orchestration of three transporters and distinct vascular ... · pdf fileorchestration of...
TRANSCRIPT

Orchestration of three transporters and distinctvascular structures in node for intervasculartransfer of silicon in riceNaoki Yamajia,1, Gen Sakuraib,1, Namiki Mitani-Uenoa, and Jian Feng Maa,2
aInstitute of Plant Science and Resources, Okayama University, Kurashiki 710-0046, Japan; and bEcosystem Informatics Division, National Institute forAgro-Environmental Sciences, Tsukuba 305-8604, Japan
Edited by Maarten J. Chrispeels, University of California, San Diego, La Jolla, CA, and approved July 27, 2015 (received for review May 7, 2015)
Requirement of mineral elements in different plant tissues is notoften consistent with their transpiration rate; therefore, plantshave developed systems for preferential distribution of mineralelements to the developing tissues with low transpiration. Herewe took silicon (Si) as an example and revealed an efficient systemfor preferential distribution of Si in the node of rice (Oryza sativa).Rice is able to accumulate more than 10% Si of the dry weight inthe husk, which is required for protecting the grains from waterloss and pathogen infection. However, it has been unknown for along time how this hyperaccumulation is achieved. We found thatthree transporters (Lsi2, Lsi3, and Lsi6) located at the node areinvolved in the intervascular transfer, which is required for thepreferential distribution of Si. Lsi2 was polarly localized to thebundle sheath cell layer around the enlarged vascular bundles,which is next to the xylem transfer cell layer where Lsi6 is local-ized. Lsi3 was located in the parenchyma tissues between en-larged vascular bundles and diffuse vascular bundles. Similar toLsi6, knockout of Lsi2 and Lsi3 also resulted in decreased distribu-tion of Si to the panicles but increased Si to the flag leaf. Further-more, we constructed a mathematical model for Si distribution andrevealed that in addition to cooperation of three transporters, anapoplastic barrier localized at the bundle sheath cells and devel-opment of the enlarged vascular bundles in node are also requiredfor the hyperaccumulation of Si in rice husk.
node | rice transporter | apoplastic barrier | silicon distribution |mathematical model
Plants have different requirement for mineral elementsdepending on organs and tissues. For examples, developing
tissues such as new leaves and reproductive organs require moremineral elements for their active growth (1). However, these tissuesusually have low transpiration; therefore, transpiration-dependentdistribution of mineral elements is not sufficient to meet their highrequirements. Plants must have developed systems for preferentialdistribution of mineral elements depending on the requirements.However, the molecular mechanisms for these systems are poorlyunderstood. Recently, several studies have shown that nodes func-tion as a hub for distribution of mineral elements in graminaceousplants (2).Node is a key component of the phytomer in graminaceous
plants, which is connected with a leaf, a tiller (or a tiller bud), andcrown roots (or these primordia) to the culm or panicle (2, 3).Therefore, graminaceous plants are composed of repeated nodes.Nodes have apparently complicated but regularly organized vas-cular bundles (VBs). No VBs are continuously connected from thebottom to the top of the culm, but a unit of VB is repeated with atwo-thirds overlapped rule; one axial VB is connected with threenodes and a leaf and specialized at different nodes as diffuse VB(DVB), transit VB (TVB), and enlarged VB (EVB) from basal toapical (2, 3). In a node, three different axial VBs with differentphases coexist and connected transversely by nodal vascular anas-tomosis (NVA). TVB is a passage phase, but its appearance andfunction within the node are similar to one of the multiple DVBs.
After EVB phase, the VB is continued to a VB of the leaf asso-ciating the node (2–4). As an exception, DVBs in the uppermosttwo nodes (nodes I and II), in which VBs have no correspondingleaves, become VBs in the panicle without EVB phase as detailedin ref. 2. Based on these repetitive vascular constructions, inter-vascular transfers between axial VBs are required for preferentialdelivery of nutrients to apical tissues such as developing leaf orpanicle (2). Furthermore, the node also shows distinct structures atcellular and subcellular level, which are characterized by (i) ex-panded xylem area of EVBs; (ii) differentiation of xylem transfercells (XTCs), which have expanded cell surface due to cell wallingrowth facing to the xylem vessel; (iii) increased number ofDVBs around the EVBs; and (iv) parenchyma cell bridge (PCB)with dense plasmodesmata between EVB and DVB (2, 4, 5).However, the role of these structures in intervascular transfer ofnutrients has not been examined.In the present study, we examined the preferential distribution
system of silicon (Si) in rice. Silicon, the most abundant mineralelement, shows beneficial effects for plant growth (6). Thebeneficial effects are characterized by protecting plants fromabiotic and biotic stresses (6). Silicon is especially important forhigh and sustainable production of rice (Oryza sativa), a typical Siaccumulating species. Rice is able to accumulate Si up to 10% ofthe shoot dry weight, which is severalfold more than macronu-trients such as nitrogen, phosphorus, and potassium (7). Lowaccumulation of Si in rice results in significant reduction of theyield (8); therefore, Si fertilization has been considered as a
Significance
Requirement of mineral elements differs with different organsand tissues; therefore, plants have developed systems forpreferentially delivering mineral elements to tissues with highrequirement. However, the molecular mechanisms for thesesystems are poorly understood. We took silicon (Si) as an ex-ample and revealed an efficient distribution system occurringin the node of rice, which is a hub for distribution. We foundthat hyperaccumulation of Si in the husk (more than 10%) isachieved by cooperation of three different Si transporters lo-calized at the different cell layers in the node. Furthermore,mathematical modeling showed that an apoplastic barrier anddevelopment of enlarged vascular bundles are also required.Our work revealed a set of players for efficient distributioncontrol in node.
Author contributions: N.Y. and J.F.M. designed research; N.Y., G.S., N.M.-U., and J.F.M. per-formed research; N.Y., G.S., N.M.-U., and J.F.M. analyzed data; and N.Y., G.S., and J.F.M. wrotethe paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1N.Y. and G.S. contributed equally to this work.2To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1508987112/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1508987112 PNAS | September 8, 2015 | vol. 112 | no. 36 | 11401–11406
PLANTBIOLO
GY

practice of rice production in many countries, especially underheavy application of nitrogen fertilizers (7).Silicon in soil solution is present in the form of silicic acid, an
uncharged molecule (7). Silicic acid is taken by the roots throughtwo different transporters: Lsi1 and Lsi2 in rice (9, 10). Lsi1 be-longs to NIP subgroup of aquaporin and preamble to silicic acid,whereas Lsi2 is an efflux transporter of silicic acid (9, 10). BothLsi1 and Lsi2 are localized at the exodermis and endodermis cellsof rice roots, but in contrast to Lsi1, which is localized at the distalside, Lsi2 is localized at the proximal side (9, 10). Knockout ofeither Lsi1 or Lsi2 results in defect of Si uptake (9, 10). Recently, insilico mathematical modeling revealed that in addition to co-operation of Lsi1 and Lsi2, presence of the Casparian strips at theexodermis and endodermis is also required for efficient Si uptakeand high accumulation (11).After uptake, more than 95% of Si is rapidly translocated from
the roots to the shoots (7). Silicon is also present in the form ofsilicic acid in the xylem sap (12), which is unloaded by Lsi6, ahomolog of Lsi1 (13). Lsi6 is polarly localized in the adaxial side ofthe xylem parenchyma cells in the leaf sheaths and leaf blades (13).With water loss due to transpiration, silicic acid is gradually con-centrated and polymerized to amorphous silica, which is depositedbeneath the cuticle, forming cuticle–silica double layers, and insideof particular cells of leaf epidermis (motor cells and short silicacells) (7). Knockout of Lsi6 results in leakage of large amount ofsilicic acid into the leaf guttation and altered deposition of Si in theleaves (13).At the reproductive stage, higher Si is finally deposited in the
husk, which can reach more than 10% of the dry weight (7).Silicon in the husk is also deposited between the epidermal cellwall and the cuticle, forming a cuticle–silica double layer (7).This heavy deposition of Si in the husk is very important for ricegrain fertility because it prevents water loss and pathogen (e.g.,panicle blast) infection (6–8). However, the question arises onhow Si is highly deposited in the husk. The rice husk does nothave stoma at the outside. Furthermore, compared with theleaves, the surface area is much smaller. Therefore, transpira-tion-dependent distribution is unlikely responsible for the high Siaccumulation in the husk.Previously, Lsi6 highly expressed in the nodes was reported to be
involved in the intervascular transfer for preferential distribution ofSi. Lsi6 in nodes is polarly localized at the xylem parenchyma cells(mainly XTCs) of EVBs (14). Knockout of Lsi6 resulted in de-creased Si accumulation in the husk but increased Si in the flag leaf(14). However, Lsi6 is a channel-type passive transporter, whichtransports silicic acid following concentration gradient (13, 15).Therefore, Lsi6 alone cannot explain hyperaccumulation of Si inthe husk. In the present study, we found that the other two activeefflux transporters are also involved in the intervascular transferrequired for high accumulation of Si in the husk. Furthermore, ourmathematical modeling revealed that the distinct nodal structuresincluding EVBs, XTCs, and apoplastic barrier are also required forthe efficient intervascular transfer of Si in rice.
ResultsExpression Profiles of Si Transporters in Nodes. To find out trans-porters involved in the intervascular transfer of Si in nodes, weinvestigated the expression profile of both passive Si channel (Lsi1-like) and active Si efflux transporter (SIET/Lsi2-like) homologgenes using node I RNA at heading stage. In rice genome, there isonly one homolog of Lsi1 (Lsi6) (9) but four homologs of Lsi2/SIET1 (Lsi3/SIET2 and SIET3–5) (10). Among them, Lsi6, Lsi2,and Lsi3 showed high-level expression in the node I (Fig. 1A). Bycontrast, expression of Lsi1 and SIET3 was not detected, and onlytrace expression of SIET4 and SIET5 was found in node I(Fig. 1A).A detailed expression pattern was further investigated by quan-
titative RT-PCR using various organs of rice at the flowering stage.
These samples were also used for analysis of Lsi6 previously (14).Similar to Lsi6, the highest expression of both Lsi2 and Lsi3 wasfound in the uppermost node I (Fig. 1 B and C). Expression of Lsi2and Lsi3 was also detected in lower nodes including nodes II and III(Fig. 1 B and C). In contrast to Lsi3, which was also expressed in thepeduncle (internode I) and rachis, there was no expression of Lsi2 inthese organs (Fig. 1 B andC). Expressions of both Lsi2 and Lsi3 werealso not detected in caryopsis (Fig. 1 B and C).
Functional Characterization of Lsi3. Among Si transporter geneshighly expressed in node I, Lsi6 and Lsi2 proteins have beenfunctionally characterized, but Lsi3 is uncharacterized in termsof transport activity and subcellular localization. Lsi3 shares 80%identity with Lsi2 at amino acid sequence (Fig. S1). To examinewhether Lsi3 also has efflux transport activity for silicic acid, weexpressed Lsi3 in Xenopus laevis oocyte and determined its effluxtransport activity. Similar to Lsi2, Lsi3 also showed efflux transportactivity for Si (Fig. S2).Furthermore, we expressed Lsi3 in the lsi2 rice mutant under
the control of Lsi2 promoter. Analysis of two independenttransgenic lines showed that introduction of Lsi3 significantlyincreased Si uptake (Fig. S2).To investigate subcellular localization of Lsi3 protein, we pre-
pared a fusion gene of GFP and Lsi3 (GFP-Lsi3) and transientlyintroduced it into onion epidermal cells together with DsRed as amarker for cytosol and nucleus by particle bombardment. Fluores-cence signal from GFP-Lsi3 was observed mainly at outer region ofthe cell, which is located outside of fluorescence of DsRed (Fig. S3).
B
C
A
Lsi2
Lsi3
Lsi1
Lsi6
SIET3
SIET4
SIET5
m500400300200100
CaryopsisRachis
PeduncleNode INode IINode III
Unelongated stem
Relative expression
0.0 0.5 1.0 1.5
CaryopsisRachis
PeduncleNode INode IINode III
Unelongated stem
Relative expression
0.0 0.5 1.0 1.5
Fig. 1. Expression analysis. (A) Semiquantitative RT-PCR of Lsi1-like genes(Lsi1 and Lsi6) and Lsi2-like genes (Lsi2, Lsi3, SIET3, SIET4, and SIET5) in nodeI of rice sampled at the heading stage. PCR products (25 cycle) with laddermarker (m) were electrophoresed and stained by ethidium bromide. (B andC) Expression pattern of Lsi2 (B) and Lsi3 (C) determined by quantitative real-time RT-PCR in various organs at flowering stages. Expression level relativeto node I was shown. Actinwas used as an internal standard. Data are means ±SD of three biological replicates.
11402 | www.pnas.org/cgi/doi/10.1073/pnas.1508987112 Yamaji et al.

In situ subcellular localization of Lsi3 protein was also investigatedby immunohistochemical staining in rice node I. No signal in thelsi3 knockout mutant as described below was detected (Fig. S4),indicating high specificity of this antibody against Lsi3. Doublestaining with 4′,6-diamidino-2-phenylindole (DAPI) for nucleishowed that the fluorescence signal from Lsi3 antibody was lo-calized mainly at the peripheral region of the cells, which wascircumscribed but did not envelope the nuclei (Fig. S4). Takentogether, these results show that Lsi3 is a plasma membrane-localized efflux transporter of silicic acid.
Tissue-Specific Localization of Si Transporters in Node. Lsi6 is local-ized at the xylem parenchyma cells of EVBs of node I with polarityfacing toward the xylem vessels (14), but the tissue localizations ofLsi2 and Lsi3 in nodes are unknown. To examine the tissue spec-ificity of localization of Lsi2 and Lsi3, we performed coimmuno-staining of Lsi2 or Lsi3 with Lsi6. Lsi2 protein (magenta) waslocalized at the bundle sheath (bs) cells around the EVBs (Fig.2A), which is next to the cell layer where Lsi6 is localized (green).Furthermore, Lsi2 showed a polar localization at the distal side ofthe vasculature (Fig. 2B), which is the opposite side of Lsi6. On theother hand, Lsi3 protein (magenta) was detected in the paren-chyma cells between the bundle sheath cell layer of EVBs andDVBs (Fig. 2 C and D). Unlike Lsi2 and Lsi6, Lsi3 did not showpolarity (Fig. 2D).
Detection of Apoplastic Barrier in Node. In rice roots, presence ofCasparian strips at the exodermis and endodermis preventsapoplastic flow of water and solutes; therefore, cooperation ofpolarly localized Lsi1 and Lsi2 at these cell layers is required forefficient Si uptake (9–11). In node, Lsi2 and Lsi6 also showed
back-to-back polar localization (Fig. 2 A and B), although atdifferent cell layers next to each other. To understand the sig-nificance of this polar localization, we examined whether apo-plastic barrier is also present in node. We fed periodic acid, atracer of apoplastic flow (16), from cut end of the internodebelow node II, followed by Schiff’s staining. At the cross sectionof node I, the xylem regions of the EVBs were heavily stained,but the phloem regions of EVBs and DVBs were hardly stained(Fig. S5). No staining was observed at the bundle sheath celllayer of EVBs and outer parenchyma cells between EVBs andDVBs (Fig. S5). This result shows that the apoplastic tracer wasblocked inside the bundle sheath cell layer, indicating the pres-ence of an apoplastic barrier at the bundle sheath of EVBs.
Stem-Fed Si Distribution to Different Organs. To investigate theexact roles of three Si transporters highly expressed in node, wecompared the distribution pattern of Si to different organs be-tween knockout lines and their wild-type rice.Knockout lines of Lsi2 and Lsi6 were used in previous studies
(6, 9), whereas lsi3mutant is a T-DNA insertion line (Fig. S1). Theexpression of Lsi3 at the 3′-untranslated region was hardly detectedin the lsi3 mutant (Fig. S1). Because Lsi2 and Lsi6 are alsoexpressed in the roots (10, 13), to rule out the effect from root Siuptake, we fed Si as well as Rb and Sr as tracer of symplastic andapoplastic transfer from the cut end of the internode II for 24 h. Sris a phloem-immobile and xylem-mobile element, whereas Rb istransported through the phloem (17). Knockout of either Lsi6,Lsi2, or Lsi3 resulted in decreased distribution of Si to the panicleorgans including spikelet, rachis, and peduncle but increased dis-tribution to the flag leaf (Fig. 3A). Among Lsi6, Lsi2, and Lsi3,knockout of Lsi6 showed the largest effect in decreasing Si distri-bution to the panicles. By contrast, there was almost no differencein the distribution of Rb and Sr to different organs between mu-tants and their corresponding WTs (Fig. 3 B and C). Based on Srdistribution, it was estimated that 70–80% of transpiration oc-curred from the flag leaf in this experiment (Fig. 3B).
Construction of Mathematical Model for Si Distribution in Node I. Tocomprehensively understand the distribution system of Si innode, we constructed a mathematical model. We simulated thetransfer of Si in the node I by using a diffusion equation onthe 2D grids (11) and a 2D explicit finite difference method.The simulations represent a 2D longitudinal section of node I.For simplicity, the simulation region includes only one EVB andone DVB connected by NVA (Fig. S6). Areas of EVB and DVB(2), the location of the Si transporters (Fig. 2), apoplastic barrier(Fig. S5), and transpiration rate (i.e., distribution of Sr) frompanicle and flag leaf (Fig. 3C) were estimated according to theexperimental data. Then, we calibrated the model parameters, Sitransport activity of three transporters, and background of Sipermeability of cell membrane per unit simulation area based onthe results of the stem-fed in vivo experiment shown in Fig. 3.Using these parameter sets, we established a model for Si dis-tribution in node I. Using this model, we were able to reproducethe Si distribution pattern in lsi6, lsi2, and lsi3 mutants as ob-served in the feeding experiment (Fig. S7).
In Silico Simulation Experiments of Si Distribution. Based on themodel constructed, we conducted in silico modeling experimentsto estimate the importance of the cooperative effect of threetransporters and the contribution of nodal structural features tothe distribution. In the normal setting of the model (corre-sponding to the wild-type rice), the Si concentration in the DVBwas much higher than that in EVB, indicating a preferentialtransfer of Si to DVB in node I (Fig. 4A). However, if Lsi6 wasabsent, the difference in Si concentration between DVB andEVB disappeared, indicating the significant role of Lsi6 in theintervascular transfer of Si (Fig. 4A). If Lsi2 or Lsi3 was absent,
DVBDVBDVB
DVB
DVB
DVB
DVBDVBDVB
DVB
EVxEVp
EVx
EVp
EVx
DVB DVB DVB
DVBDVB
DVBDVB
DVBDVB DVB
EVxEVp
EVx
EVx
A B
C D
DVp
DVx
DVxDVxDVx
DVB
EVx
EVp
bs
EVx
bs
Fig. 2. Tissue specificity of localization of Lsi6, Lsi2, and Lsi3 in node I. NodeI of the wild-type rice at heading stage was sampled for immunostaining.Coimmunostaining using Lsi6 antibody (green) and Lsi2 antibody (magentain A and B) or Lsi3 antibody (magenta in C and D) was performed. Auto-fluorescence of the cell wall is shown in blue/cyan color. Nonspecific signalsin nuclei by Lsi2 antibody was also observed in A and B. B and D are mag-nified image of yellow boxes area in A and C, respectively. DVB, xylem andphloem regions of EVBs (EVx and EVp), xylem and phloem regions of DVBs(DVx and DVp), and bundle sheath cell layer of EVBs (bs) are indicated. (Scalebar, 100 μm.)
Yamaji et al. PNAS | September 8, 2015 | vol. 112 | no. 36 | 11403
PLANTBIOLO
GY
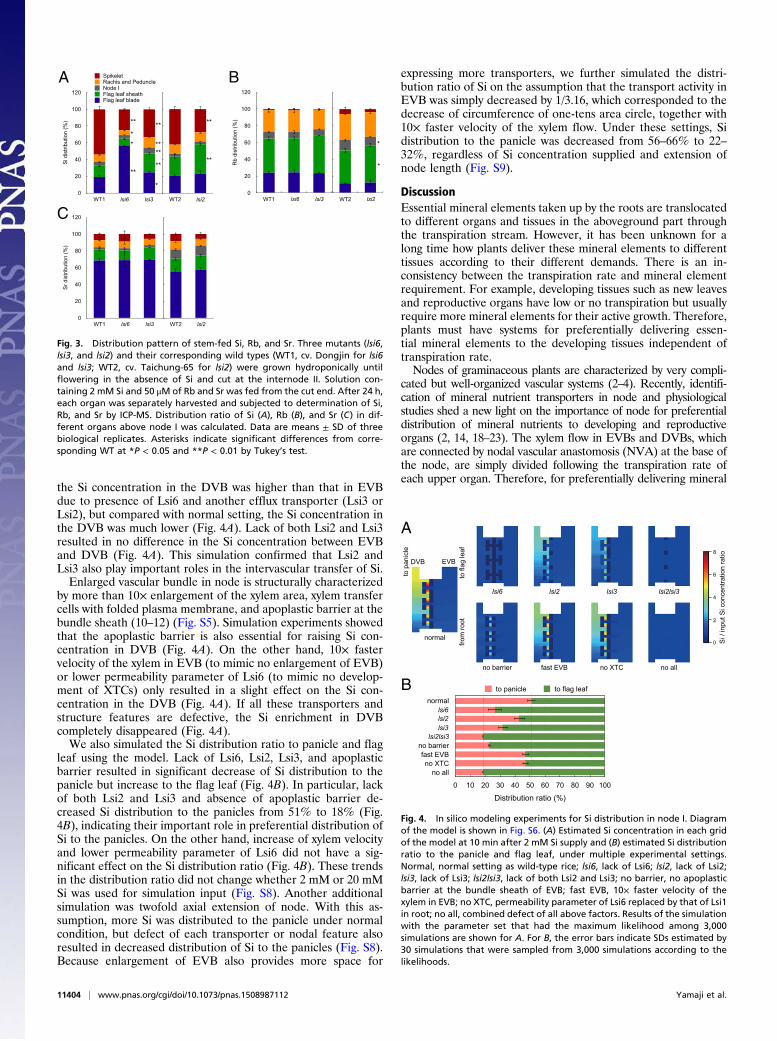
the Si concentration in the DVB was higher than that in EVBdue to presence of Lsi6 and another efflux transporter (Lsi3 orLsi2), but compared with normal setting, the Si concentration inthe DVB was much lower (Fig. 4A). Lack of both Lsi2 and Lsi3resulted in no difference in the Si concentration between EVBand DVB (Fig. 4A). This simulation confirmed that Lsi2 andLsi3 also play important roles in the intervascular transfer of Si.Enlarged vascular bundle in node is structurally characterized
by more than 10× enlargement of the xylem area, xylem transfercells with folded plasma membrane, and apoplastic barrier at thebundle sheath (10–12) (Fig. S5). Simulation experiments showedthat the apoplastic barrier is also essential for raising Si con-centration in DVB (Fig. 4A). On the other hand, 10× fastervelocity of the xylem in EVB (to mimic no enlargement of EVB)or lower permeability parameter of Lsi6 (to mimic no develop-ment of XTCs) only resulted in a slight effect on the Si con-centration in the DVB (Fig. 4A). If all these transporters andstructure features are defective, the Si enrichment in DVBcompletely disappeared (Fig. 4A).We also simulated the Si distribution ratio to panicle and flag
leaf using the model. Lack of Lsi6, Lsi2, Lsi3, and apoplasticbarrier resulted in significant decrease of Si distribution to thepanicle but increase to the flag leaf (Fig. 4B). In particular, lackof both Lsi2 and Lsi3 and absence of apoplastic barrier de-creased Si distribution to the panicles from 51% to 18% (Fig.4B), indicating their important role in preferential distribution ofSi to the panicles. On the other hand, increase of xylem velocityand lower permeability parameter of Lsi6 did not have a sig-nificant effect on the Si distribution ratio (Fig. 4B). These trendsin the distribution ratio did not change whether 2 mM or 20 mMSi was used for simulation input (Fig. S8). Another additionalsimulation was twofold axial extension of node. With this as-sumption, more Si was distributed to the panicle under normalcondition, but defect of each transporter or nodal feature alsoresulted in decreased distribution of Si to the panicles (Fig. S8).Because enlargement of EVB also provides more space for
expressing more transporters, we further simulated the distri-bution ratio of Si on the assumption that the transport activity inEVB was simply decreased by 1/3.16, which corresponded to thedecrease of circumference of one-tens area circle, together with10× faster velocity of the xylem flow. Under these settings, Sidistribution to the panicle was decreased from 56–66% to 22–32%, regardless of Si concentration supplied and extension ofnode length (Fig. S9).
DiscussionEssential mineral elements taken up by the roots are translocatedto different organs and tissues in the aboveground part throughthe transpiration stream. However, it has been unknown for along time how plants deliver these mineral elements to differenttissues according to their different demands. There is an in-consistency between the transpiration rate and mineral elementrequirement. For example, developing tissues such as new leavesand reproductive organs have low or no transpiration but usuallyrequire more mineral elements for their active growth. Therefore,plants must have systems for preferentially delivering essen-tial mineral elements to the developing tissues independent oftranspiration rate.Nodes of graminaceous plants are characterized by very compli-
cated but well-organized vascular systems (2–4). Recently, identifi-cation of mineral nutrient transporters in node and physiologicalstudies shed a new light on the importance of node for preferentialdistribution of mineral nutrients to developing and reproductiveorgans (2, 14, 18–23). The xylem flow in EVBs and DVBs, whichare connected by nodal vascular anastomosis (NVA) at the base ofthe node, are simply divided following the transpiration rate ofeach upper organ. Therefore, for preferentially delivering mineral
normal
no barrierfast EVBno XTC
no all
Distribution ratio (%)0 10 20 30 40 50 60 70 80 90 100
to panicle to flag leaf
A
B
normal
lsi6 lsi3lsi2 lsi2lsi3
no barrier fast EVB no XTC no all
DVB EVB
to p
anic
le
to fl
ag le
affro
m ro
ot
lsi6lsi2lsi3
lsi2lsi3
Si /
inpu
t Si c
once
ntra
tion
ratio
0
2
4
6
8
Fig. 4. In silico modeling experiments for Si distribution in node I. Diagramof the model is shown in Fig. S6. (A) Estimated Si concentration in each gridof the model at 10 min after 2 mM Si supply and (B) estimated Si distributionratio to the panicle and flag leaf, under multiple experimental settings.Normal, normal setting as wild-type rice; lsi6, lack of Lsi6; lsi2, lack of Lsi2;lsi3, lack of Lsi3; lsi2lsi3, lack of both Lsi2 and Lsi3; no barrier, no apoplasticbarrier at the bundle sheath of EVB; fast EVB, 10× faster velocity of thexylem in EVB; no XTC, permeability parameter of Lsi6 replaced by that of Lsi1in root; no all, combined defect of all above factors. Results of the simulationwith the parameter set that had the maximum likelihood among 3,000simulations are shown for A. For B, the error bars indicate SDs estimated by30 simulations that were sampled from 3,000 simulations according to thelikelihoods.
Si d
istri
butio
n (%
)
0
20
40
60
80
100
120
WT1
Flag leaf bladeFlag leaf sheathNode IRachis and PeduncleSpikelet
WT2
*
**
lsi6 lsi3 lsi2
*
**
**
****
**
*
**
**
A
WT2
Sr d
istri
butio
n (%
)
0
20
40
60
80
100
120
WT1 lsi6 lsi3 lsi2
B
CWT2
*
*
Rb
dist
ribut
ion
(%)
0
20
40
60
80
100
120
WT1 lsi6 lsi3 lsi2
Fig. 3. Distribution pattern of stem-fed Si, Rb, and Sr. Three mutants (lsi6,lsi3, and lsi2) and their corresponding wild types (WT1, cv. Dongjin for lsi6and lsi3; WT2, cv. Taichung-65 for lsi2) were grown hydroponically untilflowering in the absence of Si and cut at the internode II. Solution con-taining 2 mM Si and 50 μM of Rb and Sr was fed from the cut end. After 24 h,each organ was separately harvested and subjected to determination of Si,Rb, and Sr by ICP-MS. Distribution ratio of Si (A), Rb (B), and Sr (C) in dif-ferent organs above node I was calculated. Data are means ± SD of threebiological replicates. Asterisks indicate significant differences from corre-sponding WT at *P < 0.05 and **P < 0.01 by Tukey’s test.
11404 | www.pnas.org/cgi/doi/10.1073/pnas.1508987112 Yamaji et al.

elements, an intervascular transfer from EVBs to DVBs in node isrequired (2). Based on the structure of the node, this intervasculartransfer includes at least three steps: unloading of a mineral ele-ment from xylem flow in EVB, transfer through symplast or apo-plast of PCB, and reloading to xylem or phloem of DVB (2) (Fig.5). Although different transporters are supposed to be involved inthese steps, only a few of them in some steps have been identified.For instance, Lsi6 is involved in Si unloading from EVB (14, 18);OsHMA5 and OsYSL16 are involved in Cu exclusion from xylemand loading to phloem of DVB, respectively (19, 20), whereasOsHMA2 is required for Zn/Cd reloading to phloem of DVB (21);and OsNramp3 has a dual role for both xylem unloading fromEVB and phloem reloading to DVB of Mn (22). In the presentstudy, by integrating in vivo experiments with mathematical mod-eling, we were able to identify all factors involved in the inter-vascular transfer for preferential distribution of Si in node (Fig. 5).First, we identified two Si transporters (Lsi2 and Lsi3) highly
expressed in the node in addition to Lsi6 reported previously (Fig.1) (14). Both of them are efflux transporters of Si but localized atthe different cell layers between EVB and DVB of node (Figs. 2and 5). Lsi2 was localized at the distal side of bundle sheath cells ofEVB having apoplastic barrier, which is the next cell layer whereLsi6 was localized (14) (Fig. 2), whereas Lsi3 was localized at theparenchyma cells without polarity between the bundle sheath andDVB (Fig. 2). The parenchyma cells between EVBs and DVBs aresymplastically connected by dense plasmodesmata, so-called PCB(5) (Fig. 5). Therefore, Si in the xylem of EVB is taken up by Lsi6and symplastically moved to the next cell layer, the bundle sheaththrough the plasmodesmata (Fig. 5). Part of Si is released to theapoplastic space of parenchyma cells by Lsi2, which is transferredto the xylem of DVB through apoplastic pathway, whereasremaining Si is moved to the parenchyma cells through the plas-modesmata, followed by releasing to the xylem of DVB by Lsi3(Fig. 5). Knockout of either Lsi2 or Lsi3 resulted in decreaseddistribution of Si to the panicles but increased distribution to the
flag leaf (Fig. 3), indicating that the cooperation of these threetransporters is required for the preferential distribution of Si to thepanicles and subsequently high accumulation of Si in the husk(Fig. 5).Second, we constructed a mathematical model for distribution
of Si in node I (Fig. S6), which is well fit to the experimental data(Fig. S7). We found that the apoplastic barrier identified at thebundle sheath of EVBs in this study is also required for efficientintervascular transfer of Si in node based on the mathematicmodeling (Fig. 4 and Fig. S8). This barrier is essential for effi-cient cooperation of three transporters by generating a concen-tration gradient from xylem of EVB to XTC–PCB for promotionof Si transport via Lsi6 and further directional transport by Lsi2and Lsi3 at the opposite side of the barrier (Figs. 4 and 5). Thisaxisymmetric arrangement of passive channel (Lsi6) and activeefflux transporters (Lsi2 and Lsi3) with an apoplastic barrier isvery similar to the roots, where Lsi1 and Lsi2 for Si uptake arelocalized at the exodermis and endodermis with Casprian strips(9, 10). A previous modeling study also demonstrated that theseapoplastic barriers in root are required for the efficient uptakeand enrichment of Si to the stele (11). However, there are twodifferences in these cooperative systems between the root andnode. One is that two efflux transporters (Lsi2 and Lsi3) areinvolved in node, but only one efflux transporter Lsi2 is involvedin root. The second difference is that Lsi1 and Lsi2 are localizedat the same cells with different polarity in root, but in node, Lsi6,Lsi2, and Lsi3 are localized at the different cell layers (Fig. 5).However, these two cooperative systems show similar function inroot and node. This is because different from root, in node, celllayers expressing Lsi6, Lsi2, and Lsi3 are tightly connected toeach other by plasmodesmata (5) (Fig. 5). These cell layerstherefore behave like a single cell in the model. This connectedmultilayer structure by cells specialized for different functions ispresumably more suitable for the more integrated multivascularstructure of nodes compared with multifunctional single-celllayer(s) in the root. In fact, single knockout of Lsi2 or Lsi3 hadsmaller effect on the Si distribution to the panicle compared withthat of Lsi6 (Fig. 3A), indicating that both Lsi2 and Lsi3 areinvolved in generation of the Si concentration gradient for mo-tive force of Lsi6 channel (Figs. 4 and 5). This is supported by thefact that Si enrichment in the DVB was completely lost if bothLsi2 and Lsi3 were knocked out in the model (Fig. 4).Xylem transfer cells where Lsi6 is located are characterized by
folded plasma membrane due to cell wall ingrowth, resulting inincreased uptake surface (2, 5). We simulated the importance ofthis distinct structure in Si distribution by using the model con-structed. In our model, the estimated transport activity param-eter of Lsi2 per unit modeling grid (7.05 ± 2.10 μm·s−1) in node Iwas almost the same as that estimated in the root in a previousstudy (4.48 ± 0.26 μm·s−1) (11). These are also comparable to theparameter of Lsi3 in node I (6.19 ± 2.17 μm·s−1). By contrast, theestimated permeability of Lsi6 (136.25 ± 68.49 μm·s−1) was muchhigher than that estimated for Lsi1 in the root (16.38 ± 3.40 μm·s−1)(11). When the permeability parameter of Lsi6 was replaced bythat of Lsi1 in the root to mimic no XTC development (noXTC), the estimated contribution of XTCs to Si distribution wasrelatively small (Fig. 4). Lsi6 is an aquaporin-like passive chan-nel, which has much faster transport capacity compared withother types of active transporters (15). Therefore, transport ca-pacity of Lsi6 for Si in the node is possibly high enough even ifthe XTC was not developed.The xylem area of EVB is a mosaic of numerous vessels and
parenchyma cells, which has more than 10× larger area com-pared with connecting VBs in both internode and leaf (2–4). Thisenlargement has two meanings: slowdown of the xylem mass flowand increase of space for expressing transporters. Both of themmay contribute to efficient intervascular transfer. However, oursimulation showed that if the velocity of xylem mass flow in EVB
Xyl
em o
f DV
B
Xyl
em o
f EV
B
NVA
Flag leafPanicle
XTCPCB BS
Lsi2Lsi3
Apoplastic barrier
Lsi6
: Apoplast
: Influx transport of Si by Lsi6
: Parenchyma cell bridgePCB
: Xylem transfer cellXTC
: Xylem/apoplastic flow of Si
: Symplastic flow of Si
Nod
e I
: Symplast
: efflux transport of Si by Lsi2
: efflux transport of Si by Lsi3
: Plasmodesma
: Enlarged vascular bundleEVB: Diffuse vascular bundleDVB
: Bundle sheath of EVBBS
: Nodal vascular anastomosisNVA
: Apoplastic barrier
EVBDVB
Fig. 5. Schematic presentation of intervascular transfer pathway of Si in node Isilicon in the xylem of EVB is unloaded by Lsi6 polarly localized at the XTCs,followed by symplastical transfer through plasmodesmata to the bs cells of EVBwith an apoplastic barrier. Part of Si is then released by Lsi2 localized at thedistal side of bs to the apoplast of PCB and transported to the xylem of DVB,whereas the remaining Si is simplistically transferred to the PCB and exportedand reloaded to the xylem of DVB by Lsi3. The cooperation of these threetransporters across the apoplastic barrier is required for the intervasculartransfer of Si and subsequently for hyperaccumulation of Si in the rice husk.Modeling shows that a physical barrier on the bs and development of EVB arealso required for efficient intervascular transfer of Si. For details, see text.
Yamaji et al. PNAS | September 8, 2015 | vol. 112 | no. 36 | 11405
PLANTBIOLO
GY

was increasing 10×, the distribution ratio of Si to the panicles wasonly slightly decreased, probably due to the invariable concen-tration of Si input in this model (Fig. 4 and Fig. S8). By contrast,when the transport capacity was simply decreased to 1/3 to mimicreduced space for transporters, the distribution of Si to thepanicles was significantly decreased (Fig. S9). This simulationsuggests that increased transport capacity due to enlargementof EVB is also important for the efficient intervascular transfer of Si.In conclusion, we identified a complete set of players involved
in the intervascular transfer of Si in the node, which is requiredfor the preferential distribution of Si to the husk. Transporterslocalized at the different cell layers are responsible for move-ment of Si from EVB to DVB, and the distinct structure of thenode helps to enhance this movement. Although we focused onSi distribution in uppermost node I in this study, distributioncontrols of all mineral nutrients at each node throughout thedeveloping processes are required. Furthermore, graminaceousplants are composed of repeated nodes, and the structure ofeach node is basically similar. Therefore, results shown in thisstudy provide a model for studying the systems of preferentialdistribution of all minerals in graminaceous plants in future.
Materials and MethodsSeeds of T-DNA insertion mutants of Lsi6 (PFG_4A-01373) (13) and Lsi3(PFG_3C-00346; Fig. S1) [Rice T-DNA Insertion Sequence Database (RISD), cbi.khu.ac.kr/RISD_DB.html] and their wild type (WT1, cv. Dongjin) and lsi2 single-nucleotide substitution mutant (10) and its wild type (WT2, cv. Taichung-65)were grown hydroponically in a half-strength Kimura B nutrient solution(24). For expression analysis of Si transporter genes, different organs of WT1including unelongated stem, node I to III, peduncle, rachis, and caryopsiswere sampled at the flowering stage and subjected to semiquantitative andquantitative RT-PCR with gene-specific primers shown in Table S1. Effluxtransport activity assay of Lsi3 for Si in Xenopus oocytes was performed as
described previously (10, 25). Furthermore, the transport activity of Lsi3 wasdetermined by transforming the promoter region of Lsi2 (2.0 kb) (25) andORF of Lsi3 into lsi2 mutant using an Agrobacterium-mediated trans-formation system. The subcellular localization of Lsi3 was investigated byintroducing plasmids carrying GFP-Lsi3 fusion gene and DsRed-monomer(Clontech) into onion epidermal cells using Helios Gene Gun system (Bio-Rad). Immunostaining was performed for observing localization of Lsi6, Lsi2,and Lsi3 in node I as described previously (22). Fluorescence signals wereobserved with a confocal laser-scanning microscopy (LSM700; Carl Zeiss).Apoplastic permeability test in node I was performed according to Shionoet al. (16). For stem feeding experiment, WT1, WT2, and three mutants (lsi6,lsi2, and lsi3) were grown hydroponically without Si until flowering and cutat the internode II below the node I. Nutrient solution containing 2 mM Siand 50 μM of Rb and Sr was fed from the cut end for 24 h. Concentrations ofSi, Rb, and Sr in each part were determined by inductively coupled plasmamass spectrometry (7700X; Agilent Technologies).
For mathematical modeling of Si transport in node I, we divided the node Iby 32 grid points in a transverse direction and 48 grid points in a longitudinaldirection, with each grid point representing 4 × 40 μm (Fig. S6). We simulatedthe transfer of Si in the region by using a diffusion equation on the 2D grids(11, 26) and a 2D explicit finite difference method.
For more detail, see SI Materials and Methods.
ACKNOWLEDGMENTS. We thank Sanae Rikiishi and Akemi Morita for theirtechnical assistance. We also thank Drs. Akiko Satake and Masayuki Yokozawafor their valuable discussion and Drs. Shunsaku Nishiuchi and Mikio Nakazonofor kindly providing the protocol of the permeability staining. This research wassupported by a Grant-in-Aid for Scientific Research on Innovative Areas fromthe Ministry of Education, Culture, Sports, Science and Technology of Japan[22119002 and 24248014 to J.F.M., 70452737 to G.S.], and Joint Research Pro-gram implemented at the Institute of Plant Science and Resources, OkayamaUniversity, Japan (G.S.). Computations were carried out on a cluster system atthe Agriculture, Forestry and Fisheries Research Information Technology Centerfor Agriculture, Forestry and Fisheries Research, Ministry of Agriculture, Forestryand Fisheries, Japan.
1. Marschner P (2011) Mineral Nutrition of Higher Plants (Academic, London), 3rd Ed.2. Yamaji N, Ma JF (2014) The node, a hub for mineral nutrient distribution in grami-
naceous plants. Trends Plant Sci 19(9):556–563.3. Hoshikawa K (1989) The Growing Rice Plant: An Anatomical Monograph (Nobunkyo,
Tokyo).4. Kawahara H, Chonan N, Matsuda T (1974) Studies on morphogenesis in rice plants 7.
The morphology of vascular bundles in the vegetative nodes of the culm. Jpn J CropSci 43(3):389–401.
5. Chonan N, Kawahara H, Matsuda T (1985) Ultrastructure of elliptical and diffusebundles in the vegetative nodes of rice. Jpn J Crop Sci 54(4):393–402.
6. Ma JF, Yamaji N (2006) Silicon uptake and accumulation in higher plants. Trends PlantSci 11(8):392–397.
7. Ma JF, Takahashi E (2002) Soil, Fertilizer, and Plant Silicon Research in Japan (Elsevier,Amsterdam).
8. Tamai K, Ma JF (2008) Reexamination of silicon effects on rice growth and productionunder field conditions using a low silicon mutant. Plant Soil 307(1-2):21–27.
9. Ma JF, et al. (2006) A silicon transporter in rice. Nature 440(7084):688–691.10. Ma JF, et al. (2007) An efflux transporter of silicon in rice. Nature 448(7150):209–212.11. Sakurai G, et al. (2015) In silico simulation modeling reveals the importance of the
Casparian strip for efficient silicon uptake in rice roots. Plant Cell Physiol 56(4):631–639.
12. Mitani N, Ma JF, Iwashita T (2005) Identification of the silicon form in xylem sap of rice(Oryza sativa L.). Plant Cell Physiol 46(2):279–283.
13. Yamaji N, Mitatni N, Ma JF (2008) A transporter regulating silicon distribution in riceshoots. Plant Cell 20(5):1381–1389.
14. Yamaji N, Ma JF (2009) A transporter at the node responsible for intervasculartransfer of silicon in rice. Plant Cell 21(9):2878–2883.
15. Mitani N, Yamaji N, Ma JF (2008) Characterization of substrate specificity of a ricesilicon transporter, Lsi1. Pflugers Arch 456(4):679–686.
16. Shiono K, et al. (2014) RCN1/OsABCG5, an ATP-binding cassette (ABC) transporter, is re-quired for hypodermal suberization of roots in rice (Oryza sativa). Plant J 80(1):40–51.
17. Kuppelwieser H, Feller U (1991) Transport of Rb and Sr to the ear in mature, excised
shoots of wheat: Effects of temperature and stem length on Rb removal from the
xylem. Plant Soil 132(2):281–288.18. Yamaji N, Chiba Y, Mitani-Ueno N, Feng Ma J (2012) Functional characterization of a
silicon transporter gene implicated in silicon distribution in barley. Plant Physiol
160(3):1491–1497.19. Zheng L, Yamaji N, Yokosho K, Ma JF (2012) YSL16 is a phloem-localized transporter
of the copper-nicotianamine complex that is responsible for copper distribution in
rice. Plant Cell 24(9):3767–3782.20. Deng F, Yamaji N, Xia J, Ma JF (2013) A member of the heavy metal P-type ATPase
OsHMA5 is involved in xylem loading of copper in rice. Plant Physiol 163(3):
1353–1362.21. Yamaji N, Xia J, Mitani-Ueno N, Yokosho K, Feng Ma J (2013) Preferential delivery of
zinc to developing tissues in rice is mediated by P-type heavy metal ATPase OsHMA2.
Plant Physiol 162(2):927–939.22. Yamaji N, Sasaki A, Xia JX, Yokosho K, Ma JF (2013) A node-based switch for pref-
erential distribution of manganese in rice. Nat Commun 4:2442.23. Song WY, et al. (2014) A rice ABC transporter, OsABCC1, reduces arsenic accumulation
in the grain. Proc Natl Acad Sci USA 111(44):15699–15704.24. Ma JF, Goto S, Tamai K, Ichii M (2001) Role of root hairs and lateral roots in silicon
uptake by rice. Plant Physiol 127(4):1773–1780.25. Mitani N, Chiba Y, Yamaji N, Ma JF (2009) Identification and characterization of maize
and barley Lsi2-like silicon efflux transporters reveals a distinct silicon uptake system
from that in rice. Plant Cell 21(7):2133–2142.26. Grieneisen VA, Xu J, Marée AFM, Hogeweg P, Scheres B (2007) Auxin transport is suf-
ficient to generate a maximum and gradient guiding root growth. Nature 449(7165):
1008–1013.27. Ma JF, Higashitani A, Sato K, Takeda K (2003) Genotypic variation in silicon concen-
tration of barley grain. Plant Soil 249(2):383–387.
11406 | www.pnas.org/cgi/doi/10.1073/pnas.1508987112 Yamaji et al.