optical properties of lysozyme - journal of biological ... · the optical properties of lysozyme...
TRANSCRIPT

THE JOURNAL OF BIOLO(IICAL CHEMISTRY Vol. 250. No. 20,Iasue of October 25, PP. 82754282, 1975
Printed in U.S.A.
Optical Properties of Lysozyme
pH AND SACCHARIDE BINDING DIFFERENCE SPECTRA*
(Received for publication, January 30, 1975)
TAIJI IMOTO,$ LEONARD J. ANDREWS,~ SURATH K. BANERJEE,~ ANDREW SHRAKE,/~ LESLIE S. FORSTER, AND JOHN A. RUPLEY**
From the Department of Chemistry, The University of ArizoTla, Tucson, Arizona 85721
SUMMARY
Difference spectra associated with changes in pH and with binding of saccharides have been recorded for hen egg white (HEW) lysozyme, turkey egg white (TEW) lysozyme, and for the derivatives of the hen protein in which Trp-62 or Trp-108 had been oxidized specifically to oxindolealanine to give the Oxa- or Oxa-lo&proteins. Identical pH difference spectra were obtained for HEW, TEW, and Oxa-62-lyso- zymes. Oxidation of Trp-108 is reflected in both the high and low pH (pH 7 versus 5 and pH 2 uersus 5) difference spectra. The magnitude of the low pH difference spectrum is en- hanced by binding of saccharide for HEW and Oxa-62-lyso- zymes but not for TEW lysozyme. The shapes and magni- tudes of saccharide binding difference spectra are affected by oxidation of residues 62 or 108. These results can be inter- preted in terms of the perturbations responsible for the lyso- zyme difference spectra. The pH 7 versus 5 difference spec- trum results from perturbation by Glu-35 of Trp-108 and another tryptophan, probably Trp-63. Perturbation of Trp-108 and one or more other tryptophan residues by several car- boxylate groups is responsible for the low pH difference spectra of the unliganded HEW and TEW lysozyme mole- cules. Perturbation of Trp-108 makes a principal contribution to the saccharide-binding difference spectrum. Perturbation of the Oxa- chromophore by ionization of Glu-35 or by saccharide binding produces absorbance changes in the 250 to 265 nm region.
Near-ultraviolet (240 to 310 nm) absorption spectra of proteins are complex with overlapping contributions from disulfide link-
* This research was supported by grants from the National Science Foundation and the National Institutes of Health.
$ Present address, Laboratory of Biochemistry, Faculty of Agriculture, Yamaguchi University, Yamaguchi, Japan.
0 Present address, Department of Chemistry, Brandeis Uni- versity, Waltham, Mass.
1 Present address, Department of Internal Medicine, Division of Cardiolonv. Universitv of Arizona Medical School, Tucson, Ariz. 85721.-” ’
I/ Present address, National Heart and Lung Institute, National Institutes of Health, Building 3, Room 208, Bethesda, Md. 20014.
** To whom requests for reprints should be addressed.
ages and the aromatic side chains of phenylalanine, tyrosine, and tryptophan. Difference spectra are often used to separate the several contributions so that only the absorbing moieties affected by a perturbation are measured. Although interpretation of a protein difference spectrum is less uncertain than analysis of a direct spectrum, such difference spectra nevertheless are complex and varied in shape.
The optical properties of lysozyme are particularly attractive to study because the structure of this protein is known (l), a large body of chemical data exists for it (for a review, see Ref. 2), and it is possible to prepare derivatives lacking one or more chromo- phores, or groups responsible for perturbation of chromophores, or both.
Donovan et al. (3) and Ogasahara and Hamaguchi (4) have reported pH difference spectra for lysozyme. Difference spectra associated with the formation of enzgme.saccharide complexes have been used to study the binding of various ligands (5).
The aim of the present work is to analyze the pH and sac- charide-binding difference spectra of HEW and TEW lysozymes and of oxindolealanine derivatives of the HEW enzyme (Oxa-62, Oxa-108, and ester-35,108.HEW lysozgmes) in terms of solvent and charge perturbation difference spectra of tryptophan model compounds (6). This study is complementary to recent work on the steady state fluorescence (7), fluorescence lifetimes (8), and circular dichroism spectrum (9) of lysozyme and its oxindoleala- nine derivatives. For interpretation of comparative studies like those described in this paper, the conformations of the proteins examined must be closely similar, or if there are differences these must have been defined. Previous studies have indicated that Oxa-62, Oxa-108, and ester-35,108-HEW lysozymes have struc- tures like that of the native protein.2 The sequence of TEW lysozyme differs from that of the HEW protein at seven positions (10). All replacements are of surface residues and should not result in differences in conformation. There is only one difference in the active site, i.e. the replacement of Asp-101 of HEW lyso- zyme by glycine. The cat.alytic properties of the HEW and TEW proteins are essentially identical (1 l), which supports the conclu- sion that these molecules have identical conformations.
1 The abbreviations used are: HEW, hen egg white; TEW, turkey egg white; Oxa, oxindolealanine; ester-35,108.lysozyme, the derivative in which the side chain of Glu-35 is esterified with the enol form of Oxa-108; (GlcNAc),, p(1 + 4).linked Tj-mer of %acetyl-D-gfucosamine; GlcNAc-CH3, p-methyl-A-acetyl-n-glu- cosaminide; SucNBr, iV-bromosuccinimide.
2 For discussion, see Ref. 7.
8275
by guest on March 22, 2020
http://ww
w.jbc.org/
Dow
nloaded from
5276
EXPERIMENTAL PROCEDURES
Materials-Crystalline HEW lysozyme was obtained from Worthington. Oxa-62-HEW lysozyme was prepared by SucNBr oxidation of HEW lysozyme (12). For some experiments the Oxa- derivative was purified further by chromatography on Bio-llex 70 (13) and affinity chromatography (14). Purity of the product was determined to be greater than 95 y0 by chromatographic analysis
4 . /
* .
PH
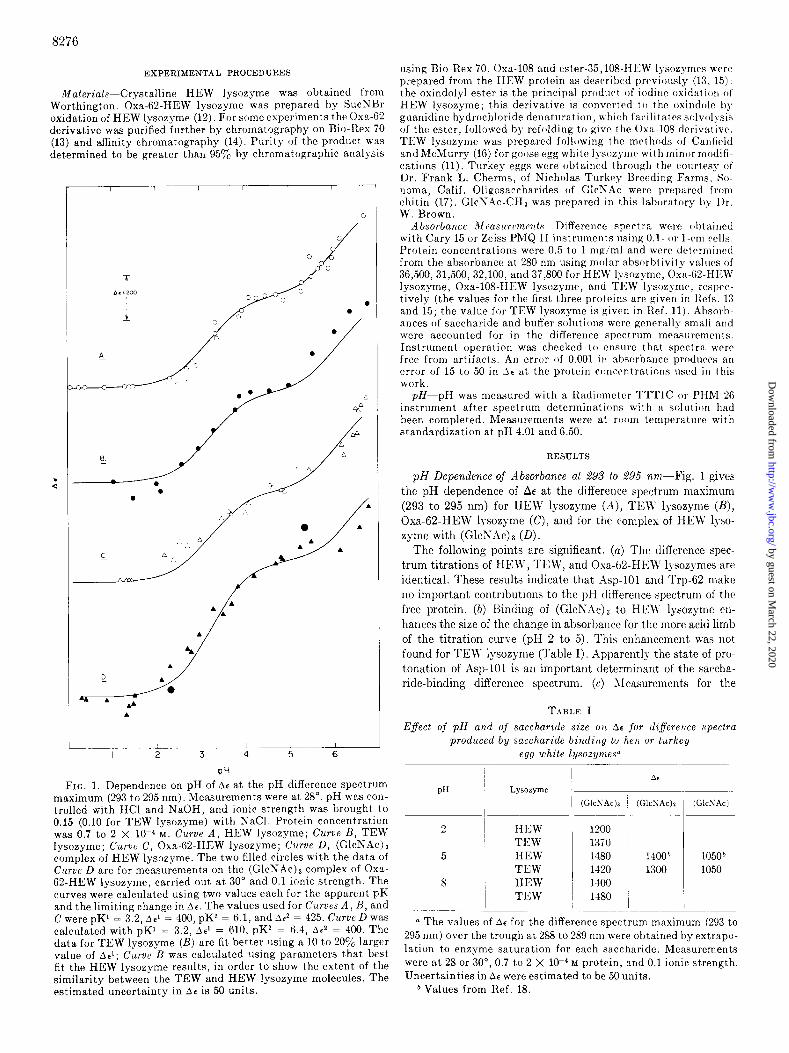
FIG. 1. Dependence on pH of ~.r at the pH difference spectrum maximum (293 to 295 nm). Measurements were at 28”. pH was con- trolled with HCl and NaOH, and ionic strength was brought to 0.15 (0.10 for TEW lysozyme) with NaCl. Protein concentration was 0.7 to 2 X 10-d M. Curve A, HEW lysozyme; Curve B, TEW lysozyme; Curve C, Oxa-62.HEW lysozyme; Curve D, (GlcNAc), complex of HEW lysozyme. The two filled circles with the data of C UT~C D are for measurements on the (GlcNAc)p complex of Oxa- 62.HEW lysozyme, carried out. at 30” and 0.1 ionic strength. The curves were calculated using two values each for the apparent pK and the limiting change in At. The values used for Curves A, B, and C were pK1 = 3.2, be1 = 400, pK* = 6.1, and Ar2 = 425. Curve D was calculated with pK1 = 3.2, Ae’ = 610, pK* = 6.4, A? = 400. The data for TEW lysozyme (H) are fit better using a 10 to 200/G larger value of Ael; C’~lrue n was calculated using parameters that best fit the HEW lysozyme results, in order to show the extent of the similarity between the TEW and HEW lysozyme molecules. The estimated uncertainty in be is 50 units.
using Bio-Rex 70. Oxa- and ester-35,108.HEW lgsozymcs were prepared from the HEW protein as described previously (13, 15) : the oxindolyl ester is the principal produrt of iodine oxidation of HEW lysozyme; this derivative is convc>rtcd to thr oxindolc b) guanidine hydrochloride denaturation, which facilitates solvol?sis of the ester, followed by refolding to givcx the Ox:t-108 dcrivatlve. TEW lysozyme was prepared following the methods of Canfield and McMurry (16) for goose egg white lgsozgmr with minor modifi- cations (11). Turkey eggs were obtained through the courtesy of Dr. Frank L. Cherms, of Nicholas Turkey Breeding Farms, So- noma, Calif. Oligosaccharides of GlcNAc were prepared from chitin (17). GlcNAc-CH, was prepared in this laboratory by Dr. W. Brown.
Absorbance nlcasur~molts-Differerlce spectra wrrc obtained with Cary 15 or Zeiss PMQ II instruments using 0.1. or l-cm cells. Protein concentrations were 0.5 to 1 mgjml and wrrc dct(,rmincd from the absorbance at 280 nm using molar absorbtiviiy valr~cs of 36,500, 31,500, 32,100, and 37,800 for HEW lysozymc, Oxa-(YZIII’W lysozyme, Oxa-108.HEW lysozyme, and TEW lysozymr, rcspcc- tively (the values for the first three proteins are given in Itcfs. 13 and 15; the value for TEW lysozyme is given in Ref. 11). Absorh- antes of saccharide and buffer solutions were generally small and were accounted for in the differ-rice spectrum mcasllrcmrnts. Instrument operation was checked to rnsrlrc that spectra wprc free from artifacts. An error of 0.001 in absorbance prodllcrs an error of 15 to 50 in me at. the protein conrrntrations used in this work.
pH-pH was measured with a Radiometer TTTlC or PHnl 26 instrument after spectrum determinations with a solution had been completed. Measurements were at room temperature with standardization at pH 4.01 and 6.50.
RESULTS
pH Dependence of Absorbance at 295 to 295 nm-Fig. 1 gives
t,he pH dependence of AE at the difference spectrum maximum
(293 to 295 nm) for HEW lysozgme (A), ‘I%\?’ lgsozyme (B), Oxa-62.HEW lysozyme (C), and for the complex of HEW lyso-
zyme with (GlcS.4~)~ (D). The following points are significant. (a) The difference spec-
trum titrations of HEW, TER, and Oxa-62.HEW lgsozymes are
identical. These results indicate that Asp-101 and Trp-62 make
no important contributions to the pH difference spectrum of the free protein. (b) Binding of (G~cNAc)~ to HE\V lgsozyme erl-
hances the size of the change in absorbance for the more acid limb
of the titration curve (pH 2 to 5). This enhancement was not
found for TEW lysozyme (Table I). Apparently the state of pro- tonation of Asp-101 is an important determinant of the saccha-
ride-binding difference spectrum. (c) Jleasurements for the
TABLE I
Effect of pH and of saccharide size OR At for differerrce spectra produced by saccharide birldirlg to hen or turkey
egg white lysozymes”
PH Lysozyme
HEW 1200 TEW 1370 HEW 1480 1400” TEW 1420 ~ 1300 HEW 1400 , TEW 1480
/ At
(ClcSAc)r ~ (GlcSAc)z (&SAC)
10506 1050
” The values of AE for the difference spectrum maximum (293 to 295 nm) over the trough at 288 to 289 nm were obtained by extrapo- lation to enzyme saturation for each saccharide. Measurements were at 28 or 30”, 0.7 to 2 X lo-’ M protein, and 0.1 ionic strength. Uncertainties in Ae were estimated to be 50 units.
* Values from Ref. 18.
by guest on March 22, 2020
http://ww
w.jbc.org/
Dow
nloaded from

8277
lOOOr 1
500
I
ot
FIG. 2. Low and high pH difference spectra for native (A), Oxa- (R), Oxa- (C), and ester-35,108-HEW lysozymes (D). Solid CUTVFS are for pH 7 versus 4.5; dashed curves are for pH 2 versus 4.5. Measurements were made at 28”, ionic strength 0.05, and 7 X 1O-s M protein. The reference solution pH of 4.5 is close to the average of the apparent pK defined by the results of Fig. 1.
-500i ,’ I L L I i
240 260 280 300 320 260 280 300 320
WAVELENGTH (n m)
I I I I I I I
240 260 280 300 320
WAVELENGTH (nm)
FIG. 3. Low and high pH difference spectra for the complex of (G~cNAc)~ with HEW Iysozyme. The solid curve is for pH 6.8 versus 4.8; the dashed curve is for pH 1.5 versus 4.8. Measurements were made at 28”, 5 X 1OP M (GlcNAc)a, and 7 X 10m6 M protein. pH was controlled using HCl and NaOH, with ionic strength brought to 0.15 with NaCl. The sample and reference pH levels of these experiments are slightly different from those of Fig. 2, which accounts for the different balance between the intensities of the low and high pH difference spectra.
(G~cNAc)~ complex of Oxa-62.HEW lysozyme at pH 2.3 and 5.3 gave the AE values shown in Fig. lD, which suggest that the pH dependence of the spectrum of the saccharide .enzyme complex has no principal contribution from Trp-62. (d) Apparent pK values of 3.2 and 6.1 fit the spectrum titration results for HEW, TEW, and Oxa-62-HEW lysozymes (Curves A to C). These values agree well with the values of 3.2 and 6.2 reported b> Donovan et al. (3) for HEW lysozyme. Curve D of Fig. 1 for the complex with (G~cNAc)~ is drawn with apparent pK values of 3.2 and 6.4.
pH IXflerence Spectra-Fig. 2 gives two pH difference spectra for each of four proteins: HEW lysozyme (A); Oxa-62-HEW
L I I I 1 , I 260 280 300 320
WAVELENGTH (nm)
FIG. 4. Low and high pH difference spectra for TEW lysozyme. The solid curve is for pH 6.8 versus 5; the dashed curve is for pH 2 versus 5. Measurements were made under the conditions of Fig. 1.
lysozyme (B); Oxa-108-HEW lysozyme (C); and ester-35,108. HEW lysozyme (D). The pH 7 versus 4.5 and pH 2 versus 4.5
difference spectra bracket the apparent pK values defined by the titration results of Fig. 1. Figs. 3 and 4 give corresponding pairs of difference spectra for the complex of (G~cNAc)~ with HEW lysozyme and for TEW lysozyme, respectively.
The following are the principal results of these measurements. (a) The pH 7 versus 4.5 difference spectra of HEW and Oxa-62- HEW lysozymes are nearly identical and show maxima at 302, 294, and 286 nm. (b) The ester-35,108 derivative shows no significant high pH difference spectrum; thus this feature is asso- ciated with deprotonation of Glu-35. (c) Conversion of the ester to the Oxa- derivative results in a significant pH 7 versus 4.5 difference spectrum and a unique new band at 250 to 265 nm. This latter feature is characteristic of perturbation of an oxindole- alanine chromophore and presumably results from interaction of
by guest on March 22, 2020
http://ww
w.jbc.org/
Dow
nloaded from

8278
Glu-35 with Oxa-108. The 290 nm band reflects perturbation by Glu-35 of some tryptophan residue other than Trp-108. (d) The pH 2 versus 4.5 difference spectra are in shape unlike the high pH difference spectra: the 294- and 286nm maxima remain, but the 302 nm peak is reduced to a shoulder. The acid pH difference spectra for HEW and Oxa-62.HEW lysozymes are similar. Al- though a difference in intensity is seen in the data of Fig. 2, com- paring the native and Oxa-62.HEW lysozyme spectra, this is an artifact that reflects the sample of Oxa-62.HEW protein, which was prepared according to procedures (12) that produce a small amount of denatured material. The close similarity of the spectra was shown by a comparison between native protein and specially purified Oxa-62.HEW lysozyme (see “Experimental Procedures”) which gave identical difference spectra for pH 2.1 versus 5.3. The Oxa-62-HEW lysozyme spectra of Fig. 2, which should be viewed as demonstrating spectrum shape and only approximately spec- trum intensity, were included because they were measured at the same time and with the same instrument and settings as the other spectra of Fig. 2. (e) Oxidation of Trp-108 reduces generally the
I 1 I I I I I
A PH 2
PH 5
‘\ \..-.. .-L .,~”
I I 1 1 I I I
240 260 280 300 320 WAVELENGTH (nm)
FIG. 5. Difference spectra for binding of (GlcNAc)z to HEW lysozyme and derivatives at pH 2, 5, and 8. Solid CUTUBS, native HEW lysozyme; dotted curves, Oxa-62.HEW lysozyme; dashed curves, Oxa-10%HEW lysozyme. Measurements were at 28”, 6 X
intensity of the acid pH difference spectrum and in particular the intensity of the 302 nm shoulder. We emphasize that these differ- ences are significant and that the sample of protein studied was homogeneous by ion exchange chromatography. (f) The shapes of the pH difference spectra of the complex of (GlcNAc)a with HEW lysozyme resemble those of the free protein (compare Figs. 2 and 3), although the intensity in the acid region is greater (see also Table I). (g) The shapes and intensities of the pH difference spectra for TEW lysozyme are identical with those of HEW lyso- zyme, except the 302 nm shoulder in the acid spectrum is absent.
Saccharide-binding Difference Spectra-Fig. 5 gives the differ- ence spectra resulting from complex formation of (G~cNAc)~ with Oxa-62.HEW, Oxa-108.HEW, and native HEW lysozymes at pH 2, 5, and 8. Different association constants characterize the binding of (G~cNAc)~ to these three proteins (approximately 105, 103, and lo2 for HEW, Oxa-62-HEW, and Oxa-108.HEW lyso- zymes, respectively). Although the (GlcNAc) 3 concentration used in each of these measurements was chosen to be nearly sufficient to saturate, the spectra of Fig. 5 should be viewed principally as showing spectrum shape and not spectrum in- tensity.
The following are the principal results of these measurements. (a) Oxidation of Trp-108 has a generally strong effect on the in- tensity of the saccharide-binding difference spectrum. In particu- lar, the 302 nm band found at pH 8 for the native protein is com- pletely lost following oxidation of Trp-108. (b) The increased absorbance at 260 nm for the complex of the Oxa- derivative at pH 8 probably reflects perturbation by saccharide of the Oxa- 108 chromophore, which is also perturbed by ionization of Glu-35 (Fig. 2). (c) Oxidation of Trp-62 produces a small change in shape but little change in intensity.
The saccharide-binding difference spectrum for TEW lysozyme at pH 8 is shown in Fig. 6. It resembles closely in shape and in- tensity the corresponding difference spectrum of HEW lysozyme, except the 302 nm band is not a separate peak, but a shoulder. Less intense 302.nm intensity was also found for the low pH difference spectrum of TEW lysozyme.
E$ect of Saccharide Size on Difference Spectrum-Fig. 7 gives saccharide-binding difference spectra at pH 8 and ligand concen- trations sufEcient for near-saturation of the HEW enzyme with mono-, di-, and trisaccharides of GlcNAc. The 302 nm band develops from a shoulder into a peak with increase in saccharide
I I I I I I I
)-
t I I I I I 260 280 300
WAVELENGTH (nm)
1 32 10
FIG. 6. Difference spectrum for binding of (GlcNAc)t to TEW lysozyme, measured at 30”, pH 8, 2 X 10m3 M (GlcNAc)z, 5.5 X
1W6 M enzyme, 5 X 10mS to 10m2~ (GlcNAc) 3, and 0.1 ionic strength. 10mK M enzyme, and 0.1 ionic strength.
by guest on March 22, 2020
http://ww
w.jbc.org/
Dow
nloaded from

I I I I
10 300 32( 3 WAVELENGTH (nm)
FIG. 7. Dependence of the saccharide-binding difference spec- trum for HEW lysozyme on l&and size. Measurements were with 0.1 M sodium phosphate bufferof pH 8, at 28”, 6 X 10m5 M enzyme, and concentrations of (GIcNAc)~, (G~cNAc)~, and GlcNAc-CH3 of 10e3, 10-a, and 10-i M, respectively.
size. The intensity of the 293 nm peak increases with saccharide size for both the HEW and TEW lysozyme molecules (Table I). The difference spectrum developed through binding of monosac- charide to native HEW lysozyme is remarkably similar to that found for binding of (G~cNAc)~ to the Oxa- derivative (com- pare Figs. 5 and 7).
DISCUSSION
pH Difference Spectrum Shapes
Andrews and Forster (6) have analyzed charge and solvent perturbation difference spectra of tryptophan model compounds. The several shapes arise from the varying IL, and ‘La energies of the indole moiety and the extent of the differential shifting of these states induced by the perturbant. These results can be used to interpret the pH and saccharide binding difference spectra of lysozyme.
The 302 nm peak seen in high pH difference spectra would arise from absorption around 300 nm in the direct spectrum. The ab- sence of the 302~nm feature in all difference spectra of Oxa- and ester-35,108HEW lysozymes shows that Trp-108 is the source of this band. Trp-108 is hydrogen-bonded to Leu-56 and the side chain is held rigidly in position. Hydrogen bonding pro- duces a substantial lL, red shift (19). In addition, Trp-108 is buried nearly completely in a hydrophobic environment (20). Limited solvent access coupled with reduction in torsional mo- bility might be expected to lead to sharpening of the ‘L, + ‘A O-O band with the observed intensification. We note that the IL, + ‘A O-O band in horseradish peroxidase has been located at 300 nm by Strickland et al. (21).
The acid pH difference spectra show the features expected for perturbation of an indole by neighboring carboxylate groups and thus do not require further discussion.
Saccharide-binding Difference Spectrum Shapes
A detailed analysis of difference spectra presumes knowledge
8279
good evidence that the structure of uncomplexed HEW lysozyme is not affected significantly by oxidation of Trp-62 or Trp-108, it is not clear that the structures for the complexes of (GlcNAc) 8 with the oxindole derivatives are the same as for the complex with the native protein. Varied modes of saccharide binding have been observed for native HEW lysozyme (2). It is likely that the mode of binding of (GlcNAc)a to Oxa-62-HEW lysozyme differs from that for the native HEW proteina In contrast, studies with TEW lysozyme indicate that this molecule binds (GlcNAc)a like HEW lysozyme (11). Keeping these cautionary comments in mind, one can arrive at significant correlations based upon the shapes of the saccharide-binding difference spectra.
Crystallographic evidence (1, 2) indicates that (GlcNAc)a in- teracts with Trp-62, Trp-63, and Trp-108, and that the extent of these interactions decreases with decrease in saccharide size. In accord with the crystallographic results, the difference spectrum intensities for HEW and TEW lysozymes also decrease with decrease in saccharide size (Table I). The close similarity between the difference spectra produced by binding of (G~cNAc)~ to Oxa- 62-HEW lysozyme and of GlcNAc-CH3 to native HEW lysozyme (compare Figs. 5 and 7) is understood if the latter process per- turbs only Trp-108 and Trp-63. Trp-62 is believed to hydrogen bond to the hydroxymethyl group of a monosaccharide bound at the active site region C (1). The data of Table I suggest that this interaction does not perturb the absorbance of Trp-62.
Perturbation of Trp-108 makes a principal contribution to the saccharide-binding difference spectrum. This conclusion is based on the following two observations. First, a substantial reduction in difference spectrum intensity and change in difference spectrum shape follow oxidation of Trp-108. Second, the prominent 302 nm band that is found in the saccharide-binding difference spectrum for pH 8 corresponds to the 302-nm feature in the pH difference spectrum that was shown to be associated with perturbation of Trp-108 by Glu-35. The discussion above made in connection with the 302~nm feature of the pH difference spectrum is suffi- cient to indicate that this band is unusual and is unlikely to be associated with any active site tryptophan except residue 108.
The development of the 302 nm band with increasing saccha- ride size at pH 8 is of interest in view of the location of Trp-108 and Glu-35 near region C of the lysozyme active site (1). Because crystallographic studies (1) at pH near 5 have shown that region C is filled similarly by mono-, di-, and trisaccharides, this result suggests that there may be an effect of pH upon the mode of sac- charide binding that varies with saccharide size. This conclusion is in accord with equilibrium binding measurements that show different shifts in the pK of Glu-35 upon binding of GlcNAc-CH, or the di- or trisaccharide of GlcNAc (18,22).
In view of the fact that Trp-108 is a principal source of lyso- zyme difference spectra, saccharide-binding difference spectra for Oxa-62-HEW lysozyme should show a large contribution from perturbation of Trp-108. The spectra for pH 2 and pH 5 in Fig. 5 resemble the spectrum of MelSO-perturbed yohimbine hydro- chloride and are expected for change in tryptophan environment from aqueous to hydrophobic.* Conversion of yohimbine hydro-
3 Dr. A. Shrake, unpublished results obtained in this laboratory. 4 In this and the following paragraphs, difference spectrum
results for lysozyme are compared with similar data obtained using a model compound, yohimbine, which was studied by An- drews and Forster (6). Yohimbine is an indole alkaloid in which a protonatable nitrogen is fixed on the heterocyclic side of the indole ring in approximately the same position as Glu-35 is located about Trp-108 of lysozyme. The results produced by placing a charge about the indole ring, or by changing solvent, or both, are de-
of the structure of the molecules compai’ed. Although there is scribed in Ref. 6.
by guest on March 22, 2020
http://ww
w.jbc.org/
Dow
nloaded from

8280
chloride to the free base lowers the IL, energy. In accord with a reduction of IL, energy produced by ionization of Glu-35, the sac- charide-binding difference spectrum of Oxa-62.HEW lysozyme at pH 8 resembles the difference spectrum for MenSO-perturbed yohimbine.
The saccharide-binding difference spectra of Oxa-108-HEW lysozyme must reflect perturbation of Trp-62 and Trp-63. The difference spectrum at pH 5 looks like the methanol-perturbed spectrum of yohimbine hydrochloride. The 302-nm minimum results from a narrowing of the IL, band. As the IL, energy is lowered the effect of this narrowing is obliterated, and the pH 8 difference spectrum resembles methanol-perturbed yohimbine.
Carboxylate-Tryptophan Interactions
pH difference spectra can arise directly from a charge perturba- tion, or from a change in chromophore environment (an indirect effect of pH), or both. If only charge perturbation is important, the model of Andrews and Forster (6) may be used as a basis for discussion. This assumption is likely true for the free HEW and TEW lysozyme molecules, but the uncertainties indicated in the previous section should be kept in mind when considering the pH dependence of saccharide-binding difference spectra.
The interaction energy according. to a charge perturbation model can be expressed in the dipole approximation as:
Ap.r
q/T13
where Ap is the vector difference between the ground and excited state dipole moments and r is the vector joining the center of the dipole to the center of charge q. The sign of the difference spec- trum depends upon the orientation of the charge with respect to Ap and the magnitude varies as 1/ 1 r Iz. Although two transitions contribute to the near-ultraviolet difference spectrum of trypto- phan, the IL, c ‘A is much the more intense transition, and IL, energies are more sensitive to environment than iLb energies (6, 19). Ap corresponds to this transition.
Table II lists distances and orientations for all carboxylate- tryptophan pairs that might contribute to the carboxylate-de- pendent perturbations of lysozyme. Those viewed as most likely to be important are boldface. We note that the dipole approxima- tion is inadequate at the short distances within a protein and also that the effect of solvent and adjacent polarizable groups on the “through space” charge effect is uncertain. Thus quantitative calculations should not be based on the values of Table II. Most of the pH difference spectrum results can be understood, how- ever, in terms of qualitative arguments that follow from t,he simple charge perturbation model.
pH 7 versus &?--The sign of the difference spectrum in the wavelength region 280 to 300 nm requires a carboxylate group located on the five-membered ring side of the indole. Glu-35 is the only residue that has pK between 4.5 and 7 and is adjacent to any of the tryptophans and oriented appropriately. The effect of ionization of Glu-35 on Trp-108 is responsible for the bulk of the free lysozyme pH difference spectrum. The smaller 290-nm con- tribution found for the Oxa-108-HEW derivative, which pre- sumably also is present for the native protein, must reflect per- turbation by Glu-35 of another tryptophan. The only likely candidate for this interaction is the Trp-63’Glu-35 pair. The Trp-28. Glu-35 interaction would lead to an absorption difference of the wrong sign. The implication of these results is that two chromophores contribute to the pH 7 versus 5 difference spectrum
TABLE II
Distances arid direction parameters for carboxylate- tryptophan interactious=
Tryptophan Asp-18 Glu-35 Asp-48 Asp-52 ASP-66 Asp-101’ Asp-119
28: r cos a
62:* r co5 a
63: r cos a
108: r cos Ly
111: r cos Ly
123: r cos a
pK (free) pK (com-
plex)
8.9 0.95
12.4 -0.66 -
13.0 0.88 6.6 0.91
10.0 0.16
13.4 0.06 6 1 6.4
8.6 0.56
12.9 0.72
10.9 0.99
10.3 1 .oo 9.7 0.86
12.0 0.5
12.5 0.52
9 1 -0.18
7.4 - 0 .58 11.8
-0.61 14.5
0.68 13.7
0.73 8.2 0.76
3.5 1.9 4.3 3.8 1.5 3.4
- a The geometry of a carboxylate-tryptophan interaction is de-
scribed using the transition moment vector Ap and the vector r joining the center of the indole ring and the midpoint, of the car- boxylate oxygens. The values listed are the magnitude of T and Cos 01 = r.Aw/(l Ap 1 1 r I). Distances are in angstrom. The sign of the difference spectrum produced by a negative charge corre- sponds to the sign of Cos (Y. All interactions for which 1 r 1 is less than 15 A are given. Values for interactions for which Cos a/l r I2 is large are in boldface. The calculations were made using coor- dinates based on the crystallographic analyses of Blake et al. (1) and given in Imoto et al. (2). The bottom two rows give apparent pK values for those groups for which pK estimates have been made for the free HEW lysozyme molecule and its complex with (Glc- XAc), (2, 18, 22). The pK values have an estimated error of ~0.2 unit.
* The positions of the side chains of Trp-62 and Asp-101 are de- fined poorly in crystals of unliganded Hl<W lysozyme but are de- fined in the complex with (GlcNAc)r (1). The coordinates given in Imoto et al. (2) for Asp-101 are for the (Glcl;Ac)r complex. The coordinates used for Trp-62 are those estimated for the unliganded protein.
and that both chromophores are perturbed by ionization of Glu-35.5
The intense 250- to 265.nm structure in the Oxa-10%HER lysozyme pH difference spectrum undoubtedly arises from the perturbation of Oxa- by Glu-35. This feature can be repro- duced by summing the observed difference spectrum for native lysozyme and a calculated difference soectrum for a 1-nm red shift of the spectrum of oxindole-3-propionic acid. It is of interest that Oxa- is not perturbed by ionizable groups or saccharide (there are no low wavelength features in the difference spectra for Oxa-62.HEW lysozyme).
pH d versus 4.5-The results listed in Table II can be used to develop a list of the carboxylate-indole interactions that possibly contribute to the acid pH difference spectrum. First, we note that Trp-62 does not contribute to the acid pH difference spectrum. Oxidation of Trp-62 affects neither the shape of the pH difference spectrum (as shown in Fig. 2) nor its magnitude (as shown in Fig. 1 and discussed under “Results”). The side chain of this
L His-15 is too distant from all tryptophan residues of lysozyme for protonation of it to be reflected in absorbance properties. Also, the pK of His-15 has been shown to be near 5 (for a review discussion, see Ref. 2), and no apparent pK of this value has been observed in studies of the optical properties of lysozyme.
by guest on March 22, 2020
http://ww
w.jbc.org/
Dow
nloaded from

8281
residue was not well located by the crystallographic analysis of the unliganded lysozyme molecule (1). It is possible that vibra- tions of Trp-62 lead to a near-zero average value for the interac- tion energy (i.e. for the cosine term given in Table II), which would explain the absence of a perturbation of Trp-62 by Asp-52. If interactions of Glu-35, which has a high pK, are excluded, then in order of size of expected contribution we find Trp-28’Asp-18, Trp-123 .Asp-119, Trp-63 .Asp-101, Trp-63 .Asp-52, and Trp- 108 .Asp-52. The observation that oxidation of Trp-108 reduces the intensity of the acid pH difference spectrum indicates that perturbation of this chromophore makes a contribution. The only carbosylate residue that could be responsible for this effect is Asp-52. Because oxidation of Trp-108 does not abolish the low pH difference spectrum, at least one other chromophore must be perturbed by a carboxylate. Because the pK of Asp-52 in the free protein is 3.5,6 significantly above the pK of 3.2 observed in the titrations of Fig. 1, there must be a principal contribution in- volving Asp-18, Asp-48, and/or Asp-l 19, residues for which pK values have not been estimated and which might have pK 3.2 or below. Therefore, at least two chromophores must be perturbed by at least two acid groups that ionize in the pH range 2 to 5, and the apparent pK of 3.2 observed in the spectrum titration re- flects more than one ionization.
In view of the close proximity of Asp-101 to Trp-63, this inter- action merits discussion. The value of Cos cy (Table II) for the Trp-63.Asp-101 interaction is negative. This would lead to a difference spectrum contribution of sign opposite to that ob- served in the acid range. Although it may be that such a perturba- tion is present but masked by other contributions of opposite sign, the similarity of the low pH difference spectra for HEW and TEW lgsozgmes shows that any effect of the ionization of ASP- 101 must be small. In esplanation of this observation, the side chain of Asp-101 is not located by crystallographic analysis of the unliganded lysozyme molecule (1), and in some orientations of the side chain the value of Cos a! is near-zero. The enhancement of the low pH difference spectrum that follows complexation of (G~cNAc)~ with HEW but not TEW lgsozyme must reflect the presence of Asp-101. This contribution could be indirect, i.e. an effect produced by change in environment such as in the mode of binding of saccharide to the protein.
Comparison of Resul@ of Absorbance, Fluorescence, and CD dleasurements
The absorbance results obtained in this work are intended to complement previous studies of the fluorescence (7, 8) and CD spectra (9, 23) of HEW lysozyme and its derivatives. It is there- fore of interest to compare the contributions of the Trp-62 and Trp-108 chromophores to the several optical properties.
Trp-&?-This residue makes a principal contribution to the fluorescence properties of the free protein, some contribution to the CD changes associated with saccharide binding, and no significant contribution to the CD spectrum or absorbance pH difference spectrum of the free protein. It is not clear whether the absorbance of Trp-62 is affected by saccharide binding.
Trp-108-Trp-108 and Trp-62 together account for more than three-quarters of the steady state fluorescence of lysozyme. Energy transfer involving Trp-108 and Trp-62 has been demon-
s A pK value higher than 3.5 has been suggested for Asp-52, but recent results from several laboratories indicate that the pK is near 3.5 (for a discussion, see Ref. 11). Furthermore, the conclu- sion important for the present work is that the pK of Asp-52 is above 3.2, which would follow from all literature reports on the pK of this residue.
strated through steady state fluorescence and fluorescence life- time measurements. The contribution of Trp-108 to the CD spectrum of the free protein is small. Trp-108 makes a principal contribution to the high pH absorbance difference spectrum and makes a significant but not dominant contribution to the acid pH difference spectrum. Its absorbance and fluorescence properties are perturbed by saccharide binding.
Other Tryptophan Residues-Residues other than Trp-62 and Trp-108 can make only small contributions to the fluorescence of the protein, but they must dominate the near-ultraviolet CD spectrum. It is likely that the absorbance of Trp-63 is affected by saccharide binding and that Trp-63 and one other tryptophan are perturbed by one or more neighboring ionizable groups.
Carboxylate-Tryptophan Interactions-The effect of Glu-35 on Trp-108 is reflected strongly in the fluorescence of lysozyme. saccharide complexes and in the absorbance of both free protein and complexes. Asp-101 enters directly or indirectly into the determination of the saccharide-binding absorbance difference spectrum, and it is probably principally responsible for the pH dependence of the fluorescence of the HEW lysozyme . (G~cNAc)~ complex between pH 2 and 5 (reported in Ref. 24), which is ab- sent for the TEW lysozyme.(GlcNA~)~ comples (11). Asp-52 and at least one other carboxylate group affect chromophores that contribute to the acid pH absorbance difference spectrum. A contribution from Asp-52 is also consistent with the pH de- pendence of the Huorescence properties of unliganded lysozyme, which is displayed more prominently by the oxindolealanine derivatives than by the native protein (7).
Concluding Remarks
The interpretation of protein difference spectra is seldom un- ambiguous. HEW lgsozyme provides a stringent test for anJ scheme of interpretation based upon model compound studies, in view of the six aromatic chromophores and correspondingl) large number of neighboring ionizable groups present in this molecule. The charge perturbation effects upon tryptophan ab- sorption in lysozyme can be accounted for qualitatively by refer- ence to results obtained using suitable model compounds. A principal conclusion derived from the present work is that even a relatively simple-appearing pH difference spectrum reflects a complex set of interactions involving several chromophores and several ionizable groups. The possible involvement of multiple perturbing interactions which may produce opposing affects bvill make it particularly difficult to locate the source ot perturbations for proteins for which the structure is unknown.
REFERENCES 1. BLAKE, C. C. F., JOHNSON, L. N., MXR, G. A., NOIETH, A. C.
T., PHILLIPS, D. C , AND SHARM,~, V. It. (1967) Proc. Izo7~. Sot. Loud. Ser. B Biol. Sci. 167, 378-388
2. IMOTO, T., JOHNSON, L. N., NORTH, A. C. T., PHILLIPS, D. C.. :\ND RUPLI,:Y. J. A. (1972) in The Eurumcs (Bover. P. U.; ed) 3rd Ed, Gal. VII; pp.’ 665-868, Acjdemic Prkss; New York
3. DONOVAN, J. W., L.ISKOWSKI, M., JR., ASD SCHI:IUG.~, H. A. (1961) J. Am. Chem. Sot. 83, 2686-2694
4. OGASAHARA, K., AND H.\M.\GUCHI, K. (1967) J. 11iochem. (Tokvo) 61, 199-210
R;PLIS;, >. A., BUTLER, L., GERRING, M., H.~RTDIZGICN, F. J., AND PECOR.IRO, Ii. (1967) Proc. Nail. Acad. Sci. U. S. A. 67,1088-1095
ANDREWS, L. J., .~ND FORSTER, L. S. (1972) Biochemistry 11, 1875-1879
IMOTO, T., FORSTER, L. S., RUPLXY, J. A., AND TANAKA, F. (1972) Proc. Xatl. Acad. Sci. U. S. A. 69, 1151-1155
FORMOSO, C., .~ND FORSTXR, L. S. (1975) J. Biol. Chem., 260, 3738-3745
by guest on March 22, 2020
http://ww
w.jbc.org/
Dow
nloaded from

9.
10.
11.
12.
13.
14.
15. 16.
17.
TANAKA, F., FORSTER, L. S., PAL, P. K., AND RUPLEY, J. A. 18. BANERJEE, S. K., VANDENHOFF, G. E., AND RUPLXY, J. A. (1975) J. Biol. Chem., 260,6977-6982 (1974) J. Riol. Chem. 249, 1439-1444
LARUE, J. N., AND SPECK, J. C., JR. (1970) J. Biol. Chem. 246, 19. STRICKLAND, E. H., BILLUPS, C., AND K.~Y, E. (1972) Bio- 1985-1991 chemistry 11, 3657-3662
B.~NERJEE. S. K., AND RUPLEY, J. A. (1975) J. Biol. Chem. 260, 20. SHRAKE, A., AND RUPLEY, J. A. (1973) J. Mol. Biol. 79, 351- 371 8267-82f4
HAYASHI, K., IMOTO, T., FuNa~su, G., .IND FUNATSU, M. 21. (1965) J. Biochem. (Tokyo) 68, 227-235
I&To,‘T., HARTDEG~N, @. i., AND RUPLEY, J. A. (1973) J. Mol. Biol. 80, 637-648 22.
CORNELIUS, D. A., BROWN, W., SHR~ZICE, A., .~ND KUPI‘EY, J. A. (1974) Methods Enzymol. 34, 639-645 23.
hOTO, T., .IND RUPLEY, J. A. (1973) J. Mol. Biol. 80,657-667 CANFIELD, Ii. E., AND MCMUHRY, S. (1967) Biochem. Biophys.
Res. Commzcn. 26, 38-42 24. RUPLEY, J. A. (1964) Biochim. Biophys. Acla 83, 245-255
STRICKLAND, E. H., HOR~ITZ, J., KAY, E., SHANNON, L. M., WILCHEK, M.. AND BILLUPS. C. (1971) Biochemistru 10. 2631-2638’ ’
I ” I
BANERJEE, S. K., AND RUPLEY, J. A. (1973) J. Rio!. Chew 248, 2117-2124
KURAMITSU, S., IKED.~, K., HAM.~GU~HI,K., FUJIO, H., AMANO, T., MIWA, S., AND NISHINA, T. (1974) J. B’ochem. (Tokyo) 76, 671-683
LEHRER, S. S., .IND FASMAN, G. D. (1967) J. Riol. Chem. 242. 4644-4651
by guest on March 22, 2020
http://ww
w.jbc.org/
Dow
nloaded from

T Imoto, L J Andrews, S K Banerjee, A Shrake, L S Forster and J A RupleyOptical properties of lysozyme. pH and saccharide binding difference spectra.
1975, 250:8275-8282.J. Biol. Chem.
http://www.jbc.org/content/250/20/8275Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/250/20/8275.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on March 22, 2020
http://ww
w.jbc.org/
Dow
nloaded from