overexpression of human aspartyl (asparaginyl) b...
TRANSCRIPT

[CANCER RESEARCH 60, 1261–1266, March 1, 2000]
Overexpression of Human Aspartyl (Asparaginyl)b-Hydroxylase Is Associated withMalignant Transformation 1
Nedim Ince, Suzanne M. de la Monte, and Jack R. Wands2
Molecular Hepatology Laboratory, Massachusetts General Hospital Cancer Center, Departments of Medicine and Pathology, Harvard Medical School, Charlestown,Massachusetts 02129
ABSTRACT
The human aspartyl (asparaginyl) b-hydroxylase (HAAH) is a highlyconserved enzyme that hydroxylates epidermal growth factor-like do-mains in transformation-associated proteins. We previously reportedoverexpression of theHAAH gene in human hepatocellular carcinomasand cholangiocarcinomas (L. Lavaissiereet al., J. Clin. Investig., 98:1313–1323, 1996). In the present study, we determined whether HAAHprotein overexpression was linked to cellular proliferation or malignanttransformation of bile ducts by using a human disease and rat model ofbile duct proliferation. In addition, the transforming properties of theAAH genes were assessed by transient and stable transfection of NIH-3T3cells with human and murine wild-type as well as mutant cDNA constructsthat lacked hydroxylation activity. Cellular characteristics of the malig-nant phenotype were assessed by formation of transformed foci, growth insoft agar, and tumor development in nude mice. We found thatHAAHgene expression was undetectable during bile duct proliferation in bothhuman disease and rat models as compared with cholangiocarcinoma.Overexpression of HAAH in NIH-3T3 cells was associated with generationof a malignant phenotype, and enzymatic activity was required for cellulartransformation. These findings suggest that overexpression of HAAH islinked to cellular transformation of biliary epithelial cells.
INTRODUCTION
To identify novel molecules that are specifically overexpressed intransformed malignant cells of human hepatocyte origin, the FOCUSHCC3 cell line was used as an immunogen to generate mAbs thatspecifically or preferentially recognize proteins associated with themalignant phenotype (1). AgGT11 cDNA expression library derivedfrom HepG2 HCC cells was screened and the FB-50 mAb producedagainst the FOCUS cell line was found to recognize an epitope on aprotein encoded by an AAH (HAAH) cDNA. The HAAH enzyme hassubsequently been found to be up-regulated in several different humantransformed cell lines and tumor tissues as compared with the adjacenthuman tissue counterpart. More important was the finding that theoverexpressed enzyme in different human malignant tissues was cat-alytically active (2).
The HAAH is a protein belonging to thea-ketoglutarate-dependentdioxygenase family of prolyl and lysyl hydroxylases, which play a keyrole in collagen biosynthesis. This molecule hydroxylates asparticacid or asparagine residues in certain EGF-like domains of severalproteins in the presence of ferrous iron. These EGF-like domainscontain conserved motifs that form repetitive sequences in diverseproteins, such as clotting factors, extracellular matrix proteins, low-density lipoprotein receptor, Notch homologues or Notch ligand ho-mologues (3–5). It is believed that EGF-like sequences play an im-
portant role in protein-protein interactions, as shown by mutations inEGF-like domains of fibrillin that cause Marfan’s syndrome or factorIX, which produces hemophilia B (6, 7).
In this study,HAAHgene expression was examined in proliferatingbile ducts because our previous findings demonstrated that this genewas overexpressed in 100% of human cholangiocarcinomas. We alsodetermined whether overexpression of HAAH in NIH-3T3 cells led tothe generation of the malignant phenotype, as measured by the for-mation of transformed foci, growth in soft agar as an index ofanchorage independent growth, and tumor formation in nude mice.Furthermore, we explored the role of enzymatic activity in the induc-tion of transformed phenotype by using a cDNA construct with amutation in the catalytic site that abolished hydroxylase activity, asdescribed previously (8). Taken together, our results are consistentwith the hypothesis that overexpression of theHAAH gene is associ-ated with malignant transformation of bile ducts.
MATERIALS AND METHODS
Antibodies. The FB-50 mAb was generated by cellular immunization ofBalb/C mice with FOCUS HCC cells. A monoclonal anti-Dengue virus anti-body served as a nonrelevant control. The HBOH2 mAb that was generatedagainst aMr 52,000 recombinant protein derived from an HAAH cDNA, whichrecognizes the catalytic domain ofb-hydroxylase from mouse and humanproteins as well as polyclonal anti-HAAH antibodies that cross-react with rathydroxylase protein, were a gift from DuPont Research Laboratories. Anti-Erk-1 was purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA).Sheep antimouse and donkey antirabbit antisera labeled with horseradishperoxidase were obtained from Amersham (Arlington Heights, IL).
Constructs. The murine full-length AAH construct (pNH376) and thesite-directed mutation construct (pNH376-H660) with abolished catalytic ac-tivity were cloned into the eukaryotic expression vector pcDNA3 (InvitrogenCorp., San Diego, CA), the full-length HAAH was cloned into prokaryoticexpression vector pBC-SK1 (Stratagene, La Jolla, CA), and these constructswere a generous gift of Dr. Joseph Dinchuk (DuPont Pharmaceuticals, Wilm-ington, DE). The full-length human AAH construct has been described previ-ously (2), and it was subcloned into theEcoRI site of the pcDNA3 vector. Theprotein sequence homology between murine and human AAH is 80%. ThepLNCX-UP1 construct that encodes v-src has been described previously (9).
Animal Model of Bile Duct Proliferation. Rats were divided into nineseparate groups of three animals each except for group 9, which contained fiverats. Group 1 served as the nonsurgical and group 2 as sham-operated surgicalcontrols. The remaining groups underwent common bile duct ligation to induceintrahepatic bile duct proliferation and were evaluated at 6, 12, 24, and 48 hand 4, 8, and 16 days, as shown in Table 1. Animals were asphyxiated withCO2, and liver samples were taken from left lateral and median lobes, fixed in2% paraformaldehyde, and embedded in paraffin. Liver samples (5mm) werecut and stained with H&E to evaluate intrahepatic bile duct proliferation.Immunohistochemistry was performed with polyclonal anti-HAAH antibodiesthat cross-react with the rat protein to determine levels of protein expression.
Bile Duct Proliferation Associated with PSC. Liver biopsy samples wereobtained from seven individuals with PSC and associated bile duct prolifera-tion. These individuals have been evaluated in the Division of Gastroentero-hepatology of Istanbul Medical School (Istanbul University, Istanbul, Turkey).Patients were 22–46 years of age, four males and three females. Four hadassociated inflammatory bowel disease (three ulcerative colitis and oneCrohn’s colitis). All patients underwent a radiological evaluation includingabdominal ultrasonography and endoscopic retrograde cholangiopancreaticog-
Received 7/28/99; accepted 1/5/00.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby markedadvertisementin accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1 Supported in part by NIH Grants CA-35711 and AA-02666.2 To whom requests for reprints should be addressed, at The Liver Research Center,
Rhode Island Hospital and Brown University School of Medicine, 55 Claverick Street,Providence, RI 02903. Phone: (401) 444-2795; Fax: (401) 444-2939; Email: [email protected].
3 The abbreviations used are: HCC, hepatocellular carcinoma; HAAH, human aspartyl(asparaginyl)b-hydroxylase; EGF, epidermal growth factor; mAb, monoclonal antibody;PSC, primary sclerosing cholangitis.
1261
on July 14, 2018. © 2000 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

raphy to exclude the diagnosis of extrahepatic biliary obstruction. Tissuesections were prepared from paraffin-embedded blocks and were evaluated byH&E staining for bile duct proliferation. Expression of HAAH was determinedby immunohistochemistry using the FB-50 mAb (2).
Immunohistochemistry. Liver tissue sections (5mm) were deparaffinizedin xylene and rehydrated in graded alcohol. Endogenous peroxidase activitywas quenched by a 30-min treatment with 0.6% H2O2 in 60% methanol.Endogenous biotin was masked by incubation with avidin-biotin blockingsolutions (Vector Laboratories, Burlingame, CA). The FB-50 mAb (for PSCsamples) and polyclonal anti-AAH-hydroxylase antibodies (for rat liver sam-ples) were added to slides in a humidified chamber at 4°C overnight. Immu-nohistochemical staining was performed using the avidin-biotin horseradishperoxidase complex (ABC) method using Vectastain kits with diaminobenzi-dine as the chromogen, according to the manufacturer’s instructions (VectorLaboratories, Inc.). Tissue sections were counterstained with hematoxylin,followed by dehydration in ethanol. Sections were examined by light micros-copy for bile duct proliferation and AAH protein expression. Paraffin sectionsof cholangiocarcinoma and placenta served as positive control (2), and hepa-tosteatosis samples served as a negative control. To control for antibodybinding specificity, adjacent sections were immunostained with the primaryantibody omitted or using nonrelevant antibody to Dengue virus (10). As apositive control for tissue immunoreactivity, adjacent sections of all specimenswere immunostained with monoclonal antibody to glyceraldehyde 3-phosphatedehydrogenase.
Western Blot Analysis. Cell lysates were prepared in RIPA buffer con-taining protease inhibitors as described previously (2). The total amount ofprotein in the lysates was determined by Bio-Rad colorimetric assay (Bio-Rad,Hercules, CA), followed by 10% SDS-PAGE and subsequent transfer topolyvinylidene difluoride membranes, and subjected to Western blot analysisusing FB-50, HBOH2, and anti-Erk-1 (used as an internal control for proteinloading) as primary antibodies and sheep antimouse and donkey antirabbitantisera labeled with horseradish peroxidase as secondary antibodies, as de-scribed (11). Antibody binding was detected with enhanced chemilumines-cence reagents (SuperSignal; Pierce Chemical Co., Rockford, IL) and filmautoradiography. The levels of immunoreactivity were measured by volumedensitometry using NIH Image software.
Enzymatic Activity Assay. AAH activity was measured in cell lysatesusing the first EGF-like domain of bovine protein S as substrate, where14C-labeleda-ketogluterate hydroxylates the domain, releasing14C-containingCO2, as described previously (4, 12, 13). Incubations were carried out at 37°Cfor 30 min in a final volume of 40ml containing 48mg of crude cell extractprotein and 75mM EGF substrate.
Cell Transfection Studies. The NIH-3T3 cells were cultured in DMEM(Mediatech, Washington, DC) supplemented with 10% heat-inactivated FCS(Sigma Chemical Co., St. Louis, MO), 1%L-glutamine, 1% nonessentialamino acids, and 1% penicillin-streptomycin (Life Technologies, Inc., GrandIsland, NY). Subconfluent NIH-3T3 cells (33 105 cells/60-mm dish) weretransfected with 10mg of one of the following plasmids: (a) nonrecombinantpcDNA3 vector (Invitrogen Corp., San Diego, CA) as a negative control; (b)pNH376-H660, the murine AAH cDNA was mutated at histidine position 660to lysine in the catalytic domain and cloned into the pcDNA3 vector driven bya cytomegalovirus promoter; (c) pNH376, the wild-type murine AAH cDNAcloned into the pcDNA3 vector; (d) pCDHH, wild-type human AAH cDNAcloned into the pcDNA3 vector; or (e) pLNCX-UP1, a cDNA that encodes
v-Src oncogene (positive control). Cells were transfected using the calciumphosphate transfection kit according to the manufacturer’s instructions (5Prime-3 Prime, Inc., Boulder, CO). Comparison of cellular transfection effi-ciency was assessed with the various constructs. For this procedure, confluentplates obtained 48 h after transfection were split and reseeded into 12 separate6-cm dishes, and 6 of them were made to grow in the presence of 400mg/mlG-418 (Life Technologies, Inc.)-containing medium. The number of G-418-resistant foci was determined at 14 days after transfection and used to correctfor any variability in transfection efficiency.
Transformation Assay. The NIH-3T3 cells were transfected with thevarious constructs and allowed to reach confluence after 48 h as describedabove. Each 6-cm dish was split and seeded into 12 different 6-cm dishes.While six of them were made to grow in the presence of G-418 to detecttransfection efficiency, the other six were grown in complete medium withoutG-418 and with a medium change every fourth day. The number of trans-formed foci were counted in these plates without G-418 and expressed astransformed foci/mg of transfected DNA. Transfection efficiency was cor-rected for as described above.
Anchorage-independent Cell Growth Assay.A limiting dilution tech-nique (0.15 cell/well of a flat-bottomed 96-well-plate) was performed ontransfectants grown in G-418 to isolate cell clones with different levels ofHAAH activity, as measured by Western blot analysis and enzymatic assay ofhydroxylase activity. Cloned cell lines (1.03 104 cells) were suspended incomplete medium containing 0.4% low-melting agarose (SeaPlaque GTGAgarose; FMC Bioproducts, Rockland, ME) and laid over a bottom agarmixture consisting of complete medium with 0.53% low-melting agarose. Eachclone was assayed in triplicate. The clones were seeded under these conditions,and 10 days later the size (positive growth.0.1 mm in diameter) and numberof foci were determined.
Tumorigenicity in Nude Mice. The same clones, as assessed in the an-chorage-independent growth assay, were injected into nude mice and observedfor tumor formation. Tumorigenicity was evaluated using 10 animals in eachof four groups (Charles River Laboratory, Wilmington, MA) as describedpreviously (14). Group 1 received 13 107 cells stably transfected with mockDNA, groups 2–4 received 13 107 cells of clones stably transfected withpNH376 and expressing various levels of murine AAH protein. Nude micewere kept under pathogen-free conditions in the animal facility of the Massa-chusetts General Hospital. Thirty days after tumor cell inoculation, the animalswere sacrificed using isofluorane (Aerrane, Anaquest, NJ)-containing cham-bers, and the tumors were carefully removed and the weights were determined.
RESULTS
Animal Model of Bile Duct Proliferation. After ligation of thecommon bile duct, intrahepatic bile duct proliferation was evident at48 h. Tissue samples obtained 8 and 16 days after common bile ductligation revealed extensive bile duct proliferation, as shown in Table1. Immunohistochemical staining failed to detect the presence of AAHin proliferating bile ducts at any time, as shown by the representativeexample depicted in Fig. 1,E andF. Analysis of AAH expression inbile ducts derived from sham surgical controls was also negative,whereas all samples exhibited positive immunoreactivity with anti-bodies to glyceraldehyde 3-phosphate dehydrogenase (data notshown). Thus, bile duct proliferation was not associated with in-creased HAAH expression in this animal model system.
The HAAH Expression in PSC. The liver biopsy specimens frompatients with PSC exhibited bile duct proliferation accompanied byperiductal fibrosis and a mononuclear inflammatory cell infiltrate, andwithout evidence of dysplasia (Fig. 1C). Adjacent sections immuno-stained with the FB-50 mAb had no detectable HAAH immunoreac-tivity in proliferating bile ducts (Fig. 1D). In contrast, sections ofcholangiocarcinoma that were immunostained simultaneously usingthe same antibody and detection reagents manifested intense levels ofHAAH immunoreactivity in nearly all tumor cells (Fig. 1A) as de-scribed previously, whereas adjacent sections of the cholangiocarci-nomas exhibited a negative immunostaining reaction with mAb to
Table 1 Bile duct proliferation and AAH expression in rat bile ducts at different timeintervals after common bile duct ligation
GroupSurgical
procedure MicroscopyaImmunohisto-
chemistry
1 No surgery Normal bile ducts Negative2 Sham surgery Normal bile ducts Negative3 6 hb Normal bile ducts Negative4 12 hb Normal bile ducts Negative5 24 hb Normal bile ducts Negative6 48 hb Minimal bile duct proliferation Negative7 4 daysb Moderate bile duct proliferation Negative8 8 daysb Extensive bile duct proliferation Negative9 16 daysb Extensive bile duct proliferation Negative
a Investigation performed under light microscopy after H&E staining.b After common bile duct ligation.
1262
HAAH AND MALIGNANT TRANSFORMATION OF BILE DUCTS
on July 14, 2018. © 2000 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
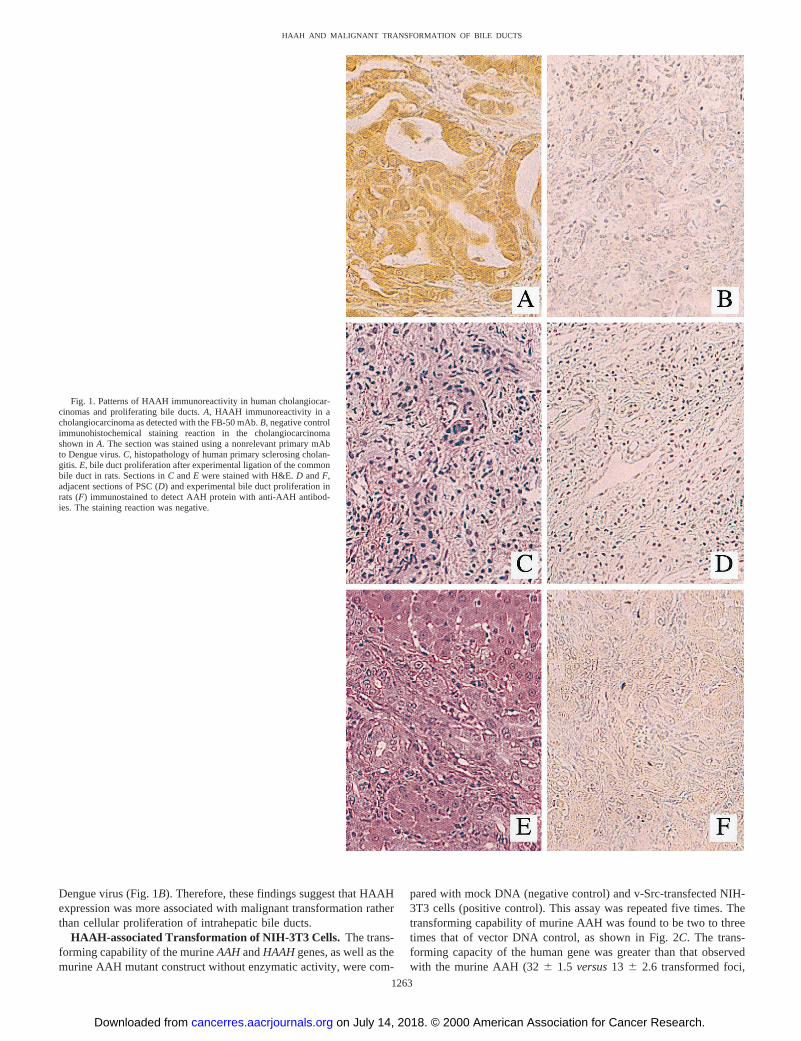
Dengue virus (Fig. 1B). Therefore, these findings suggest that HAAHexpression was more associated with malignant transformation ratherthan cellular proliferation of intrahepatic bile ducts.
HAAH-associated Transformation of NIH-3T3 Cells. The trans-forming capability of the murineAAHandHAAHgenes, as well as themurine AAH mutant construct without enzymatic activity, were com-
pared with mock DNA (negative control) and v-Src-transfected NIH-3T3 cells (positive control). This assay was repeated five times. Thetransforming capability of murine AAH was found to be two to threetimes that of vector DNA control, as shown in Fig. 2C. The trans-forming capacity of the human gene was greater than that observedwith the murine AAH (326 1.5 versus13 6 2.6 transformed foci,
Fig. 1. Patterns of HAAH immunoreactivity in human cholangiocar-cinomas and proliferating bile ducts.A, HAAH immunoreactivity in acholangiocarcinoma as detected with the FB-50 mAb.B, negative controlimmunohistochemical staining reaction in the cholangiocarcinomashown inA. The section was stained using a nonrelevant primary mAbto Dengue virus.C, histopathology of human primary sclerosing cholan-gitis. E, bile duct proliferation after experimental ligation of the commonbile duct in rats. Sections inC andE were stained with H&E.D andF,adjacent sections of PSC (D) and experimental bile duct proliferation inrats (F) immunostained to detect AAH protein with anti-AAH antibod-ies. The staining reaction was negative.
1263
HAAH AND MALIGNANT TRANSFORMATION OF BILE DUCTS
on July 14, 2018. © 2000 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
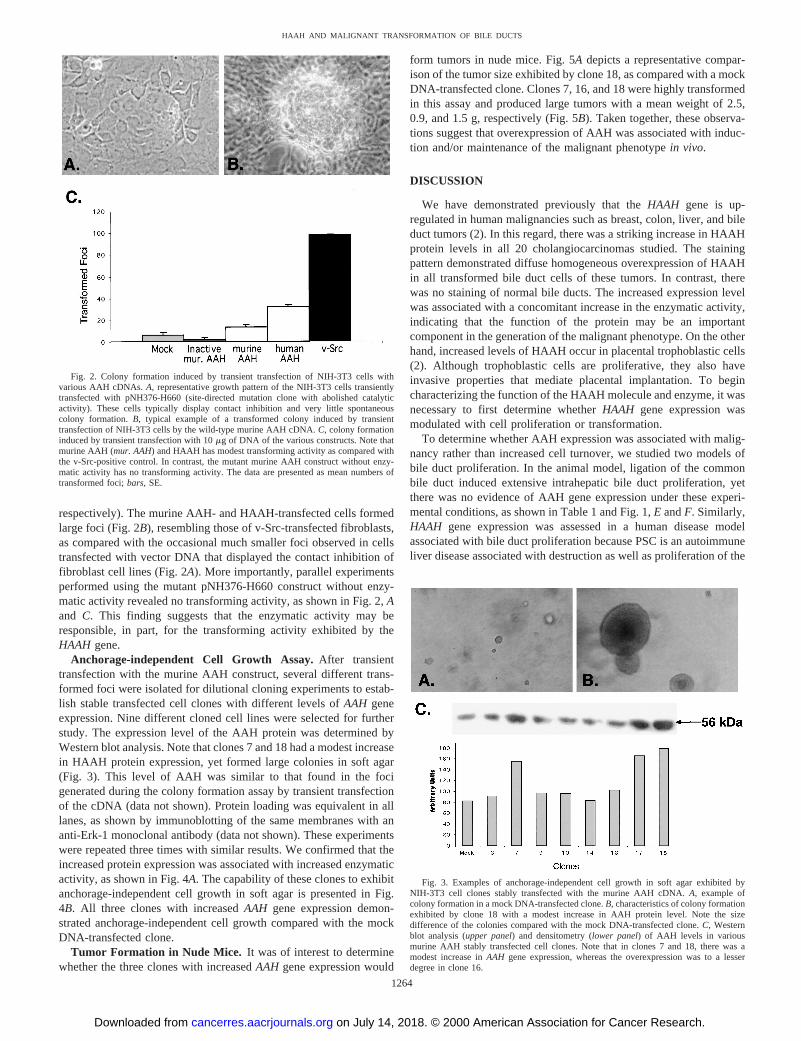
respectively). The murine AAH- and HAAH-transfected cells formedlarge foci (Fig. 2B), resembling those of v-Src-transfected fibroblasts,as compared with the occasional much smaller foci observed in cellstransfected with vector DNA that displayed the contact inhibition offibroblast cell lines (Fig. 2A). More importantly, parallel experimentsperformed using the mutant pNH376-H660 construct without enzy-matic activity revealed no transforming activity, as shown in Fig. 2,Aand C. This finding suggests that the enzymatic activity may beresponsible, in part, for the transforming activity exhibited by theHAAH gene.
Anchorage-independent Cell Growth Assay.After transienttransfection with the murine AAH construct, several different trans-formed foci were isolated for dilutional cloning experiments to estab-lish stable transfected cell clones with different levels ofAAH geneexpression. Nine different cloned cell lines were selected for furtherstudy. The expression level of the AAH protein was determined byWestern blot analysis. Note that clones 7 and 18 had a modest increasein HAAH protein expression, yet formed large colonies in soft agar(Fig. 3). This level of AAH was similar to that found in the focigenerated during the colony formation assay by transient transfectionof the cDNA (data not shown). Protein loading was equivalent in alllanes, as shown by immunoblotting of the same membranes with ananti-Erk-1 monoclonal antibody (data not shown). These experimentswere repeated three times with similar results. We confirmed that theincreased protein expression was associated with increased enzymaticactivity, as shown in Fig. 4A. The capability of these clones to exhibitanchorage-independent cell growth in soft agar is presented in Fig.4B. All three clones with increasedAAH gene expression demon-strated anchorage-independent cell growth compared with the mockDNA-transfected clone.
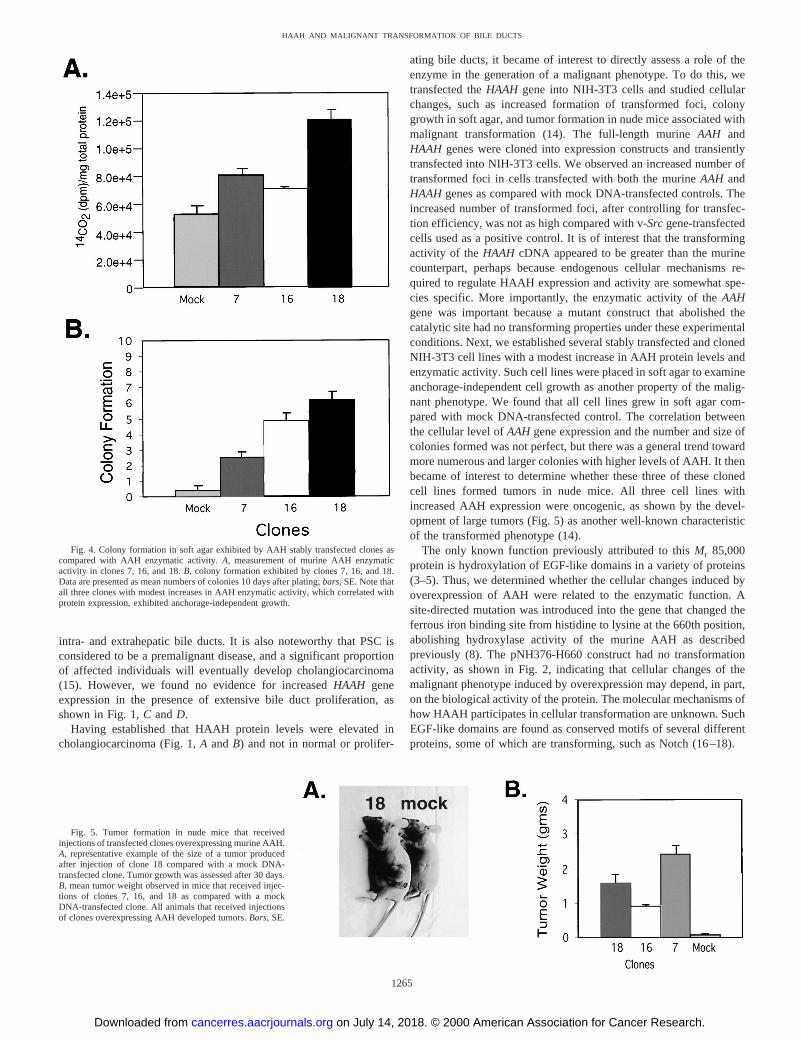
Tumor Formation in Nude Mice. It was of interest to determinewhether the three clones with increasedAAH gene expression would
form tumors in nude mice. Fig. 5Adepicts a representative compar-ison of the tumor size exhibited by clone 18, as compared with a mockDNA-transfected clone. Clones 7, 16, and 18 were highly transformedin this assay and produced large tumors with a mean weight of 2.5,0.9, and 1.5 g, respectively (Fig. 5B). Taken together, these observa-tions suggest that overexpression of AAH was associated with induc-tion and/or maintenance of the malignant phenotypein vivo.
DISCUSSION
We have demonstrated previously that theHAAH gene is up-regulated in human malignancies such as breast, colon, liver, and bileduct tumors (2). In this regard, there was a striking increase in HAAHprotein levels in all 20 cholangiocarcinomas studied. The stainingpattern demonstrated diffuse homogeneous overexpression of HAAHin all transformed bile duct cells of these tumors. In contrast, therewas no staining of normal bile ducts. The increased expression levelwas associated with a concomitant increase in the enzymatic activity,indicating that the function of the protein may be an importantcomponent in the generation of the malignant phenotype. On the otherhand, increased levels of HAAH occur in placental trophoblastic cells(2). Although trophoblastic cells are proliferative, they also haveinvasive properties that mediate placental implantation. To begincharacterizing the function of the HAAH molecule and enzyme, it wasnecessary to first determine whetherHAAH gene expression wasmodulated with cell proliferation or transformation.
To determine whether AAH expression was associated with malig-nancy rather than increased cell turnover, we studied two models ofbile duct proliferation. In the animal model, ligation of the commonbile duct induced extensive intrahepatic bile duct proliferation, yetthere was no evidence of AAH gene expression under these experi-mental conditions, as shown in Table 1 and Fig. 1,E andF. Similarly,HAAH gene expression was assessed in a human disease modelassociated with bile duct proliferation because PSC is an autoimmuneliver disease associated with destruction as well as proliferation of the
Fig. 2. Colony formation induced by transient transfection of NIH-3T3 cells withvarious AAH cDNAs.A, representative growth pattern of the NIH-3T3 cells transientlytransfected with pNH376-H660 (site-directed mutation clone with abolished catalyticactivity). These cells typically display contact inhibition and very little spontaneouscolony formation.B, typical example of a transformed colony induced by transienttransfection of NIH-3T3 cells by the wild-type murine AAH cDNA.C, colony formationinduced by transient transfection with 10mg of DNA of the various constructs. Note thatmurine AAH (mur. AAH) and HAAH has modest transforming activity as compared withthe v-Src-positive control. In contrast, the mutant murine AAH construct without enzy-matic activity has no transforming activity. The data are presented as mean numbers oftransformed foci;bars, SE.
Fig. 3. Examples of anchorage-independent cell growth in soft agar exhibited byNIH-3T3 cell clones stably transfected with the murine AAH cDNA.A, example ofcolony formation in a mock DNA-transfected clone.B, characteristics of colony formationexhibited by clone 18 with a modest increase in AAH protein level. Note the sizedifference of the colonies compared with the mock DNA-transfected clone.C, Westernblot analysis (upper panel) and densitometry (lower panel) of AAH levels in variousmurine AAH stably transfected cell clones. Note that in clones 7 and 18, there was amodest increase inAAH gene expression, whereas the overexpression was to a lesserdegree in clone 16.
1264
HAAH AND MALIGNANT TRANSFORMATION OF BILE DUCTS
on July 14, 2018. © 2000 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
intra- and extrahepatic bile ducts. It is also noteworthy that PSC isconsidered to be a premalignant disease, and a significant proportionof affected individuals will eventually develop cholangiocarcinoma(15). However, we found no evidence for increasedHAAH geneexpression in the presence of extensive bile duct proliferation, asshown in Fig. 1,C andD.
Having established that HAAH protein levels were elevated incholangiocarcinoma (Fig. 1,A andB) and not in normal or prolifer-
ating bile ducts, it became of interest to directly assess a role of theenzyme in the generation of a malignant phenotype. To do this, wetransfected theHAAH gene into NIH-3T3 cells and studied cellularchanges, such as increased formation of transformed foci, colonygrowth in soft agar, and tumor formation in nude mice associated withmalignant transformation (14). The full-length murineAAH andHAAH genes were cloned into expression constructs and transientlytransfected into NIH-3T3 cells. We observed an increased number oftransformed foci in cells transfected with both the murineAAH andHAAH genes as compared with mock DNA-transfected controls. Theincreased number of transformed foci, after controlling for transfec-tion efficiency, was not as high compared with v-Srcgene-transfectedcells used as a positive control. It is of interest that the transformingactivity of theHAAH cDNA appeared to be greater than the murinecounterpart, perhaps because endogenous cellular mechanisms re-quired to regulate HAAH expression and activity are somewhat spe-cies specific. More importantly, the enzymatic activity of theAAHgene was important because a mutant construct that abolished thecatalytic site had no transforming properties under these experimentalconditions. Next, we established several stably transfected and clonedNIH-3T3 cell lines with a modest increase in AAH protein levels andenzymatic activity. Such cell lines were placed in soft agar to examineanchorage-independent cell growth as another property of the malig-nant phenotype. We found that all cell lines grew in soft agar com-pared with mock DNA-transfected control. The correlation betweenthe cellular level ofAAH gene expression and the number and size ofcolonies formed was not perfect, but there was a general trend towardmore numerous and larger colonies with higher levels of AAH. It thenbecame of interest to determine whether these three of these clonedcell lines formed tumors in nude mice. All three cell lines withincreased AAH expression were oncogenic, as shown by the devel-opment of large tumors (Fig. 5) as another well-known characteristicof the transformed phenotype (14).
The only known function previously attributed to thisMr 85,000protein is hydroxylation of EGF-like domains in a variety of proteins(3–5). Thus, we determined whether the cellular changes induced byoverexpression of AAH were related to the enzymatic function. Asite-directed mutation was introduced into the gene that changed theferrous iron binding site from histidine to lysine at the 660th position,abolishing hydroxylase activity of the murine AAH as describedpreviously (8). The pNH376-H660 construct had no transformationactivity, as shown in Fig. 2, indicating that cellular changes of themalignant phenotype induced by overexpression may depend, in part,on the biological activity of the protein. The molecular mechanisms ofhow HAAH participates in cellular transformation are unknown. SuchEGF-like domains are found as conserved motifs of several differentproteins, some of which are transforming, such as Notch (16–18).
Fig. 4. Colony formation in soft agar exhibited by AAH stably transfected clones ascompared with AAH enzymatic activity.A, measurement of murine AAH enzymaticactivity in clones 7, 16, and 18.B, colony formation exhibited by clones 7, 16, and 18.Data are presented as mean numbers of colonies 10 days after plating;bars, SE. Note thatall three clones with modest increases in AAH enzymatic activity, which correlated withprotein expression, exhibited anchorage-independent growth.
Fig. 5. Tumor formation in nude mice that receivedinjections of transfected clones overexpressing murine AAH.A, representative example of the size of a tumor producedafter injection of clone 18 compared with a mock DNA-transfected clone. Tumor growth was assessed after 30 days.B, mean tumor weight observed in mice that received injec-tions of clones 7, 16, and 18 as compared with a mockDNA-transfected clone. All animals that received injectionsof clones overexpressing AAH developed tumors.Bars, SE.
1265
HAAH AND MALIGNANT TRANSFORMATION OF BILE DUCTS
on July 14, 2018. © 2000 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

It will be important in the future to develop assays reflecting thestate of Notch protein hydroxylation in the context of overexpressionof AAH because EGF-like domains may be involved in receptor-ligand interactions. Indeed, point mutations affecting aspartic acid orasparagine residues in EGF-like domains that are the targets forb-hydroxylation by AAH reduce calcium binding and therefore couldinfluence protein-protein interactions and thereby influence activationof downstream signal transduction pathways (19).
The data presented herein provide substantial evidence that high-level HAAH expression is linked to malignant transformation. Wedemonstrated that overexpression of the AAH cDNAs in NIH-3T3cells induces a transformed phenotype manifested by increased num-bers of transformed foci, anchorage-independent growth, and tumor-igenesis in nude mice. In addition, intact AAH enzyme was found tobe required for AAH-associated transformation. Of particular interestwere the findings that only modest increases in AAH expression andenzyme activity were required for cellular transformation. Small dif-ferences in the levels of an extracellular signaling molecule canspecify cell fate during development and possibly carcinogenesis.Threshold responses are often determined at the level of transcription,and previous studies have demonstrated that HAAH has been up-regulated at the transcriptional level (2, 20). These results suggest thatincreasedAAH gene expression and enzyme activity contribute to thegeneration or maintenance of the transformed phenotype (1) and arenot strictly related to cellular proliferation. Finally, there is very littleavailable data concerning genes that are up-regulated with malignanttransformation of bile duct epithelium, and HAAH immunoreactivityis detectable on tumor cell surface membranes (2). Assay of HAAHprotein levels in either biological fluids such as bile or cells obtainedby fine-needle aspiration deserves further attention as a possiblediagnostic marker of human cholangiocarcinoma in future.
REFERENCES
1. Wilson, B., Ozturk, M., Takahashi, H., Motte, P., Kew, M., Isselbacher K. J., andWands, J. R. Cell-surface changes associated with transformation of human hepato-cytes to the malignant phenotype. Proc. Natl. Acad. Sci. USA,85: 3140–3144, 1988.
2. Lavaissiere, L., Jia, S., Nishiyama, M., de la Monte, S., Stern, A. M., Wands, J. R.,and Friedman, P. A. Overexpression of human aspartyl(asparaginyl)b-hydroxylase inhepatocellular carcinoma and cholangiocarcinoma. J. Clin. Investig.,98: 1313–1323,1996.
3. Gronke, R. S., Van Dusen, W. J., Garsky, V. M., Jacobs, J. W., Sardana, M. K., Stern,A. M., and Friedman, P. A. Aspartylb-hydroxylase:in vitro hydroxylation of asynthetic peptide based on the structure of the first growth factor-like domain ofhuman factor IX. Proc. Natl. Acad. Sci. USA,86: 3609–3613, 1989.
4. Jia, S., Van Dusen, W. J., Diehl, R. E., Kohl, N. E., Dixon, R. A., Elliston, K. O.,Stern, A. M., and Friedman, P. A. cDNA cloning and expression of bovine aspartyl(asparaginyl)b-hydroxylase. J. Biol. Chem.,267: 14322–14327, 1992.
5. Jia, S., McGinnis, K., Van Dusen, W. J., Burke, C. J., Kuo, A., Griffin, P. R., Sardana,M. K., Elliston, K. O., Stern, A. M., and Friedman, P. A. A fully active catalyticdomain of bovine aspartyl (asparaginyl)b-hydroxylase expressed inEscherichia coli:characterization and evidence for the identification of an active-site region in verte-brate a-ketoglutarate-dependent dioxygenases. Proc. Natl. Acad. Sci. USA,91:7227–7231, 1994.
6. Downing, A. K., Knott, V., Werner, J. M., Cardy, C. M., Campbell, I. D., andHandford, P. A. Solution structure of a pair of calcium-binding epidermal growthfactor-like domains: implications for the Marfan syndrome and other genetic disor-ders. Cell,85: 597–605, 1996.
7. Nishimura, H., Takeya, H., Miyata, T., Suehiro, K., Okamura, T., Niho, Y., andIwanaga, S. Substitution of ASN92 by His in the second epidermal growth factor-likedomain results in defective interaction with factors VIIa/X. J. Biol. Chem.,268:24041–24046, 1993.
8. McGinnis, K., Ku, G. M., Van Dusen, W. J., Fu, J., Garsky, V., Stern, A. M., andFriedman, P. A. Site-directed mutagenesis of residues in a conserved region of bovineaspartyl (asparaginyl)b-hydroxylase: evidence that histidine 675 has a role in bindingFe21. Biochemistry,35: 3957–3962, 1996.
9. Maroney, A. C., Qureshi, S. A., Foster, D. A., and Brugge, J. S. Cloning andcharacterization of a thermolabile v-srcgene for use in reversible transformation ofmammalian cells. Oncogene,7: 1207–1214, 1992.
10. Monath, T. P., Wands, J. R., Hill, L. J., Gentry, M. K., and Gubler, D. J. Multisitemonoclonal immunoassay for dengue viruses: detection of viraemic human sera andinterference by heterologous antibody. J. Gen. Virol.,67: 639–650, 1986.
11. Sambrook, J., Fritsch, E. F., and Maniatis, T. Molecular Cloning: A LaboratoryManual, Ed. 2. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory, 1989.
12. Wang, Q. P., Van Dusen, W. J., Petroski, C. J., Garsky, V. M., Stern, A. M., andFriedman, P. A. Bovine liver aspartyl beta-hydroxylase. Purification and character-ization. J. Biol. Chem.,266: 14004–14010, 1991.
13. Gronke, R. S., Welsch, D. J., Van Dusen, W. J., Garsky, V. M., Sardana, M. K., Stern,A. M., and Friedman, P. A. Partial purification and characterization of bovine liveraspartylb-hydroxylase. J. Biol. Chem.,265: 8558–8565, 1990.
14. Ito, T., Sasaki, Y., and Wands, J. R. Overexpression of human insulin receptorsubstrate 1 induces cellular transformation with activation of mitogen-activatedprotein kinases. Mol. Cell. Biol.,16: 943–951, 1996.
15. Bergquist, A., Glaumann, H., Persson, B., and Broome, U. Risk factors and clinicalpresentation of hepatobiliary carcinoma in patients with primary sclerosing cholan-gitis: a case-control study. Hepatology,27: 311–316, 1998.
16. Ellisen, L. W., Bird, J., West, D. C., Soreng, A. L., Reynolds, T. C., Smith, S. D., andSklar, J. TAN-1, the human homolog of theDrosophila notchgene, is broken bychromosomal translocations in T lymphoblastic neoplasms. Cell,66: 649–661, 1991.
17. Pear, W. S., Aster, J. C., Scott, M. L., Hasserjian, R. P., Soffer, B., Sklar, J., andBaltimore, D. Exclusive development of T cell neoplasms in mice transplanted withbone marrow expressing activatedNotchalleles. J. Exp. Med.,183:2283–2291, 1996.
18. Joutel, A., and Tournier-Lasserve, E. Notch signalling pathway and human diseases.Semin. Cell. Dev. Biol.,9: 619–625, 1998.
19. Handford, P. A., Mayhew, M., Baron, M., Winship, P. R., Campbell, I. D., andBrownlee, G. G. Key residues involved in calcium-binding motifs in EGF-likedomains. Nature (Lond.),351: 164–167, 1991.
20. Mannervik, M., Nibu, Y., Zhang, H., and Levine, M. Transcriptional coregulators indevelopment. Science (Washington DC),284: 606–609, 1999.
1266
HAAH AND MALIGNANT TRANSFORMATION OF BILE DUCTS
on July 14, 2018. © 2000 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

2000;60:1261-1266. Cancer Res Nedim Ince, Suzanne M. de la Monte and Jack R. Wands -Hydroxylase Is Associated with Malignant Transformation
βOverexpression of Human Aspartyl (Asparaginyl)
Updated version
http://cancerres.aacrjournals.org/content/60/5/1261
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/60/5/1261.full#ref-list-1
This article cites 19 articles, 10 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/60/5/1261.full#related-urls
This article has been cited by 5 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/60/5/1261To request permission to re-use all or part of this article, use this link
on July 14, 2018. © 2000 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from