optochin-resistant streptococcus pneumoniae : phenotypic...
TRANSCRIPT

1
Optochin-Resistant Streptococcus pneumoniae: Phenotypic and Molecular 1
Characterization of Isolates from Brazil with Description of Five Novel 2
Mutations in the atpC Gene 3
4
Tatiana C.A. Pinto,a Aline R.V. Souza,a Sandrine E.C.M. de Pina,a Natália S. Costa,a 5
Armando A. Borges Neto,a Felipe P.G. Neves,b Vânia L.C. Merquior,c Cícero A.G. 6
Dias,d José M. Peralta,a Lúcia M. Teixeiraa 7
8
Instituto de Microbiologia Paulo de Goes, Universidade Federal do Rio de Janeiro, Rio de 9
Janeiro, RJ 21941-902, Brazila; Instituto Biomédico, Universidade Federal Fluminense, 10
Niterói, RJ 24210-130, Brazilb; Departamento de Microbiologia, Imunologia e 11
Parasitologia, Universidade do Estado do Rio de Janeiro, Rio de Janeiro, RJ 20551-030, 12
Brazilc; Departamento de Microbiologia e Parasitologia, Universidade Federal de Ciências 13
da Saúde de Porto Alegre, Porto Alegre, RS 90050-170, Brazild 14
15
Address correspondence to Lúcia M Teixeira, [email protected] 16
17
Running Title: Mutations in Optochin-Resistant S. pneumoniae 18
Copyright © 2013, American Society for Microbiology. All Rights Reserved.J. Clin. Microbiol. doi:10.1128/JCM.01168-13 JCM Accepts, published online ahead of print on 24 July 2013
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

2
ABSTRACT 19
Optochin (Opt) susceptibility is largely used for the identification of Streptococcus 20
pneumoniae (Sp) in diagnostic laboratories. Opt-resistant (Optr) Sp isolates have been 21
reported, however, indicating the potential for misidentification of this important pathogen. 22
Point mutations in the atpC gene have been associated with the emergence of Optr Sp but 23
data on the characterization of such atypical variants of Sp are still limited. The present 24
report describes the results of a polyphasic approach to identify and characterize twenty-six 25
Optr Sp isolates recovered from patients or carriers living in Brazil. Sixteen isolates 26
consisted of heterogeneous populations and ten isolates were homogeneously Optr. They 27
had different serotypes and antimicrobial susceptibility profiles. They also presented 28
diverse genetic characteristics, as indicated by PFGE, MLVA and pspA gene typing. Except 29
for Opt MICs (4 to 64-fold higher among Optr variants), Optr and Opts subpopulations 30
originating from the same culture had identical characteristics. Sequencing of the atpC gene 31
of the Optr variants revealed thirteen different nucleotide changes distributed among eight 32
different codons. Changes in codon 49 were the most frequent, suggesting that this might 33
be a hot spot for optochin resistance-conferring mutations. On the other hand, five novel 34
types of mutations in the atpC gene (Met13Ile, Gly18Ser, Gly20Ala, Ala31Val, Ala49Gly) 35
were identified. In silico prediction modeling indicated that the atpC gene mutations 36
corresponded to alterations in the transmembrane region of the ATPase, leading to a higher 37
hydrophobicity profile in α-helix 1 and to a lower hydrophobicity profile in α-helix 2. 38
39
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

3
INTRODUCTION 40
Streptococcus pneumoniae (Sp) remains as one of the most important agents of infectious 41
diseases worldwide and a leading cause of invasive and non-invasive infections in children 42
and in the elderly (1). Nasopharyngeal carriage is considered the main reservoir of Sp in 43
nature and a condition of paramount importance for the continuous circulation of this 44
bacterial species among human populations. Due to the significant morbidity and mortality 45
rates associated with pneumococcal diseases, rapid and accurate diagnosis is essential and 46
frequently relies on the isolation and identification of the etiological agent. For this 47
purpose, optochin (Opt) susceptibility testing has been widely used as the primary or even 48
the only test for the presumptive identification of pneumococci, due to the ability to 49
differentiate Sp from the other α-hemolytic streptococci (2-4). 50
Nevertheless, thirty years after the introduction of the Opt susceptibility test as a 51
diagnostic tool (5), the first optochin-resistant (Optr) Sp isolate was described (6). Since 52
then, the isolation of Optr Sp from a variety of clinical sources has sporadically been 53
reported from different locations, with an apparent increasing incidence in the last decade 54
(2, 3, 7-12). The occurrence of such atypical variant is a potential cause of misidentification 55
of this important agent, raising questions about the accuracy of laboratory diagnosis when a 56
single identification procedure based on Opt susceptibility testing is used. 57
Previous studies have shown that single point mutations in the c-subunit (Table 1), 58
and less commonly in the a-subunit, of the atpCAB operon that codes for the molecular 59
target of optochin, the transmembrane F0 F1 ATPase, are present in Optr Sp strains and may 60
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

4
be associated with this unusual phenotype (2, 7, 9, 10, 12-14). Although still uncertain, it 61
has been suggested that use of antimalarial chemotherapy in endemic areas (14), as well as 62
exposition to subinhibitory concentrations of penicillin (15) may be related to the 63
emergence of such mutations. 64
Studies concerning the phenotypic and genetic characteristics of atypical Sp variants 65
expressing resistance to Opt are still limited and may contribute to a better understanding of 66
the mechanisms involved in the emergence, as well as the biological and epidemiological 67
impacts of such resistance trait. The present report describes the results of a polyphasic 68
approach applied to characterize twenty-six Optr Sp isolates recovered from patients and 69
nasopharyngeal carriers living in Brazil. The study encompasses extensive phenotypic and 70
genotypic characterization of Optr Sp isolates presenting a variety of atpC gene mutations, 71
including the description of five novel types of mutations associated with resistance to Opt. 72
Additionally, in silico prediction modeling was performed to evaluate the impact of 73
mutations on the structure and hydrophobicity profile of the c-subunit of ATPase. 74
75
MATERIALS AND METHODS 76
Bacterial strains and identification tests. Twenty-six Optr Sp strains were included in the 77
present study. Among them, four (Sp 910, Sp 913, Sp 917 and Sp 1008) were previously 78
characterized by using a variety of conventional and genetic tests (9). They were included 79
in the present study for evaluation of additional characteristics (determination of Opt MIC, 80
MLVA type and PspA family). All the isolates were recovered during surveillance studies 81
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

5
performed by our group, or were received from different health institutions for 82
confirmation of the identification. Isolates obtained from diseased individuals were 83
recovered from clinical samples taken as part of standard patient care procedures and they 84
do not need an ethical approval for their use. Isolates from carriage studies were recovered 85
from clinical samples collected as approved by the ethics committees of the institutions 86
involved. The isolates were subjected to phenotypic identification tests, including 87
observation of colonial morphology and type of hemolytic activity on blood agar plates; 88
cellular characteristics as observed after Gram stain; optochin susceptibility, bile-solubility, 89
and latex agglutination tests. PCR tests for detection of the lytA, ply and psaA genes were 90
also performed. S. pneumoniae strains ATCC 49619 and ATCC BAA-255 (R6), and S. 91
mitis SS-429 were included as reference strains in all the assays. 92
Optochin susceptibility testing. Opt susceptibility was determined by disk 93
diffusion testing according to standard procedures (4). Optochin disks (BBL Taxo P Discs, 94
BD, Sparks, MD, USA) were applied to the surface of 5% sheep blood agar plates (Plast 95
Labor, Rio de Janeiro, RJ, Brazil) streaked with the isolate being tested. After overnight 96
incubation at 35°C under both 5% CO2 and conventional atmospheres (16), growth 97
inhibition zones around the disks were measured. Isolates displaying inhibition zones ≥14 98
mm in diameter were identified as susceptible, while strains showing zones �14 mm, or 99
zones ≥14 mm but containing colonies inside, were considered resistant. The optochin 100
(Sigma Chemical Co., St. Louis, MO, USA) minimal inhibitory concentration (MIC) values 101
were determined by the agar dilution method (2). Briefly, bacterial suspensions were 102
prepared in 0.9% saline (Sigma) from overnight cultures and adjusted to achieve the 0.5 103
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

6
McFarland turbidity standard. Suspensions were inoculated onto Müeller-Hinton agar 104
containing 5% sheep blood (Plast Labor, Rio de Janeiro, RJ, Brazil) and varying 105
concentrations of optochin (0.5 to 256 μg/mL), followed by incubation at 35°C for 20–24h 106
under 5% CO2 atmosphere. The MIC was defined as the lowest concentration of optochin 107
that inhibited visible growth of the isolate. S. pneumoniae ATCC 49619 and R6 were 108
included as susceptible controls, while S. mitis SS-429 was included as a resistant control in 109
the Opt susceptibility tests. 110
Bile-solubility testing. Bile-solubility tube testing was performed as previously 111
described (4). Briefly, heavy bacterial suspensions were prepared in 0.9% saline (Sigma) 112
from overnight cultures. The suspensions were placed into 2 tubes, and an equal volume of 113
2% deoxycholic acid (Sigma) solution was added to one tube (test tube) while an equal 114
volume of 0.9% saline was added to the other (control tube). Tubes were incubated at 35°C 115
for up to 2 h. Complete visual clearing of the turbidity in the deoxycholate-containing tube, 116
but not in the saline control tube, indicated a positive test. 117
Latex agglutination tests. The isolates were tested for the presence of capsular 118
polysaccharide antigens by latex agglutination testing using the Slidex Pneumo kit 119
according to the manufacturer’s instructions (bioMérieux, Marcy l'Etoile, France). 120
Determination of capsular type. The capsular types were determined either by 121
multiplex PCR using the Latin America scheme (17) or by the standard Quellung reaction 122
with type-specific antisera prepared at the Centers for Disease Control and Prevention 123
(CDC), as earlier described (18). 124
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

7
Antimicrobial susceptibility testing. Antimicrobial susceptibility was evaluated by 125
the agar diffusion method according to the CLSI recommendations and interpretative 126
criteria (19, 20). Antimicrobials tested included chloramphenicol, clindamycin, 127
erythromycin, levofloxacin, oxacillin, rifampicin, trimethoprim/sulfamethoxazole, 128
tetracycline and vancomycin (Oxoid, Basingstoke, Hampshire, United Kingdom). Penicillin 129
MICs were determined by using Etest strips (AB Biodisk, Solna, Stockholm, Sweden). S. 130
pneumoniae ATCC 49619 was used for quality control. 131
Preparation of bacterial DNA. DNAs for all PCR reactions were obtained by 132
using the Chelex® 100 resin (Bio-Rad, Hercules, CA, USA) method. Briefly, a loopful of 133
overnight cultures was suspended in a solution containing 5% Chelex® and proteinase K 20 134
mg/mL (Invitrogen, Life Technologies, Carlsbad, CA, USA). After incubation at 56°C for 1 135
h, followed by incubation at 95°C for 10 min, they were centrifuged and the supernatant 136
was used as the template for all the subsequent PCR reactions. 137
Detection of resistance- and virulence-associated genes. The presence of the 138
macrolide-resistance determinants ermA, ermB and mef (21), the tetracycline-resistance 139
genes tetK, tetL, tetM and tetO (22), as well as the virulence-associated genes ply (coding 140
for pneumolisin), lytA (coding for autolysin) and psaA (coding for pneumococcal surface 141
antigen A) (23, 24) was investigated by PCR using protocols previously described and an 142
automated Veriti 96-well thermal cycler (Applied Biosystems, Inc, Carlsbad, CA, USA). 143
Pulsed-Field Gel Electrophoresis (PFGE) analysis. Genomic DNA was prepared 144
in agarose plugs as described by Teixeira et al. (25), with the following modifications: 145
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

8
bacteria were grown in blood agar plates for 8 to 10 h, plugs were prepared with low-146
melting-temperature agarose at 2.5% (NuSieve GTG agarose, FMC BioProducts, Rockland, 147
ME, USA) and were washed 8 times before restriction. After macrorestriction using SmaI 148
(New England Biolabs, Ipswich, MA, USA), the fragments were separated in a CHEF-DR 149
III system (Bio-Rad) using the parameters recommended by the Pneumococcal Molecular 150
Epidemiology Network (PMEN; http://www.sph.emory.edu/PMEN/index.html). The 151
restriction profiles were analyzed by using the BioNumerics software version 6.6 (Applied 152
Maths, Ghent, East Flanders, Belgium). A dendrogram was created using the unweighted-153
pair group method with arithmetic mean (UPGMA) and the Dice similarity coefficient, 154
with optimization and position tolerance settings of 0.5% and 1.3%, respectively. Profiles 155
showing more than 80% of similarity were considered as belonging to the same cluster. 156
PFGE profiles of the 43 clones described by the PMEN were also included in the in silico 157
analysis for comparative purposes. 158
Multiple Locus VNTR Analysis (MLVA). Eight VNTRs (Spneu15, Spneu17, 159
Spneu25, Spneu33, Spneu36, Spneu37, Spneu39 and Spneu42), selected (N. S. Costa et al., 160
unpublished data) among an initially suggested panel of 18, were amplified by PCR using 161
primers and a protocol described earlier (26). MLVA profiles were analyzed by using 162
BioNumerics software version 6.6 (Applied Maths), in order to construct a dendrogram by 163
the UPGMA and a diagram based on the Minimum Spanning Tree (MST) method. 164
Reference strain S. pneumoniae R6 was used as quality control for the PCR reactions. 165
pspA gene family typing. The pspA genes were detected by PCR using primers and 166
protocols previously described (27). Control strains for pspA family 1 (strains BG9739, 167
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

9
DBL1 and EF10197), family 2 (strains AC122, BG11703, BG7591 and ATCC 6303) and 168
family 3 (strain BG6380) were also included. Strains were categorized as non typeable 169
(NT) if negative results were obtained in 3 attempts of amplification. 170
Sequencing of the atpC gene. The gene coding for the c-subunit of ATPase was 171
amplified by PCR using primers and protocols previously described (2, 9, 13). The 172
fragment was purified using ExoSAP-IT® (USB Affymetrix, Cleveland, OH, USA) and the 173
sequences were obtained using an ABI 3130 Genetic Analyzer (Applied Biosystems). 174
Edition and alignment were performed with the BioEdit software version 7.0.9.0 (28), as 175
well as the construction of charts based on the Kyte & Doolittle Scale Mean 176
Hydrophobicity Profile. Using the amino acid sequences translated from the nucleotide 177
sequences obtained in the study, a predicted model for the c-subunit was designed by using 178
the resources available at the Protein Structure PREDition (PSIPRED) server 179
(http://bioinf.cs.ucl.ac.uk/psipred/; 29). Opts reference strains (S. pneumoniae ATCC 49619 180
and R6) were also included for comparative purposes. 181
182
RESULTS AND DISCUSSION 183
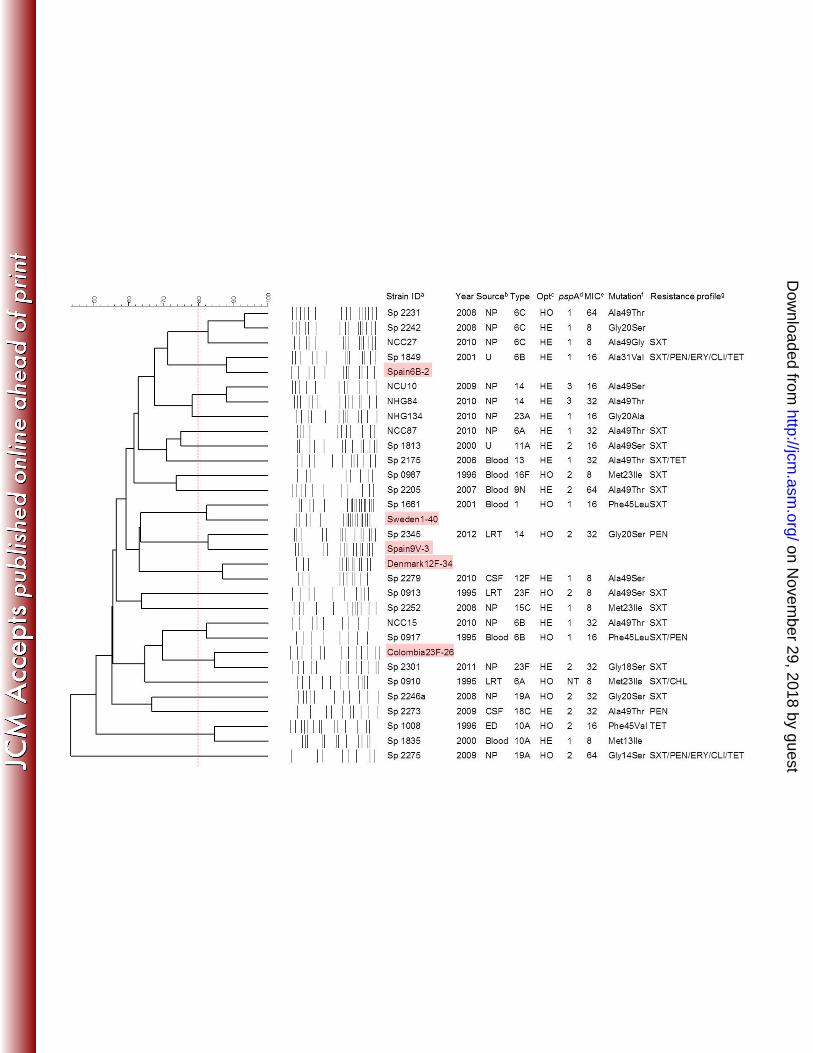
Identification of the isolates and characterization of Opt resistance. The twenty-six 184
Optr Sp isolates included in this study were recovered from different clinically significant 185
sources (14 isolates) or from nasopharyngeal secretions of healthy carriers (12 isolates) 186
between 1995 and 2012, as shown in Fig. 1. All the isolates had the following 187
characteristics: they were gram-positive catalase-negative cocci; presented α-hemolysis on 188
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

10
blood agar plates; gave positive results in the bile solubility and in the latex agglutination 189
tests; possessed the lytA, ply and psaA genes; and were resistant to Opt. 190
Opt susceptibility testing, performed either under CO2-enriched or conventional 191
atmospheres, revealed the occurrence of two phenotypes among the Optr Sp isolates 192
analyzed (Fig. 1). The prevalent phenotype comprised sixteen strains and was expressed as 193
a typical inhibition zone of ≥ 14mm around the Opt disk with colonies inside, representing 194
a heterogeneous population. The subpopulations were discriminated based on the Opt 195
susceptibility and the subsequent experiments were carried out separately for both 196
subpopulations. The second phenotype was observed for the remaining ten Optr Sp strains 197
that had no zone of inhibition around the Opt disk, representing homogeneous resistant 198
populations. The finding of two different Optr phenotypes among Sp isolates has been 199
reported before, among isolates from Portugal (11) and from the United States (2) with 200
different distributions. Among isolates from Portugal, a predominance of homogeneous 201
Optr populations was observed, while the majority of the isolates from the United States 202
were reported to be composed of heterogeneous populations as also observed in the present 203
study. Opt MICs ranged from 8 to 64 μg/mL for the Optr Sp strains (Fig. 1), which is in 204
agreement with earlier reports on Opt resistance (2, 7, 11). On the other hand, the sixteen 205
Opts subpopulations derived from heterogeneous cultures, as well as the susceptible 206
reference strains S. pneumoniae ATCC 49619 and R6, showed MIC values of 1 to 2 207
μg/mL, which were 4- to 64-fold lower than those obtained for their respective Optr 208
counterparts. Except for the unusual Optr phenotype, all the Sp strains analyzed showed 209
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

11
phenotypic and genetic characteristics of typical pneumococcal isolates, as they were bile-210
soluble, produced capsular antigens, and carried the ply, lytA and psaA genes. 211
Distribution of capsular types. The isolates tested belonged to a variety of 212
capsular types (Fig. 1). Most of the sixteen different capsular types (1, 6A, 6B, 6C, 9N, 213
10A, 11A, 12F, 13, 14, 15C, 16F, 18C, 19A, 23A, 23F) observed among the isolates 214
included in the present study correspond to types commonly found in our region (30, 31), 215
reinforcing the concept that serotype distribution among Optr Sp strains is mostly dependent 216
on the epidemiological profile of each area (2, 3, 10-12). Identical serotypes were observed 217
for the Optr and Opts subpopulations derived from the same culture. Interestingly, among 218
the eight isolates belonging to serogroup 6, three belonged to the recently described variant 219
6C (32), that seems to have emerged during the post-vaccination era in developed countries 220
(33, 34). Moreover, five of the capsular types identified (1, 6B, 14, 18C and 23F), 221
comprising a total of ten strains, are included in the 10-valent pneumococcal conjugate 222
vaccine (PCV-10), that has been incorporated into the Brazilian Immunization Programme 223
in 2010 (http://portal.saude.gov.br/portal/saude/visualizar_texto.cfm?idtxt=21462). 224
Antimicrobial susceptibility profiles. The Optr Sp strains were susceptible to most 225
of the antimicrobial agents tested (Fig. 1) with only two of them presenting a multidrug-226
resistance profile (including simultaneous resistance to clindamycin, erythromycin, 227
penicillin, tetracycline and trimethoprim/sulfamethoxazole), suggesting that Opt resistance 228
is not associated with increasing or specific resistance to other drugs. Resistance to both 229
erythromycin and clindamycin was detected in 2 (7.7%) strains, and it was associated with 230
the simultaneous presence of the ermB and mef genes. Resistance to tetracycline was 231
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

12
observed in four (15.4%) isolates carrying the tetM gene. All the resistance-associated 232
genes detected are commonly found among conventional Opts Sp strains as well (35). A 233
total of five (19.2%) isolates showed non-susceptibility to penicillin, displaying MICs 234
ranging from 0.32 to 2 μg/mL. Isolates belonging to serotypes 14 and 6B, recovered from 235
blood or sputum, had the higher penicillin MICs (0.5 or 2 μg/mL), while serotypes 18C and 236
19A isolates, obtained from nasopharynx or cerebrospinal fluid (CSF), had the lower MICs 237
(0.32 or 0.38 μg/mL). As observed for serotype distribution, antimicrobial susceptibility 238
among Optr Sp isolates included in this study is probably reflecting the epidemiological 239
profile of the region. In addition, Optr and Opts subpopulations originating from the same 240
culture showed identical profiles of antimicrobial susceptibility. 241
Genetic diversity evaluated by PFGE, MLVA and pspA gene family typing. 242
PFGE (Fig. 1) and MLVA (Fig. 2) revealed a high degree of genetic diversity, generating 243
26 and 25 different profiles, respectively, among the twenty-six Optr isolates analyzed, and 244
indicating that Opt resistance is not due to clonal dissemination, in accordance with data 245
from other regions (2, 3, 9, 10, 12). Moreover, PFGE revealed that five Optr isolates were 246
respectively related to five internationally disseminated clones recognized by the PMEN 247
(http://www.sph.emory.edu/PMEN/index.html) (Fig. 1). Among them, four (Spain6B-2, 248
Spain9V-3, Colombia23F-26 and Sweden1-40) are known to be prevalent in Brazil (31, 36, 249
37). Except for one, those five Optr Sp strains were recovered from cases of invasive 250
pneumococcal disease, and two of them were non-susceptible to penicillin, with one being 251
multidrug resistant. Likewise, the occurrence of strains related to some of the PMEN 252
clones, including Spain9V-3 and Colombia23F-26, were also reported among Portuguese Optr 253
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

13
Sp isolates (3, 11). Heterogeneity was also revealed by analysis of the pspA genes. Thirteen 254
Optr Sp isolates were typed as pspA family 1, while ten belonged to pspA family 2. The 255
pspA family 3 was observed for two strains and one strain had no detectable pspA genes 256
(Fig. 1). These results show that distribution of pspA gene families among Optr Sp strains 257
follows a profile quite similar to that seen among Sp isolates in general, with predominance 258
of pspA families 1 and 2 (38). Moreover, Opts subpopulations derived from heterogeneous 259
cultures showed PFGE profiles, MLVA types and pspA gene family types identical to their 260
respective Optr counterparts. Although this is, to our knowledge, the first report on the use 261
of MLVA and pspA gene typing to characterize Optr isolates, indistinguishable PFGE 262
profiles between Optr and Opts subpopulations deriving from a single culture have been 263
observed by others (2, 8, 11). 264
Nucleotide sequences of the atpC gene. Sequencing of the atpC gene, which codes 265
for the c-subunit of the pneumococcal ATPase, revealed single-base substitutions leading to 266
amino acid modifications in all the twenty-six Optr Sp strains analyzed (Fig. 1). Thirteen 267
different nucleotide changes distributed among eight different codons were observed (see 268
Fig. S1 in the supplemental material). Five of them, comprising six strains, consisted of 269
novel alterations: Met13Ile, Gly18Ser, Gly20Ala, Ala31Val, and Ala49Gly. Among them, 270
three mutation locations (codons 13, 18 and 31) were never reported before among Optr Sp 271
isolates obtained from clinical sources. A summary with all the mutations described among 272
Optr Sp strains until this date is presented in Table 1, including those described for the first 273
time in the present study. Mutations obtained by in vitro transformation or induction 274
experiments (10, 14) were not included. 275
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

14
The mutations observed were randomly distributed among Optr Sp strains derived 276
from either heterogeneous or homogeneous populations (Fig. 1). Therefore, no correlation 277
was observed between the Optr phenotype and the type of mutation. Likewise, no 278
correlation with Opt MICs was noted either. On the other hand, no mutations were 279
observed in the atpC sequences of Opts subpopulations derived from heterogeneous 280
cultures, as their sequences were indistinguishable from those obtained for S. pneumoniae 281
strains ATCC 49619 and R6 (see Figure S1 in the supplemental material), reinforcing the 282
association between atpC mutations and Opt resistance. The fact that Optr and Opts 283
subpopulations were only differentiated on the basis of the Opt MICs and the atpC gene 284
sequencing suggests that an originally homogeneous Opts population might have undergone 285
a point mutation resulting in two different types of cells. 286
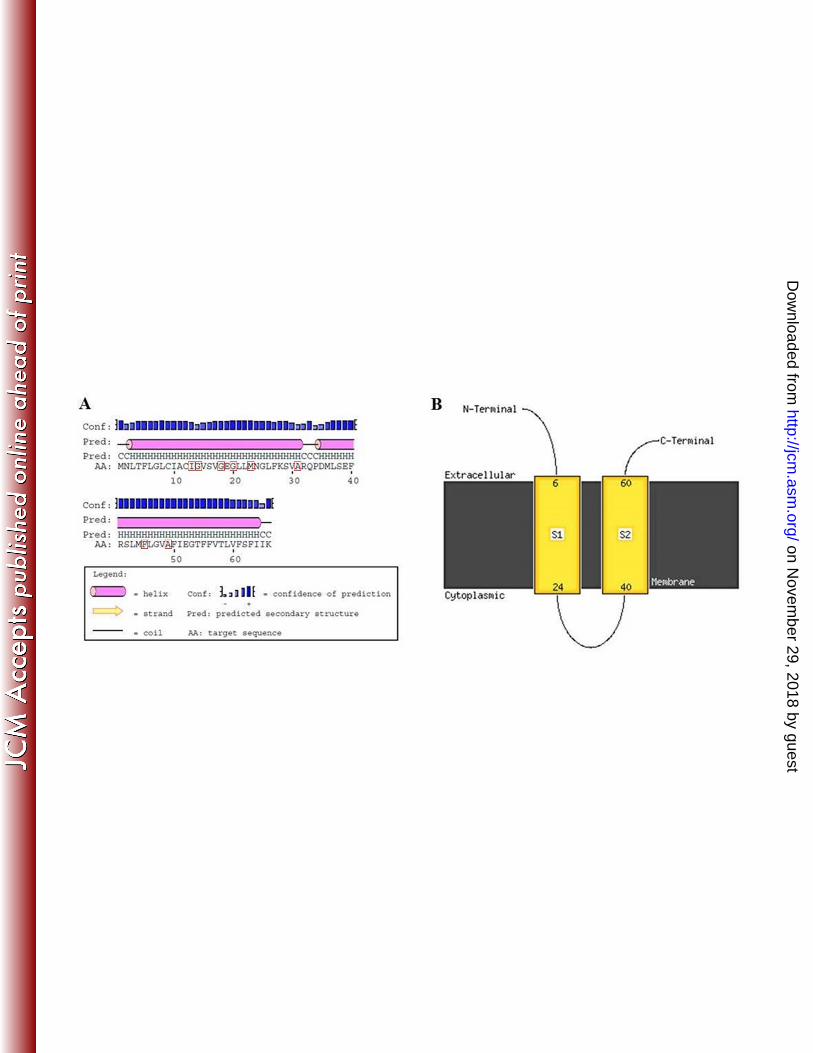
The c-subunit of the ATPase is known to consist of 66 amino acids distributed in 287
two antiparallel α-helixes linked by a conserved cytoplasmic loop (13). Previous studies 288
have shown that mutations leading to Opt resistance usually occur in the transmembrane 289
portion of the α-helixes, as it does not interfere with the activity of the whole enzyme (14). 290
Indeed, all the altered codons identified in this study are located in the transmembrane 291
region according to the predicted model of the subunit (Fig. 3). Eleven strains presented 292
alterations in the α-helix 1 of the c-subunit (codons 13, 14, 18, 20, 23 or 31) and fifteen in 293
the α-helix 2 (codons 45 or 49), among which twelve isolates (accounting for 48% of all 294
Optr Sp strains analyzed) had mutations in codon 49 (Table 1 and Fig. 1). 295
It was noticeable that all the atpC gene mutations identified in the present study 296
generated codifying sequences, involving amino acid replacement, and no termination 297
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

15
codons that would inhibit expression of atpC due to early termination were detected. Single 298
base substitutions are likely to occur randomly along the atpC gene sequence over time, 299
including synonymous or even deleterious mutations. However, those that are evolutionary 300
neutral, or maybe advantageous, and that lead to phenotypic variation, as illustrated by 301
optochin resistance, are possibly the only ones eventually detected. 302
The prevalence of mutations in codon 49 (Table 1) is noteworthy, leading to 303
speculations on its role as a hot-spot for mutations associated with resistance to Opt. No 304
peculiar characteristics around codon 49 that could predispose this part of the atpC gene to 305
frequent mutations, such as a higher incidence of repeated nucleotides, were observed in 306
the present work. On the other hand, in the predicted folded structure of ATPase, certain 307
amino acids, including those designated by codon 49, are placed next to residues of 308
glutamic or aspartic acid, two non-essential amino acids, that are believed to represent the 309
central portion of α-helix 2 and the binding site for optochin molecules (13). In terms of 310
natural selection and evolution, especially when considering optochin and its analogues as 311
selective pressures, codons producing such amino acids, as codon 49, could represent 312
preferential positions for preserved non-synonymous genetic alterations associated with 313
phenotypic advantages. 314
Although the exact mechanisms leading to Opt resistance remain to be elucidated, it 315
is reasonable to infer that structural alterations in the folding of the protein generated as a 316
result of a mutated atpC sequence, rather than the alterations in the amino acid sequences 317
solely, may, at least in part, account for resistance to optochin. It has also been suggested 318
that the alterations probably change the hydrophobicity profile of the molecule, causing the 319
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

16
loss of affinity to the drug (14). Indeed, among Optr Sp strains included in the present 320
study, changes in the α-helix 1 led, in general, to substitutions for amino acids with higher 321
hydrophobicity profiles (Fig. 4A), while in the α-helix 2 the replacements were for amino 322
acids with lower hydrophobicity profiles (Fig. 4B). Additional studies, however, are 323
required to elucidate the direct or indirect consequences of these changes in the interaction 324
with optochin. 325
Our data indicate that emergence of Optr Sp strains is not due to a clonal spread, 326
since they may possess a diversity of serotypes, phenotypes and genotypes. Also, it seems 327
to be related to a random mutational event, which is more likely to occur in the c-subunit of 328
ATPase, and more frequently in codon 49, near the center of the α-helix 2. The occurrence 329
of two types of Optr Sp strains additionally increases the complexity of this phenomenon, 330
although their individual importance or consequences are still unclear. Overall, our results 331
provide additional information and reinforce previous observations about Opt resistance 332
among Sp strains, contributing to the global and local pool of data on the characteristics of 333
these atypical isolates. Healthcare-associated professionals should be aware of the 334
occurrence of the unusual Optr Sp strains, especially considering that they can be isolated 335
from a variety of clinical sources, including asymptomatic carriers and patients with 336
invasive and non-invasive infections, leading to incorrect bacterial identification, and 337
ultimately to misinterpretation of carriage studies or imprecise diagnosis and treatment of 338
infectious conditions. 339
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

17
Nucleotide sequence accession numbers. The atpC gene sequences of the 22 340
isolates originally reported in the present study were deposited in the GenBank database 341
under accession numbers KC513927 to KC513948, as indicated in Fig. 2A. 342
343
ACKNOWLEDGMENTS 344
This work was supported in part by Coordenação de Aperfeiçoamento de Pessoal de Nível 345
Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico 346
(CNPq), Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), and 347
Ministério da Ciência e Tecnologia (MCT/PRONEX), Brazil. The authors would like to 348
thank Dr. Maria Cristina C. Brandileone (Instituto Adolfo Lutz, SP, Brazil) for providing 349
most of the control strains used in the pspA gene typing experiments, and Dr. Lesley 350
McGee (Centers for Disease Control and Prevention, Atlanta, GA, USA) for providing 351
representative strains of the PMEN clones. The authors declare that they have no competing 352
interests. 353
354
REFERENCES 355
1. Tan TQ. 2012. Pediatric invasive pneumococcal disease in the United States in the era of 356
conjugate pneumococcal vaccines. Clin. Microbiol. Rev. 25:409-419. 357
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

18
2. Pikis A, Campos JM, Rodriguez WJ, Keith JM. 2001. Optochin resistance in 358
Streptococcus pneumoniae: mechanism, significance and clinical implications. J. Infect. 359
Dis. 184:582-590. 360
3. Aguiar SI, Frias MJ, Santos L, Melo-Cristino J, Ramirez M, Portuguese Surveillance 361
Group for Study of Respiratory Pathogens. 2006. Emergence of optochin resistance 362
among Streptococcus pneumoniae in Portugal. Microb. Drug Resist. 12:239-245. 363
4. Spellerberg B, Brandt C. 2011. Gram-positive cocci, p 340-344. In Versalovic J, Carroll 364
KC, Jorgensen JH, Funke G, Landry ML, WarnockDW (ed), Manual of clinical 365
microbiology, 10th ed, vol 2. ASM Press, Washington, DC. 366
5. Bowen MK, Thiele LC, Stearman BD, Schaub IG. 1957. The optochin sensitivity test: a 367
reliable method for identification of pneumococci. J. Lab. Clin. Med. 49:641-642. 368
6. Kontiainen S, Sivonen A. 1987. Optochin resistance in Streptococcus pneumoniae strains 369
isolated from blood and middle ear fluid. Eur. J. Clin. Microbiol. 6:422-424. 370
7. Cogné N, Claverys JP, Denis F, Martin C. 2000. A novel mutation in the α-helix 1 of the 371
C subunit of the F1/F0 ATPase responsible for optochin resistance of a Streptococcus 372
pneumoniae clinical isolate. Diagn. Microbiol. Infect. Dis. 38:119-121. 373
8. Tsai HI, Hsueh PR, Teng LJ, Lee PI, Huang LM, Lee CY, Luh KT. 2000. Bacteremic 374
pneumonia caused by a single clone of Streptococcus pneumoniae with different optochin 375
susceptibilities. J. Clin. Microbiol. 38:458-459. 376
9. Dias CA, Agnes G, Frazzon APG, Kruger FD, d’Azevedo PA, Carvalho MG, Facklam 377
RR, Teixeira LM. 2007. Diversity of mutations in the atpC gene coding for the c subunit 378
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

19
of F0F1ATPase in clinical isolates of optochin-resistant Streptococcus pneumoniae from 379
Brazil. J. Clin. Microbiol. 45:3065-3067. 380
10. Cortes PR, Orio AGA, Regueira M, Piñas GE, Echenique JR. 2008. Characterization of 381
in vitro-generated and clinical optochin-resistant strains of Streptococcus pneumoniae 382
isolated from Argentina. J. Clin. Microbiol. 46:1930-1934. 383
11. Nunes S, Sá-Leão R, De Lancastre H. 2008. Optochin resistance among Streptococcus 384
pneumoniae colonizing healthy children in Portugal. J. Clin. Microbiol. 46:321-324. 385
12. Nagata M, Ueda O, Shobuike T, Muratani T, Aoki Y, Miyamoto H. 2012. Emergence 386
of optochin resistance among Streptococcus pneumoniae in Japan. Open J. Med. Microbiol. 387
2:8-15. 388
13. Fenoll A, Muñoz R, Garcia E, de la Campa AG. 1994. Molecular basis of the optochin-389
sensitive phenotype of pneumococcus: characterization of the genes encoding the F0 390
complex of the Streptococcus pneumoniae and Streptococcus oralis H(+)-ATPases. Mol. 391
Microbiol. 12:587-598. 392
14. Martín-Galiano AJ, Gorgojo B, Kunin CM, De La Campa AG. 2002. Mefloquine and 393
new related compounds target the F0 complex of the F0F1 ATPase of Streptococcus 394
pneumoniae. Antimicrob. Agents Chemother. 46:1680-1687. 395
15. Cortes PR, Piñas GE, Albarracin Orio AG, Echenique JR. 2008. Subinhibitory 396
concentrations of penicillin increase the mutation rate to optochin resistance in 397
Streptococcus pneumoniae. J. Antimicrob. Chemother. 62:973-977. 398
16. Arbique JC, Poyart C, Trieu-Cuot P, Quesne G, Carvalho MG, Steigerwalt AG, 399
Morey RE, Jackson D, Davidson RJ, Facklam RR. 2004. Accuracy of phenotypic and 400
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

20
genotypic testing for identification of Streptococcus pneumoniae and description of 401
Streptococcus pseudopneumoniae sp. nov. J. Clin. Microbiol. 42:4686-4696. 402
17. Dias CA, Teixeira LM, Carvalho MG, Beall B. 2007. Sequential multiplex PCR for 403
determining capsular serotypes of pneumococci recovered from Brazilian children. J. Med. 404
Microbiol. 56:1185-1188. 405
18. Sorensen UB. 1993. Typing of pneumococci by using 12 pooled antisera. J. Clin. 406
Microbiol. 31:2097-2100. 407
19. Clinical and Laboratory Standards Institute. 2009. Performance standards for 408
antimicrobial disk susceptibility tests; approved standard: M02-A10. CLSI, Wayne, PA. 409
20. Clinical and Laboratory Standards Institute. 2011. Performance standards for 410
antimicrobial susceptibility testing. Twenty-first informational supplement: M100-S21. 411
CLSI, Wayne, PA. 412
21. Sutcliffe J, Tait-Kamradt A, Wondrack L. 1996. Streptococcus pneumoniae and 413
Streptococcus pyogenes resistant to macrolide but sensitive to clindamycin: a common 414
resistance pattern mediated by an efflux system. Antimicrob. Agents Chemother. 40:1817-415
1824. 416
22. Ng LK, Martin I, Alfa M, Mulvey M. 2001. Multiplex PCR for the detection of 417
tetracycline resistant genes. Mol. Cell Probes 15:209-215. 418
23. Rudolph KM, Parkinson AJ, Black CM, Mayer LW. 1993. Evaluation of polymerase 419
chain reaction for diagnosis of pneumococcal pneumonia. J. Clin. Microbiol. 31:2661-2666. 420
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

21
24. Morrison KE, Lake D, Crook J, Carlone GM, Ades E, Facklam R, Sampson JS. 2000. 421
Confirmation of psaA in all 90 serotypes of Streptococcus pneumoniae by PCR and 422
potential of this assay for identification and diagnosis. J. Clin. Microbiol. 38:434-437. 423
25. Teixeira LM, Carvalho MG, Merquior VL, Steigerwalt AG, Brenner DJ, Facklam 424
RR. 1997. Phenotypic and genotypic characterization of Vagococcus fluvialis, including 425
strains isolates from human source. J. Clin. Microbiol. 35:2778-2781. 426
26. Koeck JL, Njanpop-Lafourcade BM, Cade S, Varon E, Sangare L, Valjevac S, 427
Vergnaud G, Pourcel C. 2005. Evaluation and selection of tandem repeat loci for 428
Streptococcus pneumoniae MLVA strain typing. BMC Microbiol. 5:66. doi:10.1186/1471-429
2180-5-66. 430
27. Pimenta FC, Ribeiro-Dias F, Brandileone MC, Miyaji EN, Leite LCC, Andrade ALS. 431
2006. Genetic diversity of PspA types among nasopharyngeal isolates collected during 432
ongoing surveillance study of children in Brazil. J. Clin. Microbiol. 44:2838-2843. 433
28. Hall T. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis 434
program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 41:95-98. 435
29. Buchan DW, Ward SM, Lobley AE, Nugent TC, Bryson K, Jones DT. 2010. Protein 436
annotation and modeling servers at University College London. Nucleic Acids Res. 437
38:W563-568. 438
30. Gouveia EL, Reis JN, Flannery B, Cordeiro SM, Lima JB, Pinheiro RM, Salgado K, 439
Mascarenhas AV, Carvalho MG, Beall BW, Reis MG, Ko AI. 2011. Clinical outcome of 440
pneumococcal meningitis during the emergence of pencillin-resistant Streptococcus 441
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

22
pneumoniae: an observational study. BMC Infect. Dis. 11:323. doi:10.1186/1471-2334-11-442
323. 443
31. Barroso DE, Godoy D, Castiñeiras TM, Tulenko MM, Rebelo MC, Harrison LH. 444
2012. β- Lactam resistance, serotype distribution, and genotypes of meningitis-causing 445
Streptococcus pneumoniae, Rio de Janeiro, Brazil. Pediatr. Infect. Dis. J. 31:30-36. 446
32. Park IH, Park S, Hollingshead, SK, Nahm MH. 2007. Genetic basis for the new 447
pneumococcal serotype, 6C. Infect. Immun. 75:4482-4489. 448
33. Demczuk WH, Martin I, Griffith A, Lefebvre B, McGeer A, Shane A, Zhanel GG, 449
Tyrrell GJ, Gilmour MW, Toronto Invasive Bacterial Diseases Network; Canadian 450
Public Health Laboratory Network. 2012. Serotype distribution of invasive 451
Streptococcus pneumoniae in Canada during the introduction of the 13-valent 452
pneumococcal conjugate vaccine, 2010. Can. J. Microbiol. 58:1008-1017. 453
34. Nunes S, Valente C, Sá-Leão R, de Lencastre H. 2009. Temporal trends and molecular 454
epidemiology of recently described serotype 6C of Streptococcus pneumoniae. J. Clin. 455
Microbiol. 47:472-4. 456
35. Varaldo PE, Montanari MP, Giovanetti E. 2009. Genetic elements responsible for 457
erythromycin resistance in streptococci. Antimicrob. Agents Chemother. 53:343-353. 458
36. Wolf B, Rey LC, Brisse S, Moreira LB, Milatovic D, Fleer A, Roord JJ, Verhoef J. 459
2000. Molecular epidemiology of penicillin-resistant Streptococcus pneumoniae colonizing 460
children with community-acquired pneumonia and children attending day-care centers in 461
Fortaleza, Brazil. J. Antimicrob. Chemother. 46:757-765. 462
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

23
37. Chiou AC, Andrade SS, Almeida SC, Zanella RC, Andrade AL, Brandileone MC. 463
2008. Molecular assessment of invasive Streptococcus pneumoniae serotype 1 in Brazil: 464
evidence of clonal replacement. J. Med. Microbiol. 57:839-844. 465
38. Ochs MM, Bartlett W, Briles DE, Hicks B, Jurkuvenas A, Lau P, Ren B, Millar A. 466
2008. Vaccine-induced human antibodies to PspA augment complement C3 deposition on 467
Streptococcus pneumoniae. Microb. Pathog. 44:204-214. 468
469
470
471
472
473
474
475
476
477
478
479
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

24
TABLE 1 Mutations in the atpC gene described to date among optochin-resistant Streptococcus pneumoniae isolates 480
Target codon (amino acid alteration)a Number of strains reported Country of origin (reference)
13 (Met→Ile) 1 Brazil (This study)
14 (Gly→Ser) 2 Brazil (This study); France (7)
18 (Gly→Ser) 2 Brazil (This study)
20 (Gly→Ser) 3 Brazil (This study); United States (2)
20 (Gly→Ala) 1 Brazil (This study)
23 (Met→Ile) 4 Brazil (9, this study); United States (2)
31 (Ala→Val) 1 Brazil (This study)
44 (Met→Ile) 1 Japan (12)
45 (Phe→Leu) 2 Brazil (9, this study)
45 (Phe→Val) 1 Brazil (This study)
47 (Gly→Val) 1 Argentina (10)
48 (Val→Phe) 3 Japan (12)
49 (Ala→Ser) 10 Argentina (10); Brazil (9, this study); Japan (12)
49 (Ala→Thr) 10 Argentina (10); Brazil (This study); Japan (12); United States (2)
49 (Ala→Gly) 1 Brazil (This study)
50 (Phe→Leu) 1 Japan (12)
aSubstitutions first and only detected in Brazil are outlined in bold. 481
482
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

25
LEGENDS TO FIGURES 483
484
FIG 1 Genetic relationship among optochin-resistant Streptococcus pneumoniae (Optr Sp) isolates 485
included in the study as evaluated by PFGE analysis. The 80% cut-off considered for clonal groups 486
delineation is illustrated by a red dotted line. 487 aStrain identification; those highlighted in red are reference strains of internationally disseminated 488
clones that were genetically related to some of the Optr Sp strains. 489 bSource of isolation: NP, nasopharynx; U, unknown; LRT, lower respiratory tract; CSF, 490
cerebrospinal fluid; ED, eye discharge. 491 cOptochin susceptibility phenotype: HE, heterogeneous population; HO, homogeneous population 492 dpspA gene family: NT, non typeable. 493 eMinimal inhibitory concentration (MIC) of optochin expressed in μg/mL. 494 fMutation in the atpC gene, including codon number and the respective deduced amino acid 495
substitution. 496 gSXT, trimethoprim/sulfamethoxazole; PEN, penicillin; ERY, erythromycin; CLI, clindamycin; 497
TET, tetracycline; CHL, chloramphenicol. 498
499
500
501
FIG 2 Genetic relationship among optochin-resistant Streptococcus pneumoniae (Optr Sp) 502
isolates included in this study as evaluated by MLVA analysis. A. Dendrogram showing the 503
genetic relatedness, the alleles detected for each VNTR, and the GenBank accession 504
numbers (a) of the atpC gene sequences of the 26 Optr Sp strains. The 80% cut-off 505
considered for clonal groups delineation is illustrated by a red dotted line. B. Diagram 506
constructed by using the Minimum Spanning Tree (MST) method. Node colors represent 507
the different serotypes listed in Fig 1. Higher levels of genetic relationship between nodes 508
are indicated by darker lines. 509
510
511
512
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

26
FIG 3 Predicted model of the c-subunit of ATPase based on the atpC gene sequences 513
obtained in this study. A. Protein secondary structure prediction scheme obtained by using 514
the Protein Secondary PREDiction (PSIPRED) method. Amino acids showing alterations 515
in the present study are highlighted with red boxes. B. Transmembrane protein structure 516
prediction cartoon obtained by using the MEMbrane protein Structure And Topology 517
(MEMSAT) method. S1, helix 1; S2, helix 2. The illustrations shown were designed using 518
amino acid sequence of strain Sp 1835, since all the optochin-resistant Streptococcus 519
pneumoniae isolates included in this study generated identical predictions. 520
521
522
523
FIG 4 Hydrophobicity profiles of the c-subunit of ATPase of optochin-resistant 524
Streptococcus pneumoniae (Optr Sp) isolates. A. Mean hydrophobicity profiles obtained for 525
seven representative Optr Sp isolates with alterations in six different codons belonging to 526
the helix 1 of c-subunit of ATPase. B. Mean hydrophobicity profiles obtained for five 527
representative Optr Sp isolates with alterations in two different codons belonging to the 528
helix 2 of c-subunit of ATPase. Reference strain ATCC 49619 was also included. Codons 529
with alterations are outlined with a vertical black line. 530
531
532
533
534
535
536
537
538
on Novem
ber 29, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from