overexpression of mdvha-b, a v-atpase gene from apple, confers tolerance to drought in transgenic...
TRANSCRIPT

Od
DNU
a
ARRA
KAVTAD
1
cnptieprppbpJm
aaif
s
0h
Scientia Horticulturae 145 (2012) 94–101
Contents lists available at SciVerse ScienceDirect
Scientia Horticulturae
journa l h o me page: www.elsev ier .com/ locate /sc ihor t i
verexpression of MdVHA-B, a V-ATPase gene from apple, confers tolerance torought in transgenic tomato
a-Gang Hu, Shu-Hui Wang, Hua Luo, Qi-Jun Ma, Yu-Xin Yao, Chun-Xiang You, Yu-Jin Hao ∗
ational Key Laboratory of Crop Science, National Research Center for Apple Engineering and Technology, College of Horticulture Science and Engineering, Shandong Agriculturalniversity, Taian 271018, Shandong, China
r t i c l e i n f o
rticle history:eceived 23 May 2012eceived in revised form 2 August 2012ccepted 7 August 2012
eywords:
a b s t r a c t
The survival of plant cells strongly depends on maintaining or adjusting the activity of the V-ATPase underdrought conditions. In this study, a V-ATPase gene, VHA-B, was identified in an apple cDNA library, and ananalysis of the deduced MdVHA-B protein sequence showed that it clusters with VHA-B proteins of otherspecies. The expression of MdVHA-B was induced by abiotic stress, including abscisic acid (ABA), salinity,drought and low and high temperatures. Furthermore, the over-expression of MdVHA-B in transgenic
pple-ATPase gene MdVHA-Bransgenic tomatoBArought stress
tomato plants resulted in a high tolerance to drought stress, reduced relative water loss and malondi-aldehyde contents and increased levels of free proline and H+-ATPase activity compared to the wild-typeplants (WT). Additionally, it has similar sensibility to exogenous ABA in transgenic MdVHA-B tomato seedsand seedlings, while more sensitive to polyethylene glycol in transgenic tomato compared with WT. Thisstudy indicates that MdVHA-B plays an important role in drought stress, yet may act independently of
y.
the ABA signaling pathwa. Introduction
The vacuolar H+-ATPase (V-ATPase) is a well-described andomplicated enzyme that is indispensable for plant growth underormal conditions due to its role in energizing secondary trans-ort, in the maintenance of ion homeostasis and in abiotic stressolerance (Klein, 1992; Lüttge et al., 2001; Dietz et al., 2001). Sim-lar to other enzymes in this class, the V-ATPases are multisubunitnzymes composed of two subcomplexes. The peripheral V1 com-lex, which comprises eight subunits (VHA-A to -H), is mainlyesponsible for ATP hydrolysis, and the membrane-integral V0 com-lex, which contains VHA-a, -c, -c′, -c′′, -d, and -e, is responsible forroton translocation from the cytosol into the lumen of endomem-rane compartments (Cipriano et al., 2008). Most of these protonump subunits are abundant tonoplast proteins (Carter et al., 2004;
aquinod et al., 2007), indicating that they are an essential invest-ent of energy for vacuolar transport.In Arabidopsis, the 13 subunits (no subunit c′ is found in plants)
re encoded by a total of 27 VHA-genes (Sze et al., 2002); there
re also possible isoform combinations that have not yet beendentified. Researchers have begun to elucidate the potential andunctional significance of these subunits. For instance, VHA-A andAbbreviations: V-ATPase, vacuolar-type adenosine triphosphatase; VHA-B, Bubunit of vacuolar-type H+-ATPase; MDA, malondialdehyde; ABA, abscisic acid.∗ Corresponding author. Tel.: +86 538 824 6692; fax: +86 538 824 2364.
E-mail address: [email protected] (Y.-J. Hao).
304-4238/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.scienta.2012.08.010
© 2012 Elsevier B.V. All rights reserved.
VHA-c are among the most highly conserved subunits, thereforeit is not surprising that the isoforms found within a species showvery little, if any, difference at the protein level. The two Arabidop-sis genes VHA-c1 and VHA-c3 encode identical proteins: VHA-c1is expressed ubiquitously, whereas VHA-c3 expression is limitedto the root caps (Padmanaban et al., 2004). Similarly, VHA-A1 oftomato is ubiquitous, whereas VHA-A2 is restricted to the roots andfruits (Bageshwar et al., 2005). In such cases of differential expres-sion, it seems more likely that changes in the regulatory sequencesfollowing gene duplications has allowed the evolution of morecomplex and adaptive expression patterns. However, althoughthe two tomatos VHA-A isoforms share 95% identity and are co-expressed in some tissues, the isoforms do not occur in the sameV-ATPase complexes, indicating differences in protein function. Ithas been reported that the functional differentiation between VHA-B1 and VHA-B3 (Cho et al., 2006), which originated from a recentduplication in the Arabidopsis genome (Sze et al., 2002), are 98%similarity. In addition, differences in the V-ATPase activity betweenplant organs and tissues or after changes in the nutritional statusmay either be correlated with changes in the structure of the com-plex or with the presence of individual protein isoforms (Dietz et al.,2001). Determining how the Arabidopsis isoforms affect the kineticand regulatory properties of the holocomplex and the functions ofthese subunits remain a challenging task.
There is also evidence that the V-ATPase subunits may haveadditional functions independent of proton pumping, thus addinganother layer of complexity to the biology of V-ATPases. Forexample, VHA-B1 has been identified in nuclear complexes with

orticul
hc2ptitt2t2id
gtpvattlt
2
2
ta‘I
Mcoam3(u
uaawc
2
bbtoDlrm
2
d
D.-G. Hu et al. / Scientia H
exokinase, and a series of phenotypes of a vha-B1 mutant indi-ate that this subunit is involved in glucose signaling (Cho et al.,006). These results indicate that V-ATPases are not merely “sim-le” proton-pumps. In addition, transgenic plants overexpressinghe wheat H+-PPase gene TVP1 (Brini et al., 2007) exhibitedmproved salt- and drought-stress tolerance, and transgenic toma-oes expressing AVP1 were more resistant to a soil water deficit thanhe controls because of their more robust root systems (Li et al.,005; Park et al., 2005) and proton pumps could transport waterhat allowed fruits to grow to a large size (Shiratake and Martinoia,007). These results indicate that enhanced vacuolar H+ pumping
n transgenic plants could be an effective approach to improve therought tolerance of crops.
The ectopic expression of the apple vacuolar H+-PPase MdVHP1ene enhances stress tolerance in both transgenic apple callus andomato (Dong et al., 2011), In the present study, we isolated anotherroton pump gene, MdVHA-B, which encodes the B subunit of theacuolar H+-ATPase. Its expression patterns in different tissuesnd in response to various stresses were analyzed using real-ime quantitative RT-PCR, and its functions were characterized inransgenic tomato plants. In addition, we discuss the potential uti-ization of this gene in the genetic improvement of plant toleranceo stress.
. Materials and methods
.1. Plant materials, growth conditions and stress treatments
The young leaves and other tissues of ‘Gala’ apple (Malus domes-ica Borkh) were used for the gene cloning and gene expressionssays, respectively. The tissues were obtained from 12-year-old
Gala’ apple trees planted in the Experimental Orchard of Shandongnstitute of Fruit Tree Science (Taian, China).
In vitro tissue cultures were used for analyzing the effects ofdVHA-B expression in response to abiotic stresses. The tissue
ultures were submerged in MS liquid medium at 4 ◦C or 40 ◦Cr with the inclusion of NaCl, polyethylene glycol (PEG 2000) orbscisic acid (ABA) under a maximum photoperiod of approxi-ately 600 mmol m−2 s−1 at 25 ◦C. Samples were collected at 0, 1,
, 6, 9, 12, 24, and 48 h after treatment for the expression analysisthe primers used are listed in Table 1S). MS liquid medium wassed as the control.
Tomato (Solanum lycopersicum M. cv. Tianjinbaiguo) plants weresed for the gene transformation and the drought tolerance assaysnd were maintained under a 14/10-h (light/dark) photoperiod atpproximately 600 mmol m−2 s−1 at 25 ◦C. For the drought assay,ater was withheld from 10-week-old tomato plants in a growth
hamber for 2 weeks, and the plants were then rewatered for 10 h.
.2. Molecular cloning of the MdVHA-B gene
Total RNA was extracted from apple fruit as previously describedy Yao et al. (2007). A partial MdVHA-B fragment was obtainedy reverse transcription-polymerase chain reaction (RT-PCR) usinghe degenerate primers. The flanking 3′ and 5′ regions werebtained using the rapid amplification of cDNA ends kit (TaKaRa,alian, China). The total genomic DNA was extracted from young
eaves described by Doyle and Doyle (1990), and the promoteregion was amplified using the PCR-based genomic walkingethod described by Wu and Liu (2006).
.3. Real-time quantitative RT-PCR analysis of gene expression
Apple total RNAs were extracted using a hot borate methodescribed in our previous report (Yao et al., 2007). Arabidopsis total
turae 145 (2012) 94–101 95
RNAs were extracted using Trizol reagent (Invitrogen, Carlsbad,USA). Total RNAs were treated with RNase free DNase. Two micro-grams of total RNAs were used to synthesize first-strand cDNAusing PrimeScript First Strand cDNA Synthesis Kit (Takara, Liaon-ing, China). Real-time quantitative RT-PCR was conducted usingcDNA templates to detect the expression level of MdVHA-B in apple.Apple 18S rRNA was used as loading controls. The specific primerssequences (Table S1) were used for PCR analysis. Fluorescence-quantitative PCR reactions were repeated three times.
2.4. Transformation of MdVHA-B into tomato plants
The MdVHA-B ORF was obtained by PCR, primers see Table S1.The forward primer contained a BamHI digestion site, and thereverse primer contained a SalI site (both underlined). The ORFregion of the cDNA was used for sense overexpression. The PCRproducts were digested with BamHI/SalI and cloned into the binaryvector pBI121, downstream of the CaMV 35S promoter. The con-structs were introduced into Agrobacterium strain LBA4404 andtransformed into tomato leaves as described by Holsters et al.(1978).
2.5. Relative electronic conductivity (Rec)
Fresh leaves, 0.4 g, were immersed in 4 ◦C and 40 ◦C for 4 h,were then separated into two equal groups. The first group wasshaken in 5 mL double-distilled water at 170 rpm for 2 h at 25 ◦C,the second group was boiled for 30 min. The relative electronic con-ductivity was measured as described by Tang (1999) using a DDS-12conductometer (Hangzhou Wanda Instrument Factory, China). Theconductivity in test leaves was desinated as Rc and boiled leaves asRc’. The relative conductivity was calculated by Rc/Rc’ × 100%.
2.6. Enzyme extraction and activity assays
The isolation of tonoplast membranes and VHA-B activityassays were carried out as described by Terrier et al. (2001) withminor modifications. Ten grams of fresh tissue were crushed ina buffer composed of 0.2% bovine serum albumin (BSA) (w/v),1% defatted casein (w/v), 1% polyvinyl pyrrolidone (PVP) (w/v),5 mM dithiothreitol (DTT), 5 mM ethylene diamine tetraacetic acid(EDTA), 0.25 M glucose, 0.25 M fructose, 1 mM phenylmethanesul-fonyl fluoride (PMSF), 20 mM �-mercaptoethanol, and adjustedto pH 8.2 with ascorbic acid. After filtration on a 100 �m nylonmesh, high-density organelles were removed by centrifugation at8000 × g for 20 min, and the resulting supernatant was sedimentedat 100,000 × g for 40 min. The pellet was washed by resuspen-sion in 0.3 M sorbitol, 0.2% bovine serum albumin (BSA) (w/v),0.5% polyvinyl pyrrolidone (PVP) (w/v), 1 mM ethylene diaminetetraacetic acid (EDTA), 0.2 M KCl, 5 mM dithiothreitol (DTT), 1 mMphenylmethanesulfonyl fluoride (PMSF), 25 mM bis-tris-propane-2-(N-morpholine)-ethanesulphonic acid (BTP-Mes) (pH 7.5) andthen centrifuged at 100,000 × g for 40 min. Tonoplast vesicles werepurified by floating the resuspended pellet on a 0%/30% discontinu-ous sucrose gradient containing 5 mM DTT and 5 mM BTP-Mes (pH7.5) for 2 h at 100,000 × g. The interface was collected and washedby centrifugation at 100,000 × g for 40 min in 0.3 M mannitol,0.5 mM EDTA, 5 mM DTT, 10 mM BTP-Mes (pH 7.5). The resultingpellet was resuspended in 20% glycerol (v/v), 0.5 mM EDTA, 10 mMBTP-Mes (pH 7.5) and stored in liquid nitrogen. VHA activity wasmeasured in the reaction mixture comprised of 50 mM KCl, 3 mM
MgSO4, 1 mM Na2MoO4, 0.25 mM Na3VO4, 2.5 mM ATP-Na2, 0.02%(w/v) TritonX-100, 10 mM BTP-Mes (pH 7.5), and 100 �L of extract.The Pi liberated was determined according to Ames (1966) after30 min incubation at 30 ◦C.
96 D.-G. Hu et al. / Scientia Horticulturae 145 (2012) 94–101
Fig. 1. Analysis of the deduced amino acid sequence of MdVHA-B. (a) Schematic diagram of the MdVHA-B genomic structure. The genomic structure prediction was performeda dVHAm
2
H114mroUwF(v
2
eiwarasJdcYeo
t the website http://gsds.cbi.pku.edu.cn/index.php. (b) Phylogenetic tree of the Method with the amino acid sequences of MdVHA-B and VHA-Bs in other species.
.7. Determination of the malondialdehyde (MDA) level
The MDA content was determined according to the method ofodges et al. (1998). The tomato leaves were homogenized in 5 mL0% trichloroacetic acid (TCA) and centrifuged at 12,000 × g for0 min. A volume of 2 mL of the clear supernatant was added to
mL of 0.6% thiobarbituric acid (TBA, in 10% TCA), and the reactionixture was incubated at 100 ◦C in a water bath for 15 min. The
eaction was terminated at room temperature, and the absorbancef the supernatant at 450, 532 and 600 nm was determined using aV–vis spectrophotometer (UV-2450). The concentration of MDAas calculated by the following formula: MDA content (�mol g−1
W) = CMDA (�mol L−1) × V (mL)/fresh weight (g FW), where CMDA�mol L−1) = 6.45 (OD532 − OD600) − 0.56OD450 and V refers to theolume (mL) of extraction solution.
.8. Proline content determination
The free proline content was determined as described by Batest al. (1973). Proline was extracted from tomato leaves by grind-ng the leaves in 5 mL 3% sulfosalicylic acid; 2 mL of the extract
as then reacted with 3 mL acid ninhydrin and 2 mL glacial aceticcid for 40 min at 100 ◦C. An ice bath was used to terminate theeaction. The reaction mixture was extracted with 5 mL toluenend vortexed. The absorbance of the toluene layer was mea-ured at 520 nm using a spectrophotometer (Shimadzu UV-2450,apan). The proline concentration was determined using a stan-ard curve and calculated on an FW basis as follows: proline
ontent (�g g−1 FW) = YV1 (mL)/V2 (mL) fresh weight (g FW), where= 21.594OD520 − 0.0785, V1 refers to the volume (mL) of totalxtraction solution and V2 refers to the measurement of the volumef extraction solution.
-B protein. The tree was constructed using MEGA 4.0 by the Neighbor-Joining (NJ)
2.9. Relative water loss rate
Transgenic and WT tomato plants were grown in the samepots under normal conditions for 10 weeks. The leaves were thendetached at the cotyledonary node, and the relative water loss ratewas measured as described by Dhanda and Sethi (1998).
3. Results
3.1. Isolation and sequence analysis of the MdVHA-B gene inapple
The full-length cDNA of MdVHA-B from the apple (M. domes-tica B.) cultivar ‘Gala’ was obtained using RT-PCR and RACE. TheMdVHA-B cDNA contains a 1473-bp-long open reading frame (ORF)that encodes 491 amino acid residues. The genome sequence(MDP0000945182) of MdVHA-B was obtained by searching (BLAST)the apple genome database with the MdVHA-B cDNA sequence. Thesequence was subsequently confirmed by PCR amplification andsequencing. The results showed that there are 13 introns and 14exons in the MdVHA-B ORF (Fig. 1a).
In addition, according to alignment of apple genome sequence,we found 2 sequences (MDP0000205940 and MDP0000631168)that similar to our cloning sequence MdVHA-B (MDP0000945182).And there are three isoforms for the VHA-B subunit (VHA-B1, -B2,and -B3) in Arabidopsis. Maybe the number of VHA-B subunit inapple is same to Arabidopsis. As MdVHA-B (MDP0000945182) wascloned in our study, so focus on it only.
To investigate the evolutionary context of the plant VHA-B
proteins, a phylogenetic tree of the VHA-B proteins (Fig. 1b) wasconstructed using the online software MEGA4.1 (http://www.megasoftware.net/). The results showed that the deduced MdVHA-B protein exhibits the highest sequence similarity (96.19% identity)
D.-G. Hu et al. / Scientia Horticulturae 145 (2012) 94–101 97
Fig. 2. Expression analysis of the MdVHA-B gene using real-time quantitative PCR. (a) The expression levels of MdVHA-B in apple young leaves (growth for 10 d), old leaves(growth for 60 d), stems (growth for 30 d), flowers (3 d after blooming) and fruits (60 d after bearing fruit). (b–f) The expression pattern of MdVHA-B under abiotic stress: (b)1 00 mM
tV
asAMA8BA
3r
eitMl(
wtwa(rh
00 �M ABA; (c) low temperature 4 ◦C; (d) high temperature 40 ◦C; (e) 2% PEG; (f) 1
o Arabidopsis AtVHA-B1 and that it is also closely analogous toHA-B proteins from other plant species (Fig. 1b).
There are three isoforms for the VHA-B subunit (VHA-B1, -B2,nd -B3) in Arabidopsis (Sze et al., 2002). Our phylogenetic analy-is of VHA-B proteins, including apple MdVHA-B and ArabidopsistVHA-B1, -B2, and -B3, is shown in Fig. S1, demonstrating thatdVHA-B is highly similar to AtVHA-B1 (96.19% identity) and
tVHA-B3 (96.27% identity). In contrast, MdVHA-B showed only3.58% similarity to the AtVHA-B2 protein. Therefore, MdVHA-
is the apple counterpart of AtVHA-B1 or AtVHA-B3 but nottVHA-B2.
.2. Expression analysis of MdVHA-B in different tissues and inesponse to abiotic stresses
Real-time quantitative RT-PCR was performed to determine thexpression patterns of MdVHA-B in different tissues. The resultndicated that MdVHA-B was constitutively expressed in all ofhe tissues tested; however, its expression level was variable.dVHA-B transcripts were expressed strongly in fruits and young
eaves but were relatively weak in old leaves, flowers and stemsFig. 2a).
MdVHA-B expression in response to different abiotic stressesas examined in apple in vitro shoot cultures. The result of real-
ime quantitative RT-PCR showed that the expression of MdVHA-Bas induced by abiotic stresses, including ABA (100 �M), low (4 ◦C)
nd high (40 ◦C) temperatures, PEG (2%) and salinity treatment100 mM NaCl) (Fig. 2b–f). Therefore, it is suggested that MdVHA-Besponds to abiotic stress, including ABA, salt, drought and cold andeat stresses.
NaCl.
3.3. The response of seed germination to PEG and ABA intransgenic tomato ectopically expressing MdVHA-B
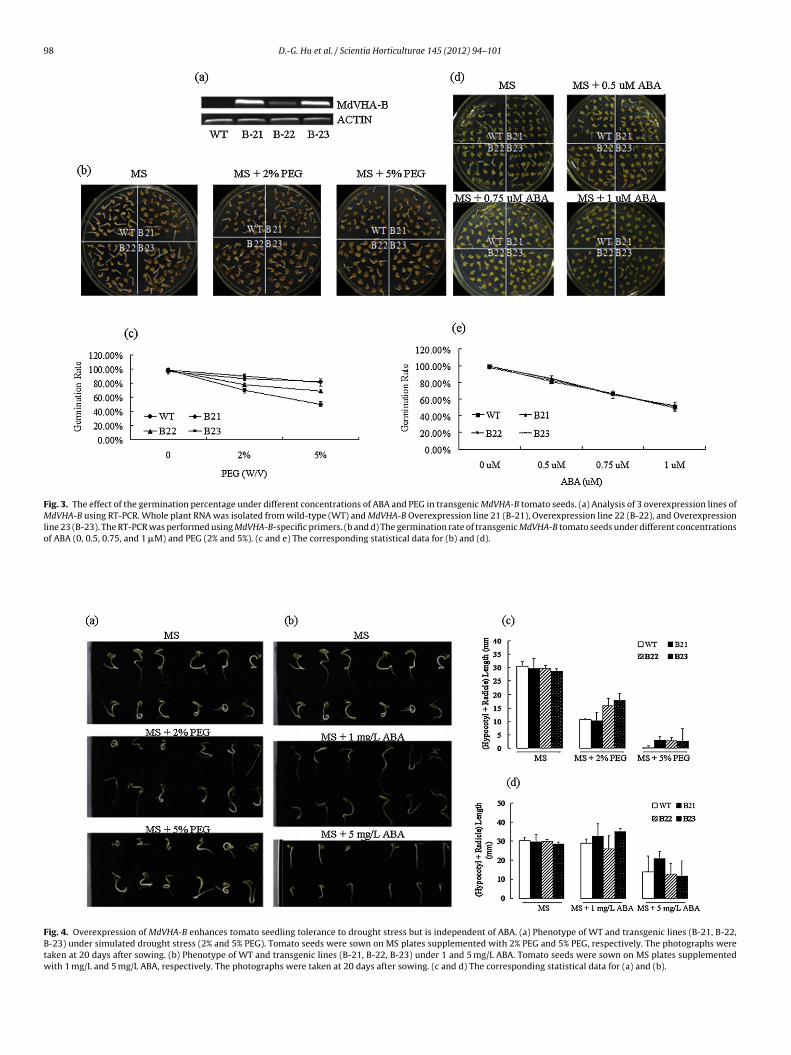
To characterize the in planta function of MdVHA-B, a plantexpression construct 35S::MdVHA-B was introduced into wild-typeplants (WT) tomato. Three independent homozygous transgeniclines, B-21, B-22 and B-23, were chosen for the functional analy-sis. RT-PCR analysis indicated that all three of the transgenic linesproduced markedly more MdVHA-B transcript than the WT con-trol, indicating that the MdVHA-B gene was ectopically expressedin those transgenic tomatoes (Fig. 3a). Subsequently, the sensitiv-ity of seed germination to ABA and PEG was determined in threeof the MdVHA-B transgenic tomato lines. The results showed thatthe 3 transgenic lines exhibited much higher germination ratesthan the WT control on MS medium containing a high concen-tration of PEG (Fig. 3b and c). Therefore, the ectopic expression ofMdVHA-B enhanced the tolerance to PEG treatment in the trans-genic tomatoes. In contrast, the ectopic expression of MdVHA-Bexerted no influence on seed germination on MS medium contain-ing ABA when compared with the WT control (Fig. 3d and e). Takentogether, we found that seed germination in transgenic tomatoplants ectopically expressing MdVHA-B respond to PEG but mayindependent of ABA.
3.4. MdVHA-B transgenic tomato seedlings are tolerant to PEGand low or high temperature but not to ABA
Furthermore, 7-day-old transgenic tomato seedlings were usedto examine whether the ectopic expression of MdVHA-B improvesthe tolerance to PEG and ABA treatments. The results showed thatMdVHA-B transgenic seedlings exhibited similar phenotypes to the
98 D.-G. Hu et al. / Scientia Horticulturae 145 (2012) 94–101
Fig. 3. The effect of the germination percentage under different concentrations of ABA and PEG in transgenic MdVHA-B tomato seeds. (a) Analysis of 3 overexpression lines ofMdVHA-B using RT-PCR. Whole plant RNA was isolated from wild-type (WT) and MdVHA-B Overexpression line 21 (B-21), Overexpression line 22 (B-22), and Overexpressionline 23 (B-23). The RT-PCR was performed using MdVHA-B-specific primers. (b and d) The germination rate of transgenic MdVHA-B tomato seeds under different concentrationsof ABA (0, 0.5, 0.75, and 1 �M) and PEG (2% and 5%). (c and e) The corresponding statistical data for (b) and (d).
Fig. 4. Overexpression of MdVHA-B enhances tomato seedling tolerance to drought stress but is independent of ABA. (a) Phenotype of WT and transgenic lines (B-21, B-22,B-23) under simulated drought stress (2% and 5% PEG). Tomato seeds were sown on MS plates supplemented with 2% PEG and 5% PEG, respectively. The photographs weretaken at 20 days after sowing. (b) Phenotype of WT and transgenic lines (B-21, B-22, B-23) under 1 and 5 mg/L ABA. Tomato seeds were sown on MS plates supplementedwith 1 mg/L and 5 mg/L ABA, respectively. The photographs were taken at 20 days after sowing. (c and d) The corresponding statistical data for (a) and (b).

orticul
WeHtettAt
iaMWctedtccm
3a
saocgcet
dma
blucalpitwletfgtifig
aabtHd
D.-G. Hu et al. / Scientia H
T control on MS medium, indicating that MdVHA-B does not influ-nce the growth of tomato seedlings under normal conditions.owever, when those seedlings were exposed to PEG treatment,
he transgenic lines were much more tolerant to PEG treatment,specially at high concentrations of PEG (Fig. 4a and c). In con-rast, the lengths of the hypocotyls and roots of both the MdVHA-Bransgenic and WT seedlings were inhibited to similar extents byBA (Fig. 4b and d), indicating that the influence of the MdVHA-B
ransgene is independent of ABA.In addition, we noticed that the expression of MdVHA-B was
nduced by low (4 ◦C) and high (40 ◦C) temperatures in apple (Fig. 2cnd d) and we were interested to know whether overexpression ofdVHA-B enhanced the resistance of low and high temperature.e found that the leaves of transgenic plants did not significantly
url and wilt compared with wild-type plants under low and highemperature tolerance (Fig. S2a and c). Furthermore, the relativelectric conductivity was measured to reflect the level of cellularamage after low and high temperature treatment. As a result, theransgenic plants had much higher relative electric conductivityompared with wild-type plants, indicating cold and heat stressaused severe ion leakage in the wild-type plants, whereas wereuch less affected (Fig. S2b and d).
.5. MdVHA-B ectopic expression confers tolerance to drought vian ABA-independent manner
To confirm the function of MdVHA-B in the tolerance to droughttress, 40-day-old tomato plants were used for a drought tolerancessay. All of the transgenic plants only started to wilt after 2 weeksf drought treatment, whereas the WT control plants were almostompletely wilted. Following the rewatering for 10 h, the trans-enic lines recovered and began to grow, but most of the WT plantsontinued to wilt and even died (Fig. 5a), indicating that the ectopicxpression of MdVHA-B confers a tolerance to drought in transgenicomato plants.
To explain why the transgenic plants are more tolerant torought than the WT control, the relative water loss rate was deter-ined. The result indicated that the transgenic plants have a greater
bility to retain water than the WT plants (Fig. 5b).Furthermore, the level of proline or MDA reflected the mem-
rane damage in some extent. And it was found that the transgenicines produced the same level of proline and MDA as the WT controlnder normal conditions (Fig. 5c and d). However, under droughtonditions, the tomato seedlings produced proline and MDA inmounts that varied with their MdVHA-B expression levels. Theevel of MdVHA-B transcripts was positively correlated with prolineroduction but negatively correlated with MDA accumulation. That
s to say, the transgenic lines produced more proline but less MDAhan the WT control under drought stress (Fig. 5c and d). Mean-hile, to further identify the membrance damage degree of these
ines, we detected the ion leakage. As shown in Fig. S3, the relativelectronic conductivity of transgenic lines is lower than WT con-rol. These results indicated that a high level of MdVHA-B transcriptacilitated the production of more protective proline in the trans-enic plants than the WT control under drought. As a result, theransgenic lines suffered less damage in the membrane system, asndicated by the lower MDA accumulation. Taken together, thesendings demonstrated that the ectopic MdVHA-B-expressing trans-enic lines were more tolerant to drought stress than the control.
Moreover, we detected the total H+-ATPase activity in the WTnd transgenic plants, and the results showed that the H+-ATPasectivity of the transgenic plants was higher than the WT control
oth under normal and drought conditions (Fig. 5e). Therefore,he increased MdVHA-B expression and the resulting enhanced+-ATPase activity are positively correlated with the tolerance torought in the transgenic tomatoes.turae 145 (2012) 94–101 99
Lastly, RT-PCR was performed to determine the expression ofABA and drought-related genes in the WT and MdVHA-B trans-genic plants. The results showed that the transcript level of theABA-related gene ABI2 was similar in the transgenic plants andthe WT control. In contrast, the transgenic plants produced sig-nificantly more transcripts of drought-inducible genes, includingERF3b, ERF5 and DREB3, compared with the WT control (Fig. 5f).Therefore, VHA-B may be involved in drought tolerance indepen-dent of ABA pathway.
4. Discussion
Vacuolar H+ pumps play key roles in the maintenance of the H+
electrochemical gradient across the vacuolar membrane in plantstress responses. In this study, a gene encoding a V-H+-ATPase,MdVHA-B, was isolated from apple. Its expression was significantlyinduced by high salinity, drought, low and high temperature andABA application (Fig. 2), which is highly similar to that of MdVHP1 inapple (Dong et al., 2011). Generally, proton pump genes are inducedby various abiotic stresses, and it has been reported that the VHA-Bgene is induced in response to abiotic stresses, such as high salinity,in various plants (Kluge et al., 2003a; Kader et al., 2006). Simi-larly, the V-ATPase activity is considered to be regulated at thetranscriptional level in response to abiotic stresses. For example,the expression of V-ATPase subunit genes is up-regulated undersalt stress (Maeshima, 2001; Kluge et al., 2003b), and the ectopicexpression of MdVHA-B enhanced the V-ATPase activity in tomato(Fig. 5e). These results further prove the relationship between thetranscriptional regulation of V-ATPase genes and V-ATPase activity.
The ectopic expression of a vacuolar H+-pyrophosphatase genefrom the halophyte Suaeda corniculata could improve salt, saline-alkali and drought tolerance in Arabidopsis (Liu et al., 2011). Asa regulatory subunit, the B subunit, participates in the formationof the catalytic site, and the existence of multiple isozymes indi-cates the complexity and importance of its regulatory role, whichis reflected in the changes in the expression of B subunits under saltstress (Li et al., 2004). In addition, it is reported that expression ofthe wheat vacuolar H+-ATPases (V-ATPase) subunit B (TaVB) geneenhanced plant tolerance to salt stress (Wang et al., 2011). Theseresults imply that the proton pump gene MdVHA-B is also involvedin the tolerance responses of plants to various stresses.
In Arabidopsis, numerous genes that respond to dehydrationstress have been identified and categorized as responsive to dehy-dration (rd) and early response to dehydration (erd) genes (Shinozakiand Yamaguchi-Shinozaki, 1996). There are at least four inde-pendent regulatory systems for gene expression in response towater-deficit stress. Two are abscisic acid (ABA)-dependent andABA-independent (Shinozaki and Yamaguchi-Shinozaki, 2000).ABA plays a crucial role during plant growth and developmentand responses to environmental stresses in plants (Xiong et al.,2001). ABA is defined as a stress hormone due to its rapid accumu-lation in response to stresses. Subsequently, its accumulation hasa highly protective role that confers osmotic adjustment, togetherwith an increase in the concentrations of other osmolytes in plants(Valliyodan and Nguyen, 2006).
With regard to the relationship between ABA and H+-ATPaseactivity, a recent study shows that the exogenous applicationof ABA on leaves has an inhibitory effect on the PM H+-ATPaseactivity (Zhang et al., 2004), whereas AHA1 (ost2) mutations inthe plasma membrane H+-ATPase generate a constitutively activeprotein with reduced sensitivity to ABA during stomatal move-
ments (Merlot et al., 2007). However, our findings indicated thatthe increased drought resistance due to the ectopic expressionof MdVHA-B might be independent of ABA in transgenic tomato.It is well known that dehydration activates ABA-dependent and
100 D.-G. Hu et al. / Scientia Horticulturae 145 (2012) 94–101
Fig. 5. MdVHA-B overexpression confers improved drought tolerance that is independent of ABA in tomato. (a) The drought tolerance of transgenic plants and WT. The5-week-old transgenic plants (T1 generation) and WT were not watered for 2 weeks and then watered, and photographs were taken after 10 h. (b) The relative water loss ratein MdVHA-B transgenic and WT tomato leaves. (c–e) The free proline, MDA and H+-ATPase activity in transgenic tomato leaf and WT tomato were measured under normalconditions, drought treatment and recovery. The results were the mean value of 3 experiments. Means and standard deviation of Rec of 9 replicates from 3 experiments. (f)The relative expression of ABA-dependent (SlABI2)- and ABA-independent (SlERF3b, SlERF5 and SlDREB3)-related genes in transgenic tomato.

orticul
AbM(2cvd
udat
A
cH(T
A
f2
R
A
A
B
B
B
C
C
C
D
D
D
D
H
D.-G. Hu et al. / Scientia H
BA-independent gene expression systems involving ABF (ABRE-inding factor)/AREB (ABA-responsive element binding protein),YC/MYB, DREB2 (drought–responsive element binding) and NAC
NAM, A TAF1, 2 and CUC) transcription factors (Agarwal and Jha,009). In addition, all of these H+-ATPase genes, which are asso-iated with ABA, are located in the PM, whereas MdVHA-B is aacuolar-type H+-ATPase. It is possible that MdVHA-B increasesrought resistance by an ABA-independent pathway.
In summary, the enhanced expression of the V-H+-ATPase sub-nit B gene under drought stress is an important factor in therought tolerance of some plants. In the present case, the droughtssociation of MdVHA-B demonstrates the potential use of theseransgenic plants for future agricultural use.
cknowledgements
This work was supported by the National Public Benefit (Agri-ultural) Research Foundation of China (200903044), Nationaligh Technology Research and Development Program of China
2011AA100204) and Changjiang Scholars and lnnovative Researcheam in University (IRT1155).
ppendix A. Supplementary data
Supplementary data associated with this article can beound, in the online version, at http://dx.doi.org/10.1016/j.scienta.012.08.010.
eferences
garwal, P.K., Jha, B., 2009. Transcription factors in plants and ABA dependent andindependent abiotic stress signaling. Biol. Plant. 54, 201–212.
mes, B.N., 1966. Assay of inorganic phosphate, total phosphate and phosphatase.Method Enzymol. 8, 115–118.
ageshwar, U.K., Taneja-Bageshwar, S., Moharram, H.M., 2005. Two isoforms ofthe A subunit of the vacuolar H(+)-ATPase in Lycopersicon esculentum: highlysimilar proteins but divergent patterns of tissue localization. Planta 220,632–643.
ates, L.S., Waldren, R.P., Teare, I.D., 1973. Rapid determination of free proline forwater-stress studies. Plant Soil 39, 205–207.
rini, F., Hanin, M., Mezghani, I., 2007. Overexpression of wheat Na+/H+ antiporterTNHX1 and H+-pyrophosphatase TVP1 improve salt and drought stress tolerancein Arabidopsis thaliana plants. J. Exp. Bot. 58, 301–308.
arter, C., Pan, S.Q., Zouhar, J., 2004. The vegetative vacuole proteome of Arabidopsisthaliana reveals predicted and unexpected proteins. Plant Cell 16, 3285–3303.
ho, Y.H., Yoo, S.D., Sheen, J., 2006. Regulatory functions of nuclear hexokinase1complex in glucose signaling. Cell 127, 579–589.
ipriano, D.J., Wang, Y.R., Bond, S., Forgac, M., 2008. Structure and regulation of thevacuolar ATPases. Biochim. Biophys. Acta 1777, 599–604.
handa, S.S., Sethi, G.S., 1998. Inheritance of excised-leaf water loss and relativewater content in bread wheat (Triticum aestivum). Euphytica 104, 39–47.
ietz, K.J., Tavakoli, N., Kluge, C., 2001. Significance of the V-type ATPase for theadaptation to stressful growth conditions and its regulation on the molecularand biochemical level. J. Exp. Bot. 52, 1969–1980.
ong, Q.L., Liu, D.D., Hao, Y.J., 2011. MdVHP1 encodes an apple vacuolar H+-PPaseand enhances stress tolerance in transgenic apple callus and tomato. J. PlantPhysiol. 168, 2124–2133.
oyle, J.J., Doyle, J.L., 1990. Isolation of plant DNA from fresh tissue. Focus 12,552–555.
odges, D.M., John, M., De, L., 1998. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containinganthocyanin and other interfering compounds. Planta 207, 604–611.
turae 145 (2012) 94–101 101
Holsters, M., Waele, D., Depicker, A., 1978. Transfection and transformation ofAgrobacterium tumefaciens. Mol. Gen. Genet. 163, 181–187.
Jaquinod, M., Villiers, F., Kieffer-Jaquinod, S., 2007. A proteomics dissection of Ara-bidopsis thaliana vacuoles isolated from cell culture. Mol. Cell. Proteomics 6,394–412.
Kader, M.A., Seidel, T., Lindberg, S., 2006. Expressions of OsHKT1, OsHKT2, and OsVHAare differentially regulated under NaCl stress in salt-sensitive and salt-tolerantrice (Oryza sativa L.) cultivars. J. Exp. Bot. 57, 4257–4268.
Klein, U., 1992. The insect V-ATPase, a plasma membrane proton pump energizingsecondary active transport: immunological evidence for the occurrence of a V-ATPase in insect ion-transporting epithelia. J. Exp. Biol. 172, 345–354.
Kluge, C., Lahr, J., Hanitzsch, M., 2003a. New insight into the structure and regulationof the plant vacuolar H+-ATPase. J. Bioenerg. Biomembr. 35, 377–388.
Kluge, C., Lamkemeyer, P., Tavakoli, N., 2003b. cDNA cloning of 12 subunits of the V-type ATPase from Mesembryanthemum crystallinum and their expression understress. Mol. Membr. Biol. 20, 171–183.
Li, J., Yang, H., Peer, W.A., Richter, G., 2005. Arabidopsis H+-PPase AVP1 regulatesauxin-mediated organ development. Science 310, 121–125.
Li, P.H., Wang, Z.L., Wang, B.S., 2004. Cloning and expression analysis of the B subunitof V-H+-ATPase in leaves of Halophyte Suaeda salsa under salt stress. Acta Bot.Sin. 46, 93–99.
Liu, L., Wang, Y., Li, H.Y., 2011. Cloning of a vacuolar H+-pyrophosphatase gene fromthe Halophyte Suaeda corniculata whose heterologous overexpression improvessalt, saline-alkali and drought tolerance in Arabidopsis. J. Integr. Plant Biol. 53,731–742.
Lüttge, U., Wang, B.S., Ratajczak, R., 2001. Effects of salt treatment and osmotic stresson V-ATPase and V-PPase in leaves of the halophyte Suaeda salsa. J. Exp. Bot. 52,2355–2365.
Maeshima, M., 2001. Tonoplast transporters: organization and function. Annu. Rev.Plant Physiol. Plant Mol. Biol. 52, 469–497.
Merlot, S., Leonhardt, N., Leung, J., 2007. Constitutive activation of a plasma mem-brane H+-ATPase prevents abscisic acid-mediated stomatal closure. EMBO J. 26,3216–3226.
Padmanaban, S., Lin, X., Sze, H., 2004. Differential expression of vacuolar H+-ATPasesubunit c genes in tissues active in membrane trafficking and their roles in plantgrowth as revealed by RNAi. Plant Physiol. 134, 1514–1526.
Park, S., Li, J., Gaxiola, R.A., 2005. Up-regulation of an H+-pyrophosphatase (H+-PPase)as a strategy to engineer drought resistant crop plants. Proc. Natl. Acad. Sci. U.S.A.102, 18830–18835.
Shiratake, K., Martinoia, E., 2007. Transporters in fruit vacuoles. Plant Biotechnol.24, 127–133.
Shinozaki, K., Yamaguchi-Shinozaki, K., 1996. Molecular responses to drought andcold stress. Curr. Opin. Biotechnol. 7, 161–167.
Shinozaki, K., Yamaguchi-Shinozaki, K., 2000. Molecular responses to dehydrationand low temperature: differences and cross-talk between two stress signalingpathways. Curr. Opin. Plant Biol. 3, 217–223.
Sze, H., Schumacher, K., Taiz, L., 2002. A simple nomenclature for a complex pro-ton pump: VHA genes encode the vacuolar H+-ATPase. Trends Plant Sci. 7,157–161.
Tang, Z.C., 1999. Modern Experiment Procotols in Plant Physiology. Science Press,Beijing, pp. 302–308.
Terrier, N., Sauvage, F.X., Romieu, C., 2001. Changes in acidity and in protontransport at the tonoplast of grape berries during development. Planta 213,20–28.
Valliyodan, B., Nguyen, H.T., 2006. Understanding regulatory networks and engi-neering for enhanced drought tolerance in plants. Curr. Opin. Plant Biol. 9,189–195.
Wang, L., He, X., Zhao, Y., Shen, Y., Huang, Z., 2011. Wheat vacuolar H+-ATPasesubunit B cloning and its involvement in salt tolerance. Planta 234, 1–7.
Wu, A.M., Liu, J.Y., 2006. An improved method of genomoc walking for promotersequences cloning. Chin. J. Biochem. Mol. Biol. 22, 243–246.
Xiong, L.M., Lee, H., Zhu, J.K., 2001. The Arabidopsis LOS5/ABA3 locus encodes amolybdenum cofactor sulfurase and modulates cold stress- and osmotic stress-responsive gene expression. Plant Cell 13, 2063–2083.
Yao, Y.X., Li, M., Zhai, H., 2007. A novel gene, screened by cDNA-AFLP approach,contributes to lowering the acidity of fruit in apple. Plant Physiol. Biochem. 45,
139–145.Zhang, X., Wang, H., Shimazaki, K., 2004. Inhibition of blue light-dependent H+
pumping by abscisic acid through hydrogen peroxide-induced dephosphoryla-tion of the plasma membrane H+-ATPase in guard cell protoplasts. Plant Physiol.136, 4150–4158.