on the similarity of tetanus and botulinum toxins
TRANSCRIPT

Naunyn-Schmiedeberg's Arch. Pharmacol. 276, 303--310 (1973) �9 by Springer-Verlag 1973
On the Similarity of Tetanus and Botulinum Toxins
Jane Mellanby, P. A. Thompson, and Nieola H a m p d e n
Department of Experimental Psychology, University of Oxford
Received June 22, 1972
Summary. 1. Botulinum toxin blocks neuromuscular transmission to the abduc- tor superficialis muscle of the goldfish pectoral fin. Since the botulinum-paralysed muscles are still responsive to direct electrical stimulation and to earbachol it is probable that the toxin acts (at least partly) presynaptieMly. The actions of botu- linum and tetanus toxins at this site are similar.
2. Tetanus toxin is fixed by nervous tissue and this fixation is due to ganglio- sides, in particular to those gangliosides eontaining two sialie acid residues joined to each other by a sialidase-sensitive bond. In contrast, botulinum toxin is not fixed by either nervous tissue or by gangliosides.
3. The implication of these findings for the role of the fixation of tetanus toxin by ganglioside in the mode of action of tetanus toxin are discussed.
Key words: Tetanus -- Botulinum -- Toxin -- Ganglioside --Fish.
When the clinical impressions of te tanus and botulism are considered, it is the differenees ra ther than the similarities t ha t s tand out. I n mam- mals the most obvious effect of te tanus intoxication is a spastic paralysis with convulsions, whereas botul inum toxin produces a flaccid paralysis. These two contrasting effects are generally believed to be produced by te tanus toxin blocking inhibition in the spinal cord while botul inum toxin blocks neuromuscular transmission. However, the clinical picture of spastic paralysis with te tanus toxin m a y conceal a botulinum-like action of the toxin. Under certain special circumstances it has been reported tha t the injection of te tanus toxin m a y lead to the development of flaccid ra ther than spastic symptoms. Thus Davies and Wright (1955) found t h a t when mice were injected int ravenously with a large dose of te tanus toxoid (330 Lf) short ly before they were given tetanus toxin (36 LDs0 ) they could be protected from the toxin. I f however the dose of toxoid was insufficient to protect the mice completely, then the mice exhibited symptoms of flaccid paralysis. Miyasaki et al. (1967) reported tha t a predominant ly flaccid paralysis could be produced in rabbits if intra- muscular injection of te tanus toxin was followed 2 h later by intravenous injection of antitoxin. Similar flaccidity could also be produced when the toxin was injected into the epineurium of the ischiadicus nerve. Tha t te tanus toxin co~:ld block neuromuscular transmission in a similar way to

304 J .H. Mellanby et al. :
botul inum toxin, at least at a parasympathe t ic junction, had previously been demonst ra ted by Ambache et al. (1948). I n their experiments it was found tha t both te tanus and botul inum toxins when injected into the anterior chamber of the rabbi t eye blocked transmission to the sphincter papillae muscle, thus abolishing the light reflex. Since then a number of workers have found evidence for a neuromuscular blocking action of te tanus toxin at other junctions, notably on mammal ian and fish skeletal muscle (Prabhu and Oester, 1962; P rabhu et al., 1962; Kaeser and Saner, 1970; Diamond and Me]lanby, 1970, 1971 ; Mellanby, 1971 ; Mellanby and Thompson, 1972; Duchen et al., 1972; Kryzhanovsky , 1972). Where this action has been investigated in detail, it appears t ha t the action is p re synap t i e - -bo th toxins block the release of t ransmit ter f rom the presh~naptie terminals (Harris and Miledi, 1971).
The similarity of the modes of action of te tanus and botul inum toxins at several neuromuscular junctions is in marked contrast to the difference between the two toxins in the context of fixation by nervous tissue or ganglioside. Tetanus toxin is fixed in an avid and specific way by nervous tissue and this has been shown to be due to those gangliosides which contain two sialic acid residues joined together by a sialidase-sensitive bond (van Heyningen, 1963; see also van Heyningen, 1973). Botul inum toxin does no t appear to be fixed by nervous tissue (Coleman, 1924; van t Ieyningen, unpublished).
The present paper presents experiments concerning the act ion of botul inum toxin at the neuromuscular junct ion in the goldfish, where it apparent ly acts similarly to te tanus toxin, and on the abil i ty of various preparat ions to fix te tanus toxin bu t not botul inum toxin.
Materials and Methods
Tetanus toxin was kindly supplied by Dr. R. O. Thomson of the Wellcome Research Laboratories: it contained 40~ protein of which 75 ~ was toxin (see van Heyningen, 1959). Dilutions of the toxin were made in Fish Ringer solution (see Diamond and Mellanby, 1971) containing 0.2~ gelatin. Toxin was assayed by injecting serial twofold dilutions into pairs of swiss albino mice. One LDh0 was defined as the amount of toxin which killed half the mice injected with it within 7 days.
Botulinum toxin was an ammonium sulphate precipitate of a crude culture filtrate of Clostridium botulinum type A. I t contained approximately 5000 LDh0 per rag. I t was also assayed by injecting serial twofold dilutions into pairs of Swiss albino mice. One LDh0 killed half the mice injected with it within 3 days.
The isolated nerve.muscle prepc~ration consisted of the abductor superficialis muscle still attached to the pectoral girdle at one end and to the fin at the other, with its nerve running through the pectoral girdle (Diamond and Mellanby, 1971).
Toxin-/ixing capacity was measured by adding constant volumes (usually 1 ml, containing 40 LDa0/ml ) of toxin to equal volumes of serial dilutions of suspensions of nervous or muscle tissue, or ganglioside-cerebroside complexes (van Heyningen and Mellanby, 1968), standing ibr 10 min at room temperature, centrifuging at

Similarity of Tetanus and Botulinum Toxins 305
20000 RPM for 15 min (40.2 head in Spinco model L) and injecting 0.5 ml volumes of the supernatant fluids into pairs of Swiss albino mice. In this way, the least amount of suspension required to fix 10 LDs0 doses of toxin could be determined (van I{eyningen, 1959).
Gangliosides and cerebrosides from beef brain were prepared according to the methods described by Mellanby et al. (1968). Ganglioside was prepared /tom fish muscle by extracting with 20 vols. of chloroform-methanol (1:2, v/v). 20 vols. of chloroform was then added to the combined extracts to bring the final proportion of chloroform to 2:1 and the gangliosides were separated into an upper phase by the procedure of Folch-Pi et at. (1957). After partitioning, the upper phase was dialysed against distilled water overnight at 4 ~ and then freeze-dried.
Results
E//ect o /Bo tu l inum Toxin on Neuromuscular Transmission in the Gold/ish
Local paralysis of the abductor superficialis muscle of the goldfish pectoral fin was produced by injection of 2 LDs0 (mouse) of botulinum toxin in gelatin Fish Ringer solution. The corresponding muscle of the opposite pectoral fin which had been injected with the gelatin Ringer solution alone served as a control. The fish were killed at a time when the paralysis of the muscle (as observed by fin movement) was just complete. The abductor superficialis muscle from the two fins was dissected out with its nerve and set up for recording tension in response to stimulation of either the nerve or of the muscle directly. Fig. 1 shows the tension recorded in the control and toxin-injected muscles in one fish 2 days after injection of the toxin. I t can be seen that while the response to nerve-stimulation is virtually absent in the toxin-injected muscle, the response to electrical stimulation of the muscle itself can still be elicited, although it is weaker.
The postsynaptic sensitivity to carbachol was also investigated in such just-paralysed muscles. It was ibund to be fairly similar both in magnitude and in time-course in the pairs of muscles.
I t would thus appear that botulinum toxin blocks neuromuscular transmission in the goldfish and that this effect involves a presynaptic action (though there is probably also some reduction in the response of the muscle to direct electrical stimulation).
Fixation o /Te tanus and Botul inum Toxins in vitro
I t was shown by van I-Ieyningen and Miller (1961) that the Wasser- mann-Takaki phenomenon of the fixation of tetanus toxin by nervous tissue was specific to tetanus toxin. Botulinum toxin (and a number of other toxins and enzymes) was not fixed by nervous tissue. Since the physiological actions of tetanus and botulinum toxins are apparently similar at some sites, it might be expected, if the fixation of tetanus toxin by ganglioside is relevant to its action, that under suitable conditions botulinnm toxin would also be fixed by ganglioside.
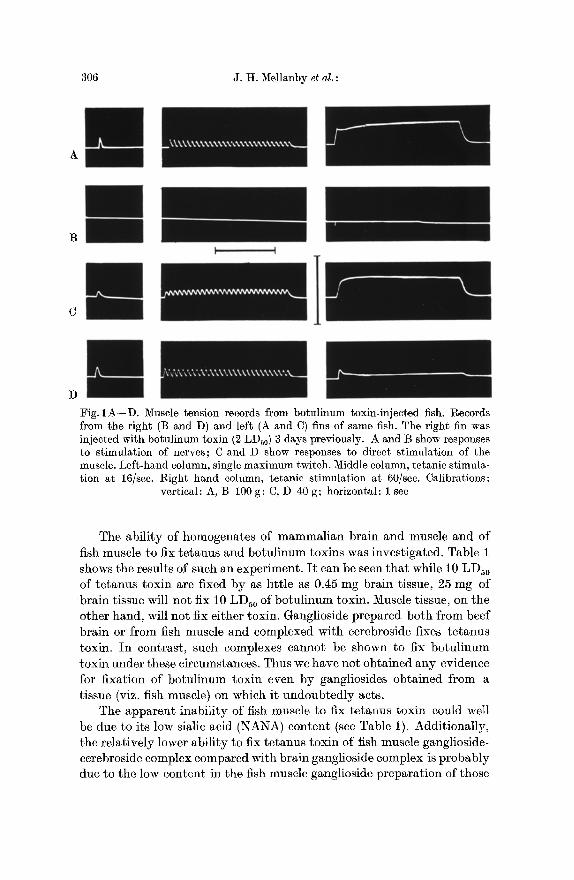
306 J .H . Mellanby et al . :
A
B
D
Fig. 1A--D. Muscle tension records from botulinum toxin-injected fish. Records from the right (B and D) and left (A and C) fins of same fish. The right fin was injected with botulinum toxin (2 LDs0 ) 3 days previously. A and B show responses to stimulation of nerves; C and D show responses to direct stimulation of the muscle. Left-hand column, single maximum twitch. Middle column, tetanic stimula- tion at 16/see. Right hand column, tetanie stimulation at 60/see. Calibrations:
vertical: A, B 100 g ; C, D 40 g ; horizontal: 1 see
The ab i l i ty of homogenates of m a m m a l i a n bra in and muscle and of fish muscle to fix t e t anus and bo tu l inum toxins was inves t iga ted . Table 1 shows the resul ts of such an exper iment . I t can be seen t h a t while 10 LD~0 of t e t anus t ox in are fixed b y as l i t t le as 0.45 mg bra in t issue, 25 mg of b r a i n t issue will not fix 10 LDs0 of bo tu l inum toxin. Muscle t issue, on the o ther hand, will no t fix e i ther toxin. Ganglioside p repa red bo th from beef b ra in or f rom fish muscle and complexed wi th cerebroside fixes t e t anus toxin. I n contrast , such complexes cannot be shown to fix bo tu l inum tox in under these circumstances. Thus we have no t ob ta ined any evidence for f ixat ion of bo tu l inum tox in even b y gangliosides ob ta ined f rom a t issue (viz. fish muscle) on which i t u n d o u b t e d l y acts.
The a p p a r e n t i nab i l i t y of fish muscle to fix t e t anus t ox in could well be due to i ts low sialic acid (NANA) conten t (see Table 1). Addi t iona l ly , the re la t ive ly lower ab i l i ty to fix t e t anus tox in of fish muscle ganglioside- cerebroside complex compared with bra in ganglioside complex is p r o b a b l y due to the low content in the fish muscle ganglioside p repa ra t ion of those
Similarity of Tetanus and Botul inum Toxins 307
Table 1. Comparison of the abil i ty of tissue homogenates, and of complexes of the gangliosides derived from them, to fix te tanus and botul inum toxins
Amount needed to fix 10 LDa0 toxin
Tetanus toxin Botul inum toxin
wet wt. lipid wet wt. lipid tissue NANA a tissue NANA ~ mg ~g mg ~g
Tissue homogenates Beef brain Guineapig muscle Goldfish muscle
Ganglioside-cerebroside somplexes Source of Ratio ganglioside C: G
Beef brain 1 : 3 Goldfish muscle 1 : 3 Goldfish muscle 4:1
0.45 0.225 > 25 > 12.5 > 25 > 0.225 > 25 > 0.225 > 25 > 0.150 > 25 > 0.150
wt. complex NANA wt. complex NANA mg izg mg tzg 0.001 0.06 > 2.5 > 150
0.25 1.3 -- - -
0.03 0.5 > 1.4 > 23
NANA (N-acetyl-neuraminic acid, sialic acid).
Brain Fishmusc[e
Fig.2. Thin layer chromatograms of gangliosides (Penick el al., 1966) from brain and fish muscle. On the fish muscle chromatogram only those bands m a r k e d , stained
positively with the resorcinol reagent for gangliosides

308 J .H. Mellanby et al . :
gangliosides which are specifically involved in tetanus toxin fixation (see van Heyningen, 1963). Fig.2 shows a thin layer chromatogram of fish muscle and beef brain gangliosides, and demonstrates the paucity of these gangliosides in the fish muscle ganglioside preparation. The fish ganglioside preparation appears to consist mainly of the disialoganglio- side (SGGnSLC) which does not fix tetanus toxin, and another ganglioside with a much lower Rf value than the two tetanus toxin-fixing gangliosides (SGGnSSLC and GGnSSLC) which might be a tetrasialoganglioside or the other trisialoganglioside (SSGGnSLC) found in brain by Penick and McCluer (1965).
Discussion
The present experiments indicate tha t botulinum toxin blocks neuro- muscular transmission in the goldfish, as it is well known to do in mam- mals. The effect is at least par t ly presynaptic as the botulinum-paralysed muscles are still responsive to carbachol and to direct electrical stimula- tion. Tetanus toxin has been shown to block transmission to this muscle, presynaptically (Mellanby, 1971; Mellanby and Thompson, 1972) and thus at this site tetanus and botulinum toxins act similarly. I f the action of tetanus toxin at the neuromuscular junction in the fish (Mellanby and Thompson, 1972) is compared with the action of botulinum toxin at the neuromuscular junction in the frog (Harris and Miledi, 1972) the actions appear similar in detail except that in the botulinum-poisoned frog muscles a few, small miniature end plate potentials persist whereas these have not been detected in the fish muscle poisoned with tetanus toxin.
One practical difference between the two toxins is that the blocking actions of botulinum toxin may readily be demonstrated on addition of the toxin to the bathing medium of a muscle whereas it has usually been found necessary to inject tetanus toxin into the body of the muscle (or as in the case of the rabbit eye to introduce it into a small volume of fluid in especially close contact with the muscle, the anterior chamber of the eye) in order to produce a neuromuscular block.
The inability of nervous tissue and of ganglioside-cerebroside com- plexes to fix botulinum toxin shows that only one of the two clostridial neurotoxins which block neuromuscular transmission presynaptically possesses an affinity for this "receptor" substance. In view of the similar- i ty of the actions of the two toxins this might be interpreted as indicating tha t the avid and specific fixation of tetanus toxin by nervous tissue and by ganglioside is irrelevant to its mode of action. We have made the preliminary observation that although rat cerebral cortex tissue will fix tetanus toxin, yet the injection of toxin into the motor or visual cortex does not apparently lead to any electroencephalographic abnormalities. This suggests tha t the toxin can be fixed by a tissue without apparently

Similarity of Tetanus and Botulinum Toxins 309
act ing on it. The possibil i ty t ha t the fixation of t e tanus tox in was no t
necessarily par t of its act ion has always been borne in mind since it was
originally shown tha t the fixation of tox in was t ime and tempera ture independen t (van t t eyn ingen , 1959) whereas the lethal act ion of the tox in in frogs and fish was s t rongly t empera ture dependent (l~owson, 1962; Diamond and Mellanby, 1971).
We would like to thank Dr. W. E. van I-Ieyningen for helpful discussions and the Medical Research Council and the Muscular Dystrophy Group of Great Britain for financial support.
References
Ambache, ~., Morgan, R. S., Wright, G. P. : The action of tetanus toxin on the rabbit's iris. J. Physiol. (Lond.) 107, 45--53 (1948).
Coleman, G. E. : Action of leucocytes and brain tissue on toxin of B. botulinus. J. infect. Dis. 34, 614--620 (1924).
Davies, J. R., Wright, E. A. : The specific precocious protective action of tetanus r Brit. J. exp. Path. 86, 487--493 (1955).
Diamond, J., Mellanby, J. H. : The effect of tetanus toxin in the goldfish. J. Physiol. (Lond.) 210, 186--1871 ) (1970).
Diamond, J., Mellanby, J. H. : The effect of tetanus toxin in the goldfish. J. Physiol. (Lond.) 215, 727--741 (1971).
Duchen, L. W., Stolkin, C., Tonge, D. A. : Changes in neuromuscular transmission in slow and fast skeletal muscles of the mouse after local injection of tetanus toxin. J. Physiol. (Lond.) 222, 147 p--148P (1972).
Folch-1)i, J., Lees, M., Sloane-Stanley, G. H. : A simple method for the isolation and purification of total lipids from animal tissues. J. biol. Chem. 226, 497--509 (1957).
Harris, A. J., Miledi, R. : The effect of type D botulinum toxin on frog neuromuscular junctions. J. 1)hysiol. (Lond.) 217, 495--515 (1971).
Kaeser, H. E., Saner, A. : The effect of tetanus toxin on neuromuscular transmission. Europ. Neurol. 8, 193--205 (1970).
Kryzhanovsky, G. ~. : The mechanism of action of tetanus toxin: effect on synaptic processes and some particular features of toxin binding by the nervous tissue. Naunyn-Schmiedeberg's Arch. 1)harmacol. 276, 247--270 (1973).
Mellanby, J. H. : 1)resynaptic effect of tetanus toxin at the neuromuscular junction. J. Physiol. (Lond.) 218, 68--69P (1971).
Mellanby, J. H., Pope, D., Ambache, N. : The effect of the treatment of crude teta- nus toxin with ganglioside-cerebroside complex on sphincter paralysis in the rabbit's eye. J. gen. Microbiol. 50, 479--486 (1968).
Mellanby, J. tt., Thompson, 1). A. : The effect of tetanus toxin at the neuromuscular junction in the goldfish. J. Physiol. 224, 407--421 (1972).
Miyasaki, S., Okada, K., Muto, S., Itokazu, T., ~atsui, M., Ebisawa, I., Kagabe, K., Kimuro, T. : On the mode of action of tetanus toxin in rabbit. I. Distribution of tetanus toxin in vivo and development of paralytic signs under some conditions. Jap. J. exp. ~Ied. 37, 217--225 (1967).
1)enick, R. J., McCluer, R. H. : Evidence for a second trisialoganglioside. Biochim. biophys. Acta. (Amst.) 106, 435--438 (1965).
1)enick, R. J., Meisler, M. H., McCluer, R. H. : Thin layer chromatographic studies of human brain gangliosides. Biochim. biophys. Acta (Amst.) 116, 279--287 (1966).

310 J .H. Mellanby et al. : Similarity of Tetanus and Botulinum Toxins
Prabhu, V. G., Oester, Y. T. : Electromyographic changes in skeletal muscle due to tetanus toxin. J. Pharmacol. exp. Ther. 188, 241--248 (1962).
Prabhu, V. G., Oester, Y. T., Karczmar, A. G. : Some observations on the actions of certain pharmacological agents on the fibrillation potentials and on the sensitivity of the skeletal muscle of rabbit in advanced local tetanus. Int. J. Neuropharma- col. I , 371--379 (1962).
P~owson, I~2. E. K. : The action of tetanus toxin in frogs. J. gem Mierobiol. 25, 315--329 (1961).
Van Heyningen, W. E. : Chemical assay of the tetanus toxin receptor in nervous tissue. J. gen. Microbiol. 20, 301--309 (1959).
Van Heyningen, W. E.: The fixation of tetanus toxin, strychnine, serotonin and other substances by ganglioside. J. gen. Microbiol. 81,375--389 (1963).
Van Heyningen, W. E. : On the similarity of tetanus and cholera toxins. Naunyn- Sehmiedeberg's Arch. PharmacoI. 276, 289--295 (1973).
Van Heyningen, W. E., Mellanby, J. H. : The effect of cerebroside and other lipids on the fixation of tetanus toxin by gangliosides. J. gen. Microbiol. 52, 447--454 (1968).
Van Heyningen, W. E., Mellanby, J. H.: Tetanus Toxin. In: Microbial Toxins, vol. l lA. S. Kadis, D. C. ~ontie and S. J. Ajl, ed. New York: Academic Press 1971.
Van Heyningen, W. E., Miller, P. A. : The fixation of tetamls toxin by ganglioside. J. gen. Microbiol. 24, 107--119 (1961).
J. Mellanby Department of Experimental Psychology University of Oxford South Parks lload Oxford OX 1 3PS, England