on the relationship between two indices (“bulk density” and “dry-matter content”) of...
TRANSCRIPT

Bot. Mag. Tokyo 98 : 335-345, 1985 The Botanical Magazine, Tokyo ~) by The Botanical Society of Japan 1985
On the Relationship between Two Indices
("Bulk Density" and "Dry-Matter Content")
of Dry-Matter Accumulation
in Plant Organs
YOTA YOKOI AND AKIHIKO KISHIDA
Department of Biology, Faculty of Science, Ibaraki University, Mito 310
"Bulk density" and "dry-matter content" are useful indices of dry-matter accumu- lation in plant organs. A theoretibal equation describing the relationship between these two indices was put forward. To examine the reliability of this equation, the seasonal changes of these two indices were investigated in the leaves and stems of different ages of Aucuba japonica. In each organ both indices varied seasonally almost parallel to each other, but the seasonal changes of dry-matter content were less obvious than those of bulk density. The observed bulk density was always larger than that calculated from the observed dry-matter content by the theoretical equation. A drying experi- ment showed that this discrepancy was caused by the decrement of the volume of the plant material by water loss during the period from the weight measurement to the volume measurement. When the water loss was negligible, the equation described well the relationships between the two indices of the experimental materials. It was also shown that this equation was useful for the estimation of the amount of air space in plant materials.
Key words : Air space -- Aucuba japonica -- Bulk density -- Dry-matter content Dry-matter economy -- Reserve material.
I t is impor tant in studies of dry-matter economy of plants to investigate the
accumulat ion of dry mat ter in the organs. In the s tudy of dry-mat ter production in
Miscanthus sinensis, Midorikawa et al. (1963) measured "bulk density", i.e., the weight
of dry mat ter per unit fresh volume of plant material, of the rhizome. From the
seasonal change in this index, they estimated the amount of accumulation and
consumption of reserve materials in the organ. This method was recommended as
useful for studies of the product ion process in higher plants ( Iwaki and Midorikawa,
1968 ; Kimura, 1976), and has been used in some ecological studies (Mutoh et al., 1968 ;
Fiala, 1973; Ino, 1977; Masuzawa, 1977; Kobayashi, 1981; Yamamura, 1984).
Masuzawa and Hogetsu (1977) measured nonstructural carbohydrate content of
the rhizomes of Miscanthus sacchariflorus, and observed a parallel relationship between
the seasonal changes in bulk density and nonstructural carbohydrate content. This
observation confirmed tha t bulk density is a good index of the content of reserve
materials in a plant organ.

336 Y. YOKOI AND A. KISHIDA
On the other hand, in the growth-analytical studies by the English school, dry weight/fresh weight ratio (dry weight percentage) has been used as an anatomical index of plant materials (cf. Evans, 1972). This ratio gives the content of dry matter in fresh plant material, so it is called "dry-matter content", and is also used in some studies of matter economy of plants (e.g., van Andel, 1975 ; Kimura et al., 1983).
Thus, we have two indices, "bulk density" and "dry-matter content", to express the degree of dry-matter accumulation in plant organs. However, the properties of these two indices and the relationship between them have not yet been investigated thoroughly.
First in this study, we have introduced theoretically an equation describing the relationship between these two indices. Next, in order to examine the reliability of this equation, two surveys were done with A u e u b a j a p o n i c a . They were an observa- tion of the seasonal changes in the two indices of stems and leaves of different ages, and a drying experiment with the stems.
Equation of Relationship between Bulk Density and Dry-matter Content
Fresh volume (Vf ; ml) of a plant material can be described by the following equation,
V+-= W + - W~ + W~ + V ~ (1) P~ P~
[ ~ ar ~ l where, We and Wj are the dry and fresh weights (g) of the material, Pw and pe are the densities (g/m/) of
T the matter lost by drying and of the dry matter respectively, and Va is the volume of air space (m/) included in the fresh plant material.
'. vo ,. r - , br In some cases, the values of Vj and Va of the ,. . . . . . , l same material may vary with the methods of volume
• , " - , measurement. For example, we may consider a cylindrical material having a diameter of a em and
~ _ ~ a height of bem, and which has an open air space (%
Fig. 1. A schematic plant m/) and a closed one (Vc m/) (Fig. 1). When the material which is cylindri- shape of this material is approximated to a right eal and has an open (%) cylinder, fresh volume (Vi) is given geometrically as and a closed air spaces (vc). ~r �9 a 2 �9 b /4 and V~ is vo+Vc. On the other hand,
when we use another volumetric method excluding the open air space, e.g., an air-comparison pycnometer, then VI and V~ are ~ �9 a 2 �9 b /
4 - % and v~, respectively. From Eq. (1),
Bulk Density and Dry-Matter Content 337
w~ w,, _ m , . p~,. w , (2)
wd Va Vs pd-(p,~-p,,,) " ~ + p ~ , * Pal" Ws
where Va/We is an index of the amount of air space included in a plant material, we
call this "air-space index", and use the symbol " y " to represent it.
Bulk density (B ; g /m/) and dry-matter content (D ; g/g) are defined as
B _ Wd vs (3).
D - Wd Ws
Then, we get the following equation describing the relationship between B and D ( B -
D relationship) from Eq. (2).
B = P w ' P d " D (4) Pd--(Pd--Pw) " D + p w �9 Pd * $
We can assume that p~ is 1.00, because the main component of the matter lost by
drying should be water. Thus, Eq. (4) can be approximately expressed by the
following equation.
D B = 1 1 (4') 1-( --Pd)'D+r
Eq. (4') shows that two parameters, the density of dry matter (Pd) and air-space
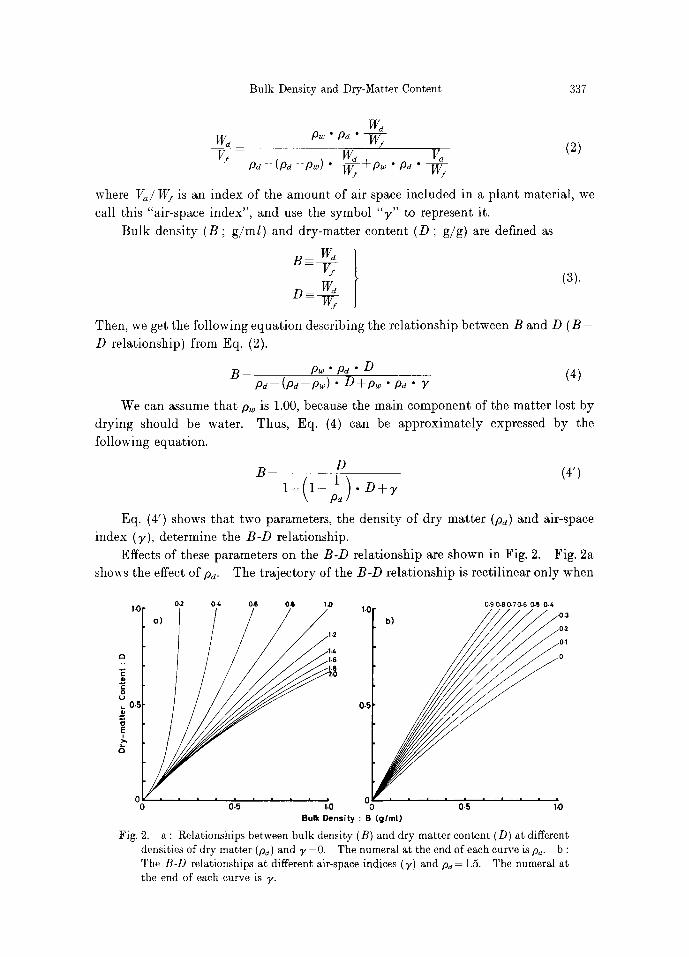
index (y), determine the B - D relationship. Effects of these parameters on the B - D relationship are shown in Fig. 2. Fig. 2a
shouts the effect of Pa. The trajectory of the B - D relationship is rectilinear only when
0.2 0*4 0-6 0-8 1.0 0.90-80,?0-6 0-5 0,4
b) //, 02 1.2 0-1
0 1.6 0
O5
~>" 0 ~ 01 . . . . OiS . . . . 110 0 0.5 1.0
Bulk Density : B (g lml )
Fig. 2. a : Relationships between bulk density (B) and dry-matter content (D) at different densities of dry matter (pd) and 3'- 0. The numeral at the end of each curve is pa. b : The B-D relationships at different air-space indices (7) and pa= 1.5. The numeral at tile end of each curve is y.

338 Y. YoKoi AND A. KISHIDA
Pd = 1. When Pd > 1, the traiectories are convex, and when there is no air space ( y -
0), B is always larger than D. When p d < l , then the traiectories are concave. The
degree of concavity or convexity increases with the increment of discrepancy of ps
from unity. The effect of ~, is shown in Fig. 2b. Existence of air space reduces only the value
of B, not affecting the value of D. Thus, the trajectory of the B-D relationship shifts
upward with the increment of ~,.
Observations and Experiments
Material and Methods Aucuba japonica Thunb., which is a broad-leaved evergreen woody plant, was
used as a plant material. This plant is dioecious and usually an understory species.
The collection site of the material was a floor of a Cryptomeria japonica forest near
Ibaraki University in Mito, Ibaraki Pref. (36 ~ 24" N, 140 ~ 26' E). On this site, A.
japonica is the dominant understory tree species, and other tree species are scarce.
Mean relative light intensity at breast height on this forest floor ranged from about
three to four percent of open site in summer and from seven to eight percent in winter.
For the observation of seasonal changes in the bulk density and dry-matter
content of the steins and leaves of different ages, male plants of A. japonica of about
1.5 m high were labelled in early May, 1979. The above ground part of one plant was
harvested at random from these plants at intervals of about one month from May 1979
to February 1980. The harvested plant was quickly carried into a laboratory of Ibaraki University.
At first the current year branches (0-year-old organs) were cut off. Then the branches
were subdivided into leaves and stems. Each organ was cut into small pieces, and a
part of them was used as a sample. Three replicated samples were taken for each
organ, and their fresh weight was determined. Subsequently, 1-, 2-, and 3-year-old
organs were treated in the same manner as 0-year-old organs. These samples were
packed in paper bags after their fresh weight determinations, and left at room tempera-
ture until their volume measurements. An air-comparison pycnometer (Beckman
model 930) was used for the volume measurement. Then, all the samples were dried
at 80 C to constant weights to determine dry weights. The dry samples were ground
into powder by a cutt ing mill (Wiley type) and a porcelain mortar. These powdered
samples were dried again at 80 C and their actual densities were estimated by the
pycnorneter. A drying experiment was carried out on March 9, 1980 with 0- and 2-year-old
stems. Two replicated samples were prepared for each stem. The volume and weight
of each sample were measured successively. Then, the sample was put in a drying oven with air circulation at 80 C. Each sample was taken out of the oven at intervals
of about one hour, and its volume and weight were rapidly determined. The integrat-
ed drying time was recorded at the time of each measurement. All samples were
ground into powder after the experiment, and their densities of dry matter were
estimated.
Bulk Density and Dry-Matter Content 339
Results and Discussion
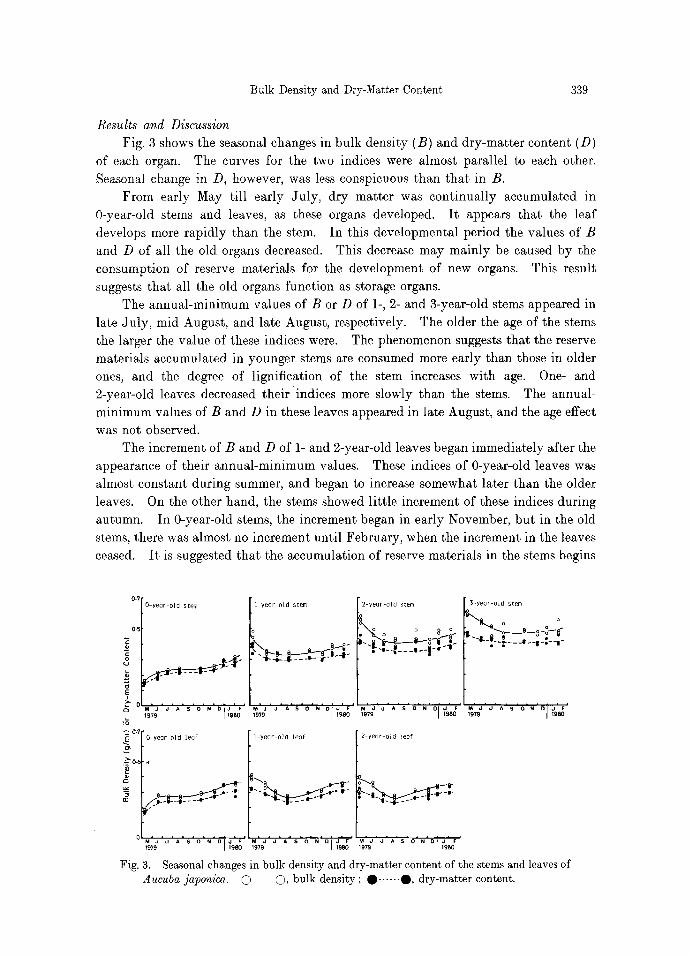
Fig. 3 shows the seasonal changes in bulk density (B) and dry-matter content (D) of each organ. The curves for the two indices were almost parallel to each other. Seasonal change in D, however, was less conspicuous than that in B.
From early May till early July, dry matter was continually accumulated in 0-year-old stems and leaves, as these organs developed. It appears that the leaf develops more rapidly than the stem. In this developmental period the values of B and D of all the old organs decreased. This decrease may mainly be caused by the consumption of reserve materials for the development of new organs. This result suggests that all the old organs function as storage organs.
The annual-minimum values of B or D of 1-, 2- and 3-year-old stems appeared in late July, mid August, and late August, respectively. The older the age of the stems the larger the value of these indices were. The phenomenon suggests that the reserve materials accumulated in younger stems are consumed more early than those in older ones, and the degree of lignification of the stem increases with age. One- and 2-year-old leaves decreased their "indices more slowly than the stems. The annual- minimum values of B and D in these leaves appeared in late August, and the age effect was not observed.
The increment of B and D of 1- and 2-year-old leaves began immediately after the appearance of their annual-minimum values. These indices of 0-year-old leaves was almost constant during summer, and began to increase somewhat later than the older leaves. On the other hand, the stems showed little increment of these indices during autumn. In 0-year-old stems, the increment began in early November, but in the old stems, there was almost no increment until February, when the increment in the leaves ceased. It is suggested that the accumulation of reserve materials in the stems begins
0-7
0-5
O-year-old stem
.d~..~____=~4'-4
1 year old stem
)
\ - s J ' ~ g " w- -o - -g - - - - - /
o2-year -old stern 3-year-aid stem
r ~ ' ~ ~ ~ 8 ~ o
o-~
M ' J ' J ' A ' S ' O ' N ' D ' J ' F . . . . 01979 r199o 1979zJ'J A S O" N' DI d' F I 1 [ 1980 1979' J" J " A" $' O" N" D'11980J" F" 1979M" J" J" A" 5 "O" N" D" J" F I J 1990
E 0 year-old leof 1 year old leaf E-year aid leaf
f Y - >
Fig. 3. Seasonal changes in bulk density and dry-matter content of the stems and leaves of Aucuba japonica. (2) �9 bulk density ; �9 ...... O, dry-matter content.
340 Y. YOKO[ AND A. KISHIDA
0.5 (1) O-ye<:lr-otd s t e m / / "" / 1.43
0.3 , ~ l . S Z
/ / / t x I I
Q - - / / o
~ ~ ~
o I 0,3 0.5 ~' o.s 0 r O-year-old leaf / i E ~,, / / 1.31
j /c /" / "41
0.3 / / / x~
, / I zz ////// % ; ' o.'3 ' o:s
0.6 b ) 2-year-old stem ,, "" / /
/ / 142 /rz~1,51
/ / / 0
0.4 / "
~~xxo n
/ /
, /
0.20.2 0.4 0-6 0.f / d } 2-year-old leaf ,"
/ t /
/ / / 1 - 3 ~ , / / / 1 . ~ 0
///
O.Z* /I/ / ///~ / / / / / / / 0
~/"~ . . . . 020.2 0.4 0.6
Bulk Density : B (glml) Fig. 4. Relationships between bulk density and dry-matter content in stems and leaves of
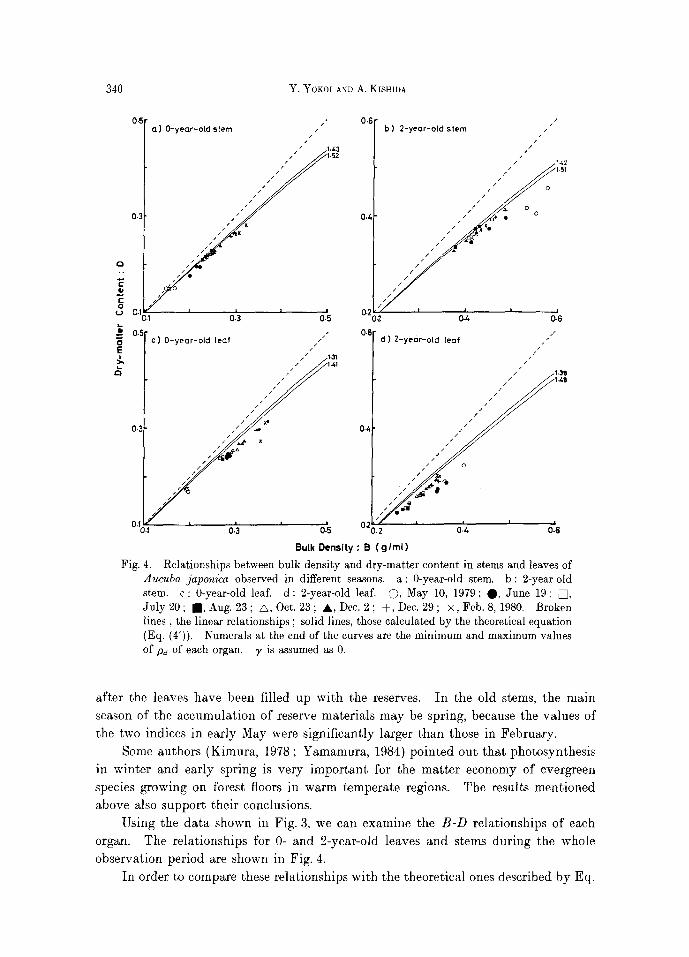
Aucuba japonica observed in different seasons, a: 0-year-old stem. b: 2-year-old stem. c: 0-year-old leaf. d: 2-year-old leaf. O, May 10, 1979; O, June 19; [] , July 20; re, Aug. 23; A, Oct, 23; &,Dec. 2; + ,Dec . 29; • 8,1980. Broken l ines, the linear relationships ; solid lines, those calculated by the theoretical equation (Eq. (4')). Numerals at the end of the curves are the minimum and maximum values of pd of each organ. ~, is assumed as 0.
after the leaves have been filled up with the reserves. In the old stems, the main season of the accumulation of reserve materials may be spring, because the values of the two indices in early May were significantly larger than those in February.
Some authors (Kimura, 1978 ; Yamamura, 1984) pointed out that photosynthesis in winter and early spring is very important for the matter economy of evergreen species growing on forest floors in warm temperate regions. The results mentioned above also support their conclusions.
Using the data shown in Fig. 3, we can examine the B-D relationships of each organ. The relationships for 0- and 2-year-old leaves and stems during the whole observation period are shown in Fig. 4.
In order to compare these relationships with the theoretical ones described by Eq.
Bulk Density and Dry-Matter Content 341
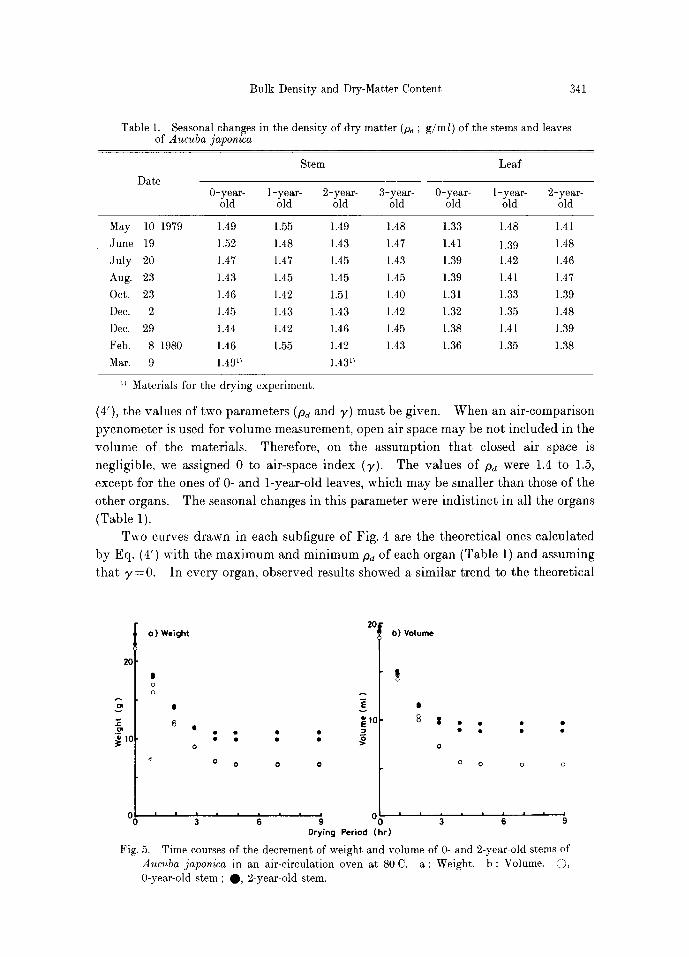
Table 1. Seasonal changes in the density of dry matter (pd ; g/m/) of the stems and leaves of Aucuba japonica
Date
Stem Leaf
0-year- 1-year- 2-year- 3-year- 0-year- 1-year- 2-year- old old old old old old old
May 10 1979 1.49 1.55 1.49 1.48 1.33 1.48 1.41
June 19 1.52 1.48 1.43 1.47 1.41 1.39 1.48
July 20 1.47 1.47 1.45 1.43 1.39 1.42 1.46
Aug. 23 1.43 1.45 1.45 1.45 1.39 1.41 1.47
Oct. 23 1.46 1.42 1.51 1.40 1.31 1.33 1.39
Dec. 2 1.45 1.43 1.43 1.42 1.32 1.35 1.48
Dec. 29 1.44 1.42 1.46 1.45 1.38 1.41 1.39
Feb. 8 1980 1.46 1.55 1.42 1.43 1.36 1.35 1.38 Mar. 9 1.4911 1.4311
~ Materials for the drying experiment.
(4'), the va lues of two p a r a m e t e r s (Pd and y) mus t be given. W h e n an a i r -compar i son
p y e n o m e t e r is used for v o l u m e measurement , open ai r space m a y be no t inc luded in the
vo lume of the mater ia l s . Therefore , on the a s sumpt ion t h a t c losed air space is
negl ig ib le , we ass igned 0 to a i r -space index (:y). The va lues of p~ were 1.4 to 1.5,
excep t for the ones of 0- and l - y e a r - o l d leaves, which m a y be smal le r t h a n those of the
o ther organs. The seasonal changes in th is p a r a m e t e r were i n d i s t i n c t in all the organs
(Table 1).
Two curves d r a w n in each subfigure of Fig. 4 are the theore t i ca l ones ca l cu l a t ed
b y Eq. (4') w i th the m a x i m u m and m i n i m u m pd of each organ (Tab le 1) and assuming
t h a t ~, = 0. In eve ry organ, obse rved resul ts showed a s imi l a r t r e n d to the theore t ica l
20
o~ v
Jr ot
0
a ) Weight
| o o
@
o
O o o
, ~ , , ;
E
o Drying Period ( h r )
Fig. 5. Aucuba japonica in an air-circulation oven at 80 C. 0-year-old stem ; O, 2-year-old stem.
b ) Volume
O
8 s :
o
o O o 0
Time courses of the decrement of weight and volume of 0- and 2-year-old stems of a : Weight. b : Volume. �9
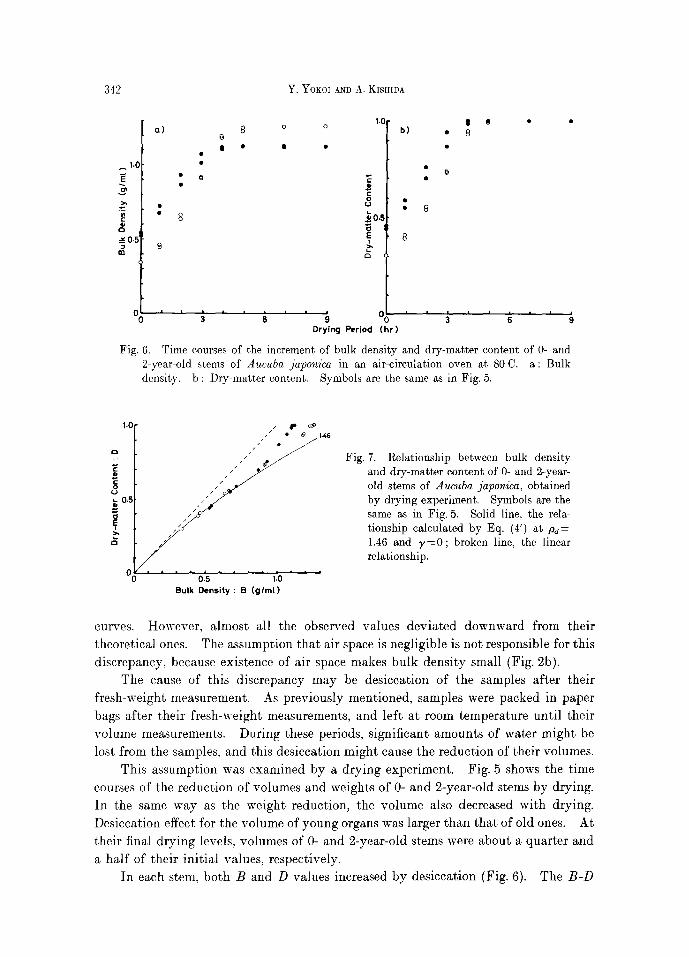
342 Y. YOKOI AND A. KISHIDA
1.0
_~ 0.51
a )
�9 0
8 o o
O �9 �9 �9
1.0 b)
0.5
E 8
0
| �9 8
Drying Period ( h r )
Time courses of the increment of bulk density and dry-matter content of 0- and a : Bulk
Fig. 6. 2-year-old stems of Aucuba japonica in an air-circulation oven at 80 C. density, b : Dry-matter content. Symbols are the same as in Fig. 5.
1.0 + ~p +, ] �9 @ 146
~ 0.5 +" + ] /
S
o ~ + . . . . 0 : s . . . . i o " " "
Bulk Density: B ( g l m l )
Fig. 7. Relationship between bulk density and dry-matter content of 0- and 2-year- old stems of Aucuba japonica, obtained by drying experiment. Symbols are the same as in Fig. 5. Solid line, the rela- tionship calculated by Eq. (4') at p,~= 1.46 and ~,=0; broken line, the linear relationship.
curves. However, almost all the observed values deviated downward from their
theoretical ones. The assumption tha t air space is negligible is not responsible for this
discrepancy, because existence of air space makes bulk density small (Fig. 2b).
The cause of this discrepancy may be desiccation of the samples after their
fresh-weight measurement. As previously mentioned, samples were packed in paper
bags after their fresh-weight measurements, and left at room temperature until their
volume measurements. During these periods, significant amounts of water might be
lost from the samples, and this desiccation might cause the reduction of their volumes.
This assumption was examined by a drying experiment. Fig. 5 shows the time
courses of the reduction of volumes and weights of 0- and 2-year-old stems by drying.
In the same way as the weight reduction, the volume also decreased with drying.
Desiccation effect for the volume of young organs was larger than that of old ones. At
their final drying levels, volumes of 0- and 2-year-old stems were about a quarter and
a half of their initial values, respectively.
In each stem, both B and D values increased by desiccation (Fig. 6). The B-D
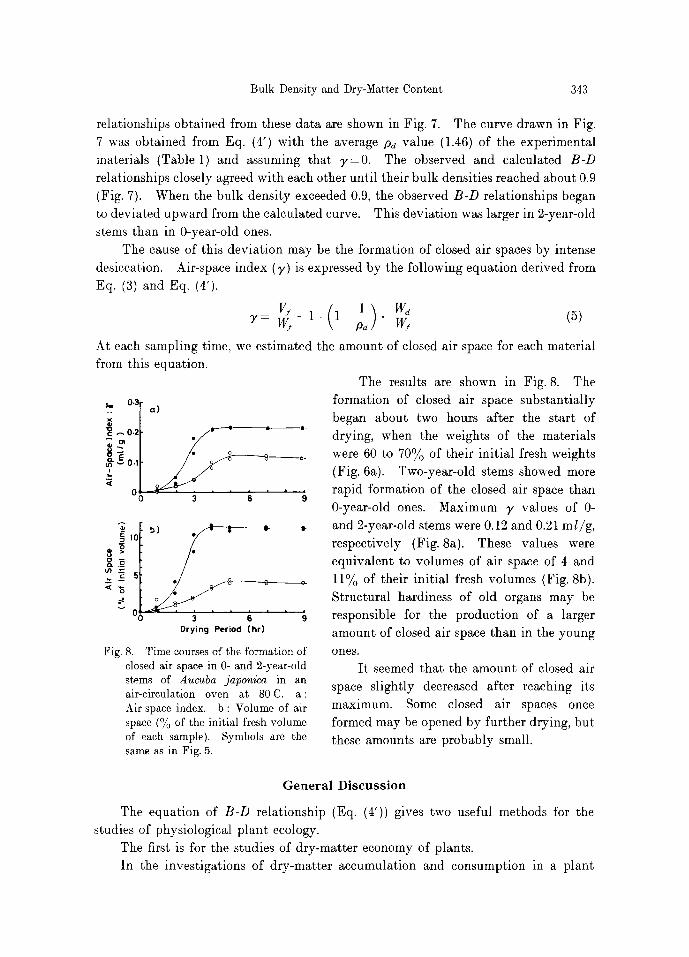
Bulk Density and Dry-Matter Content 343
relationships obtained from these data are shown in Fig. 7. The curve drawn in Fig. 7 was obtained from Eq. (4') with the average Pd value (1.46) of the experimental materials (Table l) and assuming that ~,=0. The observed and calculated B-D
relationships closely agreed with each other until their bulk densities reached about 0.9 (Fig. 7). When the bulk density exceeded 0.9, the observed B-D relationships began to deviated upward from the calculated curve. This deviation was larger in 2-year-old stems than in 0-year-old ones.
The cause of this deviation may be the formation of closed air spaces by intense desiccation. Air-space index (~) is expressed by the following equation derived from Eq. (3) and Eq. (4").
~=wT~ -I w, (5) At each sampling time, we estimated the amount of closed air space for each material from this equation.
~_ 0.3
~ o a
~e 0.,
o
~10
s
Fig. 8.
a)
3 6 9
b) .~41~II i s-
/
3 6 ; Drying Period (hr)
Time courses of the formation of closed air space in 0- and 2-year-old stems of Aucuba japonica in an air-circulation oven at 80C. a: Air-space index, b : Volume of air space (% of the initial fresh volume of each sample). Symbols are the same as in Fig. 5.
The results are shown in Fig. 8. The formation of closed air space substantially began about two hours after the start of drying, when the weights of the materials were 60 to 70% of their initial fresh weights (Fig. 6a). Two-year-old stems showed more rapid formation of the closed air space than 0-year-old ones. Maximum ~/ values of 0- and 2-year-old stems were 0.12 and 0.21 m//g, respectively (Fig. 8a). These values were equivalent to volumes of air space of 4 and 11% of their initial fresh volumes (Fig. 8b). Structural hardiness of old organs may be responsible for the production of a larger amount of closed air space than in the young ones.
It seemed that the amount of closed air space slightly decreased after reaching its maximum. Some c]osed air spaces once formed may be opened by further drying, but these amounts are probably small.
G e n e r a l D i s c u s s i o n
The equation of B-D relationship (Eq. (4')) gives two useful methods for the studies of physiological plant ecology.
The first is for the studies of dry-matter economy of plants. In the investigations of dry-matter accumulation and consumption in a plant

344 Y. YOKOI AND A. KISHIDA
organ, it is a useful approach to regard each organ as a container of dry matter and to trace the change of the content of dry matter per unit size. As an index of the container size, fresh volume minus air space ( = actual fresh volume) is most reasonable. One of the best methods of estimating the actual fresh volume is the air-comparison pycnometer method, because the volume estimated by this method excludes open air spaces, and closed air spaces in fresh plant materials are usually negligible.
Fresh weight is a less reasonable index of the container size. Kuriyama (1978) observed that the relationship between the volume and fresh weight of the bulb of Licoris sanguinea in autumn differed among plants with and without flower and/or seeds, and suggested that the difference of dry-matter accumulation in bulbs affected this relationship. Her observation and suggestion agree with the theoretical result described by Eq. (1).
Therefore, the bulk density measured by an air-comparison pycnometer (actual bulk density) is a good index of dry-matter accumulation in a plant organ. Also, delicate care is necessary in using dry-matter content as an index of it, though this index is useful in selniquantitative investigations of dry-matter economy of plants (of. Fig. 3).
The air-comparison pycnometers, however, are fairly expensive, and, when sam- ples are small, the accuracy of volume measurements using them is not satisfactory. On the other hand, we can easily measure the weights of plant material with sufficiently high accuracy even in fields. Therefore, it is desirable to estimate the actual bulk density of a plant organ without the volume measurement.
When values of ~/and pa of an organ are given, we can estimate the actual bulk density with Eq. (4'). The drying experiment showed that when the desiccation of a plant material was not intense, Eq. (4') described well the relationships between bulk density and dry-matter content with $ = 0. In A. japonica, Pd of the 0- and 1-year-old leaves were 1.35 to 1.40, and those of 2-year-old leaves and all stems were about 1.45 (Table 1). Perhaps tile similar values may be applicable also to other species.
The second usefulness of the equation of B-D relationship is that we can estimate the amount of air space in a plant organ from the equation.
Air space in a plant organ has a significant effect on some physiological processes, especially on photosynthesis and transpiration. So, quantitative estimation of the air space is required in some investigations of physiological plant ecology. When vol- ume, fresh weight, dry weight, and density of dry matter of a plant material are given, we can easily estimate the volume of the air space with Eq. (5) (of. Fig. 8).
This study was partly supported by a Grant-in-Aid for Scientific Research (No. 57540369) from the Ministry of Education, Science and Culture, Japan.
R e f e r e n c e s
EVANS, G.C. 1972. The Quantitative Analysis of Plant Growth. Blaekwell Sci. Publ., Oxford. FIALA,K. 1973. Growth and production of underground organs of Typha angustifolia L. and
Phragmites communis Trin. Pol. Arch. Hydrobiol. 20: 59-66.

Bulk Density and Dry-Matter Content 345
INO, Y. 1977�9 Studies on the seasonal growth and the life mode of Kudzu vine plant (Pueraria Thunbergiana) (2). Sci. Res. (School of Educ., Waseda Univ.) 26 : 1-11 (in Japanese).
IWAKI, H. AND B. MIDORIKAWA. 1968. Principles for estimating root production in herbaceous perennials. In : Int. Symp. USSR : Methods of Productivity Studies in Root Systems and Rhizosphere Organisms p. 72-78. Nauka, Leningrad�9
KIMURA, M. 1976. Methods of Production Measurements in Terrestrial Plant Communities. Kyoritsu-shuppan, Tokyo (in Japanese)�9 �9 1978. Analysis of production processes of some evergreen plant populations based on the harvest method. In : M. Monsi and T. Saeki, ed., Ecophysiology of Photosynthetic Productivity, J IBP Synthesis 19 : 212-222. Univ. of Tokyo Press, Tokyo�9
- - , J. SUZUKI, M. TACHIBANA AND H. TAHARA. 1983. Role of evergreen foliage in the nitrogen economy during shoot growth of Ternstroemia qymnanthera, a warm-temperate broadleaf tree. Bot. Mag. Tokyo 96 : 75-83.
KOBAYASHI, K. 1981. Studies on the growth and maintenance of patch of Miscanthus sinensis Anderss. --Formation process of the patch and its ecological characteristics in a warm region--�9 J. Jap. Grassl. Sci. 27:71-78 (in Japanese).
KURIYAMA, K. 1978. Distribution and ecology of Licoris sanguinea Maxim. in Niigata Prefec- ture, Japan. M.S. thesis, Niigata Univ. (in Japanese)�9
MASUZAWA, W. 1977. C)kologishe Beobachtungen zur standSrtlichen Variabilit~t der Volums- dichte (bulk density) yon Speicherrhizomen yon Geum montanum L. im alpinen Grasheidgnrtel der Hohen Tauern. In : Alpine Grasheide Hohe Tauern, Ergebniss der ()kosystemstudie 1976 p. 103-110. Universitatsverlag Wagner, Innsbruck. AND K. HOGETSU. 1977. Seasonal changes in the amount of carbohydrate and crude
protein in the rhizome of Miscanthus sacchariflorus. Bot. Mat. Tokyo 90 : 181-191. MIDORIKAWA, B., K. HOGETSU, H. IWAKI AND M. MONSI. 1963. Studies on the Productivity and
Nutrient Circulation in Kirigamine Grassland, Central Japan. Naganoken Noseibu, Nagano (in Japanese)�9
MUTOH, N., K.H. YOSHIDA, Y. YOKOI, M. KIMURA AND K. HOGETSU. 1968. Studies on the production processes and net production of Miscanthus sacchariflorus community. Jap. J. Bot. 2 0 : 6 7 92.
VAN ANDEL, J. 1975. A study on the population dynamics of the perennial plant species Chamaenerion angustifolium (L.) Scop. Oecologia 19: 329-337.
YAMAMURA, Y. 1984. Matter production processes of Reineckia carnea Kunth, an evergreen forest floor herb in the warm-temperate region of Japan. Bot. Mag. Tokyo 97 : 179 191.
Received August 16, 1984