ontogenetic morphometrics of some upper cretaceous ... · pdf filetechniques for biometric...
TRANSCRIPT

fore, organisms that depended on algae as a principal nutrientsource, such as bivalves, would have been adversely affected.
It is therefore being proposed that E. antarctica shows that itsgrowth was restricted, due not to the environmental tem-perature regime, but to the fact that its growing season wouldhave been very short compared to that at lower latitudes. In thiscase, bivalve growth patterns could be very useful as pal-colatitude indicators.
This research was supported by National Science Foundationgrant DPP 84-16783.
References
Eisma, D., W.G. Mook, and H.A. Das. 1976. Shell characteristics,isotopic composition and trace-element contents of some euryhaline
molluscs as indicators of salinity. Palaeogeography, Palaeoclimatology,Palaeoecology, 19, 39-62.
Kennett, J.P. 1977. Cenozoic evolution of antarctic glaciation, the cir-cum-antarctic ocean, and their impact on global paleoceanography.Journal of Geophysical Research, 82(27), 3843-3859.
Panella, C., and C. MacClintock. 1968. Biological and environmentalrhythms reflected in molluscan shell growth. Journal of Paleontology,42(5, part II), 64-80.
Tilzer, MM., and Z. Dubinsky. 1987. Effects of temperature and daylength on the mass balance of Antarctic phytoplankton. Polar Biology,7, 35-42.
Urey, H.C., H.A. Lowenstam, S. Epstein, and C.R. McKinney. 1951.Measurement of paleotemperatures and temperatures of Upper Cre-taceous of England, Denmark, and the southeastern United States.Geological Society of America Bulletin, 62, 399.
Ontogenetic morphometrics of someUpper Cretaceous foraminifera from
the southern high latitudes
BRIAN T. HUBER
Byrd Polar Research Centerand
Department of Geology and MineralogyOhio State University
Colunibus, Ohio 4321()
Several factors have impeded efforts toward improving theUpper Cretaceous planktonic foraminiferal biostratigraphy ofthe southern high latitudes: (1) rarity of thermophilic taxa usedin the standard zonal schemes, (2) apparently slow evolution-ary rates, and (3) high morphologic variability of several indige-nous planktonic species. The latter factor has caused consider-able taxonomic confusion in studies of Upper Cretaceousglobigerine foraminifera from Deep Sea Drilling Project hole511 (Falkland Plateau) and Seymour Island (Antarctic Penin-sula). Specimens from these localities, which were previouslyclassified as Rucoglobiçerina rugosa (Plummer), R. pilula Belford,R. pustulata Bro;dinnimann, R. rotundata Bro;dinnimann, andHedbergella montnouthensis (Olsson) by Sliter (1976), Krashenin-nikov and Basov (1983) and Huber, Harwood, and Webb (1983),show considerable intraspecific morphologic overlap, and theydiffer significantly from the original type descriptions (Huber inpress). The proper taxonomic designation of these mor-photypes is important because they dominate the antarctic andFalkland Plateau planktonic foraminiferal assemblages. Theiroriginal taxonomic assignments were based only on externalcharacteristics of the foraminiferal shells, without regard to theontogenetic features preserved within. To achieve taxonomicstability within this group, it became apparent that more de-tailed morphologic information was needed.
New methods were used to observe the developmental on-togeny of the Falkland Plateau and Seymour Island specimensand compare them with topotypes of the species listed. Oneprocedure involves a series of test dissections and observationsunder the scanning electron microscope using techniques sim-
ilar to those described by Huang (1981). Removal of the ventralchamber walls was achieved through the use of a very finelyground needle mounted on a Sensaur micromanipulator, whichenables precisely controlled movement in all three dimensions.Examination of the internal morphology using this methodreveals valuable information on the ontogenetic changes in (1)wall surface ornament and porosity, (2) apertural position, (3)chamber size, and (4) number of chambers per whorl (figure 1,items 1, 3, 4, 5, and 6; figure 2, items 1, 3, 4, 5, 6, and 7).
High resolution X-radiograph images of planktonic for-aminifera were produced by adapting techniques described byArnold (1982) (figure 1, item 2; figure 2, item 2). Equatorial viewsof the X-ray 1 x images were analyzed using morphometricssoftware in conjunction with a Leitz orthoplan microscope, adigitizing tablet, and a microcomputer. The chosen parametersinclude presence/absence and numerical attributes as well aslinear and areal measurements which characterize the on-togenetic morphological changes. Internal morphologic charac-teristics considered as taxonomically useful include: (1) numberof chambers in the penultimate whorl, (2) ontogenetic changesin the cross-sectional chamber area and shape, (3) position ofthe generating curve, and (4) ratio of the penultimate and ante-penultimate chamber areas. The advantage of using this meth-od is that data can be generated rapidly from large foraminiferalpopulations, and it can easily be analyzed statistically.
Results of this study indicate that the upper Campanian!Maastrichtian specimens from the Falkland Plateau andSeymour Island, which were previously included in Hedbergellaand Rugoglobigerina, are growth morphovariants of a single,new, high-latitude rugoglobigerinid species whose distributionis probably restricted to the southern high-latitude australprovince. Specimens which were previously identified as Hed-bergella mon;nouthensis at these high latitudes are now consid-ered as juvenile and kummerform adult forms of the newrugoglobigerinid species. The gerontic morphotypes of thistaxon, formerly considered as Rugoglobigerina pilula, R. rugosa,R. pustulata, and R. rotundata, show substantial variability inmaximum test diameter, cross-sectional area ratio of the ulti-mate and penultimate chambers, and position of the generatingcurve. They differ from typical species of Rugog!ohigerina in thatsurface ornamentation is predominantly composed of ran-domly arranged pustules, with only rare occurrence of mend-ionally aligned costellae. Kummerform chambers occur in mod-erate frequency and umbilical cover plates (tegilla) are rare.
1987 REVIEW 15
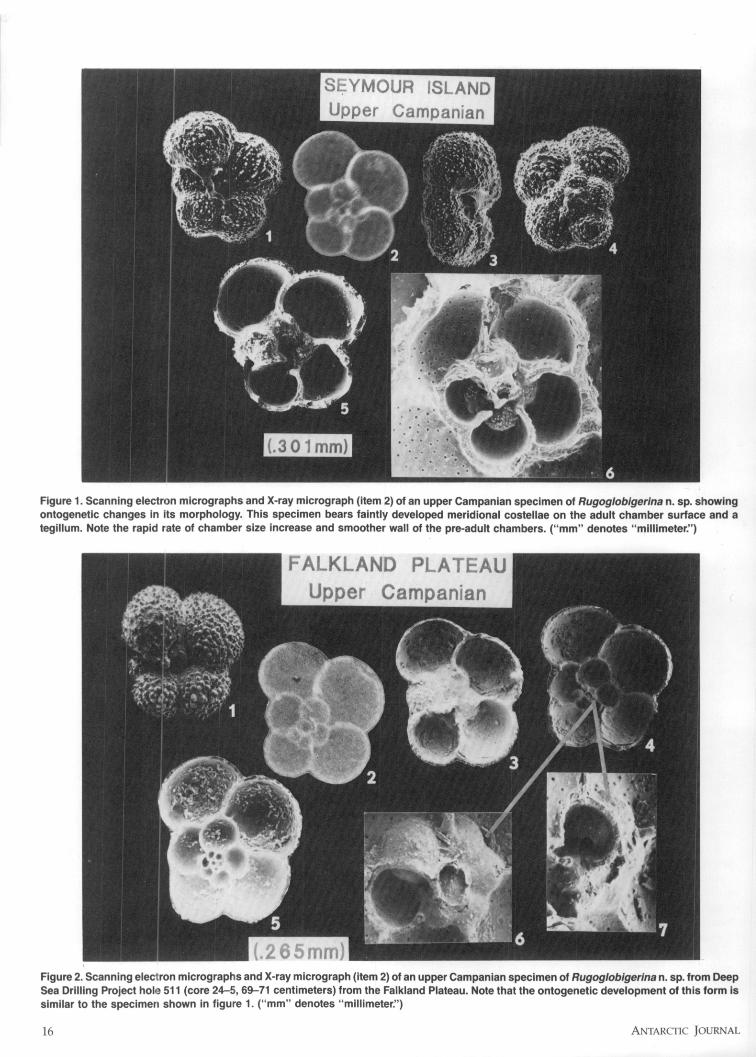
Figure 1. Scanning electron micrographs and X-ray micrograph (item 2) of an upper Campanian specimen of Rugoglobigerina n. sp. showingontogenetic changes in its morphology. This specimen bears faintly developed meridional costellae on the adult chamber surface and ategillum. Note the rapid rate of chamber size increase and smoother wall of the pre-adult chambers. ("mm" denotes "millimeter:')
Figure 2. Scanning electron micrographs and X-ray micrograph (item 2) of an upper Campanian specimen of Rugoglobigerina n. sp. from DeepSea Drilling Project hole 511 (core 24-5, 69-71 centimeters) from the Falkland Plateau. Note that the ontogenetic development of this form issimilar to the specimen shown in figure 1. ("mm" denotes "millimeter:')
16 ANTARCTIC JOURNAL

Examination of the pre-adult morphology of planktonic for-aminifera using the methods described above provides addi-tional criteria for understanding their taxonomy and phy-logenetic relationships. Although this study was concernedwith a single time-slice in the Late Cretaceous, analysis of strat-igraphic changes in the ontogenetic morphometry of particulartaxonomic groups will certainly improve high-latitude bio-stratigraphy and provide further insight to paleoceanographicand paleoclimatic evolution.
Samples provided by the Deep Sea Drilling Project (EastCoast Repository, Lamont-Doherty Geological Observatory)are gratefully acknowledged.
This research was partially funded by National Science Foun-dation grants DPP 85-17625 and Di p 84-20622. Jam grateful to thecurators at the Scripps Institution of Oceanography for makingthe Deep Sea Drilling Project samples available to me.
ReferencesArnold, A.J. 1982. Techniques for biometric analysis of foraminifera.
Third North American Paleontological Convention, Proceedings, 1, 13-15.
Huang, C. 1981. Observations on the interior of some late Neogeneplanktonic foraminifera. Journal of Foraminiferal Research, 1(3),173-190.
Huber, B.T. In press. Upper Campanian-Paleocene foraminifera fromthe James Ross Island region (Antarctic Peninsula). In R.M. Fel-dmann and M.O. Woodburne (Eds.), Geology and Paleontology ofSeymour Island, Antarctica. (Geological Society of America, MemoirSeries 169.)
Huber, B.T., D.M. Harwood, and P.N. Webb. 1983. Upper Cretaceousmicrofossil biostratigraphy of Seymour Island, Antarctic Peninsula.Antarctic Journal of the U.S., 18(5), 72-74.
Krasheninnikov, V.A., and I.A. Basov. 1983. Stratigraphy of Cretaceoussediments of the Falkland Plateau based on planktonic foraminifers,Deep Sea Drilling Project, Leg 71. In W.J. Ludwig, V.A. Krashenin-nikov, et al. (Eds.), Initial Reports of the Deep Sea Drilling Project.Washington, D.C.: U.S. Government Printing Office.
Sliter, W.V. 1976. Cretaceous foraminifera from the southwest AtlanticOcean, Leg 36, Deep Sea Drilling Project. In P.F. Barker, I.W.D.Dalziel et al. (Eds.), Initial Reports of the Deep Sea Drilling Project.Washington, D.C.: U.S. Government Printing Office.
Evidence from the Beardmore Glacierregion for a late Paleozoic/early
Mesozoic foreland basin
• An Early Permian facies transition from terrestrial to marinetoward the orogenic belt.
• The existence of two major source areas, the east antarcticcraton and calc-alkaline volcanics from a convergent paleo-
JAMES W. CoI,uNsoN and JOHN L. ISBELI.
Byrd Polar Research Ccii hrand
Department of Geology and MineralogyOhio State University
Columbus, Ohio 43220
---------
EM BASINlsrth Mtns
/ EASTANTARCTIC
£ Pensacola Mtns.CRATON-1
74'+ South Pole
New data and interpretations from our 1985-1986 field seasonin the Beardmore Glacier region lend support to the hypothesisthat the Upper Paleozoic/Lower Mesozoic sedimentary se-quence is part of a major foreland basin that paralleled themargin of the east antarctic craton. This foreland basin com-prises at least four distinct stratigraphic basins (figure 1):Ellsworth Mountains (EM), Central Transantarctic Mountains(cTM), southern Victoria Land (svL), and northern Victoria Land(NvL). The stratigraphy of these basins has been summarized atlength by Elliot (1975).
Foreland basins are elongated depressions that develop oncontinental crust, typically near the edge of a craton, inboard ofa fold/thrust belt. Lines of evidence supporting the forelandbasin hypothesis include:• The widespread similarity of stratigraphic sequences from
antarctic basins suggesting that they are genetically related.• Post-Early Permian to pre-Middle Jurassic folding in the
Ellsworth and Pensacola mountains indicating the existenceof an orogenic belt.
• The thickening of time-equivalent sequences toward theorogenic belt.
CTMLBASIN
Shackle tonBeardmore GI.
ROSS ICE SHELF
D Stratigraphic basins
500igooKilometers
Rennick GI.
Figure 1. Location map showing extent of stratigraphic basins.("EM" denotes "Ellsworth Mountains"; "CTM" denotes "centralTransantarctic Mountains"; "SVL" denotes southern VictoriaLand"; "NVL" denotes "northern Victoria Land?')
C6 NVLASIN
1987 REVIEW 17