oxidation of nitrotoluenes by toluene dioxygenase - applied and
TRANSCRIPT

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Aug. 1992, p. 2643-26480099-2240/92/082643-06$02.00/0Copyright C) 1992, American Society for Microbiology
Oxidation of Nitrotoluenes by Toluene Dioxygenase:Evidence for a Monooxygenase Reaction
Vol. 58, No. 8
JEFFREY B. ROBERTSON,' JIM C. SPAIN,`* JOHN D. HADDOCK,2 AND DAVID T. GIBSON2
Air Force Civil Engineering Support Agency, Tyndall Air Force Base, Florida 32403,1 andDepartment of Microbiology and Center for Biocatalysis and Bioprocessing,
The University of Iowa, Iowa City, Iowa 522422
Received 4 February 1992/Accepted 1 June 1992
Pseudomonas putida Fl and Pseudomonas sp. strain JS150 initiate toluene degradation by incorporatingmolecular oxygen into the aromatic nucleus to form cis-1,2-dihydroxy-3-methylcyclohexa-3,5-diene. Whentoluene-grown cells were incubated with 2- and 3-nitrotoluene, the major products identified were 2- and3-nitrobenzyl alcohol, respectively. The same cells oxidized 4-nitrotoluene to 2-methyl-5-nitrophenol and3-methyl-6-nitrocatechol. Escherichia coli JM109(pDTG601), which contains the toluene dioxygenase genesfrom P. putida Fl under the control of the tac promoter, oxidized the isomeric nitrotoluenes to the samemetabolites as those formed by P. putida Fl and Pseudomonas sp. strain JS150. These results extend the rangeof substrates known to be oxidized by this versatile enzyme and demonstrate for the first time that toluenedioxygenase can oxidize an aromatic methyl substituent.
Nitrotoluenes are used extensively in the manufacture ofazo and sulfur dyes and in the production of explosives (13).They have been detected in wastewater from paper mills andchemical plants, and it is estimated that the levels of 2- and4-nitrotoluene in wastewater from 2,4,6-trinitrotoluene man-ufacturing facilities may be as high as 16 and 9.2 ppm,respectively (24). Little is known about the environmentalfate and biodegradation of these compounds.
It has been suggested that aromatic hydrocarbons withnitro substituents are resistant to electrophilic attack byoxygenases. This is attributed to the electron-withdrawingproperties of nitro groups on the aromatic nucleus (3). Incontrast, polar nitroaromatic compounds such as nitrophe-nols (3, 20, 21, 26) and nitrobenzoic acids (4, 5) are readilybiodegradable. Several studies have shown that nitrotolu-enes can be transformed to aromatic amines by bacteria (7,13, 16), and a very recent report has shown that the methylgroup of m- and p-nitrotoluene can be oxidized by thetoluene monooxygenase encoded by the TOL plasmid (6).However, there is little information available on the struc-ture and physiological properties of metabolites that areformed by the bacterial oxidation of nitrotoluenes.The first intermediate in the degradation of toluene by
Pseudomonas putida Fl and Pseudomonas sp. strain JS150is cis-1,2-dihydroxy-3-methylcyclohexa-3,5-diene (cis-tolu-ene dihydrodiol) (8, 10). The enzyme catalyzing this reac-tion, toluene dioxygenase, has a relaxed substrate specificityand is capable of oxidizing a wide range of aromatic com-pounds (11).We now report novel reactions in the oxidation of isomeric
nitrotoluenes by the oxygenases in P. putida Fl and Pseu-domonas sp. strain JS150 and by the cloned P. putida Fltoluene dioxygenase in Escherichia coli JM109.
MATERIALS AND METHODSMaterials. The following were obtained from the sources
indicated: toluene, Fisher Scientific Co., Fair Lawn, N.J.;2-, 3-, and 4-nitrotoluene, 2- and 3-nitrobenzyl alcohol, and
* Corresponding author.
3-nitrobenzoate, Aldrich Chemical Co., Inc., Milwaukee,Wis.; 2-methyl-5-nitrophenol, Pfaltz and Bauer, Stanford,Conn.; 1802, MSD Isotopes, Montreal, Quebec, Canada;isopropyl-,-D-thiogalactopyranoside (IPTG), BoehringerMannheim Biochemicals, Indianapolis, Ind.Organisms and culture conditions. Pseudomonas sp. strain
JS150 (12), P. putida Fl (9), and E. coli JM109(pDTG601)(27) were isolated as described in the references cited.Liquid cultures of JS150 and P. putida Fl were grown inminimal salts medium (MSB) (23) with toluene as describedpreviously (9).
Transformation of nitrotoluene by JS150 and P. putida Fl.Pseudomonas strains were grown overnight on toluene,harvested by centrifugation, and suspended in 100 ml ofdilute MSB (1:4) to anA6. of 0.4 to 0.75 (0.10 to 0.23 mg ofprotein per ml). Nitrotoluenes were dissolved in the MSB toa final concentration of 10-4 M before cells were added.Flasks (140 ml) were sealed and incubated at 30°C on arotary shaker at 250 rpm. At designated intervals, samples(0.45 ml) were removed, mixed with 0.05 ml of methanol,and clarified by centrifugation. All samples were held on iceuntil analyzed by high-performance liquid chromatography(HPLC).
E. coli JM109(pDTG601) was grown at 37°C to an A600 of0.5 before IPTG was added to a final concentration of 1.0mM. After 1 h, cells were harvested by centrifugation,washed with 0.05 M KH2PO4 buffer (pH 7.3), and suspendedin 0.5 volumes of the same buffer containing 0.01 M sodiumpyruvate and 10-4 M nitrotoluene. Flasks (250 ml) contain-ing 25 ml of the amended cell suspension were incubated at30°C on a rotary shaker at 200 rpm. At intervals, samples(1.0 ml) were removed and acidified with 10 ,u of concen-trated phosphoric acid, and the cells were removed bycentrifugation. The clear supernatant solution was stored at-70°C until analyzed.Incorporation of 1802 into 4-nitrotoluene. Toluene-grown
cells of JS150 were suspended in dilute MSB (1:4) to give anA600 of 1.5 (0.54 mg of protein per ml). The cell suspension(500 ml) was transferred to a 1.0-liter round-bottom flask,sealed with a stopcock, and stirred with a magnetic stirrer at25°C. The oxygen in the headspace of the flask was adjusted
2643
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 18
6.21
6.68
.130
.

2644 ROBERTSON ET AL.
80
60
40
20
CM
a)
00
0
100
80
60
40-
20
0
100
80'
60
40
20
._
C
0)
CZ
0
0
Time (hours)FIG. 1. Oxidation of 2-, 3-, and 4-nitrotoluene by toluene-grown
cells of JS150. Cell suspensions contained protein concentrations of0.10 mg/ml (A), 0.23 mg/ml (B), and 0.20 mg/ml (C). Nitrotolueneswere added to a concentration of 10-4 M.
to contain 52% 1602, 46% 1802, and 2% 1602-)802 as de-scribed previously (22). 4-Nitrotoluene was added to a finalconcentration of 10-4 M. A control culture contained air inthe headspace. Samples from the control culture were takenas described above and analyzed by HPLC for the disap-pearance of 4-nitrotoluene and the appearance of metabo-lites. After complete converison of 4-nitrotoluene (1.75 h),the reaction mixtures from both flasks were centrifuged toremove cells and the clear supernatant solutions were ex-tracted with equal volumes of ethyl acetate. The extractswere dried over anhydrous sodium sulfate and the solventwas removed in vacuo at 40°C. The residues from each flaskwere dissolved in ethyl acetate and analyzed by gas chro-matography-mass spectrometry (GC-MS).
Isolation and purification of 3-methyl-6-nitrocatechol. Cells
Time (hours)FIG. 2. Oxidation of 2-, 3-, and 4-nitrotoluene by toluene-grown
cells of P. putida Fl. Cell suspensions contained protein concentra-tions of 0.10 mg/ml (A and B) and 0.16 mg/ml (C). Nitrotolueneswere added to a concentration of 10-4 M.
of JS150 were grown overnight on toluene in 17 liters ofMSBin a Virtis model 43-100 20-liter fermentor. Cells wereharvested with a Pellicon cassette cell concentrator (Milli-pore Corporation, Bedford, Mass.). The pore size of thecassette was 0.45 ,um. Cells were washed with MSB andsuspended in 6 liters of dilute (1:4) MSB to give an opticaldensity of 1.8 at 600 nm (0.66 mg of protein per ml).4-Nitrotoluene in acetone was added in two equal portions togive a final concentration of 3.5 x 10-4 M. The suspensionwas incubated at 30°C until 4-nitrotoluene was no longerdetected by HPLC (1.5 h). The cells were removed asdescribed above, and the supernatant solution was extractedtwice with 3 liters of methylene chloride. The extract wasdried over anhydrous sodium sulfate, and the solvent was
BCH3
&N02
CH20H
&N02 COOH(NO 0--
APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 18
6.21
6.68
.130
.
MONOOXYGENATION OF NITROTOLUENES 2645
removed to yield a brown oil. This residue was dissolved in2 ml of hexane and applied to the top of a column (2.8 by 31cm) of silica gel. The column was eluted with benzene:hexane (6:4), and 250-ml fractions were collected. Fractions13 to 23 were pooled, and the solvent was removed by flashevaporation to leave yellow crystals (mp, 60 to 61°C).Analysis by HPLC, nuclear magnetic resonance (NMR), andGC-MS revealed a single compound subsequently identifiedas 3-methyl-6-nitrocatechol (see Results).
Analytical methods. HPLC separations were done with,uBondapak C18 columns (4.6 mm by 25 cm; Waters Associ-ates Inc., Milford, Mass.). For the experiments described inFig. 1 and 2, UVA210 was monitored with an HP1040A diodearray detector (Hewlett-Packard Corp., Palo Alto, Calif.).Compounds were eluted with a mobile phase of methanol,water, and phosphoric acid (370:630:1) at a flow rate of 1.5ml/min. Retention volumes for nitroaromatic compoundswere as follows: 2-nitrotoluene, 16.1 ml; 3-nitrotoluene, 18.3ml; 4-nitrotoluene, 17.4 ml; 2-nitrobenzoyl alcohol, 6.0 ml;3-nitrobenzoyl alcohol, 5.6 ml; 3-nitrobenzoic acid, 8.4 ml;3-methyl-6-nitrocatechol, 11.4 ml; 2-hydroxy-4-nitrotoluene,9.1 ml. For the experiments shown in Fig. 4, the UVabsorbance was monitored with a 990-MS photodiode arraydetector (Waters Associates Inc.), and the flow rate was 1.0ml/min. The mobile phase described above was used for 11min, followed by a convex gradient curve where the meth-anol concentration was increased to 90% over a 10-minperiod. Identification and quantification of substrates andmetabolites was accomplished by comparison with authenticstandards. Identification of nitrobenzoyl alcohols was con-firmed by GC-MS. GC-MS analyses were conducted on aHewlett-Packard model 5890 gas chromatograph equippedwith a fused-silica capillary column (0.25 mm by 30 m) witha 1-,um DB-5 stationary phase (J&W Scientific, Folsom,Calif.). Helium was the carrier gas. The oven temperaturewas maintained at 40°C for 4 min and then increased to 250°Cat a rate of 10°C/min. Mass spectra of compounds elutingfrom the column were obtained with a Hewlett-Packardmodel 5970 mass selective detector. Proton (1H) and carbon(13C) NMR spectra were obtained with a Bruker AC-300Fourier transform NMR spectrometer at a field of 7.2 T. Thecorresponding resonance frequencies for 'H and 13C at thisfield strength are 300.135 and 75.469 MHz, respectively.Samples were dissolved in acetone-d6 (99.8%). Predicted 1 Cchemical shifts were calculated by the method described byLevy et al. (15). The protein concentration in cell suspen-sions was determined with the bicinchoninic acid assayprocedure described by Smith et al. (18), with bovine serumalbumin as the standard.
RESULTS
Oxidation of nitrotoluenes by Pseudomonas sp. strain JS150and P. putida Fl. Glucose-grown cells of Pseudomonas sp.strain JS150 and P. putida Fl did not catalyze the oxidationof 2-, 3-, or 4-nitrotoluene. In contrast, toluene-grown cellsofPseudomonas sp. strain JS150 and P. putida Fl catalyzedthe almost stoichiometric oxidation of 2-nitrotoluene to2-nitrobenzyl alcohol (Fig. 1A and 2A). Experiments with1802 showed the incorporation of one atom of molecularoxygen, indicating that 2-nitrobenzyl alcohol is formed by amonooxygenation of 2-nitrotoluene (data not shown). JS150also produced small amounts of a transient metabolite (datanot shown). The mass spectrum of this compound gave aparent ion at mlz 169 with major fragments at m/z 121 andm/z 65. These properties are consistent with the structure of
15
51
0
(I).-
a)ca1)a-
cta1)
104
51
0 65 CH3 A
3yt0H 169NO2
o - 3 121
65 B
1690 39
30507090121 173Ld1~ 51070- ~ IL -j -
30 50 70 90 110 130 150 170 190 210
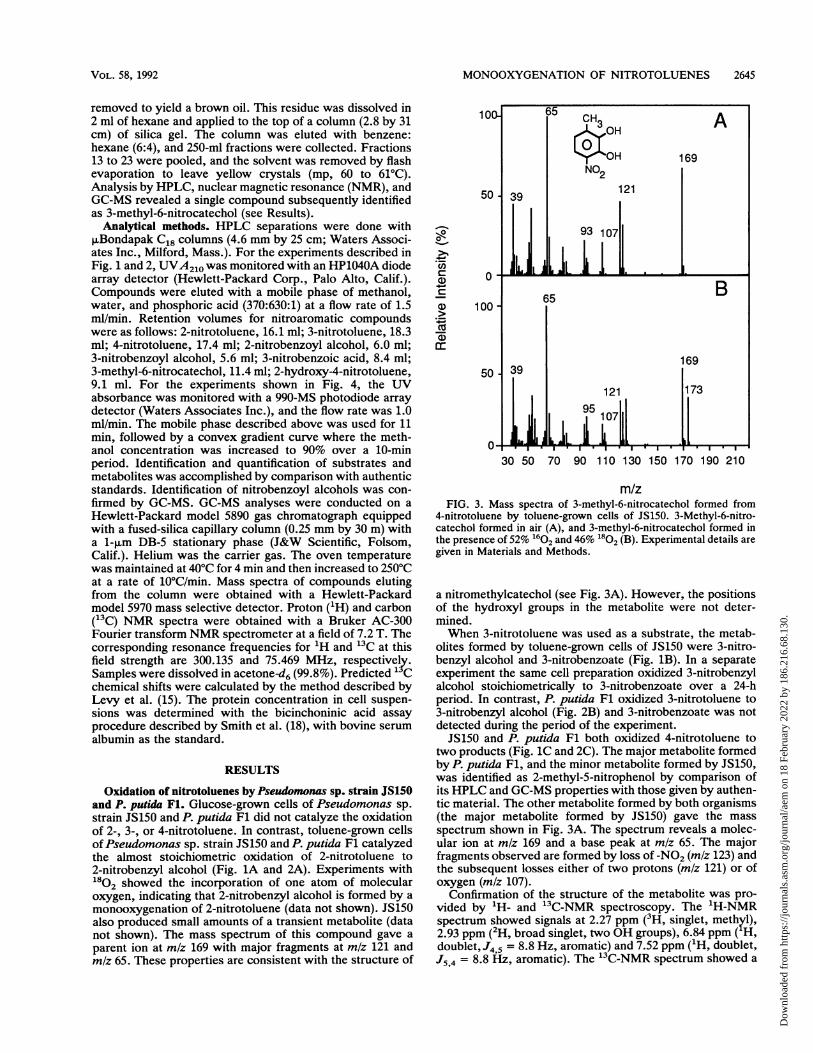
m/zFIG. 3. Mass spectra of 3-methyl-6-nitrocatechol formed from
4-nitrotoluene by toluene-grown cells of JS150. 3-Methyl-6-nitro-catechol formed in air (A), and 3-methyl-6-nitrocatechol formed inthe presence of 52% 1602 and 46% 1802 (B). Experimental details aregiven in Materials and Methods.
a nitromethylcatechol (see Fig. 3A). However, the positionsof the hydroxyl groups in the metabolite were not deter-mined.When 3-nitrotoluene was used as a substrate, the metab-
olites formed by toluene-grown cells of JS150 were 3-nitro-benzyl alcohol and 3-nitrobenzoate (Fig. 1B). In a separateexperiment the same cell preparation oxidized 3-nitrobenzylalcohol stoichiometrically to 3-nitrobenzoate over a 24-hperiod. In contrast, P. putida Fl oxidized 3-nitrotoluene to3-nitrobenzyl alcohol (Fig. 2B) and 3-nitrobenzoate was notdetected during the period of the experiment.JS150 and P. putida Fl both oxidized 4-nitrotoluene to
two products (Fig. 1C and 2C). The major metabolite formedby P. putida Fl, and the minor metabolite formed by JS150,was identified as 2-methyl-5-nitrophenol by comparison ofits HPLC and GC-MS properties with those given by authen-tic material. The other metabolite formed by both organisms(the major metabolite formed by JS150) gave the massspectrum shown in Fig. 3A. The spectrum reveals a molec-ular ion at mlz 169 and a base peak at mlz 65. The majorfragments observed are formed by loss of -NO2 (m/z 123) andthe subsequent losses either of two protons (m/z 121) or ofoxygen (m/z 107).
Confirmation of the structure of the metabolite was pro-vided by 'H- and 13C-NMR spectroscopy. The 'H-NMRspectrum showed signals at 2.27 ppm (3H, singlet, methyl),2.93 ppm (2H, broad singlet, two OH groups), 6.84 ppm ( H,doublet, J4,5 = 8.8 Hz, aromatic) and 7.52 ppm ('H, doublet,J5,4 = 8.8 Hz, aromatic). The 13C-NMR spectrum showed a
VOL. 58, 1992
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 18
6.21
6.68
.130
.

2646 ROBERTSON ET AL.
100
80
60
40
20
0100
80
0._
a)
0CD00
60-
40 - HH3
rrOHNOH
00 1 2 3 4 5 6 0 1 2 3 45 6
Time (hours)FIG. 4. Oxidation of 2-, 3-, and 4-nitrotoluene by IPTG-induced cells of E. coli JM109(pDTG601). Cell suspensions contained 0.4 mg of
protein per ml (A to C) and 0.72 mg of protein per ml (D). Substrates were added to a final concentration of 10-4 M.
singlet at 16.43 ppm (methyl) and peaks at 115.4, 122.2, 133.6134.2, 143.8, and 145.9 ppm for six aromatic carbon atoms(17). The NMR spectra along with the mass spectrumconfirm that the metabolite formed from 4-nitrotoluene byJS150 is 3-methyl-6-nitrocatechol.Toluene-grown cells of JS150 were incubated with 4-nitro-
toluene in the presence of a mixture of 1602 and 1802. Themass spectrum of the 3-methyl-6-nitrocatechol formed underthese conditions is shown in Fig. 3B. The results indicatethat both atoms of oxygen in 3-methyl-6-nitrocatechol orig-inate from a single oxygen molecule.
Oxidation of nitrotoluenes by E. coli JM109(pDTG601A).The preferential oxidation of the methyl group in 2- and3-nitrotoluene by P. putida Fl and JS150 was unexpected.The possible role of toluene dioxygenase in the formation ofnitrobenzyl alcohols was investigated by incubating 2- and3-nitrotoluene with IPTG-induced cells of E. coli JM109(pDTG601). This organism contains the toluene dioxygenasegenes (todClC2BA) from P. putida Fl in the expressionvector pKK223-3, where they are under the control of thetac promoter (27). Expression of the cloned toluene dioxy-genase genes in E. coli JM109(pDTG601) yields cells thatoxidize 2- and 3-nitrotoluene to 2- and 3-nitrobenzyl alcohol,respectively (Fig. 4). Nitrobenzoates were not detected. The
same induced cell preparations oxidized 4-nitrotoluene to2-methyl-5-nitrophenol and 3-methyl-6-nitrocatechol (Fig.4C). The latter gave a mass spectrum identical to that shownin Fig. 3A for the 3-methyl-6-nitrocatechol formed by JS150.In a separate experiment, induced cells of E. coli JM109(pDTG601) oxidized 2-methyl-5-nitrophenol to 3-methyl-6-nitrocatechol (Fig. 4D). Toluene-grown cells of P. putida Flgave similar results, whereas cells of JS150 oxidized 2-methyl-5-nitrophenol at very low rates (data not shown). Incontrol experiments, E. coli JM109(pKK223-3), which doesnot contain the todCJC2BA genes, was incubated for 6 hwith 2-, 3-, and 4-nitrotoluene. No products were detectedby HPLC under conditions where the detection limits were<0.005 mM.
DISCUSSION
The initial reaction in toluene degradation by JS150 and P.putida Fl is catalyzed by toluene dioxygenase and yieldscis-toluene dihydrodiol as the first metabolite (8, 10, 12).Subsequent oxidation of cis-toluene dihydrodiol leads to theformation of 3-methylcatechol, which undergoes ring fission(Fig. 5A). Analogous reactions were expected when toluene-grown cells of P. putida Fl and JS150 were incubated with
APPL. ENvIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 18
6.21
6.68
.130
.

MONOOXYGENATION OF NITROTOLUENES 2647
(B)
0N02
2-Nitrotoluene
CH20H
N02
2-Nitrobenzyl Alcohol
(C)
CH3
NH23-Nitrotoluene
CH2OH
N4~3-Nitrobenzyl Alcohol
COOH
NNo 2
2-Nitrobenzoate
FIG. 5. Initial reactions in the oxidation of toluene (A), 2-nitro-toluene (B), and 3-nitrotoluene (C) by JS150 and P. putida Fl. E.coli JM109(pDTG601), which contains the cloned toluene dioxygen-ase genes from P. putida Fl, catalyzes the formation of cis-toluenedihydrodiol, 2-nitrobenzyl alcohol, and 3-nitrobenzyl alcohol fromtoluene (27) and 2- and 3-nitrotoluene, respectively.
2-, 3-, and 4-nitrotoluene. However, the major productsformed from 2- and 3-nitrotoluene by both organisms were 2-and 3-nitrobenzyl alcohol, respectively. The same nitroben-zyl alcohols were formed by E. coli JM1O9(pDTG601) wherethe todCJC2BA genes, encoding toluene dioxygenase, areunder the control of the IPTG-inducible tac promoter (Fig.
CH3
NO24-Nitrotoluene
4A and B). These results clearly demonstrate that toluenedioxygenase is the enzyme responsible for the oxidation ofthe methyl substituent. This novel monooxygenase activityof toluene dioxygenase extends the range of substratesknown to be oxidized by this enzyme system (11). Earlystudies with Pseudomonas fluorescens suggested that ben-zoate dioxygenase could catalyze the oxidation of the methylgroup of o-toluate, but the activity was not rigorouslyattributed to the dioxygenase (14). Previous studies haveshown that toluene dioxygenase from P. putida Fl catalyzesthe monooxygenation of indan and indene to benzylic alco-hols (2, 25). In addition, a mutant strain (UV4) of P. putidathat oxidizes toluene to cis-toluene dihydrodiol also oxidizesthe bicyclic alkenes 1,2-dihydronaphthalene, indene, and1,2-benzocyclohepta-1,3-diene to bicyclic monols (1). Theseobservations indicate that the current distinctions betweenmonooxygenase and dioxygenase enzymes may be arbitraryand substrate dependent.
P. putida Fl and JS150 oxidized 4-nitrotoluene to 2-methyl-5-nitrophenol and 3-methyl-6-nitrocatechol. Experi-ments with 180 showed that toluene-induced cells of JS150incorporated both atoms of molecular oxygen into the aro-matic nucleus to form 3-methyl-6-nitrocatechol (Fig. 3).These results can be explained by the oxidation of 4-nitrotol-uene to 1,2-dihydroxy-3-methyl-6-nitrocyclohexa-3,5-diene(4-nitrotoluene-2,3-dihydrodiol). Enzymatic dehydrogenationof the dihydrodiol by toluene dihydrodiol dehydrogenasewould yield 3-methyl-6-nitrocatechol. These reactions areanalogous to those used by many bacteria to initiate thedegradation of aromatic hydrocarbons and related com-pounds (10). The formation of 2-methyl-5-nitrophenol byJS150 could be due to the nonenzymatic dehydration of theputative 4-nitrotoluene-2,3-dihydrodiol. The same rationalecan be used to explain the oxidation of 4-nitrotoluene to2-methyl-5-nitrophenol and 3-methyl-6-nitrocatechol by P.putida Fl. However, the formation of significant amounts of2-methyl-5-nitrophenol suggests that the toluene dihydrodioldehydrogenase in P. putida Fl is much less active than theisofunctional enzyme in JS150 with 4-nitrotoluene-2,3-dihy-drodiol as a substrate. In addition, the oxidation of 4-nitrotol-uene to 3-methyl-6-nitrocatechol by E. coli JM1O9(pDTG601)
JS150Pp Fl
"Dihydrodiol"
OH-
CH3OOH
OH
N023-Methyl-6-Nitrocatechol
t_OH-
OH3 OH3
OH 0 \
Pp Fl jH
NO2 E. coli JM109(pDTG601) NO2
2-Methyl-5-NitrophenolFIG. 6. Proposed reaction mechanism for formation of 3-methyl-6-nitrocatechol from 4-nitrotoluene.
(A)
CH3
Toluene
CH3
H
cis-dihydrodiol
CH3HOH
OH
3-methylcatechol
VOL. 58, 1992
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 18
6.21
6.68
.130
.

2648 ROBERTSON ET AL.
cannot proceed directly through 4-nitrotoluene-2,3-dihydro-diol since this recombinant organism does not express toluenedihydrodiol dehydrogenase. The observation that P. putidaFl and E. coli JM109(pDTG601) catalyze the oxidation of2-methyl-5-nitrophenol to 3-methyl-6-nitrocatechol suggeststhat the mechanism involves the initial oxidation of 4-nitro-toluene to 4-nitrotoluene-2,3-dihydrodiol by toluene dioxy-genase. Nonenzymatic dehydration of the dihydrodiol wouldyield 2-methyl-5-nitrophenol, which would undergo furtheroxidation to the equivalent of a hydrated ketone. The latterwould immediately rearomatize to 3-methyl-6-nitrocatechol(Fig. 6). Analogous reactions have been postulated to accountfor the oxidation of substituted phenols to catechols bytoluene dioxygenase (19, 22). Scenarios other than thosegiven above are possible and await the results of experimentsin progress with purified dioxygenases from P. putida Fl andJS150.
ACKNOWLEDGMENTS
This work was supported in part by grant AFOSR 88-6225 fromthe U.S. Air Force Office of Scientific Research to D.T.G.We thank Stephen Aubert and Paul Bolduc for obtaining and
interpreting the NMR spectra, Howard Mayfield, and Chris Ant-worth for the GC-MS analyses, Joe Bernardo for technical assis-tance, and Sharon Gaffney for assistance in preparing the manu-script.
REFERENCES1. Boyd, D. R., R. A. S. McMordie, N. D. Sharma, H. Dalton, P.
Williams, and R. 0. Jenkins. 1989. Stereospecific benzylichydroxylation of bicyclic alkenes by Pseudomonas putida:isolation of (+)-R-1-hydroxy-1,2-dihydronaphthalene, an arenehydrate of naphthalene from metabolism of 1,2-dihydronaphtha-lene. Chem. Commun. (J. Chem. Soc. Sect. D) 1989:339-340.
2. Brand, J. M., D. L. Cruden, and D. T. Gibson. Unpublishedresults.
3. Bruhn, D., H. Lenke, and H.-J. Knackmuss. 1987. Nitrosubsti-tuted aromatic compounds as nitrogen source for bacteria.Appl. Environ. Microbiol. 53:208-210.
4. Cain, R. B. 1958. The microbial metabolism of nitro-aromaticcompounds. J. Gen. Microbiol. 19:1-14.
5. Cartwright, N. J., and R. B. Cain. 1959. Bacterial degradation ofthe nitrobenzoic acids. J. Biochem. 71:248-261.
6. Delgado, A., M. G. Wubbolts, M.-A. Abril, and J. L. Ramos.1992. Nitroaromatics are substrates for the TOL plasmid upper-pathway enzymes. Appl. Environ. Microbiol. 58:415-417.
7. Fewson, C. A. 1981. Biodegradation of aromatics with industrialrelevance, p. 141-179. In T. Leisinger, A. M. Cook, R. Huttler,and J. Neusch (ed.), Microbiol degradation of xenobiotic andrecalcitrant compounds. Academic Press, London.
8. Gibson, D. T., M. Hensley, H. Yoshioka, and T. J. Mabry. 1970.Formation of (+)-cis-2,3-dihydroxy-1-methylcyclohexa-4,6-di-ene from toluene by Pseudomonas putida. Biochemistry9:1626-1630.
9. Gibson, D. T., J. R. Koch, and R. E. Kallio. 1968. Oxidativedegradation of aromatic hydrocarbons by microorganisms. I.Enzymatic formation of catechol from benzene. Biochemistry7:2653-2662.
10. Gibson, D. T., and V. Subramanian. 1984. Microbial degrada-
tion of aromatic hydrocarbons, p. 181-252. In D. T. Gibson(ed.), Microbial degradation of organic compounds. MarcelDekker, New York.
11. Gibson, D. T., G. J. Zylstra, and S. Chauhan. 1990. Biotrans-formations catalyzed by toluene dioxygenase from Pseudomo-nas putida Fl, p. 121-132. In S. Silver, A. M. Chakrabarty, B.Iglewski, and S. Kaplan (ed.), Pseudomonas. Biotransforma-tions, pathogenesis, and evolving technology. American Societyfor Microbiology, Washington, D.C.
12. Haigler, B. E., C. A. Pettigrew, and J. C. Spain. 1992. Biodeg-radation of mixtures of substituted benzenes by Pseudomonassp. strain 150. Appl. Environ. Microbiol. 58:2237-2244.
13. Hallas, L. E., and M. Alexander. 1983. Microbial transformationof nitroaromatic compounds in sewage effluent. Appl. Environ.Microbiol. 45:1234-1241.
14. Ichihara, A., K. Adachi, K. Hosokawa, and Y. Takeda. 1962.The enzymatic hydroxylation of aromatic carboxylic acids;substrate specificities of anthranilate and benzoate oxidases. J.Biochem. 237:2296-2302.
15. Levy, G. C., R. L. Lichter, and G. L. Nelson. 1980. Carbon-13nuclear magnetic resonance spectroscopy, p. 111-112. JohnWiley and Sons, New York.
16. McCormick, N. G., F. E. Feeherry, and H. S. Levinson. 1976.Microbial transformation of 2,4,6-trinitrotoluene and other ni-troaromatic compounds. Appl. Environ. Microbiol. 31:949-958.
17. Silverstein, R. M., G. C. Bassler, and T. C. Morrill. 1981.Spectrometric identification of organic compounds, 4th ed.,p. 22, 32-33. John Wiley and Sons, New York.
18. Smith, P. K., R. I. Krohn, G. T. Hermanson, A. K. Mallia, F. H.Gartner, M. D. Provenzano, E. K. Fujimoto, N. M. Goeke, B. J.Olson, and D. C. KDenk. 1958. Measurement of protein usingbicinchoninic acid. Anal. Biochem. 150:76-85.
19. Spain, J. C., and D. T. Gibson. 1988. Oxidation of substitutedphenols by Pseudomonas putida Fl and Pseudomonas sp.strain JS6. Appl. Environ. Microbiol. 54:1399-1404.
20. Spain, J. C., and D. T. Gibson. 1991. Pathway for biodegrada-tion ofp-nitrophenol in a Moraxella sp. Appl. Environ. Micro-biol. 57:812-819.
21. Spain, J. C., 0. Wyss, and D. T. Gibson. 1979. Enzymaticoxidation of p-nitrophenol. Biochem. Biophys. Res. Commun.88:634-641.
22. Spain, J. C., G. J. Zylstra, C. K. Blake, and D. T. Gibson. 1989.Monohydroxylation of phenol and 2,5-dichlorophenol by tolu-ene dioxygenase in Pseudomonas putida Fl. Appl. Environ.Microbiol. 55:2648-2652.
23. Stanier, R. Y., N. J. Palleroni, and M. Doudoroff. 1966. Theaerobic pseudomonads: a taxonomic study. J. Gen. Microbiol.43:159-271.
24. U.S. Environmental Protection Agency. 1986. Health and envi-ronmental effects profile for nitrotoluenes (o-, m-, p-). ReportEPA/600/X-86/143. Environmental Criteria and Assessment Of-fice, Cincinnati, Ohio.
25. Wackett, L. P., L. D. Kwart, and D. T. Gibson. 1988. Benzylicmonooxygenation catalyzed by toluene dioxygenase from Pseu-domonas putida. Biochemistry. 27:1360-1367.
26. Zeyer, J., and P. C. Kearney. 1984. Degradation of o-nitrophe-nol and m-nitrophenol by a Pseudomonasputida. J. Agric. FoodChem. 32:238-242.
27. Zylstra, G. J., and D. T. Gibson. 1989. Toluene degradationby Pseudomonas putida Fl: nucleotide sequence of thetodClCfBADE genes and their expression in Escherichia coli. J.Biol. Chem. 264:14940-14946.
APPL. ENvIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 18
Feb
ruar
y 20
22 b
y 18
6.21
6.68
.130
.