chickens experimentally challenged with strains X73, 756, and 775. Putative
biofilm material could be seen in the lungs of chickens with acute and chronic avian cholera (Fig. 6.2 F-H).
Figure 6.2: Gram stained tissue sections from the pilot study. All images are at 40x magnification.
Control lung (A), lung section with a minimal inflammation score of 1 (B), lung representative a mild or focal
inflammation score of 2 (C), lung representative of a moderate or multifocal inflammation score of 3 (D), lung
representative of a severe or widespread inflammation score of 4 (E). Lung with an inflammation score of 4 with
visible biofilm material (F, arrow). Other lung sections with putative biofilm material (G and H, arrows).

____________________________________________________________________________
Strain Challenge Dose Surviving Histopathology Scorea
Birdsb Lung Heart Trachea ____
756 104 5/5 1 0 0
756 106 5/5 2 1 2
775 104 5/5 1 0 0
775 106 5/5 1 0 1
775 108 4/5c 1 0 0
X73 104 3/5 3 1 2
X73 106 1/5 4 1 1
X73 108 0/8 3 1 2
_____________________________________________________________________________
Table 6.2. Histopathology scoring of lung, heart, or trachea of experimentally infected chickens in pilot
study. aScoring: 0 = normal, 1 = minimal, 2 = mild or focal, 3 = moderate or multifocal, 4 = severe or widespread.
These values are averages of the birds in each group. bNumber of birds that survived challenge per size of
challenge group. cStrain X73 was isolated from the one bird that died from this strain 775 challenge group.
6.4.3 Scanning Electron Microscopy (SEM).
Microscopy of lung sections revealed probable biofilm material within the lungs of chickens in the pilot
study. The most prominent and extensive putative biofilm material was evident in birds infected with strain
X73ΔhyaD (Fig. 6.3B,C), while less putative biofilm material was present in birds infected with strain 775 (Fig.
6.3A), and even less in birds infected with strain 756 (Fig. 6.3D). A small amount of putative biofilm material

was also evident in birds infected with virulent strain X73 (Fig. 6.3E). No potential biofilm material was
identified in the lungs of non-infected control birds (Fig. 6.3F). The putative biofilm material observed within the
lungs was consistent with the biofilm morphology of P. multocida 371.
Figure 6.3: SEM of chicken lungs after experimental infection. Arrows indicate the presence of putative
biofilm matrix in the airways of chickens experimentally infected with P. multocida clinical isolate 775 (A), strain
X73ΔhyaD (B, C), clinical isolate 756 (D), WT X73 (E), non-infected control (F).
6.4.4 Fluorescent-lectin staining of biofilm EPS.
P. multocida produces a glycogen EPS as part of the biofilm matrix that is reactive with the lectin GS-II 371.
Therefore, FITC-labelled GS-II was used to determine whether P. multocida EPS, and hence biofilm matrix, were
present in the pulmonary tissue of chickens experimentally infected as part of the pilot study. Glycogen was
observed in patches at high concentrations in pulmonary tissue of chickens infected with strain 775, supporting
the presence of P. multocida biofilm matrix in these specimens (Fig. 6.4B). These patches of glycogen were not
detected in non-infected controls (Fig. 6.4A).

Figure 6.4: Fluorescent lectin staining of in vivo biofilm EPS. The lectin GS-II specifically binds to glycogen,
and when tagged with FITC can be used to determine the presence of glycogen biofilm EPS in the pulmonary
tissue of chickens experimentally infected during the pilot study described in this report. Glycogen EPS was
found in the pulmonary tissue of chickens experimentally infected with clinical isolate 775 (B). Glycogen EPS
was not found in non-infected control chickens (A).
6.4.5 Follow-up experimental challenge.
During the second experimental infection, acute avian cholera was induced in 8 out of 20 (40%) chickens that
were inoculated with 104 CFU of strain X73 (1 chick died prior to challenge). Chickens that succumbed to acute
avian cholera were euthanized within 24 hours P.I. P. multocida was recovered from the lungs of all birds
showing signs of acute disease. The surviving 12 chickens inoculated with strain X73 showed signs of a subacute
infection, and appeared to recover after 3-4 days. On day 7 of the experiment, P. multocida was recovered from
the lungs of 3 of 6 euthanized chickens (50% recovery). On day 14 of the experiment, the remaining 6 chickens
that had been inoculated with strain X73 were euthanized, and P. multocida was recovered from the lungs of 1 of
the 6 chickens (16% recovery) (Table 6.3).

Acute avian cholera was induced in 1 chicken inoculated with 106 CFU of strain X73ΔhyaD. This acutely
infected chicken accidently received two doses of inoculum, and was euthanized within 24 hours P.I. On day 7,
six of the chickens were euthanized and P. multocida was recovered from 1 chicken (16% recovery). On day 14,
six additional chickens were euthanized and P. multocida was not recovered from any of the birds. The remaining
7 chickens were given a second challenge inoculum of 108 CFU of strain X73ΔhyaD on day 21 and were
euthanized 3 days later. P. multocida was recovered from the lungs of 1 chicken (14% recovery).
Similarly, P. multocida was not recovered from any chickens challenged with 106 CFU of strain 775 prior to
the second challenge. After a second challenge with 108 CFU of strain 775, P. multocida was recovered from 4 of
7 euthanized chickens (57% recovery) (Table 6.3).
___________________________________________________________________________
Challenge Day 1a Day 7 Day 14 Day 24b
Strain Percent of birds from which P. multocida was recovered from lungs
______________________________________________________________________________
X73 100% (8 of 8) 50% (3 of 6) 16% (1 of 6) --c
X73ΔhyaD --d 16% (1 of 6) 0% (0 of 6) 14% (1 of 7)
775 --d 0% (0 of 7) 0% (0 of 7) 57% (4 of 7)
______________________________________________________________________________
Table 6.3: Bacterial recovery of P. multocida X73 from challenged chickens. aDays post-inoculation (P.I.)
chickens were euthanized after the second experimental challenge. bA second challenge was given P.I. day 21
and the birds euthanized 3 days later. cNo birds survived to day 24 P.I. in this group. dSamples were not obtained
because these birds were not acutely ill at P.I. day 1 and were not euthanized.
6.4.6 Antibody response.

Sera were collected from each bird in each group from the follow-up challenge study at the time of
euthanasia. Of the eight birds challenged with strain X73 that developed acute avian cholera blood could only be
obtained from one, but an antibody response to P. multocida was not detected. At day 7 P.I. seven of the surviving
birds were euthanized, but only two made a detectable antibody response. At day 14 P.I., the seven remaining
birds were euthanized, and an antibody response that averaged about 1:400 was detected in four of those birds
(Fig. 6.5A).
Only two of seven birds in the strain X73ΔhyaD challenge group had detectable antibody titers (average
1:500) to P. multocida at 7 days P.I. At 14 days P.I., three of seven birds made a detectable response, which
averaged 1:1300, and 3 days after the second challenge 4 of seven birds made an antibody response that averaged
about 1:900 (Fig. 6.5B).
Of 21 birds challenged with strain 775, an antibody response could be detected in only eleven birds. No
bacteria were isolated from chickens inoculated with strain 775 prior to the second inoculation. At day 7 P.I., four
of seven birds inoculated with strain 775 had an average antibody titer of 1:300. At 14 days P.I., three of seven
challenged birds had detectable antibodies to P. multocida with an average titer 1:1600. The remaining 7 birds
were re-inoculated at day 21 P.I., euthanized 3 days later, and four of these birds had a detectable response of
1:600 (Fig. 6.5C). Differences in antibody titers between groups were not statistically significant (Fig. 5D),
probably due to the large standard deviation within groups. Antibody titers to cell lysates were comparable to
titers to whole cells (data not shown).

Figure 6.5: Antibody Response to experimental infection. Strains X73 (A), X73ΔhyaD (B), 775 (C), all strains
(D).
6.4.7 Detection of cytokines by qRT-PCR of spleen cells following challenge.
An initial screen of 24 chicken cytokine markers was performed using RNA isolated from infected spleen
tissues of chickens that succumbed to acute avian cholera after inoculation with strain X73 (Table 6.4). The
relative fold increase (RFI) in cytokine expression was determined by comparing cytokine expression levels of
acutely infected chickens with those of non-infected control chickens. Chemokine CCR6, cytokines IL-17A, IL-6,
IL-22, IL-12A, IL-16, IL-1β, IFN-γ, and receptor IL-17RA were chosen for further analysis (Table 6.5). Cytokine
IL-15 (RFI of 9.55) was upregulated in the spleen during acute infection, but not chosen for further analysis.
Chickens that were acutely infected with strain X73 succumbed to infection within the first 24 hours of the
experiment, and had increased expression levels of all cytokines tested, but not chemokine CCR6. The greatest

expression levels were to IFN-γ and IL-1β. Significant responses to INF-ƴ and IL-12A indicated a Th1 immune
response was present. A Th17 immune response was also apparent, as shown by the increased expression levels of
IL-17A, IL-17RA, IL-22, and IL-1β. Seven days P.I., chickens that survived inoculation with strain X73 had
increased expression levels of IL-17RA, IL-22, and CCR6, but now no response to IL-1β. Expression of Th1
cytokine IL-16 was also increased, but not IFN-γ. At 14 days P.I., the results were similar to 7 days P.I.:
Expression of IL-17RA, CCR6, and IL-16 were increased compared to the spleens of non-infected control
chickens. No chickens remained at day 21 to receive a second inoculation.
Cytokine and chemokine expression levels within the first 24 hours were not available for chickens inoculated
with strains 775 or X73ΔhyaD because the birds inoculated with these isolates were clinically asymptomatic at 1
day P.I. Seven days P.I., chickens inoculated with strain X73ΔhyaD had increased expression levels of IL-16, IL-
17A, IL-22, and IL-1β when compared to non-infected control spleens. Expression of IL-17RA (p = 0.0013) and
IL-16 (p = 0.0255), were significantly higher 7 days P.I. in chickens inoculated with strain X73 than with strain
X73ΔhyaD, indicating that the overall immune response was lower in birds inoculated with capsule-deficient
mutant X73ΔhyaD. However, the response to IL-17A at this time point was much greater than to X73. Fourteen
days P.I., expression of IL-16 (p = 0.0080) and CCR6 (p = 0.0038) were significantly higher in chickens
inoculated with strain X73 than with strain X73ΔhyaD. After the second challenge with strain X73ΔhyaD on P.I.
day 24, expression levels of IL-17RA, IL-16, IL-12A, and IFN-ƴ were increased compared to non-infected
controls.
Chickens inoculated with strain 775 had increased expression levels in the spleen at 7 days P.I. to IL-17RA,
IL-16, CCR6, IL-1β, and IFN-ƴ. However, expression levels of IL-17RA (p = 0.0025) and CCR6 (p = 0.0032)
were significantly higher at 7 days P.I. in chickens inoculated with strain X73 compared to strain 775. As for
chickens inoculated with strain X73hyaD, chickens inoculated with strain 775 produced a milder cytokine
response compared to virulent strain X73. Fourteen days P.I., expression levels of Th17 markers IL-17RA and IL-
16 were increased in birds inoculated with strain 775. After a second challenge with strain 775, expression levels
of IL-16, IL-17A, IL-22, IL-12A, and IL-1β were increased 3 days after the second challenge (Table 6.5).

______________________________________________________________________________
Cytokine/ Relative Fold Cytokine/ Relative Fold
Chemokineb Increase Chemokineb Increase
______________________________________________________________________________
CCL20 0.64 IL-12B 1.11
CCR6 1.3 IL-13 0.72
CXCR2 6.4 IL-15 9.55
CXCR4 0.73 IL-16 19.9
IL-1β 109.9 IL-17A 0.17
IL-2 0.13 IL-17RA 1.27
IL-4 0.60 IL-17RC 0.38
IL-5 0.98 IL-18 0.20
IL-6 0 IL-22 0.16
IL-8 0.02 IFN-ƴ 0.12
IL-10 0.47 GM-CSF 0
IL-12A 2.69
______________________________________________________________________________
Table 4. Cytokine or chemokine genes tested by qRT-PCR on infected spleen samples.a aThe cytokine
responses of spleen tissue of chickens with acute avian cholera were compared with the lungs of non-infected
control birds. The cytokines and relative fold increase are displayed. bCytokines shown in bold font were chosen
for further analysis.

______________________________________________________________________________
Strain Cytokine marker 1 day P.I.b 7 days P.I. 14 days P.I. 3 days post 2nd
challenge
______________________________________________________________________________
X73 CCR6 0 1.38 ± 1.07 3.15 ± 1.19 N/A
IL-1β 402.15 ± 117.63 0 0 N/A
IL-6 167 ± 65 0 0 N/A
IL-12A 3.84 ± 2.76 0 0 N/A
IL-16 7.65 ± 2.38 16.68 ± 9.05 11.86 ± 4.19 N/A
IL-17A 3.03 ± 1.85 0 0 N/A
IL-17RA 12.77 ± 3.95 16.68 ± 6.64 11.41 ± 8.83 N/A
IL-22 25.08 ± 9.74 3.58 ± 0.38 0 N/A
IFN- ƴ 370.53 ± 142.43 0 1.79 ± 0.53 N/A
X73ΔhyaD CCR6 N/A 0 0 0
IL-1β N/A 73.49 ± 15.84 0 0
IL-6 N/A 1.43 ± 0.87 0 0
IL-12A N/A 0 0 4.03 ± 0.68
IL-16 N/A 2.75 ± 1.2 3.01 ± 0.80 7.64 ± 3.19
IL-17A N/A 522 ± 105 2.75 ± 3.37 0
IL-17RA N/A 0 4.63 ± 1.16 2.3 ± 2.45

IL-22 N/A 12 ± 7.74 0 0
IFN- ƴ N/A 0 0 2.79 ± 3.40
775 CCR6 N/A 4.76 ± 0.94 0 0
IL-1β N/A 27.07 ± 5 0 7.68 ± 2.09
IL-6 N/A 0 0 0
IL-12A N/A 0 6.83 ± 1.55 1.83 ± 0.30
IL-16 N/A 6.39 ± 1.18 81.17 ± 50.13 3.07 ± 3.81
IL-17A N/A 0 0 2.97 ± 0.73
IL-17RA N/A 1.96 ± 0.54 6.00 ± 1.62 0
IL-22 N/A 0 0 4.54 ± 1.11
IFN- ƴ N/A 2.95 ± 0.32 0 0
______________________________________________________________________________
Table 6.5. Relative fold increase of cytokine gene expressiona in spleens of chickens experimentally infected
with different strains of P. multocida . aThe relative fold increase of IL-17RA, IL-16, CCR6, IL-1β, and INF-ƴ of
chickens challenged with strains X73, X73ΔhyaD, or 775 are listed for all time points tested. bBirds challenged
with strain X73 succumbed to acute avian cholera, so data points 24 hr P.I. are available for chickens challenged
with strain X73, but not any other group.
6.5 DISCUSSION
Avian cholera is a worldwide concern, causing significant economic and ecological losses each year.
Outbreaks of P. multocida reoccur frequently due to the ability of P. multocida to survive in carrier birds, insects,
water supplies, and soil for up to several months after signs of disease have disappeared 156, 162, 170. Most research

to date has focused on highly virulent strains of P. multocida and acute avian cholera outbreaks, largely
underestimating the importance of chronic infections and asymptomatic carriage. The work presented here aims
to characterize chronic avian cholera, and determine the importance of biofilm formation in disease persistence.
Additionally, we compared the immune response of the chicken host presenting with acute or chronic avian
cholera.
P. multocida strain X73 is a poultry isolate recovered from birds with acute avian cholera. Although also
isolated from birds, strains 775 and 756 were isolated from tissues of a turkey and chicken, respectively, with a
more chronic, less serious, infection. Our challenge experiments confirmed that strain X73 was more virulent and
caused more acute disease than strains 775 or 756, as well and a capsule-deficient mutant of strain X73. Strain
X73 also produced more capsule and less biofilm than the other strains. In their natural environment, bacteria
likely persist as part of a biofilm community. Biofilm formation is considered a universal mode for survival of
both pathogenic and environmental bacteria. Genetic modifications 376 that occur during transition from
planktonic to biofilm growth may explain the transition from a virulent phenotype to a less virulent phenotype,
which may be associated with transmission of P. multocida in environmental and asymptomatic populations.
Strains 775 and 756 were proficient biofilm formers in vitro, and produced less CPS than biofilm-deficient strain
X73 (noteworthy by colony morphology on blood agar). The difference in pathogenicity between strains 775 and
X73 was confirmed in both the pilot study and by a second experimental infection. Virulence increased with CPS
production and decreased with biofilm formation (determined in vitro), as previously described 371.
Putative biofilm material in respiratory specimens from our challenged birds were identified using SEM,
histological analysis, and fluorescence microscopy using a FITC-labelled lectin. SEM also revealed likely biofilm
material in the airways of birds infected with clinical strains 775 and 756. This material was not present in the
airways of control birds, and minimally present in the airways of birds that succumbed to an acute infection after
challenge with strain X73. Clusters of gram-negative bacterial cells, as well as putative biofilm matrix material,
were present in the pulmonary tissues of experimentally infected chickens. Patches of concentrated glycogen were
detected using the FITC-tagged lectin to the glycogen EPS (evidence of biofilm). Analyzing these results, we

believe that biofilm formation was important for the survival of less virulent P. multocida inside (chronic
infections, asymptomatic carriage) and outside (environmental persistence in soil and water supplies) 156, 165 of the
host.
Heterophils were noted at sites of inflammation in the lungs, trachea, and hearts of challenged birds, which is
common during avian cholera. Heterophils are important for the clearance of P. multocida, but may initially aid in
P. multocida dissemination throughout the host 377. Inflammation was most severe in the lungs of chickens that
succumbed to acute avian cholera, and was minimal in birds that developed chronic infections with less virulent
strains 775 and 756. P. multocida was observed in the tracheal lumen of chickens experimentally challenged with
strains 756, 775, or X73. Clusters of bacteria could be visualized by H&E and Gram stains of tissue sections in
the trachea of chickens inoculated with strain X73. However, bacteria were present in the trachea of birds that
survived challenge with strain X73 three weeks P.I., indicating that chronic infections had become established.
Due to the peristaltic nature of the trachea, P. multocida were likely adhered and encased in a biofilm material,
which would offer protection from the host immune response and allow P. multocida to persist in the airways.
In the second experimental challenge, the immune response of chickens to acute and chronic avian cholera
was examined. At 7 days P.I., P. multocida was recovered from approximately half of the infected chickens. At
14 days P.I., P. multocida was only recovered from chickens inoculated with strain X73 that did not succumb to
acute avian cholera. However, most of the birds in each group did not made a detectable immune response to P.
multocida at 7 and 14 days P.I., regardless of P. multocida recovery, and therefore titers shown are representative
of only those birds that made a response. The short time period of the study likely contributed to the lack of
antibody response. Nonetheless, antibody titers of challenged chickens remained low for all strains and isolates
tested. However, titers varied greatly between birds within the same group, which is in agreement with reports
that antibody responses do little to slow the progression of avian cholera 378. Phagocytic cells may be more
important for a successful response to overcome infection and prevent acute avian cholera 151. The lowest
antibody titers occurred in birds experimentally infected with highly virulent strain X73. This strain is highly
encapsulated and the hyaluronic acid CPS, which is not immunogenic, may also mask other surface antigens and

inhibit the host immune response 282. Antibody titers were generally higher at 7 and 14 days P.I. for the less
virulent and more prominent biofilm-forming strains 775 and X73ΔhyaD than for highly virulent strain X73.
A cellular immune response may be more important in protection against P. multocida serogroup A infection
in most animals. Mice are protected from disease following intranasal challenge with P. multocida by an IL-17-
dependent, antibody-independent response 379. The cytokine and chemokine response of chickens in all
experimental infection groups suggested that a Th1 and Th17 response, rather than a Th2 response, predominated.
Chickens that succumbed to acute avian cholera had increased expression levels of INF-ƴ, IL-16, IL-15, IL-1β,
IL-22, IL-17A, IL-12A, and receptor IL-17RA when compared to non-infected control chickens. Cytokines IL-15
and IL-12A were upregulated during acute avian cholera, but only IL-12A was chosen for further analysis. IL-12
and IL-15 are important for driving the Th1 cellular immune response and natural killer (NK) cell activation 380.
IL-12 induces INF-ƴ synthesis and has a proliferative effect on chicken splenocytes. Furthermore, IL-12 in
combination with INF-ƴ function to clear bacteria from the lungs 381.
Expression of IFN-ƴ was upregulated shortly after birds were challenged with strain X73, and again at 14
days P.I. IFN-γ was upregulated to a much lesser extent 7 days P.I. in birds inoculated with strains 775. IFN-ƴ is
expressed by Th1 cells, and is important for macrophage activation, non-specific immunity to microbial
pathogens, and clearance of bacterial infections 382. Additionally, INF-ƴ drives the differentiation of Th0 cells into
Th1 cells, and inhibits the differentiation of Th0 cells into Th2 cells. Activated macrophages are important for
controlling disease progression in chickens 382. However, for pathogens such as Salmonella enteritidis, INF-ƴ
expression does not increase until 2-3 weeks post-infection and is associated with clearance of the pathogen 383.
In addition to a Th1 immune response, a Th17 response was induced in chickens that developed less serious
chronic disease. In mammals, the Th17 response is important for bacterial clearance from the lungs 384, but may
be responsible for inflammation-related injuries in the host 385. Expression of pro-inflammatory cytokine IL-1β
was increased during at 7 days P.I. in birds inoculated with strain X73ΔhyaD. Chicken IL-1β shares homology
and functional similarities to mammalian IL-1β, and is most frequently produced by activated macrophages to

induce fever and activate the cytokine network. Additionally, IL-1β promotes T cell proliferation and
differentiation of Th0 cells into Th17 cells, as well as B cell maturation 386.
Th17-associated chemokine receptor 6 (CCR6) did not display an increase in expression shortly after
challenge, but was increased in expression 7 and 14 days P.I. in birds challenged with strain X73, and at 7 days
P.I. in birds challenged with strain 775. CCR6 is important for the adaptive immune response and T cell
activation. CCR6 traffics dendritic cells to the site of pathogen entry, and after encountering the pathogen, in this
case P. multocida, mature dendritic cells migrate to secondary lymphoid organs and present the antigen to T cells
387. IL-17RA expression was increased for most time points in birds challenged with each P. multocida strain.
Chicken IL-17RA shares 46% identity with human, mouse, and rat IL-17RA, and likely performs the same
functions. To date, research on chicken IL-17 cytokines and receptors have focused on oral infections and gut
immunity 388. Expression of pro-inflammatory, Th17- associated cytokine IL-16 was increased at 7 days P.I. for
all P. multocida strains. Birds inoculated with strain 775 had the greatest increase in expression of IL-16 at 14
days P.I. Overall. both chronic and acute P. multocida infections induced a Th1 and Th17 immune response,
which is consistent with experimental P. multocida intraperitoneal infections in mice 389.
6.6 Conclusion
The present study demonstrates that P. multocida strains of lesser virulence can induce chronic infection in
chickens that are associated with biofilm formation in the pulmonary tissues. P. multocida infection induced a
simultaneous Th1 and Th17 immune response in the spleen during acute and chronic avian cholera, but a
relatively poor antibody response. The cytokine response may aid in recruitment and activation of antigen-
presenting and phagocytic cells, such as macrophages and dendritic cells, and likely provide Th17-mediated,
antibody-independent, memory. No clear differences in immune response were noted between deficient and
proficient biofilm formers with the exception of strain X73ΔhyaD, which induced overall fewer mRNA copies of
the cytokine markers tested.

6.7 Acknowledgements
We would like to thank Shuo Huang, Myeongseon Park, and Anna Champion for excellent technical assistance
with portions of this work, and to Dr. Robert Briggs for providing strains X73 and X73ΔhyaD.
This work was supported by grant 2013-67015-21314 from the U.S. Department of Agriculture-National
Institute of Food and Agriculture to TJI, and funds from the Virginia-Maryland College of Veterinary Medicine
and College of Agriculture and Life Sciences. The funders had no role in study design, data collection and
interpretation, writing of the manuscript, or the decision to submit the work for publication.

REFERENCES
[1] De Alwis MC: Haemorrhagic septicaemia: Australian Centre for International Agricultural Research, 1999.
[2] Bailie W, Stowe E, Schmitt A: Aerobic bacterial flora of oral and nasal fluids of canines with reference to
bacteria associated with bites. Journal of clinical microbiology 1978, 7:223-31.
[3] Bethe A, Wieler LH, Selbitz H-J, Ewers C: Genetic diversity of porcine Pasteurella multocida strains from the
respiratory tract of healthy and diseased swine. Veterinary microbiology 2009, 139:97-105.
[4] Love DN, Vekselstein R, Collings S: The obligate and facultatively anaerobic bacterial flora of the normal
feline gingival margin. Veterinary microbiology 1990, 22:267-75.
[5] Clapper W, Meade G: Normal flora of the nose, throat, and lower intestine of dogs. Journal of bacteriology
1963, 85:643-8.
[6] Woolfrey B, Quall C, Lally R: Pasteurella multocida in an infected tiger bite. Archives of pathology &
laboratory medicine 1985, 109:744-6.
[7] Muhairwa A, Christensen J, Bisgaard M: Investigations on the carrier rate of Pasteurella multocida in healthy
commercial poultry flocks and flocks affected by fowl cholera. Avian Pathol 2000, 29:133-42.
[8] Hubálek Z: An annotated checklist of pathogenic microorganisms associated with migratory birds. Journal of
Wildlife Diseases 2004, 40:639-59.
[9] Capitini CM, Herrero IA, Patel R, Ishitani MB, Boyce TG: Wound infection with Neisseria weaveri and a
novel subspecies of Pasteurella multocida in a child who sustained a tiger bite. Clinical Infectious Diseases 2002,
34:e74-e6.
[10] Kuhnert P, Boerlin P, Emler S, Krawinkler M, Frey J: Phylogenetic analysis of Pasteurella multocida
subspecies and molecular identification of feline P. multocida subsp. septica by 16S rRNA gene sequencing.
International journal of medical microbiology 2000, 290:599-604.
[11] Lehmann KB, Neumann RO: Atlas und Grundriss der Bakteriologie und Lehrbuch der speziellen
bakteriologischen diagnostik: Text: JF Lehmann, 1899.
[12] Yamaguchi H, Tamura T, Abe M, Ogiwara S, Sai S, Kosugiyama K, Sugihara A, Nagumo K, Iwata S,
Kinugawa Y: Prolonged incubation period in neonatal Pasteurella multocida meningitis and bacteremia. Pediatrics
International 2014, 56:e79-e81.
[13] Bardou M, Honnorat E, Dubourg G, Couderc C, Fournier PE, Seng P, Stein A: Meningitis caused by
Pasteurella multocida in a dog owner without a dog bite: clonal lineage identification by MALDI-TOF mass
spectrometry. BMC research notes 2015, 8:626.
[14] Guet-Revillet H, Levy C, Andriantahina I, Kalach N, Pierre M-H, Elbez-Rubinstein A, Boniface C, Berche
P, Cohen R, Ferroni A: Paediatric epidemiology of Pasteurella multocida meningitis in France and review of the
literature. European journal of clinical microbiology & infectious diseases 2013, 32:1111-20.
[15] Tsukadaira A, Nogami A, Yanagida N, Jitsuhara M, Hoshinoo K: [A fatal case of septicemia by serotype A:
3 Pasteurella multocida in human: review of 14 septic cases in Japan]. Rinsho byori The Japanese journal of
clinical pathology 2014, 62:427-31.
[16] Lutz P, Parcina M, Bekeredjian-Ding I, Hoerauf A, Strassburg C, Spengler U: Spontaneous bacterial
peritonitis by Pasteurella multocida under treatment with rifaximin. Infection 2014, 42:175-7.
[17] Teng FY, Cardone JT, Au AH: Pasteurella multocida tubo-ovarian abscess in a virgin. Obstetrics &
Gynecology 1996, 87:883.
[18] Raffi F, Barrier J, Baron D, Drugeon HB, Nicolas F, Courtieu AL: Pasteurella multocida bacteremia: report
of thirteen cases over twelve years and review of the literature. Scandinavian journal of infectious diseases 2009.
[19] Bates HA, Controni G, Elliott N, Eitzman DV: Septicemia and meningitis in a newborn due to Pasteurella
multocida. Clinical pediatrics 1965, 4:668-70.
[20] Hubbert W, Rosen M: Pasteurella multocida infections. II. Pasteurella multocida infection in man unrelated
to animal bite. American Journal of Public Health and the Nations Health 1970, 60:1109-17.
[21] Dendle C, Looke D: Review article: Animal bites: an update for management with a focus on infections.
Emerg Med Australas 2008, 20:458-67.

[22] Murphy E: Microbiology of animal bites. Clinical Microbiology Newsletter 2008, 30:47-50.
[23] Burdge DR, Scheifele D, Speert DP: Serious Pasteurella multocida infections from lion and tiger bites.
JAMA 1985, 253:3296-7.
[24] Emami P, Kaiser TM, Regelsberger J, Goebell E, Fiehler J, Westphal M, Heese O: Case report: surviving a
tiger attack. Neurosurgical review 2012, 35:621-4.
[25] Hubbert WT, Rosen MN: Pasteurella multocida infections: I. Pasteurella multocida infection due to animal
bite. American Journal of Public Health and the Nations Health 1970, 60:1103-8.
[26] López C, Sanchez-Rubio P, Betrán A, Terré R: Pasteurella multocida bacterial meningitis caused by contact
with pigs. Brazilian Journal of Microbiology 2013, 44:473-4.
[27] Raval P, Khan W, Haddad B, Mahapatra AN: Suppl 1: Bite Injuries to the Hand-Review of the Literature.
The open orthopaedics journal 2014, 8:204.
[28] Talley P, Snippes‐Vagnone P, Smith K: Invasive Pasteurella multocida Infections–Report of Five Cases at a
Minnesota Hospital, 2014. Zoonoses and public health 2016.
[29] Talley PP: Increase in Pasteurella multocida Bacteremia in Hospital A-Minnesota 2014. 2015 CSTE Annual
Conference: Cste, 2015.
[30] Siahanidou T, Gika G, Skiathitou A-V, Oikonomopoulos T, Alexandrou-Athanassoulis H, Koutouzis EI,
Syriopoulou VP: Pasteurella multocida infection in a neonate: evidence for a human-to-human horizontal
transmission. The Pediatric infectious disease journal 2012, 31:536-7.
[31] Baillot R, Voisine P, Cote LM, Longtin Y: Deep sternal wound infection due to Pasteurella multocida: the
first case report and review of literature. Infection 2011, 39:575-8.
[32] Boyce JD, Chung JY, Adler B: Pasteurella multocida capsule: composition, function and genetics. Journal of
biotechnology 2000, 83:153-60.
[33] Rosner H, Grimmecke H-D, Knirel YA, Shashkov AS: Hyaluronic acid and a (1→ 4)-β-D-xylan,
extracellular polysaccharides of Pasteurella multocida (Carter type A) strain 880. Carbohydrate research 1992,
223:329-33.
[34] Rimler R, Rhoades K: Serogroup F, a new capsule serogroup of Pasteurella multocida. Journal of clinical
microbiology 1987, 25:615-8.
[35] DeAngelis PL, Gunay NS, Toida T, Mao W-j, Linhardt RJ: Identification of the capsular polysaccharides of
Type D and F Pasteurella multocida as unmodified heparin and chondroitin, respectively. Carbohydrate Research
2002, 337:1547-52.
[36] Rimler R, Brogden K: Pasteurella multocida isolated from rabbits and swine: serologic types and toxin
production. American journal of veterinary research 1986, 47:730-7.
[37] Moustafa AM, Seemann T, Gladman S, Adler B, Harper M, Boyce JD, Bennett MD: Comparative Genomic
Analysis of Asian Haemorrhagic Septicaemia-Associated Strains of Pasteurella multocida Identifies More than 90
Haemorrhagic Septicaemia-Specific Genes. PloS one 2015, 10:e0130296.
[38] Harper M, Boyce JD, Adler B: The key surface components of Pasteurella multocida: capsule and
lipopolysaccharide. Curr Top Microbiol Immunol 2012, 361:39-51.
[39] Boyce JD, Adler B: The capsule is a virulence determinant in the pathogenesis of Pasteurella multocida
M1404 (B:2). Infect Immun 2000, 68:3463-8.
[40] DeAngelis PL, White CL: Identification of a distinct, cryptic heparosan synthase from Pasteurella multocida
types A, D, and F. Journal of bacteriology 2004, 186:8529-32.
[41] Chung JY, Zhang Y, Adler B: The capsule biosynthetic locus of Pasteurella multocida A: 1. FEMS
microbiology letters 1998, 166:289-96.
[42] Steen JA, Harrison P, Seemann T, Wilkie I, Harper M, Adler B, Boyce JD: Fis is essential for capsule
production in Pasteurella multocida and regulates expression of other important virulence factors. PLoS Pathog
2010, 6:e1000750.
[43] Townsend KM, Boyce JD, Chung JY, Frost AJ, Adler B: Genetic organization of Pasteurella multocida cap
loci and development of a multiplex capsular PCR typing system. Journal of Clinical Microbiology 2001, 39:924-
9.

[44] Blackburn BO, Heddleston KL, Pfow CJ: Pasteurella multocida serotyping results (1971-1973). Avian
diseases 1975:353-6.
[45] Harper M, Cox AD, Adler B, Boyce JD: Pasteurella multocida lipopolysaccharide: The long and the short of
it. Veterinary microbiology 2011, 153:109-15.
[46] Harper M, Michael FS, John M, Vinogradov E, Adler B, Boyce JD, Cox AD: Pasteurella multocida
Heddleston serovars 1 and 14 express different lipopolysaccharide structures but share the same
lipopolysaccharide biosynthesis outer core locus. Veterinary microbiology 2011, 150:289-96.
[47] Harper M, St Michael F, Vinogradov E, John M, Steen JA, van Dorsten L, Boyce JD, Adler B, Cox AD:
Structure and biosynthetic locus of the lipopolysaccharide outer core produced by Pasteurella multocida serovars
8 and 13 and the identification of a novel phospho-glycero moiety. Glycobiology 2013, 23:286-94.
[48] Michael FS, Harper M, Parnas H, John M, Stupak J, Vinogradov E, Adler B, Boyce JD, Cox AD: Structural
and genetic basis for the serological differentiation of Pasteurella multocida Heddleston serotypes 2 and 5. Journal
of bacteriology 2009, 191:6950-9.
[49] Harper M, Michael FS, Vinogradov E, John M, Boyce JD, Adler B, Cox AD: Characterization of the
lipopolysaccharide from Pasteurella multocida Heddleston serovar 9: Identification of a proposed bi-functional
dTDP-3-acetamido-3, 6-dideoxy-α-d-glucose biosynthesis enzyme. Glycobiology 2012, 22:332-44.
[50] Boyce JD, Wilkie I, Harper M, Paustian ML, Kapur V, Adler B: Genomic scale analysis of Pasteurella
multocida gene expression during growth within the natural chicken host. Infection and Immunity 2002, 70:6871-
9.
[51] May BJ, Zhang Q, Li LL, Paustian ML, Whittam TS, Kapur V: Complete genomic sequence of Pasteurella
multocida, Pm70. Proc Natl Acad Sci U S A 2001, 98:3460-5.
[52] Verma S, Sharma M, Katoch S, Verma L, Kumar S, Dogra V, Chahota R, Dhar P, Singh G: Profiling of
virulence associated genes of Pasteurella multocida isolated from cattle. Vet Res Commun 2013, 37:83-9.
[53] Nakai T, Sawata A, Tsuji M, Samejima Y, Kume K: Purification of dermonecrotic toxin from a sonic extract
of Pasteurella multocida SP-72 serotype D. Infection and immunity 1984, 46:429-34.
[54] Nakai T, Sawata A, Tsuji M, Kume K: Characterization of dermonecrotic toxin produced by serotype D
strains of Pasteurella multocida. American journal of veterinary research 1984, 45:2410-3.
[55] Magyar T, Rimler R: Detection and enumeration of toxin-producing Pasteurella multocida with a colony-blot
assay. Journal of clinical microbiology 1991, 29:1328-32.
[56] Hatfaludi T, Al-Hasani K, Boyce JD, Adler B: Outer membrane proteins of Pasteurella multocida. Veterinary
microbiology 2010, 144:1-17.
[57] Ali HAH, Sawada T, Hatakeyama H, Katayama Y, Ohtsuki N, Itoh O: Invasion of chicken embryo fibroblast
cells by avian Pasteurella multocida. Veterinary Microbiology 2004, 104:55-62.
[58] Galdiero M, De Martino L, Pagnini U, Pisciotta MG, Galdiero E: Interactions between bovine endothelial
cells and Pasteurella multocida: association and invasion. Research in Microbiology 2001, 152:57-65.
[59] Dabo SM, Confer AW, Hartson SD: Adherence of Pasteurella multocida to fibronectin. Vet Microbiol 2005,
110:265-75.
[60] Maheswaran SK, Thies ES: Influence of encapsulation on phagocytosis of Pasteurella multocida by bovine
neutrophils. Infect Immun 1979, 26:76-81.
[61] Fuller TE, Kennedy MJ, Lowery DE: Identification of Pasteurella multocida virulence genes in a septicemic
mouse model using signature-tagged mutagenesis. Microb Pathog 2000, 29:25-38.
[62] Harper M, John M, Edmunds M, Wright A, Ford M, Turni C, Blackall P, Cox A, Adler B, Boyce JD:
Protective efficacy afforded by live Pasteurella multocida vaccines in chickens is independent of
lipopolysaccharide outer core structure. Vaccine 2016, 34:1696-703.
[63] Heddleston K, Gallagher J, Rebers P: Fowl cholera: gel diffusion precipitin test for serotyping Pasteurella
multocida from avian species. Avian Dis 1972:925-36.
[64] Horadagoda N, Hodgson J, Moon G, Wijewardana T, Eckersall P: Development of a clinical syndrome
resembling haemorrhagic septicaemia in the buffalo following intravenous inoculation of Pasteurella multocida
serotype B: 2 endotoxin and the role of tumour necrosis factor-α. Research in veterinary science 2002, 72:194-
200.

[65] Aderem A, Ulevitch RJ: Toll-like receptors in the induction of the innate immune response. Nature 2000,
406:782-7.
[66] Michael FS, Vinogradov E, Li J, Cox AD: Structural analysis of the lipopolysaccharide from Pasteurella
multocida genome strain Pm70 and identification of the putative lipopolysaccharide glycosyltransferases.
Glycobiology 2005, 15:323-33.
[67] Adler R, Adler B: Opsonic monoclonal antibodies against lipopolysaccharide (LPS) antigens of Pasteurella
multocida and the role of LPS in immunity. Veterinary microbiology 1991, 26:335-47.
[68] Wijewardana TG, Wilson CF, Gilmour N: Production of mouse monoclonal antibodies to Pasteurella
multocida type A and the immunological properties of a protective anti-lipopolysaccharide antibody. Journal of
medical microbiology 1990, 33:217-22.
[69] Koebnik R, Locher KP, Van Gelder P: Structure and function of bacterial outer membrane proteins: barrels
in a nutshell. Molecular microbiology 2000, 37:239-53.
[70] Carpenter T, Khalid S, Sansom MS: A multidomain outer membrane protein from Pasteurella multocida:
modelling and simulation studies of PmOmpA. Biochimica et Biophysica Acta (BBA)-Biomembranes 2007,
1768:2831-40.
[71] Smith SG, Mahon V, Lambert MA, Fagan RP: A molecular Swiss army knife: OmpA structure, function and
expression. FEMS microbiology letters 2007, 273:1-11.
[72] Dabo S, Confer A, Quijano-Blas R: Molecular and immunological characterization of Pasteurella multocida
serotype A: 3 OmpA: evidence of its role in P. multocida interaction with extracellular matrix molecules.
Microbial pathogenesis 2003, 35:147-57.
[73] Vachon V, Laprade R, Coulton JW: Properties of the porin of Haemophilus influenzae type b in planar lipid
bilayer membranes. Biochimica et Biophysica Acta (BBA)-Biomembranes 1986, 861:74-82.
[74] Luo Y, Zeng Q, Glisson JR, Jackwood MW, Cheng I-HN, Wang C: Sequence analysis of Pasteurella
multocida major outer membrane protein (OmpH) and application of synthetic peptides in vaccination of chickens
against homologous strain challenge. Vaccine 1999, 17:821-31.
[75] Luo Y, Glisson JR, Jackwood MW, Hancock R, Bains M, Cheng I, Wang C: Cloning and characterization of
the major outer membrane protein gene (ompH) of Pasteurella multocida X-73. Journal of Bacteriology 1997,
179:7856-64.
[76] Sthitmatee N, Numee S, Kawamoto E, Sasaki H, Yamashita K, Takahashi N, Kataoka Y, Sawada T:
Protection of chickens from fowl cholera by vaccination with recombinant adhesive protein of Pasteurella
multocida. Vaccine 2008, 26:2398-407.
[77] Lee J, Kim YB, Kwon M: Outer membrane protein H for protective immunity against Pasteurella multocida.
Journal of microbiology (Seoul, Korea) 2007, 45:179-84.
[78] Gong Q, Qu N, Niu M, Qin C, Cheng M, Sun X, Zhang A: Immune responses and protective efficacy of a
novel DNA vaccine encoding outer membrane protein of avian Pasteurella multocida. Vet Immunol
Immunopathol 2013, 152:317-24.
[79] Deich R, Anilionis A, Fulginiti J, Metcalf B, Quataert S, Quinn-Dey T, Zlotnick G, Green B: Antigenic
conservation of the 15,000-dalton outer membrane lipoprotein PCP of Haemophilus influenzae and biologic
activity of anti-PCP antisera. Infection and immunity 1990, 58:3388-93.
[80] Lo M, Boyce JD, Wilkie IW, Adler B: Characterization of two lipoproteins in Pasteurella multocida.
Microbes and infection 2004, 6:58-67.
[81] Green B, Vazquez M, Zlotnick G, Quigley-Reape G, Swarts J, Green I, Cowell J, Bluestone C, Doyle W:
Evaluation of mixtures of purified Haemophilus influenzae outer membrane proteins in protection against
challenge with nontypeable H. influenzae in the chinchilla otitis media model. Infection and immunity 1993,
61:1950-7.
[82] Kasten RW, Wakenell PS, Ahmad S, Yilma TD, Hirsh DC: Lack of protection against avian cholera by
vaccination with recombinant P6-like protein from Pasteurella multocida. Avian Dis 1997:972-6.
[83] Boyce JD, Cullen PA, Nguyen V, Wilkie I, Adler B: Analysis of the Pasteurella multocida outer membrane
sub‐proteome and its response to the in vivo environment of the natural host. Proteomics 2006, 6:870-80.

[84] Rimler RB: Purification of a cross-protective antigen from Pasteurella multocida grown in vitro and in vivo.
Avian Dis 2001:572-80.
[85] Tabatabai LB, Zehr ES: Identification of five outer membrane-associated proteins among cross-protective
factor proteins of Pasteurella multocida. Infection and immunity 2004, 72:1195-8.
[86] Wu J-R, Shien J-H, Shieh HK, Chen C-F, Chang P-C: Protective immunity conferred by recombinant
Pasteurella multocida lipoprotein E (PlpE). Vaccine 2007, 25:4140-8.
[87] Dabo SM, Taylor JD, Confer AW: Pasteurella multocida and bovine respiratory disease. Anim Health Res
Rev 2007, 8:129-50.
[88] Ruffolo CG, Tennent JM, Michalski WP, Adler B: Identification, purification, and characterization of the
type 4 fimbriae of Pasteurella multocida. Infection and immunity 1997, 65:339-43.
[89] Mullen LM, Bosse JT, Nair SP, Ward JM, Rycroft AN, Robertson G, Langford PR, Henderson B:
Pasteurellaceae ComE1 proteins combine the properties of fibronectin adhesins and DNA binding competence
proteins. PLoS One 2008, 3:e3991.
[90] Tatum FM, Yersin AG, Briggs RE: Construction and virulence of a Pasteurella multocida fhaB2 mutant in
turkeys. Microbial Pathogenesis 2005, 39:9-17.
[91] Tatum FM, Tabatabai LB, Briggs RE: Protection against fowl cholera conferred by vaccination with
recombinant Pasteurella multocida filamentous hemagglutinin peptides. Avian Dis 2009, 53:169-74.
[92] Tomich M, Planet PJ, Figurski DH: The tad locus: postcards from the widespread colonization island. Nature
Reviews Microbiology 2007, 5:363-75.
[93] May BJ, Zhang Q, Li LL, Paustian ML, Whittam TS, Kapur V: Complete genomic sequence of Pasteurella
multocida, Pm70. Proceedings of the National Academy of Sciences 2001, 98:3460-5.
[94] Krewulak KD, Vogel HJ: Structural biology of bacterial iron uptake. Biochimica et Biophysica Acta (BBA)-
Biomembranes 2008, 1778:1781-804.
[95] Bosch M, Garrido E, Llagostera M, de Rozas AMP, Badiola I, Barbé J: Pasteurella multocida exbB, exbD
and tonB genes are physically linked but independently transcribed. FEMS microbiology letters 2002, 210:201-8.
[96] Paustian ML, May BJ, Cao D, Boley D, Kapur V: Transcriptional response of Pasteurella multocida to
defined iron sources. Journal of bacteriology 2002, 184:6714-20.
[97] Choi-Kim K, Maheswaran SK, Felice LJ, Molitor TW: Relationship between the iron regulated outer
membrane proteins and the outer membrane proteins of in vivo grown Pasteurella multocida. Veterinary
microbiology 1991, 28:75-92.
[98] Ogunnariwo JA, Schryvers AB: Characterization of a novel transferrin receptor in bovine strains of
Pasteurella multocida. Journal of bacteriology 2001, 183:890-6.
[99] Krieg S, Huché F, Diederichs K, Izadi-Pruneyre N, Lecroisey A, Wandersman C, Delepelaire P, Welte W:
Heme uptake across the outer membrane as revealed by crystal structures of the receptor–hemophore complex.
Proceedings of the National Academy of Sciences 2009, 106:1045-50.
[100] Prado ME, Dabo S, Confer A: Immunogenicity of iron-regulated outer membrane proteins of Pasteurella
multocida A: 3 in cattle: molecular characterization of the immunodominant heme acquisition system receptor
(HasR) protein. Veterinary microbiology 2005, 105:269-80.
[101] Bosch M, Garrido ME, de Rozas AMP, Badiola I, Barbé J, Llagostera M: Pasteurella multocida contains
multiple immunogenic haemin-and haemoglobin-binding proteins. Veterinary microbiology 2004, 99:103-12.
[102] Steenbergen SM, Lichtensteiger CA, Caughlan R, Garfinkle J, Fuller TE, Vimr ER: Sialic Acid metabolism
and systemic pasteurellosis. Infect Immun 2005, 73:1284-94.
[103] Mizan S, Henk A, Stallings A, Maier M, Lee MD: Cloning and characterization of sialidases with 2-6′ and
2-3′ sialyl lactose specificity from Pasteurella multocida. Journal of bacteriology 2000, 182:6874-83.
[104] Abdullahi MZ, Gilmour NJ, Poxton IR: Outer membrane proteins of bovine strains of Pasteurella multocida
type A and their doubtful role as protective antigens. J Med Microbiol 1990, 32:55-61.
[105] Horiguchi Y: Swine atrophic rhinitis caused by pasteurella multocida toxin and bordetella dermonecrotic
toxin. Curr Top Microbiol Immunol 2012, 361:113-29.
[106] Hunt ML, Adler B, Townsend KM: The molecular biology of Pasteurella multocida. Veterinary
microbiology 2000, 72:3-25.

[107] Pullinger GD, Bevir T, Lax AJ: The Pasteurella multocida toxin is encoded within a lysogenic
bacteriophage. Molecular microbiology 2004, 51:255-69.
[108] Jarvinen L, Hogenesch H, Suckow M, Bowersock T: Induction of protective immunity in rabbits by
coadministration of inactivated Pasteurella multocida toxin and potassium thiocyanate extract. Infection and
immunity 1998, 66:3788-95.
[109] Suckow M: Immunization of rabbits against Pasteurella multocida using a commercial swine vaccine.
Laboratory animals 2000, 34:403-8.
[110] Jarvinen LZ, HogenEsch H, Suckow MA, Bowersock TL: Intranasal Vaccination of New Zealand White
Rabbits Against Pasteurellosis, Using AlginateEncapsulated Pasteurella multocida Toxin and Potassium
Thiocyanate Extract. Comparative medicine 2000, 50:263-9.
[111] Horiguchi Y: Escherichia coli cytotoxic necrotizing factors and Bordetella dermonecrotic toxin: the
dermonecrosis-inducing toxins activating Rho small GTPases. Toxicon 2001, 39:1619-27.
[112] Horiguchi Y, Nakai T, Kume K: Purification and characterization of Bordetella bronchiseptica
dermonecrotic toxin. Microbial pathogenesis 1989, 6:361-8.
[113] Baldwin MR, Lakey JH, Lax AJ: Identification and characterization of the Pasteurella multocida toxin
translocation domain. Mol Microbiol 2004, 54:239-50.
[114] Horiguchi Y: Swine atrophic rhinitis caused by Pasteurella multocida toxin and Bordetella dermonecrotic
toxin. Pasteurella multocida: Springer, 2012. pp. 113-29.
[115] Diallo IS, Frost AJ: Characteristics of a haemolytic extract from avian Pasteurella multocida. Vet Microbiol
2000, 72:37-45.
[116] Diallo IS, Frost AJ: Survival of avian strains of Pasteurella multocida in chicken serum. Vet Microbiol
2000, 72:153-61.
[117] Negrete-Abascal E, Tenorio VR, de la Garza M: Secretion of proteases from Pasteurella multocida isolates.
Current Microbiology 1999, 38:64-7.
[118] Olson ME, Ceri H, Morck DW, Buret AG, Read RR: Biofilm bacteria: formation and comparative
susceptibility to antibiotics. Can J Vet Res 2002, 66:86-92.
[119] Ross RF: Pasteurella multocida and its role in porcine pneumonia. Animal health research reviews /
Conference of Research Workers in Animal Diseases 2006, 7:13-29.
[120] Sandal I, Shao JQ, Annadata S, Apicella MA, Boye M, Jensen TK, Saunders GK, Inzana TJ: Histophilus
somni biofilm formation in cardiopulmonary tissue of the bovine host following respiratory challenge. Microbes
and infection / Institut Pasteur 2009, 11:254-63.
[121] Costerton JW, Lewandowski Z, Caldwell DE, Korber DR, Lappin-Scott HM: Microbial biofilms. Annual
review of microbiology 1995, 49:711-45.
[122] Kuramitsu HK, He X, Lux R, Anderson MH, Shi W: Interspecies interactions within oral microbial
communities. Microbiology and molecular biology reviews : MMBR 2007, 71:653-70.
[123] Costerton JW, Cheng K, Geesey GG, Ladd TI, Nickel JC, Dasgupta M, Marrie TJ: Bacterial biofilms in
nature and disease. Annual Reviews in Microbiology 1987, 41:435-64.
[124] Hall-Stoodley L, Costerton JW, Stoodley P: Bacterial biofilms: from the natural environment to infectious
diseases. Nature reviews microbiology 2004, 2:95-108.
[125] Parsek MR, Singh PK: Bacterial biofilms: an emerging link to disease pathogenesis. Annual Reviews in
Microbiology 2003, 57:677-701.
[126] Sutherland IW: The biofilm matrix–an immobilized but dynamic microbial environment. Trends in
microbiology 2001, 9:222-7.
[127] Hausner M, Wuertz S: High rates of conjugation in bacterial biofilms as determined by quantitative in situ
analysis. Applied and Environmental Microbiology 1999, 65:3710-3.
[128] Ghigo J-M: Natural conjugative plasmids induce bacterial biofilm development. Nature 2001, 412:442-5.
[129] Ziebuhr W, Krimmer V, Rachid S, Lößner I, Götz F, Hacker J: A novel mechanism of phase variation of
virulence in Staphylococcus epidermidis: evidence for control of the polysaccharide intercellular adhesin
synthesis by alternating insertion and excision of the insertion sequence element IS256. Molecular microbiology
1999, 32:345-56.

[130] Fux C, Costerton J, Stewart P, Stoodley P: Survival strategies of infectious biofilms. Trends in
microbiology 2005, 13:34-40.
[131] Déziel E, Comeau Y, Villemur R: Initiation of biofilm formation byPseudomonas aeruginosa 57RP
correlates with emergence of hyperpiliated and highly adherent phenotypic variants deficient in swimming,
swarming, and twitching motilities. Journal of Bacteriology 2001, 183:1195-204.
[132] Romby P, Vandenesch F, Wagner EGH: The role of RNAs in the regulation of virulence-gene expression.
Current opinion in microbiology 2006, 9:229-36.
[133] Sobrero P, Valverde C: The bacterial protein Hfq: much more than a mere RNA-binding factor. Critical
reviews in microbiology 2012, 38:276-99.
[134] Gottesman S, Storz G: Bacterial small RNA regulators: versatile roles and rapidly evolving variations. Cold
Spring Harbor perspectives in biology 2011, 3:a003798.
[135] Vogel J, Luisi BF: Hfq and its constellation of RNA. Nature Reviews Microbiology 2011, 9:578-89.
[136] Møller T, Franch T, Højrup P, Keene DR, Bächinger HP, Brennan RG, Valentin-Hansen P: Hfq: a bacterial
Sm-like protein that mediates RNA-RNA interaction. Molecular cell 2002, 9:23-30.
[137] Brennan RG, Link TM: Hfq structure, function and ligand binding. Current opinion in microbiology 2007,
10:125-33.
[138] Folichon M, Arluison V, Pellegrini O, Huntzinger E, Régnier P, Hajnsdorf E: The poly (A) binding protein
Hfq protects RNA from RNase E and exoribonucleolytic degradation. Nucleic acids research 2003, 31:7302-10.
[139] Chao Y, Vogel J: The role of Hfq in bacterial pathogens. Current opinion in microbiology 2010, 13:24-33.
[140] Mégroz M, Kleifeld O, Wright A, Powell D, Harrison P, Adler B, Harper M, Boyce JD: The RNA-binding
chaperone Hfq is an important global regulator of gene expression in Pasteurella multocida and plays a crucial
role in production of a number of virulence factors, including hyaluronic acid capsule. Infection and immunity
2016, 84:1361-70.
[141] Saldaña Z, Xicohtencatl‐Cortes J, Avelino F, Phillips AD, Kaper JB, Puente JL, Girón JA: Synergistic role
of curli and cellulose in cell adherence and biofilm formation of attaching and effacing Escherichia coli and
identification of Fis as a negative regulator of curli. Environmental microbiology 2009, 11:992-1006.
[142] Goldberg M, Johnson M, Hinton J, Williams PH: Role of the nucleoid‐associated protein Fis in the
regulation of virulence properties of enteropathogenic Escherichia coli. Molecular microbiology 2001, 41:549-59.
[143] Kelly A, Goldberg MD, Carroll RK, Danino V, Hinton JC, Dorman CJ: A global role for Fis in the
transcriptional control of metabolism and type III secretion in Salmonella enterica serovar Typhimurium.
Microbiology 2004, 150:2037-53.
[144] Lautier T, Nasser W: The DNA nucleoid‐associated protein Fis co‐ordinates the expression of the main
virulence genes in the phytopathogenic bacterium Erwinia chrysanthemi. Molecular microbiology 2007, 66:1474-
90.
[145] Lenz DH, Bassler BL: The small nucleoid protein Fis is involved in Vibrio cholerae quorum sensing.
Molecular microbiology 2007, 63:859-71.
[146] Steen JA, Steen JA, Harrison P, Seemann T, Wilkie I, Harper M, Adler B, Boyce JD: Fis is essential for
capsule production in Pasteurella multocida and regulates expression of other important virulence factors. PLoS
Pathog 2010, 6:e1000750-e.
[147] Liu D, Lawrence ML, Austin FW: Specific PCR identification of Pasteurella multocida based on putative
transcriptional regulator genes. Journal of microbiological methods 2004, 58:263-7.
[148] Wilkie IW, Harper M, Boyce JD, Adler B: Pasteurella multocida: diseases and pathogenesis. Current topics
in microbiology and immunology 2012, 361:1-22.
[149] Botzler RG: Epizootiology of avian cholera in wildfowl. Journal of Wildlife Diseases 1991, 27:367-95.
[150] Samuel MD, Shadduck DJ, Goldberg DR, Johnson WP: Avian cholera in waterfowl: the role of lesser snow
and ross's geese as disease carriers in the Playa Lakes Region. J Wildl Dis 2005, 41:48-57.
[151] Hunter B, Wobeser G: Pathology of Experimental Avian Cholera in Mallard Ducks. Avian Dis 1980,
24:403-14.
[152] Leotta G, Chinen I, Vigo G, Pecoraro M, Rivas M: Outbreaks of avian cholera in Hope Bay, Antarctica.
Journal of wildlife diseases 2006, 42:259-70.

[153] Harms NJ: Avian Cholera in the Eastern Canadian Arctic: Investigating Disease Origins and Reservoirs.
Arctic 2011, 64:501-5.
[154] Iverson S, Forbes M, Simard M, Soos C, Gilchrist HG: Avian Cholera emergence in Arctic-nesting northern
Common Eiders: using community-based, participatory surveillance to delineate disease outbreak patterns and
predict transmission risk. Ecology and Society 2016, 21.
[155] Quortrup E, Queen F, Merovka L: An outbreak of pasteurellosis in wild ducks. Journal of the American
Veterinary Medical Association 1946, 108:94-100.
[156] Bredy JP, Botzler RG: The effects of six environmental variables on Pasteurella multocida populations in
water. Journal of Wildlife Diseases 1989, 25:232-9.
[157] Rosen M, Bischoff A: The 1948-49 outbreak of fowl cholera in birds in the San Francisco Bay area and
surrounding counties. California Fish and Game 1949, 35:185-92.
[158] ROSEN MN: SPECIES SUSCEPTIBILITY TO AVIAN CHOLERA 1. Bulletin of the Wildlife Disease
Association 1969, 5:195-200.
[159] Simensen E, Olson L, Hahn G: Influence of Pasteurella multocida and high and low environmental
temperatures on adrenals and bursa of Fabricius in turkeys. Avian Dis 1980:844-67.
[160] Simensen E, Olson L: Aerosol transmission of Pasteurella multocida in turkeys. Avian Dis 1980:1007-10.
[161] Miguel B, Wang C, Maslin WR, Keirs RW, Glisson JR: Subacute to chronic fowl cholera in a flock of
Pharaoh breeder quail. Avian Dis 1998, 42:204-8.
[162] Petrov D: Studies on the gamasid red mite of poultry, Dermanyssus gallinae, as a carrier of Pasteurella
multocida. Veterinarnomedit︠ s︡inski nauki= 1975.
[163] Bigland C: A rabbit infestation with poultry mites and experimental mite transmission of fowl cholera.
Canadian journal of comparative medicine and veterinary science 1954, 18:213.
[164] Rosen MN, Bischoff AI: DISEASE, NUTRITION AND CONTROL PROBLEMS. Transactions of the
North American Wildlife Conference: American Wildlife Institute, 1950. p. 147.
[165] Dimov I: Survival of avian Pasteurella multocida in soils at different acidity, humidity and temperature.
Nauchni Trudove Vissh Veterinarnomeditsinski Institute ‘‘Prof G Pavlov 1964, 12:339-45.
[166] Korbel R: Epizootiology, clinical aspects and therapy of Pasteurella multocida infection in bird patients
after cat bite. Tierarztliche Praxis 1990, 18:365-76.
[167] Gregg D, Olson L, McCune E: Experimental transmission of Pasteurella multocida from raccoons to
turkeys via bite wounds. Avian Dis 1974:559-64.
[168] Mbuthia PG, Njagi LW, Nyaga PN, Bebora LC, Minga U, Christensen JP, Olsen JE: Time-course
investigation of infection with a low virulent Pasteurella multocida strain in normal and immune-suppressed 12-
week-old free-range chickens. Avian Pathol 2011, 40:629-37.
[169] Iliev T, Arsov R, Lazarov V: Can fowls, carriers of pasteurella excrete the organism in the fae-ces. Vet Bull
1965, 36:710.
[170] Curtis P, Ollerhead G: Investigation to determine whether healthy chickens and turkeys are oral carriers of
Pasteurella multocida. The Veterinary Record 1981, 108:206-7.
[171] Hughes TP: THE EPIDEMIOLOGY OF FOWL CHOLERA : II. BIOLOGICAL PROPERTIES OF P.
AVICIDA. The Journal of Experimental Medicine 1930, 51:225-38.
[172] Rimler RB, Rhoades KR: Pasteurella multocida. Pasteurella and pasteurellosis 1989, 1:37-73.
[173] Ficken M, Barnes H: Acute airsacculitis in turkeys inoculated with Pasteurella multocida. Veterinary
Pathology Online 1989, 26:231-7.
[174] Rhoades KR: The microscopic lesions of acute fowl cholera in mature chickens. Avian Dis 1964, 8:658-65.
[175] Pritchett IW, Hughes TP: The Epidemiology of Fowl Cholera : Vi. The Spread of Epidemic and Endemic
Strains of Pasteurella Avicida in Laboratory Populations of Normal Fowl. J Exp Med 1932, 55:71-8.
[176] Harper M, Boyce JD, Adler B: Pasteurella multocida pathogenesis: 125 years after Pasteur. FEMS
Microbiol Lett 2006, 265:1-10.
[177] Holman DB, Timsit E, Amat S, Abbott DW, Buret AG, Alexander TW: The nasopharyngeal microbiota of
beef cattle before and after transport to a feedlot. BMC microbiology 2017, 17:70.

[178] Snowder G, Van Vleck LD, Cundiff L, Bennett G: Bovine respiratory disease in feedlot cattle:
environmental, genetic, and economic factors. Journal of Animal Science 2006, 84:1999-2008.
[179] Ames TR: Dairy calf pneumonia: the disease and its impact. Veterinary Clinics of North America: Food
Animal Practice 1997, 13:379-91.
[180] Taylor JD, Fulton RW, Lehenbauer TW, Step DL, Confer AW: The epidemiology of bovine respiratory
disease: What is the evidence for predisposing factors. Can Vet J 2010, 51:1095-102.
[181] Cusack PM, McMeniman N, Lean IJ: The medicine and epidemiology of bovine respiratory disease in
feedlots. Aust Vet J 2003, 81:480-7.
[182] Hotchkiss EJ, Dagleish MP, Willoughby K, McKendrick IJ, Finlayson J, Zadoks RN, Newsome E,
Brulisauer F, Gunn GJ, Hodgson JC: Prevalence of Pasteurella multocida and other respiratory pathogens in the
nasal tract of Scottish calves. Veterinary Record 2010, 167:555-60.
[183] Miles DG: Overview of the North American beef cattle industry and the incidence of bovine respiratory
disease (BRD). Animal health research reviews / Conference of Research Workers in Animal Diseases 2009,
10:101-3.
[184] Griffin D: Economic impact associated with respiratory disease in beef cattle. Veterinary clinics of North
America: food animal practice 1997, 13:367-77.
[185] Stovall T, Gill D, Smith R, Ball R: Impact of bovine respiratory disease during the receiving period on
feedlot performance and carcass traits. Animal Science Research Report, Oklahoma State University http://www
ansi okstate edu/research/2000rr/16 htm 2000.
[186] Rice J, Carrasco-Medina L, Hodgins D, Shewen P: Mannheimia haemolytica and bovine respiratory
disease. Animal Health Research Reviews 2007, 8:117-28.
[187] Arcangioli M-A, Duet A, Meyer G, Dernburg A, Bézille P, Poumarat F, Le Grand D: The role of
Mycoplasma bovis in bovine respiratory disease outbreaks in veal calf feedlots. The Veterinary Journal 2008,
177:89-93.
[188] Bryson D, McFerran J, Ball H, Neill S: Observations on outbreaks of respiratory disease in housed calves--
(1) Epidemiological, clinical and microbiological findings. The Veterinary Record 1978, 103:485-9.
[189] Virtala A, Gröhn Y, Mechor G, Erb H, Dubovi E: Association of seroconversion with isolation of agents in
transtracheal wash fluids collected from pneumonic calves less than three months of age. Bovine Practitioner
2000, 34:77-80.
[190] Nikunen S, Härtel H, Orro T, Neuvonen E, Tanskanen R, Kivelä S-L, Sankari S, Aho P, Pyörälä S,
Saloniemi H: Association of bovine respiratory disease with clinical status and acute phase proteins in calves.
Comparative immunology, microbiology and infectious diseases 2007, 30:143-51.
[191] Griffin D, Chengappa M, Kuszak J, McVey DS: Bacterial pathogens of the bovine respiratory disease
complex. Veterinary Clinics of North America: Food Animal Practice 2010, 26:381-94.
[192] Gagea MI, Bateman KG, van Dreumel T, McEwen BJ, Carman S, Archambault M, Shanahan RA, Caswell
JL: Diseases and pathogens associated with mortality in Ontario beef feedlots. Journal of Veterinary Diagnostic
Investigation 2006, 18:18-28.
[193] Murray GM, More SJ, Sammin D, Casey MJ, McElroy MC, O’Neill RG, Byrne WJ, Earley B, Clegg TA,
Ball H: Pathogens, patterns of pneumonia, and epidemiologic risk factors associated with respiratory disease in
recently weaned cattle in Ireland. Journal of Veterinary Diagnostic Investigation 2017, 29:20-34.
[194] Agnes J, Zekarias B, Shao M, Anderson M, Gershwin L, Corbeil L: Bovine respiratory syncytial virus and
Histophilus somni interaction at the alveolar barrier. Infection and immunity 2013, 81:2592-7.
[195] Booker CW, Abutarbush SM, Morley PS, Jim GK, Pittman TJ, Schunicht OC, Perrett T, Wildman BK,
Fenton RK, Guichon PT: Microbiological and histopathological findings in cases of fatal bovine respiratory
disease of feedlot cattle in western Canada. Canadian Veterinary Journal 2008, 49:473-82.
[196] Sibley CD, Duan K, Fischer C, Parkins MD, Storey DG, Rabin HR, Surette MG: Discerning the complexity
of community interactions using a Drosophila model of polymicrobial infections. PLoS Pathog 2008, 4:e1000184.
[197] Murray JL, Connell JL, Stacy A, Turner KH, Whiteley M: Mechanisms of synergy in polymicrobial
infections. Journal of Microbiology 2014, 52:188-99.

[198] Burmølle M, Ren D, Bjarnsholt T, Sørensen SJ: Interactions in multispecies biofilms: do they actually
matter? Trends in microbiology 2014, 22:84-91.
[199] Boukahil I, Czuprynski CJ: Characterization of Mannheimia haemolytica biofilm formation in vitro. Vet
Microbiol 2015, 175:114-22.
[200] Simmons WL, Dybvig K: Mycoplasma biofilms ex vivo and in vivo. FEMS microbiology letters 2009,
295:77-81.
[201] Olson ME, Ceri H, Morck DW, Buret AG, Read RR: Biofilm bacteria: formation and comparative
susceptibility to antibiotics. Canadian Journal of Veterinary Research 2002, 66:86-92.
[202] Ehrlich GD, Hu FZ, Shen K, Stoodley P, Post JC: Bacterial Plurality as a General Mechanism Driving
Persistence in Chronic Infections. Clinical orthopaedics and related research 2005:20-4.
[203] Weigel LM, Donlan RM, Shin DH, Jensen B, Clark NC, McDougal LK, Zhu W, Musser KA, Thompson J,
Kohlerschmidt D: High-level vancomycin-resistant Staphylococcus aureus isolates associated with a
polymicrobial biofilm. Antimicrobial agents and chemotherapy 2007, 51:231-8.
[204] Molin S, Tolker-Nielsen T: Gene transfer occurs with enhanced efficiency in biofilms and induces enhanced
stabilisation of the biofilm structure. Current opinion in biotechnology 2003, 14:255-61.
[205] Harriott MM, Noverr MC: Candida albicans and Staphylococcus aureus form polymicrobial biofilms:
effects on antimicrobial resistance. Antimicrobial agents and chemotherapy 2009, 53:3914-22.
[206] Kramer LM, Mayes MS, Brown J, Braun L, Fritz-Waters ER, Williams J, Woolums A, Chase C, Reecy JM:
Evaluation of Responses to Vaccination of Angus Cattle for Four Viruses that Contribute to Bovine Respiratory
Disease Complex. Animal Industry Report 2017, 663:7.
[207] Fulton R, d’Offay J, Landis C, Miles D, Smith R, Saliki J, Ridpath J, Confer A, Neill J, Eberle R: Detection
and characterization of viruses as field and vaccine strains in feedlot cattle with bovine respiratory disease.
Vaccine 2016, 34:3478-92.
[208] Biswas A, Shivachandra S, Saxena M, Kumar A, Singh V, Srivastava S: Molecular variability among
strains of Pasteurella multocida isolated from an outbreak of haemorrhagic septicaemia in India. Veterinary
research communications 2004, 28:287-98.
[209] Kumar A, Shivachandra S, Biswas A, Singh V, Singh VP, Srivastava S: Prevalent serotypes of Pasteurella
multocida isolated from different animal and avian species in India. Veterinary research communications 2004,
28:657-67.
[210] Kumar P, Singh V, Agrawal R, Singh S: Identification of Pasteurella multocida isolates of ruminant origin
using polymerase chain reaction and their antibiogram study. Tropical animal health and production 2009,
41:573-8.
[211] Shivachandra S, Viswas K, Kumar A: A review of hemorrhagic septicemia in cattle and buffalo. Animal
Health Research Reviews 2011, 12:67-82.
[212] Perry BD: Investing in animal health research to alleviate poverty: ILRI (aka ILCA and ILRAD), 2002.
[213] Aalbæk B, Eriksen L, Rimler RB, Leifsson PS, Basse A, Christiansen T, Eriksen E: Typing of Pasteurella
multocida from haemorrhagic septicaemia in Danish fallow deer (Dama dama). Apmis 1999, 107:913-20.
[214] Benkirane A, De Alwis M: Haemorrhagic septicaemia, its significance, prevention and control in Asia.
VETERINARNI MEDICINA-PRAHA- 2002, 47:234-40.
[215] Thacker EL: Immunology of the porcine respiratory disease complex. Veterinary Clinics of North America:
Food Animal Practice 2001, 17:551-65.
[216] Brockmeier SL, Halbur PG, Thacker EL: Porcine respiratory disease complex. 2002.
[217] Thacker EL, Thanawongnuwech R: Porcine respiratory disease complex. Thai J Vet Med 2002, 32.
[218] Elling F, Pedersen K: The pathogenesis of persistent turbinate atrophy induced by toxigenic Pasteurella
multocida in pigs. Veterinary pathology 1985, 22:469-74.
[219] Lax AJ, Chanter N: Cloning of the toxin gene from Pasteurella multocida and its role in atrophic rhinitis.
Microbiology 1990, 136:81-7.
[220] Dominick M, Rimler R: Turbinate atrophy in gnotobiotic pigs intranasally inoculated with protein toxin
isolated from type D Pasteurella multocida. American journal of veterinary research 1986, 47:1532-6.

[221] Kimman T, Löwik C, Van De Wee-Pals L, Thesingh C, Defize P, Kamp E, Bijvoet O: Stimulation of bone
resorption by inflamed nasal mucosa, dermonecrotic toxin-containing conditioned medium from Pasteurella
multocida, and purified dermonecrotic toxin from P. multocida. Infection and immunity 1987, 55:2110-6.
[222] Horiguchi Y, Senda T, Sugimoto N, Katahira J, Matsuda M: Bordetella bronchiseptica dermonecrotizing
toxin stimulates assembly of actin stress fibers and focal adhesions by modifying the small GTP-binding protein
rho. Journal of Cell Science 1995, 108:3243-51.
[223] Harris D, Switzer W: Turbinate atrophy in young pigs exposed to Bordetella bronchiseptica, Pasteurella
multocida, and combined inoculum. American journal of veterinary research 1968, 29:777-85.
[224] De Jong M, Nielsen J: Definition of progressive atrophic rhinitis. Veterinary Record 1990, 126.
[225] Pedersen K, Elling F: The pathogenesis of atrophic rhinitis in pigs induced by toxigenic Pasteurella
multocida. Journal of comparative pathology 1984, 94:203-14.
[226] Pedersen K, Barfod K: The aetiological significance of Bordetella bronchiseptica and Pasteurella multocida
in atrophic rhinitis of swine. Nordisk veterinaermedicin 1981, 33:513.
[227] Liao C-M, Huang C, Hsuan S-L, Chen Z-W, Lee W-C, Liu C-I, Winton JR, Chien M-S: Immunogenicity
and efficacy of three recombinant subunit Pasteurella multocida toxin vaccines against progressive atrophic
rhinitis in pigs. Vaccine 2006, 24:27-35.
[228] Foged N, Nielsen J, Jorsal S: Protection against progressive atrophic rhinitis by vaccination with Pasteurella
multocida toxin purified by monoclonal antibodies. The Veterinary Record 1989, 125:7-11.
[229] DiGiacomo R, Jones C, Wathes C: Transmission of Pasteurella multocida in rabbits. Laboratory animal
science 1987, 37:621.
[230] Rougier S, Galland D, Boucher S, Boussarie D, Vallé M: Epidemiology and susceptibility of pathogenic
bacteria responsible for upper respiratory tract infections in pet rabbits. Veterinary microbiology 2006, 115:192-8.
[231] Duclos P, Caillet J, Javelot P: Flore bactérienne aérobie des cavités nasales du lapin d’élevage. Ann Rech
Vét 1986, 17:91-9.
[232] George JL, Martin DJ, Lukacs PM, Miller MW: Epidemic pasteurellosis in a bighorn sheep population
coinciding with the appearance of a domestic sheep. Journal of Wildlife Diseases 2008, 44:388-403.
[233] Weiser GC, DeLong WJ, Paz JL, Shafii B, Price WJ, Ward AC: Characterization of Pasteurella multocida
associated with pneumonia in bighorn sheep. Journal of Wildlife Diseases 2003, 39:536-44.
[234] Callan R, Bunch T, Workman G, Mock R: Development of pneumonia in desert bighorn sheep after
exposure to a flock of exotic wild and domestic sheep. Journal of the American Veterinary Medical Association
1991, 198:1052-6.
[235] Franson JC, Smith BL: Septicemic pasteurellosis in elk (Cervus elaphus) on the United States National Elk
Refuge, Wyoming. Journal of Wildlife Diseases 1988, 24:715-7.
[236] Brogden KA, Rhoades KR: Prevalence of serologic types of Pasteurella multocida from 57 species of birds
and mammals in the United States. Journal of Wildlife Diseases 1983, 19:315-20.
[237] Tessaro SV: Review of the diseases, parasites and miscellaneous pathological conditions of North American
bison. The Canadian Veterinary Journal 1989, 30:416.
[238] Hernandez J, Prado V, Torres D, Waldenström J, Haemig PD, Olsen B: Enteropathogenic Escherichia coli
(EPEC) in Antarctic fur seals Arctocephalus gazella. Polar Biology 2007, 30:1227-9.
[239] Kennedy-Stoskopf S, Stoskopf MK, Eckhaus MA, Strandberg JD: Isolation of a retrovirus and a
herpesvirus from a captive California sea lion. Journal of Wildlife Diseases 1986, 22:156-64.
[240] Bjarnsholt T: The role of bacterial biofilms in chronic infections. APMIS 2013, 121:1-58.
[241] James GA, Swogger E, Wolcott R, Pulcini Ed, Secor P, Sestrich J, Costerton JW, Stewart PS: Biofilms in
chronic wounds. Wound Repair and Regeneration 2008, 16:37-44.
[242] Sandal I, Inzana TJ, Molinaro A, De Castro C, Shao JQ, Apicella MA, Cox AD, St Michael F, Berg G:
Identification, structure, and characterization of an exopolysaccharide produced by Histophilus somni during
biofilm formation. BMC Microbiol 2011, 11:186.
[243] Corbeil LB: Histophilus somni host-parasite relationships. Animal health research reviews / Conference of
Research Workers in Animal Diseases 2007, 8:151-60.

[244] Elswaifi SF SW, Inzana TJ: The role of lipooligosaccharide phosphorylcholine in colonization and
pathogenesis of Histophilus somni in cattle. Vet Res 2012, 43:49. doi: 10.1186/297-9716-43-49.
[245] Gogolewski RP, Leathers CW, Liggitt HD, Corbeil LB: Experimental Haemophilus somnus pneumonia in
calves and immunoperoxidase localization of bacteria. Vet Pathol 1987, 24:250-6.
[246] Agnes JT, Zekarias B, Shao M, Anderson ML, Gershwin LJ, Corbeil LB: Bovine respiratory syncytial virus
and Histophilus somni interaction at the alveolar barrier. Infection and immunity 2013, 81:2592-7.
[247] Gershwin LJ: Bovine respiratory syncytial virus infection: immunopathogenic mechanisms. Animal Health
Research Reviews 2007, 8:207-13.
[248] Gershwin LJ, Berghaus LJ, Arnold K, Anderson ML, Corbeil LB: Immune mechanisms of pathogenetic
synergy in concurrent bovine pulmonary infection with Haemophilus somnus and bovine respiratory syncytial
virus. Veterinary immunology and immunopathology 2005, 107:119-30.
[249] Nyvad B, Crielaard W, Mira A, Takahashi N, Beighton D: Dental caries from a molecular microbiological
perspective. Caries research 2013, 47:89-102.
[250] Marsh PD: Dental plaque as a biofilm and a microbial community–implications for health and disease.
BMC Oral health 2006, 6:S14.
[251] Johansen LK, Koch J, Frees D, Aalbæk B, Nielsen OL, Leifsson PS, Iburg TM, Svalastoga E, Buelund LE,
Bjarnsholt T, Høiby N, Jensen HE: Pathology and Biofilm Formation in a Porcine Model of Staphylococcal
Osteomyelitis. Journal of Comparative Pathology 2012, 147:343-53.
[252] Hussain M, Steinbacher T, Peters G, Heilmann C, Becker K: The adhesive properties of the Staphylococcus
lugdunensis multifunctional autolysin AtlL and its role in biofilm formation and internalization. International
journal of medical microbiology : IJMM 2015, 305:129-39.
[253] Nguyen CT, Jung W, Kim J, Chaney EJ, Novak M, Stewart CN, Boppart SA: Noninvasive in vivo optical
detection of biofilm in the human middle ear. Proceedings of the National Academy of Sciences of the United
States of America 2012, 109:9529-34.
[254] Bakaletz LO: Bacterial biofilms in the upper airway-evidence for role in pathology and implications for
treatment of otitis media. Paediatric respiratory reviews 2012, 13:154-9.
[255] Puig C, Domenech A, Garmendia J, Langereis JD, Mayer P, Calatayud L, Liñares J, Ardanuy C, Marti S:
Increased Biofilm Formation by Nontypeable Haemophilus influenzae Isolates from Patients with Invasive
Disease or Otitis Media versus Strains Recovered from Cases of Respiratory Infections. Applied and
environmental microbiology 2014, 80:7088-95.
[256] Francoz D, Buczinski S, Bélanger AM, Forté G, Labrecque O, Tremblay D, Wellemans V, Dubuc J:
Respiratory Pathogens in Québec Dairy Calves and Their Relationship with Clinical Status, Lung Consolidation,
and Average Daily Gain. Journal of Veterinary Internal Medicine 2015, 29:381-7.
[257] Griffin D: The monster we don't see: subclinical BRD in beef cattle. Animal Health Research Reviews
2014, 15:138-41.
[258] Sandal I, Inzana TJ, Molinaro A, De Castro C, Shao JQ, Apicella MA, Cox AD, St Michael F, Berg G:
Identification, structure, and characterization of an exopolysaccharide produced by Histophilus somni during
biofilm formation. BMC microbiology 2011, 11:186.
[259] Sandal I, Hong W, Swords WE, Inzana TJ: Characterization and comparison of biofilm development by
pathogenic and commensal isolates of Histophilus somni. J Bacteriol 2007, 189:8179-85.
[260] Villarino Romero R, Osicka R, Sebo P: Filamentous hemagglutinin of Bordetella pertussis: a key adhesin
with immunomodulatory properties? Future Microbiol 2014, 9:1339-60.
[261] Ward CK, Lumbley SR, Latimer JL, Cope LD, Hansen EJ: Haemophilus ducreyi secretes a filamentous
hemagglutinin-like protein. J Bacteriol 1998, 180:6013-22.
[262] Gotschlich EC, Fraser BA, Nishimura O, Robbins JB, Liu TY: Lipid on capsular polysaccharides of gram-
negative bacteria. J Biol Chem 1981, 256:8915-21.
[263] Zarankiewicz T, Madej J, Galli J, Bajzert J, Stefaniak T: Inhibition of in vitro Histophilus somni biofilm
production by recombinant Hsp60 antibodies. Polish journal of veterinary sciences 2012, 15:373-8.
[264] Henderson B, Fares MA, Lund PA: Chaperonin 60: a paradoxical, evolutionarily conserved protein family
with multiple moonlighting functions. Biological Reviews 2013, 88:955-87.

[265] Zou Y, Feng S, Xu C, Zhang B, Zhou S, Zhang L, He X, Li J, Yang Z, Liao M: The role of galU and galE
of Haemophilus parasuis SC096 in serum resistance and biofilm formation. Veterinary microbiology 2013,
162:278-84.
[266] LiGuo S, YuHua Z, Pai G, YongLiang Y: Research progress on the function and regulatory mechanism of
CsrA, a global regulator in bacteria. Journal of Agricultural Science and Technology (Beijing) 2014, 16:79-86.
[267] Mandrell RE, McLaughlin R, Kwaik YA, Lesse A, Yamasaki R, Gibson B, Spinola SM, Apicella MA:
Lipooligosaccharides (LOS) of some Haemophilus species mimic human glycosphingolipids, and some LOS are
sialylated. Infect Immun 1992, 60:1322-8.
[268] Howard MD, Willis L, Wakarchuk W, St. Michael F, Cox A, Horne WT, Hontecillas R, Bassaganya-Riera
J, Lorenz E, Inzana TJ: Genetics and molecular specificity of sialylation of Histophilus somni lipooligosaccharide
(LOS) and the effect of LOS sialylation on Toll-like receptor-4 signaling. Veterinary microbiology 2011,
153:163-72.
[269] Inzana TJ, Balyan R, Howard MD: Decoration of Histophilus somni lipooligosaccharide with N-acetyl-5-
neuraminic acid enhances bacterial binding of complement factor H and resistance to killing by serum and
polymorphonuclear leukocytes. Veterinary microbiology 2012, 161:113-21.
[270] Inzana TJ, Glindemann G, Cox AD, Wakarchuk W, Howard MD: Incorporation of N-acetylneuraminic acid
into Haemophilus somnus lipooligosaccharide (LOS): enhancement of resistance to serum and reduction of LOS
antibody binding. Infect Immun 2002, 70:4870-9.
[271] Swords WE, Moore ML, Godzicki L, Bukofzer G, Mitten MJ, VonCannon J: Sialylation of
lipooligosaccharides promotes biofilm formation by nontypeable Haemophilus influenzae. Infection and
immunity 2004, 72:106-13.
[272] Langereis JD, Hermans PW: Novel concepts in nontypeable Haemophilus influenzae biofilm formation.
FEMS microbiology letters 2013, 346:81-9.
[273] Pan Y, Fisher T, Olk C, Inzana TJ: Detection of antibodies to the biofilm exopolysaccharide of Histophilus
somni following infection in cattle by enzyme-linked immunosorbent assay. Clinical and vaccine immunology :
CVI 2014, 21:1463-7.
[274] Miyoshi S, Hamada H, Miyoshi A, Ito R, Hamaguchi N, Murakami S, Miyamoto H, Takeuchi T, Okura T,
Higaki J: Pasteurella multocida pneumonia: Zoonotic transmission confirmed by molecular epidemiological
analysis. Geriatrics & gerontology international 2012, 12:159-63.
[275] Ewers C, Lübke-Becker A, Bethe A, Kießling S, Filter M, Wieler LH: Virulence genotype of Pasteurella
multocida strains isolated from different hosts with various disease status. Veterinary microbiology 2006,
114:304-17.
[276] Giordano A, Dincman T, Clyburn BE, Steed LL, Rockey DC: Clinical Features and Outcomes of
Pasteurella multocida Infection. Medicine 2015, 94.
[277] Patel S, Joshi D, Raval S, Patel B, Patel J, Chauhan H, Chandel B, Patel B, Shah N: Clinicopathological
studies of Pasteurella multocida B: 2 experimental infection in rabbits. Indian Journal of Animal Sciences 2016,
86:380-6.
[278] Hodgson J, Dagleish M, Gibbard L, Bayne C, Finlayson J, Moon G, Nath M: Seven strains of mice as
potential models of bovine pasteurellosis following intranasal challenge with a bovine pneumonic strain of
Pasteurella multocida A: 3; comparisons of disease and pathological outcomes. Research in veterinary science
2013, 94:634-40.
[279] Samuel MD, Goldberg DR, Shadduck DJ, Price JI, Cooch EG: Pasteurella multocida serotype 1 isolated
from a lesser snow goose: evidence of a carrier state. Journal of wildlife diseases 1997, 33:332-5.
[280] Samuel MD, Shadduck DJ, Goldberg DR: Avian cholera exposure and carriers in greater white-fronted
geese breeding in Alaska, USA. Journal of wildlife diseases 2005, 41:498-502.
[281] Samuel MD, Shadduck DJ, Goldberg DR, Johnson WP: Comparison of methods to detect Pasteurella
multocida in carrier waterfowl. J Wildl Dis 2003, 39:125-35.
[282] Chung JY, Wilkie I, Boyce JD, Townsend KM, Frost AJ, Ghoddusi M, Adler B: Role of capsule in the
pathogenesis of fowl cholera caused by Pasteurella multocida serogroup A. Infection and immunity 2001,
69:2487-92.

[283] Carter G: Pasteurellosis: Pasteurella multocida and Pasteurella hemolytica. Advances in veterinary science
1966, 11:321-79.
[284] Rimler R: Presumptive identification of Pasteurella multocida serogroups A, D and F by capsule
depolymerisation with mucopolysaccharidases. Veterinary record 1994, 134:191-2.
[285] Dargatz DA, Lombard JE: Summary of BRD data from the 2011 NAHMS feedlot and dairy heifer studies.
Animal Health Research Reviews 2014, 15:123-5.
[286] Griego RD, Rosen T, Orengo IF, Wolf JE: Dog, cat, and human bites: a review. Journal of the American
Academy of Dermatology 1995, 33:1019-29.
[287] Talan DA, Citron DM, Abrahamian FM, Moran GJ, Goldstein EJ: Bacteriologic analysis of infected dog
and cat bites. Emergency Medicine Animal Bite Infection Study Group. The New England journal of medicine
1999, 340:85-92.
[288] Narsana N, Farhat F: Septic shock due to Pasteurella multocida bacteremia: a case report. J Med Case Rep
2015, 9:159.
[289] Carter GR, Chengappa MM: Recommendations for a standard system of designating serotypes of
Pasteurella multocida. Proceedings 24th Amer Assoc Vet Lab Diagn 1981. pp. 37-42.
[290] Etienne G, Malaga W, Laval F, Lemassu A, Guilhot C, Daffé M: Identification of the polyketide synthase
involved in the biosynthesis of the surface-exposed lipooligosaccharides in mycobacteria. Journal of bacteriology
2009, 191:2613-21.
[291] Harmon B, Glisson J, Latimer K, Steffens W, Nunnally J: Resistance of Pasteurella multocida A: 3, 4 to
phagocytosis by turkey macrophages and heterophils. American journal of veterinary research 1991, 52:1507-11.
[292] Poermadjaja B, Frost A: Phagocytic uptake and killing of virulent and avirulent strains of Pasteurella
multocida of capsular serotype A by chicken macrophages. Veterinary microbiology 2000, 72:163-71.
[293] McNally DJ, Schoenhofen IC, Mulrooney EF, Whitfield DM, Vinogradov E, Lam JS, Logan SM, Brisson
JR: Identification of labile UDP-ketosugars in Helicobacter pylori, Campylobacter jejuni and Pseudomonas
aeruginosa: key metabolites used to make glycan virulence factors. Chembiochem 2006, 7:1865-8.
[294] Sandkvist M, Hough LP, Bagdasarian MM, Bagdasarian M: Direct interaction of the EpsL and EpsM
proteins of the general secretion apparatus in Vibrio cholerae. Journal of bacteriology 1999, 181:3129-35.
[295] Isobe T, White KA, Allen AG, Peacock M, Raetz CR, Maskell DJ: Bordetella pertussis waaA encodes a
monofunctional 2-keto-3-deoxy-D-manno-octulosonic acid transferase that can complement an Escherichia coli
waaA mutation. Journal of bacteriology 1999, 181:2648-51.
[296] Kingsley MT, Gabriel DW, Marlow GC, Roberts PD: The opsX locus of Xanthomonas campestris affects
host range and biosynthesis of lipopolysaccharide and extracellular polysaccharide. Journal of bacteriology 1993,
175:5839-50.
[297] Petersen KD, Christensen JP, Permin A, Bisgaard M: Virulence of Pasteurella multocida subsp. multocida
isolated from outbreaks of fowl cholera in wild birds for domestic poultry and game birds. Avian Pathol 2001,
30:27-31.
[298] Wobeser G: Avian cholera and waterfowl biology. Journal of Wildlife Diseases 1992, 28:674-82.
[299] Pritchett IW, Beaudette FR, Hughes TP: The Epidemiology of Fowl Cholera : V. Further Field
Observations of the Spontaneous Disease. J Exp Med 1930, 51:259-74.
[300] ZINKL JG, Dey N, HYLAND JM, HURT JJ, HEDDLESTON KL: An epornitic of avian cholera in
waterfowl and common crows in Phelps County, Nebraska, in the spring, 1975. Journal of Wildlife Diseases
1977, 13:194-8.
[301] Bond RE, Donahue JM, Olson LD: Colony features of Pasteurella multocida and their use in diagnosing
fowl cholera in turkeys. Avian diseases 1970, 14:24-8.
[302] Heddleston KL, Watko LP, Rebers PA: Dissociation of a fowl
cholera strain of Pasteurella multocida. Avian Dis 1964, 8:649-57.
[303] Yi K, Rasmussen AW, Gudlavalleti SK, Stephens DS, Stojiljkovic I: Biofilm formation by Neisseria
meningitidis. Infection and immunity 2004, 72:6132-8.

[304] Sherlock O, Schembri MA, Reisner A, Klemm P: Novel roles for the AIDA adhesin from diarrheagenic
Escherichia coli: cell aggregation and biofilm formation. Journal of bacteriology 2004, 186:8058-65.
[305] Danese PN, Pratt LA, Dove SL, Kolter R: The outer membrane protein, Antigen 43, mediates cell‐to‐cell
interactions within Escherichia coli biofilms. Molecular microbiology 2000, 37:424-32.
[306] Felek S, Lawrenz MB, Krukonis ES: The Yersinia pestis autotransporter YapC mediates host cell binding,
autoaggregation and biofilm formation. Microbiology 2008, 154:1802-12.
[307] Rosner H, Grimmecke HD, Knirel YA, Shashkov AS: Hyaluronic acid and a (1----4)-beta-D-xylan,
extracellular polysaccharides of Pasteurella multocida (Carter type A) strain 880. Carbohydr Res 1992, 223:329-
33.
[308] Song J-M, Im J-H, Kang J-H, Kang D-J: A simple method for hyaluronic acid quantification in culture
broth. Carbohydrate Polymers 2009, 78:633-4.
[309] Chen Y-H, Wang Q: Establishment of CTAB Turbidimetric method to determine hyaluronic acid content in
fermentation broth. Carbohydrate Polymers 2009, 78:178-81.
[310] Bitter T, Muir HM: A modified uronic acid carbazole reaction. Analytical biochemistry 1962, 4:330-4.
[311] Ogawa K, Hatano M: Circular dichroism of the complex of a (1→ 3)-β-d-glucan with Congo Red.
Carbohydrate Research 1978, 67:527-35.
[312] Nagar E, Schwarz R: To be or not to be planktonic? Self‐inhibition of biofilm development. Environmental
microbiology 2015, 17:1477-86.
[313] Schembri MA, Dalsgaard D, Klemm P: Capsule Shields the Function of Short Bacterial Adhesins. Journal
of Bacteriology 2004, 186:1249-57.
[314] Klemm P, Vejborg RM, Sherlock O: Self-associating autotransporters, SAATs: Functional and structural
similarities. International Journal of Medical Microbiology 2006, 296:187-95.
[315] Wilson WA, Roach PJ, Montero M, Baroja-Fernández E, Muñoz FJ, Eydallin G, Viale AM, Pozueta-
Romero J: Regulation of glycogen metabolism in yeast and bacteria. FEMS microbiology reviews 2010, 34:952-
85.
[316] Hart ME, Champlin FR: Susceptibility to hydrophobic molecules and phospholipid composition in
Pasteurella multocida and Actinobacillus lignieresii. Antimicrobial agents and chemotherapy 1988, 32:1354-9.
[317] Doyle RJ: Contribution of the hydrophobic effect to microbial infection. Microbes and infection 2000,
2:391-400.
[318] Herzberg M, Rezene TZ, Ziemba C, Gillor O, Mathee K: Impact of higher alginate expression on deposition
of Pseudomonas aeruginosa in radial stagnation point flow and reverse osmosis systems. Environmental science
& technology 2009, 43:7376-83.
[319] Sherlock O, Vejborg RM, Klemm P: The TibA adhesin/invasin from enterotoxigenic Escherichia coli is self
recognizing and induces bacterial aggregation and biofilm formation. Infection and immunity 2005, 73:1954-63.
[320] Sorroche FG, Spesia MB, Zorreguieta Á, Giordano W: A positive correlation between bacterial
autoaggregation and biofilm formation in native Sinorhizobium meliloti isolates from Argentina. Applied and
environmental microbiology 2012, 78:4092-101.
[321] Jackson DW, Suzuki K, Oakford L, Simecka JW, Hart ME, Romeo T: Biofilm formation and dispersal
under the influence of the global regulator CsrA of Escherichia coli. J Bacteriol 2002, 184:290-301.
[322] Fields JA, Thompson SA: Campylobacter jejuni CsrA mediates oxidative stress responses, biofilm
formation, and host cell invasion. J Bacteriol 2008, 190:3411-6.
[323] Jablonski L, Sriranganathan N, Boyle SM, Carter GR: Conditions for transformation of Pasteurella
multocida by electroporation. Microbial pathogenesis 1992, 12:63-8.
[324] Kamp EM, Bokken GC, Vermeulen TM, de Jong MF, Buys HE, Reek FH, Smits MA: A specific and
sensitive PCR assay suitable for large-scale detection of toxigenic Pasteurella multocida in nasal and tonsillar
swabs specimens of pigs. Journal of Veterinary Diagnostic Investigation 1996, 8:304-9.
[325] Livak KJ, Schmittgen TD: Analysis of relative gene expression data using real-time quantitative PCR and
the 2− ΔΔCT method. methods 2001, 25:402-8.
[326] Johnson M, Zaretskaya I, Raytselis Y, Merezhuk Y, McGinnis S, Madden TL: NCBI BLAST: a better web
interface. Nucleic acids research 2008, 36:W5-W9.

[327] Kumar A, Larsen CE, Preiss J: Biosynthesis of bacterial glycogen. Primary structure of Escherichia coli
ADP-glucose: alpha-1, 4-glucan, 4-glucosyltransferase as deduced from the nucleotide sequence of the glgA gene.
Journal of Biological Chemistry 1986, 261:16256-9.
[328] De Castro C, Parrilli M, Holst O, Molinaro A: Microbe-associated molecular patterns in innate immunity:
Extraction and chemical analysis of gram-negative bacterial lipopolysaccharides. Methods Enzymol 2010,
480:89-115.
[329] Scott Jr TA, Melvin EH: Determination of dextran with anthrone. Analytical Chemistry 1953, 25:1656-61.
[330] Inzana TJ: Electrophoretic heterogeneity and interstrain variation of the lipopolysaccharide of Haemophilus
influenzae. Journal of Infectious Diseases 1983, 148:492-9.
[331] Inzana TJ, Apicella MA: Use of a bilayer stacking gel to improve resolution of lipopolysaccharides and
lipooligosaccharides in polyacrylamide gels. Electrophoresis 1999, 20:462-5.
[332] Goncalves Mdos S, Delattre C, Balestrino D, Charbonnel N, Elboutachfaiti R, Wadouachi A, Badel S,
Bernardi T, Michaud P, Forestier C: Anti-biofilm activity: a function of Klebsiella pneumoniae capsular
polysaccharide. PLoS One 2014, 9:e99995.
[333] Heydorn A, Nielsen AT, Hentzer M, Sternberg C, Givskov M, Ersboll BK, Molin S: Quantification of
biofilm structures by the novel computer program COMSTAT. Microbiology 2000, 146 ( Pt 10):2395-407.
[334] Adler B, Bulach D, Chung J, Doughty S, Hunt M, Rajakumar K, Serrano M, van Zanden A, Zhang Y,
Ruffolo C: Candidate vaccine antigens and genes in Pasteurella multocida. Journal of biotechnology 1999, 73:83-
90.
[335] Willis LM, Whitfield C: Structure, biosynthesis, and function of bacterial capsular polysaccharides
synthesized by ABC transporter-dependent pathways. Carbohydrate research 2013, 378:35-44.
[336] Clutterbuck A, Woods E, Knottenbelt D, Clegg P, Cochrane C, Percival S: Biofilms and their relevance to
veterinary medicine. Veterinary microbiology 2007, 121:1-17.
[337] Jacques M, Aragon V, Tremblay YD: Biofilm formation in bacterial pathogens of veterinary importance.
Animal Health Research Reviews 2010, 11:97-121.
[338] Adelberg EA, Mandel M, Chen GCC: Optimal conditions for mutagenesis by N-methyl-N′-nitro-N-
nitrosoguanidine in Escherichia coli K12. Biochemical and Biophysical Research Communications 1965, 18:788-
95.
[339] Tae H, Settlage RE, Shallom S, Bavarva JH, Preston D, Hawkins GN, Adams LG, Garner HR: Improved
variation calling via an iterative backbone remapping and local assembly method for bacterial genomes.
Genomics 2012, 100:271-6.
[340] Darling AE, Mau B, Perna NT: progressiveMauve: multiple genome alignment with gene gain, loss and
rearrangement. PloS one 2010, 5:e11147.
[341] Smith RL, Gilkerson E: Quantitation of glycosaminoglycan hexosamine using 3-methyl-2-benzothiazolone
hydrazone hydrochloride. Analytical biochemistry 1979, 98:478-80.
[342] Zhang Y, Luo K, Zhao Q, Qi Z, Nielsen LK, Liu H: Genetic and biochemical characterization of genes
involved in hyaluronic acid synthesis in Streptococcus zooepidemicus. Applied microbiology and biotechnology
2016, 100:3611-20.
[343] Hossain GS, Shin H-d, Li J, Wang M, Du G, Chen J, Liu L: Metabolic engineering for amino-, oligo-, and
polysugar production in microbes. Applied microbiology and biotechnology 2016, 100:2523-33.
[344] Olsen LR, Roderick SL: Structure of the Escherichia coli GlmU pyrophosphorylase and acetyltransferase
active sites. Biochemistry 2001, 40:1913-21.
[345] Gehring AM, Lees WJ, Mindiola DJ, Walsh CT, Brown ED: Acetyltransfer precedes uridylyltransfer in the
formation of UDP-N-acetylglucosamine in separable active sites of the bifunctional GlmU protein of Escherichia
coli. Biochemistry 1996, 35:579-85.
[346] Zhang Z, Bulloch EM, Bunker RD, Baker EN, Squire CJ: Structure and function of GlmU from
Mycobacterium tuberculosis. Acta Crystallographica Section D: Biological Crystallography 2009, 65:275-83.
[347] Mochalkin I, Lightle S, Narasimhan L, Bornemeier D, Melnick M, VanderRoest S, McDowell L: Structure
of a small‐molecule inhibitor complexed with GlmU from Haemophilus influenzae reveals an allosteric binding
site. Protein Science 2008, 17:577-82.

[348] Kostrewa D, D'Arcy A, Takacs B, Kamber M: Crystal Structures of Streptococcus pneumoniaeN-
Acetylglucosamine-1-phosphate Uridyltransferase, GlmU, in Apo Form at 2.33 Å Resolution and in Complex
with UDP-N-Acetylglucosamine and Mg 2+ at 1.96 Å Resolution. Journal of molecular biology 2001, 305:279-
89.
[349] Taylor JD, Fulton RW, Lehenbauer TW, Step DL, Confer AW: The epidemiology of bovine respiratory
disease: what is the evidence for preventive measures. Can Vet J 2010, 51:1351-9.
[350] Perez AC, Pang B, King LB, Tan L, Murrah KA, Reimche JL, Wren JT, Richardson SH, Ghandi U, Swords
WE: Residence of Streptococcus pneumoniae and Moraxella catarrhalis within polymicrobial biofilm promotes
antibiotic resistance and bacterial persistence in vivo. Pathogens and disease 2014, 70:280-8.
[351] Fulton RW: Bovine respiratory disease research (1983–2009). Animal Health Research Reviews 2009,
10:131-9.
[352] Thurnheer T, Gmür R, Guggenheim B: Multiplex FISH analysis of a six-species bacterial biofilm. Journal
of Microbiological Methods 2004, 56:37-47.
[353] Hennigar R, Mayfield R, Harvey J, Ge Z-H, Sens D: Lectin detection of renal glycogen in rats with short-
term streptozotocin-diabetes. Diabetologia 1987, 30:804-11.
[354] Hennigar RA, Schulte BA, Spicer SS: Histochemical detection of glycogen using Griffonia simplicifolia
agglutinin II. The Histochemical journal 1986, 18:589-96.
[355] Angen Ø, Ahrens P, Tegtmeier C: Development of a PCR test for identification of Haemophilus somnus in
pure and mixed cultures. Veterinary microbiology 1998, 63:39-48.
[356] Yemm E, Willis A: The estimation of carbohydrates in plant extracts by anthrone. Biochemical journal
1954, 57:508.
[357] Laurentin A, Edwards CA: A microtiter modification of the anthrone-sulfuric acid colorimetric assay for
glucose-based carbohydrates. Analytical Biochemistry 2003, 315:143-5.
[358] Heydorn A, Nielsen AT, Hentzer M, Sternberg C, Givskov M, Ersbøll BK, Molin S: Quantification of
biofilm structures by the novel computer program COMSTAT. Microbiology (Reading, England) 2000,
146:2395-407.
[359] Zhu Y, Dashper SG, Chen Y-Y, Crawford S, Slakeski N, Reynolds EC: Porphyromonas gingivalis and
Treponema denticola synergistic polymicrobial biofilm development. PloS one 2013, 8:e71727.
[360] Zainal-Abidin Z, Veith PD, Dashper SG, Zhu Y, Catmull DV, Chen Y-Y, Heryanto DC, Chen D, Pyke JS,
Tan K: Differential proteomic analysis of a polymicrobial biofilm. J Proteome Res 2012, 11:4449-64.
[361] Arenas J, Cano S, Nijland R, Dongen V, Rutten L, Ende A, Tommassen J: The meningococcal
autotransporter AutA is implicated in autoaggregation and biofilm formation. Environ Microbiol 2015, 17:1321-
37.
[362] Bos R, van der Mei HC, Meinders JM, Busscher HJ: A quantitative method to study co-adhesion of
microorganisms in a parallel plate flow chamber: basic principles of the analysis. Journal of microbiological
methods 1994, 20:289-305.
[363] Rickard AH, Gilbert P, High NJ, Kolenbrander PE, Handley PS: Bacterial coaggregation: an integral
process in the development of multi-species biofilms. Trends in Microbiology 2003, 11:94-100.
[364] Karched M, Bhardwaj RG, Asikainen SE: Coaggregation and biofilm growth of Granulicatella spp. with
Fusobacterium nucleatum and Aggregatibacter actinomycetemcomitans. BMC microbiology 2015, 15:114.
[365] Castonguay M-H, Van der Schaaf S, Koester W, Krooneman J, Van der Meer W, Harmsen H, Landini P:
Biofilm formation by Escherichia coli is stimulated by synergistic interactions and co-adhesion mechanisms with
adherence-proficient bacteria. Research in Microbiology 2006, 157:471-8.
[366] Pan Y, Fisher T, Olk C, Inzana TJ: Detection of antibodies to the biofilm exopolysaccharide of Histophilus
somni following infection in cattle by enzyme-linked immunosorbent assay. Clinical and Vaccine Immunology
2014, 21:1463-7.
[367] Okshevsky M, Meyer RL: The role of extracellular DNA in the establishment, maintenance and
perpetuation of bacterial biofilms. Critical reviews in microbiology 2015, 41:341-52.
[368] Peters BM, Jabra-Rizk MA, Graeme A, Costerton JW, Shirtliff ME: Polymicrobial interactions: impact on
pathogenesis and human disease. Clinical microbiology reviews 2012, 25:193-213.

[369] Brogden KA, Guthmiller JM, Taylor CE: Human polymicrobial infections. The Lancet 2005, 365:253-5.
[370] Hunter B, Wobeser G: Pathology of experimental avian cholera in mallard ducks. Avian Dis 1980:403-14.
[371] Petruzzi B, Brigg RE, Swords WE, De Castro C, Molinaro A, Inzana TJ: Capsular Polysaccharide Interferes
with Biofilm Formation by Pasteurella multocida Serogroup A. mBio 2017:8:e01843-17.
[372] Flemming H-C, Wingender J: The biofilm matrix. Nature Reviews Microbiology 2010, 8:623-33.
[373] Pors SE, Chadfield MS, Sørensen DB, Offenberg H, Bisgaard M, Jensen HE: The origin of Pasteurella
multocida impacts pathology and inflammation when assessed in a mouse model. Research in veterinary science
2016, 105:139-42.
[374] Inzana T, Gogolewski R, Corbeil L: Phenotypic phase variation in Haemophilus somnus
lipooligosaccharide during bovine pneumonia and after in vitro passage. Infection and immunity 1992, 60:2943-
51.
[375] Olson L, McCune E, Moseley B: Gross and histopathological description of the cranial form of chronic
fowl cholera in turkeys. Avian Dis 1966, 10:518-29.
[376] Boyd CD, O'Toole GA: Second messenger regulation of biofilm formation: breakthroughs in understanding
c-di-GMP effector systems. Annual review of cell and developmental biology 2012, 28:439-62.
[377] Bojesen AM, Petersen KD, Nielsen OL, Christensen JP, Bisgaard M: Pasteurella multocida infection in
heterophil-depleted chickens. Avian Dis 2004, 48:463-70.
[378] Kodama H, Matsumoto M, Snow LM: Immunogenicity of capsular antigens of Pasteurella multocida in
turkeys. American journal of veterinary research 1981, 42:1838-41.
[379] Wang Y, Jiang B, Guo Y, Li W, Tian Y, Sonnenberg GF, Weiser JN, Ni X, Shen H: Cross-protective
mucosal immunity mediated by memory Th17 cells against Streptococcus pneumoniae lung infection. Mucosal
immunology 2017, 10:250-9.
[380] Lillehoj HS, Min W, Choi KD, Babu US, Burnside J, Miyamoto T, Rosenthal BM, Lillehoj EP: Molecular,
cellular, and functional characterization of chicken cytokines homologous to mammalian IL-15 and IL-2.
Veterinary immunology and immunopathology 2001, 82:229-44.
[381] Sun K, Salmon SL, Lotz SA, Metzger DW: Interleukin-12 promotes gamma interferon-dependent
neutrophil recruitment in the lung and improves protection against respiratory Streptococcus pneumoniae
infection. Infection and immunity 2007, 75:1196-202.
[382] Qureshi M: Avian macrophage and immune response: an overview. Poultry science 2003, 82:691-8.
[383] Withanage G, Wigley P, Kaiser P, Mastroeni P, Brooks H, Powers C, Beal R, Barrow P, Maskell D,
McConnell I: Cytokine and chemokine responses associated with clearance of a primary Salmonella enterica
serovar Typhimurium infection in the chicken and in protective immunity to rechallenge. Infection and immunity
2005, 73:5173-82.
[384] Caucheteux SM, Hu-Li J, Mohammed R, Ager A, Paul WE: Cytokine regulation of lung Th17 response to
airway immunization using LPS adjuvant. Mucosal immunology 2017, 10:361-72.
[385] Codolo G, Amedei A, Steere AC, Papinutto E, Cappon A, Polenghi A, Benagiano M, Paccani SR, Sambri
V, Del Prete G: Borrelia burgdorferi NapA–driven Th17 cell inflammation in Lyme arthritis. Arthritis &
Rheumatology 2008, 58:3609-17.
[386] Weining KC, Sick C, Kaspers B, Staeheli P: A chicken homolog of mammalian interleukin‐1β: cDNA
cloning and purification of active recombinant protein. The FEBS Journal 1998, 258:994-1000.
[387] Wu Z, Hu T, Kaiser P: Chicken CCR6 and CCR7 are markers for immature and mature dendritic cells
respectively. Developmental & Comparative Immunology 2011, 35:563-7.
[388] Kim WH, Jeong J, Park AR, Yim D, Kim S, Chang HH, Yang S-H, Kim D-H, Lillehoj HS, Min W:
Downregulation of chicken interleukin-17 receptor A during Eimeria infection. Infection and immunity 2014,
82:3845-54.
[389] Wu C, Qin X, Li P, Pan T, Ren W, Li N, Peng Y: Transcriptomic Analysis on Responses of Murine Lungs
to Pasteurella multocida Infection. Frontiers in cellular and infection microbiology 2017, 7.