overexpression of protein kinase cγ inhibits gap junctional intercellular communication in the lens...
TRANSCRIPT

Exp. Eye Res. (2000) 71, 99±102doi:10.1006/exer.2000.0847, available online at http://www.idealibrary.com on
LETTER TO THE EDITORS
Overexpression of Protein Kinase Cggggg Inhibits Gap JunctionalIntercellular Communication in the Lens Epithelial Cells
Lens epithelial cells represent the anterior monolayerof cells in the lens. They communicate through gap
junctions, which are formed of connexin 43 (Cx 43)
protein (Goodenough, 1992; Reynhout, Lamp and
0014-4835/00/070099�04 $35.00/0
PKC α Overexpression
+ Zn − Zn
PKC γ Overexpression
+ Zn − Zn
(A)
(B)
(C)
FIG. 1. Western blots and densitometric analysis of theoverexpressed PKCa and PKCg in N/N1003 cells afterinducing them with zinc; (A) Western blots of transfectedN/N1003 with PKCa + zinc for 24 hr; (B) Western blotsof transfected N/N1003 with PKCg + zinc for 24 hr;(C) Densitometric Scan intensity of overexpressed PKCaand PKCg in N/N1003 cells.
Johnson, 1992; Albert and Jakobiec, 1994; Bassnettet al., 1994; Le and Musil, 1998). Protein Kinase C(PKC) has been implicated in the inhibition ofgap junctions in different cell lines by using thetumor promoter TPA (12-O-tetradecanoylphorbol-13-acetate), which is an activator of PKC (Yada, Rose andLoewenstein, 1985; Reynhout et al., 1992; Saez et al.,1998).
In this report, rabbit lens epithelial cell line(N/N1003) was used as a model to investigate theeffect of PKC isoforms on the gap junction inter-cellular communication (GJIC) in the lens epithelialcells. In those cells, the major isoforms of PKC arePKCa and PKCg (Gonzalez, Udovichenko andTakemoto, 1993). To determine which isoform isresponsible for the regulation of gap junction activity,both PKCa and PKCg were overexpressed and
the scrape-loading/dye transfer assay was used toevaluate the gap junction activity.# 2000 Academic Press
FIG. 2. Phase contrast photographs of N/N1003 cells;(A) 40� magni®cation of the lens epithelial cells grown to90 % con¯uency to use for the SL/DT assay; (B) 10�magni®cation of the lens epithelial cells showing one of thetwo cuts after scraping them to do the SL/DT assay.
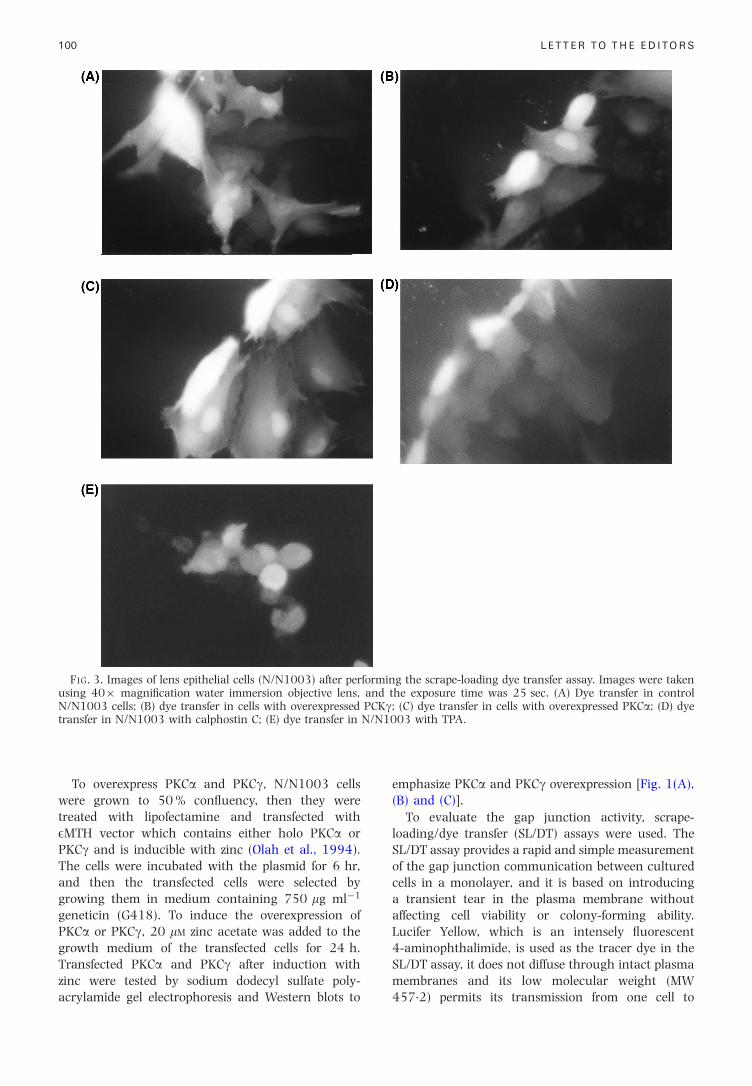
FIG. 3. Images of lens epithelial cells (N/N1003) after performing the scrape-loading dye transfer assay. Images were takenusing 40� magni®cation water immersion objective lens, and the exposure time was 25 sec. (A) Dye transfer in controlN/N1003 cells; (B) dye transfer in cells with overexpressed PCKg; (C) dye transfer in cells with overexpressed PKCa; (D) dyetransfer in N/N1003 with calphostin C; (E) dye transfer in N/N1003 with TPA.
100 LETTER TO THE EDITORS
To overexpress PKCa and PKCg, N/N1003 cellswere grown to 50 % con¯uency, then they weretreated with lipofectamine and transfected withEMTH vector which contains either holo PKCa orPKCg and is inducible with zinc (Olah et al., 1994).The cells were incubated with the plasmid for 6 hr,and then the transfected cells were selected bygrowing them in medium containing 750 mg mlÿ1
geneticin (G418). To induce the overexpression ofPKCa or PKCg, 20 mM zinc acetate was added to thegrowth medium of the transfected cells for 24 h.Transfected PKCa and PKCg after induction with
zinc were tested by sodium dodecyl sulfate poly-acrylamide gel electrophoresis and Western blots toemphasize PKCa and PKCg overexpression [Fig. 1(A),(B) and (C)].
To evaluate the gap junction activity, scrape-loading/dye transfer (SL/DT) assays were used. TheSL/DT assay provides a rapid and simple measurementof the gap junction communication between culturedcells in a monolayer, and it is based on introducinga transient tear in the plasma membrane withoutaffecting cell viability or colony-forming ability.Lucifer Yellow, which is an intensely ¯uorescent4-aminophthalimide, is used as the tracer dye in theSL/DT assay, it does not diffuse through intact plasma
membranes and its low molecular weight (MW457.2) permits its transmission from one cell to
(B)
(A)
FIG. 4. Analysis of the results of the SL/DT assay; (A)graphical representation of SL/DT assay in control N/N1003cells, control � TPA, control � DMSO (control for the TPAexperiment since TPA was dissolved in DMSO), con-trol � calphostin C, N/N1003 cells that express PKCa andthat express PKCg; (B) results of the SL/DT assay representedas the mean result + standard error of the mean (S.E.M.). n:number of the experiments which were done in differentcoverslips (three SL/DT experiments/coverslip); DMSO:dimethyl sulfoxide was used as a control for TPA becauseTPA was dissolved in DMSO, therefore, to make sure that theinhibition of gap junction was due to TPA and not DMSOcells were incubated in media with 0.004 % DMSO (which isthe same amount of DMSO which was introduced to the
101
another presumably through patent gap junctions.Rhodamine dextran has a high molecular weight(MW 10 000) and it can neither diffuse throughintact plasma membranes nor cross the gap junctions,therefore it is used as a control for the dye transfer(El-Fouly, Trosko and Chang, 1987). In the SL/DTassay, N/N1003 cells were grown to 90 % con¯uency[Fig. 2(A)] on glass coverslips, washed three timeswith Hank's balanced salt solution containing 1 %bovine serum albumin and 1 mM CaCl2 (HBC). Then2.5 ml of 1 % Lucifer Yellow (Sigma) or 0.75 %Rhodamine dextran (molecular probes, control fordye transfer), made with Phosphate buffered saline(PBS), was added at the center of the coverslip andusing a 27 gauge needle, two cuts crossing each otherwere made on the coverslip, passing through the dyein the center of the coverslip [Fig. 2(B)]. These cellswere incubated with the dye for 1 min, after that thecells were washed three times with HBC andincubated in tissue culture medium for 10 min atroom temperature to allow dye transfer. The cells werethen washed three times with PBS and ®xed with 2 %paraformaldehyde for 30 min at room temperature.Dye transfer was evaluated by examining the cells by¯uorescent microscopy (Olympus BHS) equipped with¯uorescein and rhodamine ®lter sets. Photographswere obtained using a 40� objective. For quanti-tation, the extent of dye transfer was estimated bycounting the number of ¯uorescent labeled cells in themicroscopic ®eld under 40� magni®cation in ran-domly selected areas away from the scrape line on thecoverslip (El-Fouly et al., 1987; Mckarns and Doo-little, 1992; Le and Musil, 1998).
After evaluating the gap junction activity by thescrape-loading dye transfer assay in the controlN/N1003 cells and in the transfected cells, we foundthat the cells which overexpress PKCg had a signi®-cantly decreased dye transfer (mean of 4.76 cells)[Fig. 3(B)] compared to the dye transfer in the controlN/N1003 cells shown in Fig. 3(A) (mean of10.77 cells). In the cells that overexpress PKCa, dyetransfer was almost the same as in the controlN/N1003 cells as shown in Fig. 3(C). Calphostin C(1 mM) was used as a positive control for the gapjunction activity, as it is an inhibitor of PKC(Kanashiro and Khalid, 1998). Control N/N1003cells were incubated in tissue culture medium withcalphostin C for 24 hr, then the gap junction activitywas evaluated by SL/DT assay. As shown in Fig. 3(D)dye transfer was increased to a mean of 14.55 cells.The tumor promoter TPA (12-O-tetradecanoyl-phorbol-13-acetate, 200 nM) was used as a negativecontrol for the gap junction activity, as it is anactivator of PKC (Yada et al., 1985; Asamoto et al.,1991; Reynhout et al., 1992; Lamp, 1994; Kanashiroand Khalid, 1998). For this experiment control
LETTER TO THE EDITORS
N/N1003 cells were incubated in tissue culturemedium with TPA for 30 min. After performing the
gap junction assay dye transfer was decreased[Fig. 3(E)] to a mean of 5.5 cells.
Fig. 4(B) summarizes the results of the gap junctionassay experiments that were done. It shows clearlythat gap junction activity decreased with increasingPKCg in the lens epithelial cells. On the other hand,increasing PKCa in the lens epithelial cells did notaffect the gap junction activity.
These results indicate that increasing the amountof PKCg by overexpression in the lens epithelial cellsinhibits the gap junctional intercellular communi-cation [Fig. 4(A)], while overexpression of PKCa didnot affect the gap junction activity. Therefore it isPKCg and not PKCa which regulates gating and gapjunctional channels in the lens epithelial cells. Thisregulation is most probably through phosphorylationof serine 368 residue of the carboxy-terminal onCx43 by PKCg (Musil et al., 1990; Stagg and Flecher,
cells with TPA) for 30 min and then SL/DT assay wasperformed.
1990; Laird, Puranam and Revel, 1991; Lau et al.,1996).

Acknowledgements
The authors thank John Reddan for providing the N/N1003cells, Wayne Anderson for providing PKCa and PKCgplasmids, Linda Musil, Michael Kanost, Larry Takemoto,Thomas Steinberg, Karen Gonzalaz and Peggy Zelenka fortheir useful technical advice. Also we would like to thankKSU Cancer Center for providing ®nancial assistance tolearn about gap junction assays at Dr Thomas Stenberg'slaboratory at Washington University Medical School.
SUHA M. SALEHa*DOLORES J. TAKEMOTOb
aDepartment of Anatomy and Physiology, KansasState University, Manhattan, Kansas, 66506,U.S.A. and bDepartment of Biochemistry, KansasState University, Manhattan, Kansas, 66506,U.S.A.
102
* Address correspondence to: S. M. Saleh, Department ofAnatomy and Physiology, Kansas State University, Manhattan,
Kansas 66506, U.S.A.References
Albert, D. and Jakobiec, F. (1994). Principles and practice ofophthalmology. W. B. Saunders Company, Philadelphia.
Asamoto, M., Oyamada, M., Aoumari, A., Gros, D. andYamasaki, H. (1991). Molecular mechanisms of TPA-mediated inhibition of gap-junctional intercellularcommunication: evidence for action on the assemblyor function but not the expression of connexin 43 in ratliver epithelial cells. Mol. Carcinog. 4, 322±7.
Bassnett, S., Kuszak, J., Reinisch, L., Brown, H. and Beebe,D. (1994). Intercellular communication between epi-thelial and ®ber cells of the eye. J. Cell Sci. 107,799±811.
El-Fouly, M., Trosko, J. and Chang, C. (1987). Scrap-loadingand dye transfer a rapid and simple technique to studygap junctional intercellular communication. Exp. CellRes. 168, 422±30.
Gonzalez, K., Udovichenko, I. and Takemoto, D. (1993).Protein kinase C in galactose and tolrestat treated lensepithelial cells. Curr. Eye Res. 12, 373±7.
Goodenough, D. (1992). The crystalline lens. A system
(Received Rochester 3 December 1999 and ac
Kanashiro, C. and Khalid, R. (1998). Signal transduction byprotein kinase C in mammalian cells. Clin. Exp.Pharmacol. Physiol. 25, 974±85.
Laird, D., Puranam, K. and Revel, J. (1991). Turnover andphosphorylation dynamics of connexin 43 gap junctionprotein in cultured cardiac myocytes. Biochem. J. 273,67±72.
Lamp, P. (1994). Analyzing phorbol ester effects on gapjunctional communication: a dramatic inhibition ofassembly. J. Cell Biol. 127, 1895±905.
Lau, A., Kurata, W., Kanemitsu, M., Loo, L., Warn-Cramer,B., Eckhart, W. and Lamp, P. (1996). Regulation ofconnexin 43 function by activated tyrosine proteinkinases. J. Bioenerg Biomembr. 28, 359±68.
Le, A. and Musil, L. (1998). Normal differentiation ofcultured lens cells after inhibition of gap junction-mediated intercellular communication. Dev. Biol. 204,80±96.
Mckarns, S. and Doolittle, D. (1992). Limitations ofthe scrap-loading/dye transfer technique to quantifyinhibition of gap junctional intercellular communica-tion. Cell Biol. Toxicol. 8, 89±103.
Musil, L., Cunningham, B., Edelman, G. and Goodenough,D. (1990). Differential phosphorylation of the gapjunction protein connexin 43 in junctional communi-cation-competent and -de®cient cell lines. J. Cell Biol.111, 2077±88.
Olah, Z., Lehel, C., Jakob, G. and Anderson, W. (1994). Acloning and E-epitope-tagging insert for the expressionof polymerase chain reaction-generated cDNA frag-ments in Escherichia coli and mammalian cells. Anal.Biochem. 221, 94±102.
Reynhout, J., Lamp, P. and Johnson, R. (1992). An activatorof protein kinase C inhibits gap junction communi-cation between cultured bovine lens cells. Exp. Cell Res.198, 337±42.
Saez, J., Martinez, A., Branes, M. and Gonzalez, H. (1998).Regulation of gap junctions by protein phosphorylation.Braz. J. Med. Biol. Res. 31, 593±600.
Stagg, R. and Flecher, W. (1990). The hormone-inducedregulation of contact-dependent cell-cell communi-cation by phosphorylation. Endocr. Rev. 11, 302±25.
Yada, T., Rose, B. and Loewenstein, W. (1985). Diacylgly-
LETTER TO THE EDITORS
networked by gap junctional communication. Semin.Cell Biol. 3, 49±58.
cerol downregulates junctional membrane permeability.Tmb-8 blocks this effect. J. Membr. Biol. 88, 217±33.
cepted in revised form 22 February 2000)