on: 20 october 2014, at: 19:49 biochemical and genomic ... · informa ltd registered in england and...
TRANSCRIPT

See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/274265215
Fulvic Acid (FA) for Enhanced Nutrient Uptake and Growth:
Insights from Biochemical and Genomic Studies
Article in Journal of Crop Improvement · November 2014
DOI: 10.1080/15427528.2014.923084
CITATIONS
3
READS
180
6 authors, including:
Some of the authors of this publication are also working on these related projects:
National facility for stable isotope studies in biological sciences View project
Development of alternate biofuel crops View project
Priya Vadlamudi
ITC Limited
1 PUBLICATION 3 CITATIONS
SEE PROFILE
Gurumurthy D. S.
ITC Limited
10 PUBLICATIONS 865 CITATIONS
SEE PROFILE
Bindumadhava H
World Vegetable Center, South Asia
36 PUBLICATIONS 433 CITATIONS
SEE PROFILE
Navin Sharma
The Trans-disciplinary University
41 PUBLICATIONS 123 CITATIONS
SEE PROFILE
All content following this page was uploaded by Bindumadhava H on 10 April 2015.
The user has requested enhancement of the downloaded file.

This article was downloaded by: [Bindumadhava H]On: 20 October 2014, At: 19:49Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Journal of Crop ImprovementPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/wcim20
Fulvic Acid (FA) for Enhanced NutrientUptake and Growth: Insights fromBiochemical and Genomic StudiesPriya B. N. V.a, Mahavishnan K.a, Gurumurthy D. S.a, BindumadhavaH.a, Ambika P. Upadhyaya & Navin K. Sharmaa
a ITC – Life Sciences & Technology Centre, ITC Limited, Bangalore,IndiaPublished online: 15 Oct 2014.
To cite this article: Priya B. N. V., Mahavishnan K., Gurumurthy D. S., Bindumadhava H., AmbikaP. Upadhyay & Navin K. Sharma (2014) Fulvic Acid (FA) for Enhanced Nutrient Uptake and Growth:Insights from Biochemical and Genomic Studies, Journal of Crop Improvement, 28:6, 740-757, DOI:10.1080/15427528.2014.923084
To link to this article: http://dx.doi.org/10.1080/15427528.2014.923084
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever orhowsoever caused arising directly or indirectly in connection with, in relation to or arisingout of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Journal of Crop Improvement, 28:740–757, 2014Copyright © Taylor & Francis Group, LLCISSN: 1542-7528 print/1542-7536 onlineDOI: 10.1080/15427528.2014.923084
Fulvic Acid (FA) for Enhanced Nutrient Uptakeand Growth: Insights from Biochemical and
Genomic Studies
PRIYA B. N. V., MAHAVISHNAN K., GURUMURTHY D. S.,BINDUMADHAVA H., AMBIKA P. UPADHYAY,
and NAVIN K. SHARMAITC – Life Sciences & Technology Centre, ITC Limited, Bangalore, India
Potassium (K), one of the essential elements required for plantgrowth and development, determines leaf quality in tobacco(Nicotiana tabacum L.). Potassium (K) levels are relatively high inblack soils (vertisols), but K uptake is severely hindered by the pres-ence of remarkably high levels of calcium and magnesium. Ourmajor objective was to enhance potassium uptake in black soils,which cover the major tobacco growing regions of Andhra Pradesh,India. Among several agronomic inputs such as soil amendments,fertilizer application, and plant growth regulators, we found thatfoliar application of fulvic acid (FA), one of the most bioactivehumate molecules, enhanced K levels in leaves. Using next-gen-eration sequencing (NGS), we identified changes in expressionlevels of a number of genes related to metabolic pathways impli-cated in plant growth and nutrient uptake upon FA application.Interestingly, starch levels in leaves were reduced concomitant withan increase in K attributable to FA application. We attempt toprovide plausible reasons for these observed FA-induced changes.Our results suggested that FA acts in a manner similar to theplant hormone auxin in tobacco, influencing expression of keygenes encoding transporters and enzymes involved in K uptake and
Received 26 February 2014; accepted 7 May 2014.Current affiliation for Navin K. Sharma: The World Agroforestry Centre, NASC complex,
PUSA, New Delhi, India.Address correspondence to Bindumadhava H., AVRDC – The World Vegetable Center,
South Asia Regional Office, ICRISAT campus, Patancheru 502 324, Hyderabad, AndhraPradesh, India. E-mail: [email protected]
Color versions of one or more of the figures in the article can be found online at www.tandfonline.com/wcim.
740
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

Fulvic Acid and Nutrient Uptake 741
starch metabolism. While fulvic acid has beneficial effects on plantgrowth, its mechanism of action is still unclear.
KEYWORDS FCV tobacco, humus, NGS, potassium, starch
ABBREVIATIONS. (C and N) carbon and nitrogen; (FCV) flue-cured Virginia;(FA) fulvic acid; (HA) humic acid; (NAA) naphthylacetic acid; (NGS) next-generation sequencing; (PCR) polymerase chain reaction; (K) potassium;(QC) quality control; (SBCS) southern black cotton soil.
INTRODUCTION
Potassium is the lone essential plant nutrient that is not a constituent ofany plant part (Marschner 1995; Cakmak 2005). Potassium acts as a cat-alyst for many of the necessary enzymatic processes in the plant and isalso involved in osmoregulation, i.e., regulation of water transport in thexylem and opening and closing of the stomata (Raschke 1975). Potassiumexists in several forms in the soil, including mineral K (90% to 98% of total),non-exchangeable K, exchangeable K, and dissolved or solution K (K ions)(Tisdale et al. 1985). Even though K is abundant in many soils, the bulk ofit is in the form unavailable to plants. Potassium uptake by plants is alsoinfluenced by type of soil. Calcareous soils tend to have high concentra-tions of calcium ions (Ca) that dominate clay surfaces and limit K sorption.High concentrations of Ca and magnesium (Mg) tend to limit K uptake bycompeting for binding sites on root surfaces (Havlinet al.1999). Enhancingpotassium content in plants is key to improving yield and quality in severalcrops (Geraldson 1985; Kanai et al. 2007; Lester et al. 2010). Potassium fertil-izers are either applied directly to the soil or in the form of foliar spray. Foliarapplication has been consistently shown to improve fruit quality attributesin cucumber, mango, and muskmelon; soil application had little or no effect(Tisdale et al. 1985; Brady and Weil 1999). Potassium enhanced color andglossiness of chili fruits (Prabhavathi et al. 2008). Earlier studies have shownthat foliar application of potassium fertilizers, humic acids, and their deriva-tives had improved K uptake in several crops (Wuzhong 2002; Demiral andKoseoglu 2005; Lester et al. 2005, 2006; Jifon and Lester 2009).
Many beneficial effects are attributed to foliar application of fulvic acid(FA), including stimulation of plant metabolism, increased enzyme activity(transaminase, invertase), increased bioavailability and uptake of nutrients(Pascual et al. 1999), and increased crop growth and yield (Mylonas and McCants 1980; Xudan 1986). Fulvic acid has maximum influence on chemicalreactions because of the presence of more electronegative oxygen atomsthan any other humate molecules, which enhances membrane permeability(Pascual et al. 1999).
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

742 P. B. N. V. et al.
Potassium is considered one among organic and inorganic compoundsthat influence the quality of tobacco; potassium is considered the elementof quality (Srinivas and Seshaiah 1993). Cured leaf color, grade, body, tex-ture, fire-holding capacity, and aroma are significantly influenced by Kconcentration (Krishnamurthy and Ramakrishnayya 1997; Juan et al. 2005;Mahavishnan, Priya, and Upadhyay, personal communication, July 2011).In India, flue-cured Virginia (FCV) tobacco is grown mainly in the states ofAndhra Pradesh (southern black cotton soils [SBCS]) of Prakasam district)and Karnataka.
In a previous study, we established positive effect of FA on carbontranslocation (Priya et al. 2011). In the same study, we also noted an increasein K content of leaves. K-deficient plants had several-fold increase in sucroseconcentration and marked reduction in root growth compared with controlplants (Cakmak et al. 1994; Huber 1984; Marschner et al. 1996). The accu-mulation of sugars and apparent increase in partitioning of fixed carboninto starch was associated with declined sucrose phosphate synthase activ-ity attributable to nutrient deficiency (Rufty and Huber 1983). Hence, alongwith K uptake, we determined the starch content in leaf and explainedits association with leaf K. In the present study, we explore FA-associatedphysiological, biochemical, and molecular changes that possibly enhanceK uptake and decrease starch content in FCV tobacco grown in the SBCSregion.
MATERIALS AND METHODS
Experimental Design
The SBCS soils are clay-to-clay loams throughout the profile, slightly alkalinein reaction (pH 7.5 to 8.8). The experiments were laid out in a random-ized complete-block design (RCBD) for first growing season (2007–2008).However growing seasons of 2008–09 and 2009–10 were on-farm large-scaletrials. Sixty-day-old seedlings of variety CY 135 were planted during winter(Oct–Nov) with a standard spacing of 65 cm x 65 cm. Recommended fertiliz-ers dose of 50:50:50 (N:P2O5:K2O) were applied 20 days before transplanting.Need-based prophylactic measures were taken to control pest and diseases.Commercial grade FA (81% pure: MW-308.24) was obtained from East CoastSeaweed Inc., India, and a concentration of 2mM was applied twice, i.e.,45 and 60 days after planting (DAP). The first priming was done at 60 DAPand subsequent primings were carried out at 7- to 10-day intervals. Leavesfrom each harvest were cured, stripped, conditioned, and graded separately.Analysis of variance (ANOVA) of phenotypic data was carried out using thegeneral linear model (GLM) of SAS software (version 9.3, 2011) (http://www.sas.com).
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

Fulvic Acid and Nutrient Uptake 743
Sampling
Cured leaf samples were drawn based on plant positions from the bottomof the plant (P, X, L, and T; see Figure S-1) as per the recommendation ofCentral Tobacco Research Institute, India (CTRI). Priming (P) contains leaves1–3 from the lower-most position, cutter (X) has leaves 4–9, leaf (L) positionhas leaves from 10-15, and final top (T) position consists of leaves from16–21 (Figure S-1). Laminas of the cured leaves were separated and groundto a fine powder. Leaves from all the positions were analyzed for K andstarch content. Green leaf samples from only the “X” position were collected7 days after first foliar application, snap-frozen in liquid nitrogen and usedfor RNA extraction.
Potassium and Starch Analysis
K content of the leaf samples was analyzed using Atomic AbsorptionSpectrophotometer (AAS) (Shimadzu, AA6300) in a measured quantity(100mg) of sample wet-digested with Di-acid mix (nitric acid: perchloric acid,9:4 ratio) (Jackson 1967). Estimation of starch was done as per McCreadyet al. (1950) with slight modifications. Leaf tissue (200mg) was extractedthrice with 10ml of sodium chloride-saturated methanol, followed by diges-tion with 5ml of 6M perchloric acid for 10 min. Contents were filtered(through Whatman filter paper grade 1) and filtrate was made up to 100mlwith distilled water. Ten ml of sample was mixed with 2.5ml iodine reagentand final volume made up to 25ml with distilled water. Absorbance of sam-ple was recorded at 600 nm using spectrophotometer (Agilent, 8453). Starchestimation was done using a standard curve.
Potassium Uptake Assay
Two month-old tobacco seedlings were placed in Hoagland solution withand without FA (2mM). Experiment consisted of 3 sets, i.e., absolute control(100ml Hoagland solution), FA-Solution (0.0625g FA in 100ml of Hoaglandsolution), and FA-Spray (100 ml Hoagland solution in a jar + foliar sprayof FA). Fulvic acid solution for foliar application was prepared by dissolv-ing 0.0625g FA in 100ml water, and then 10ml of solution was sprayed oneach seedling. Samples (left over Hoagland solution) were taken at 72h andanalyzed for K using AAS (Shimadzu, AA6300).
Quantitative Real-Time PCR (qRT-PCR) Analysis
Tobacco plants grown in greenhouse were sprayed with FA (2mM) 45 daysafter transplanting. Green leaf samples (X position) were collected after6, 7, 8, 12, 18, and 24 h of foliar application of FA. Total RNA was
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

744 P. B. N. V. et al.
isolated from tobacco leaf samples using RNA aqueous Kit (Ambion).Synthesis of cDNA was carried out with an equal concentration of RNA foreach sample using high-capacity cDNA synthesis kit (Applied Biosystems)after DNase treatment. Real-time PCR for AGPase small subunit (F-5′TAGCTGAGTGGAGCAGAGCA 3′and R- 5′ AGAAACAGCCTTAGGCGACA3′) and 18s rRNA (F- 5′ CGCGCTACACTGATGTATTC 3′ and R- 5′ GTACAAAGGGCAGGGACGTA 3′) was carried out using Power SYBR Green MasterMix (Applied Biosystems). The PCR was carried out with 7500 RT PCR sys-tem (Applied Biosystems) with following steps: initial denaturation at 94◦Cfor 2 min, followed by 35 cycles of denaturation at 94◦C for 15 s, annealingat 60◦C, and extension at 72◦C for 15 s. The experiment was repeated at leastthrice, along with three independent biological replicates.
Next-Generation Sequencing (NGS) Analysis
The total RNA was extracted using TRIzol (Invitrogen) and the quality of RNAchecked by Bioanalyzer (Agilent 2100). Transcriptome libraries (single librarywith average size of 250bp) for sequencing were constructed according tothe Illumina protocol outlined in “mRNA Sequencing Sample PreparationGuide” (Cat #RS-931-1001, Rev. D). Next generation sequencing was carriedout at Genotypic Pvt Ltd, India, using Illumina Genome Analyzer IIx via syn-thesis method with the read length of 72 bases single-end sequencing. Rawreads were filtered using SeqQC_V2.1, a genotypic proprietary tool (http://www.genotypictech.com/SeqQC.html?mnu=1), for quality control (Examplenumber of reads, bases, sequence length, nucleotide composition, andadapter sequence search, etc.).
Transcriptome alignment was carried out using Bowtie V0.12.7, andpost-alignment was done by SAM 0.1.7. The aligned contigs were annotatedby homology sequence search with The Arabidopsis Information Resources(TAIR) (www.arabidopsis.org) database. Expression value was calculatedusing custom Perl code, based on which fold change was calculated bycomparing the FA-treated expression values with control values. Transcriptswith a fold change of < −1 were regarded as down regulated, and the rangebetween −1 to +1 as neutral and > +2 as up regulated. Pathways used toillustrate changes in gene expression were obtained from KEGG Pathwaydatabase (http://www.genome.jp/kegg/pathway.html).
RESULTS AND DISCUSSION
Effect of FA Application on Leaf K and Starch Content
Foliar application of FA resulted in a significant increase in mean leaf K con-tent and a reduction in the mean leaf starch content at all the leaf positionsof the plant (Table 1, Figure S-1). This result was consistent across the three
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

Fulvic Acid and Nutrient Uptake 745
TABLE 1 Analysis of variance (ANOVA) using means across all leaf positions (P, X, L, and T)for leaf K content (%) and starch content (%) following application of fulvic acid in flue-curedVirginia tobacco for growing season 2007–08
Mean squares
Source of variation Df K content Starch content
Replications 2 0.01 0.05Treatments 5 0.54∗∗ 10.27∗∗
Residual 12 0.01 0.84CD (5%) 0.12 1.63
∗∗Denotes significance at the 1% probability level.
growing seasons (2007 to 2009), of which 2008 and 2009 were on-farm large-scale trials. Potassium content in these samples ranged from 0.5% to 3.6%across all three seasons with highest K content observed in P position leaf.Average K content of all the leaf positions was clearly higher in FA-treatedsamples than that in control leaves. The mean percent increase in K in FA-treated samples was 33% over the control (Figure 1A). Leaf starch contentvaried from 1.0% to 6.0% among FA-treated and control plants across allthree growing seasons, with lowest starch content in the P position leaf.The mean percent decrease in leaf starch content was 22% over the con-trol (Figure 1B). Among the other treatments explored in our earlier study(Mahavishnan et al. 2011), application of naphthylacetic acid (NAA) led to anincrease in K content by 40% and a decrease in starch content by 30% com-pared with the control. Though the humic substances (HS) have been shownto contain auxin and an “auxin-like” activity, support for this hypothesis isstill fragmentary (Trevisan et al. 2010).
Numerous studies have shown that HS, not particularly FA, enhanceroot, leaf, and shoot growth by stimulating water and nutrient uptake(Piccolo et al. 1993; Trevisan et al. 2009, and references therein). These pos-itive effects are explained as an interaction between HS and physiologicaland metabolic processes (Nardi et al. 2002; Muscolo et al. 1999). The addi-tion of HS stimulates nutrient uptake, cell permeability, and seems to regulatemechanisms involved in plant growth stimulation (Canellas et al. 2008).
However, it is not easy to distinguish between the direct and indirecteffects of HS as some of the positive effects may be ascribed to a generalimprovement of soil fertility, leading to a higher nutrient availability for plants(Quaggiotti et al. 2004). While in other cases, HS seem to positively influencemetabolic and signaling pathways involved in the plant development byacting directly on specific physiological targets (Trevisan et al. 2009; FigureS-2). It is interesting to note that, in our experiments, only foliar spray of FAbut not soil application was found to be effective (Mahavishnan, Priya, andUpadhyay, personal communication, July 2011).
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

746 P. B. N. V. et al.
FIGURE 1 Effect of fulvic acid application on mean potassium (A) and starch (B) contentof flue-cured Virginia tobacco in different growing seasons (2007–2009). Error bars representstandard deviation of the replicates (n = 24). The alphabet represents the level of significance(same alphabet indicates non-significance and different alphabet indicates significance oftreatment means for that particular growing season).
Relationship Between Leaf K And Starch Content
Foliar application of FA was found to enhance K and decrease starch con-tent of the cured leaf at all leaf positions. We observed a strong inverserelationship between K and starch in tobacco-cured leaf. Wherever K contentwas higher than 2%, starch content was lower than 2% (r2 = 0.31, Figure 2).Although the correlation coefficient was highly significant, it explained only31% of the variation. This result was highly consistent across the three
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

Fulvic Acid and Nutrient Uptake 747
FIGURE 2 Relationship between leaf potassium and starch content in flue-cured Virginiatobacco (values represent the data obtained from control and fulvic acid treated samples ofthree different growing seasons, 2007–2009). Values were significant at P = 0.001 (∗∗∗).
seasons. Elevated starch content in tobacco has a negative effect on itsquality. Potassium is a highly mobile element that helps in maintainingcell turgor (Krishnamurthy et al. 2001) and general plant growth (Taiz andZieger 2010). It is envisaged that potassium-induced cell expansion ener-gizes normal metabolism mainly associated with carbon and nitrogen. Thisactivation leads to production of more carbohydrate (mainly sucrose) andits translocation to growing parts, leaving behind less substrate for starchsynthesis in chloroplasts (Priya et al. 2011).
Similar results have been shown in cotton where low leaf K content wasassociated with elevated leaf carbohydrate concentrations because of delayin translocation (Pettigrew 1999) and high amounts of sugars under low Kcondition in all parts of peanut (Mahaboob and Rao 1980). According to Ruftyand Huber (1983), limitation in translocation of sugars from source leavesleads to enhanced partitioning of accumulated photosynthates into starch.Decreased sink capacity has been shown to inhibit sugar transport, enhanceaccumulation of sugars in leaves with concurrent increase in expression ofgenes involved in carbohydrate storage, and suppress photosynthetic genesand subsequent growth (Sheen 1990; Paul and Pellny 2003; Stitt et al. 2010).Further, reportedly K plays a major role in the regulation of the carbohydratemetabolism through control of starch-sugar balance and starch content ishigher under K starvation (White 1936).
Effect of FA Application on K Uptake
Hydroponic uptake assay revealed that the untreated control plants absorbedaround 0.5 mM of K only as against FA-treated plants (1 mM). Fulvic acidapplied either in solution or spray form increased the K uptake by 95% and
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

748 P. B. N. V. et al.
FIGURE 3 Effect of fulvic acid on potassium uptake in seedlings. Data indicates that treatedplants absorbed more K (∼twofold) from nutrient medium than control plants. Values aremeans with SD (minimum three replicates). The alphabet represents the level of significance(same alphabet indicates non significance and different alphabet indicates the significance oftreatment means).
92% over untreated control (Figure 3). Fulvic acid application on tobaccoseedlings grown in a controlled environment caused in leaves about a 16%increase in K content (Mylonas and McCants 1980). Rauthan and Schnitzer(1981) reported that FA application led to a significant increase in K con-centration in shoots of cucumber seedlings grown in hydroponics. SimilarHS-derived response of increased uptake of macro and micro-elements wasreported by several workers in other crop species (Chen and Aviad 1990;Varanini and Pinton 1995; Nardi et al. 2002; Yaofu 2005; Asik et al. 2009).Further, HS-stimulated uptake of nutrients and its involvement in regula-tion of plant growth in maize was demonstrated (Canellas et al. 2008).These studies suggested an increased ion influx was partially attributableto transcriptional activation of major gene H+-ATPase, which possibly leadsto generation of favorable electrochemical gradient (Millerand Smith 1996).Spraying humic acid has been shown to have a significant effect on physio-logical traits, including nutrient uptake, improved plant growth, and yield ofeggplant (Ebrahim et al. 2012).
Quantitative Real-Time PCR (qRT-PCR) Analysis for StarchMetabolism Gene
Expression of ADP-glucose pyrophosphorylase (AGPase) small subunit wasassayed by q-RT PCR in RNA isolated from the control and FA-treated tobaccoleaves collected at different time intervals post-FA application. The RNA fromleaves treated with NAA was used as a positive control in the experiment.Application of FA and NAA resulted in ∼threefold reduction in AGPase
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

Fulvic Acid and Nutrient Uptake 749
FIGURE 4 Gene expression profile of ADP glucose pyrophosphorylase post fulvic acid andnaphthylacetic acid application (A), and time course of gene expression profile of ADP glu-cose pyrophosphorylase (AGPase) in fulvic acid treated samples (B). Experiment was repeatedthree times and similar pattern of down-regulation was observed.
transcript level (Figure 4A). A time course study found that the AGPase tran-script levels were lowest at 12h post-application of FA (Figure 4B) and slowlyrecovered by 48h. Down-regulation of AGPase, a rate-limiting enzyme instarch biosynthesis (Taiz and Zieger 2010), reportedly reduces starch content(Kwak et al. 2007; Sanjaya et al. 2011). Similar results were obtained by appli-cation of NAA on small subunit of AGPase in tobacco cell lines (Miyazawaet al. 1999). A decrease in AGPase transcript level was observed after 6h; amaximum level was achieved at 12h and the level then slowly came downto normal after 48h of application of 2, 4 dichlorophenoxyacetic acid, asynthetic auxin (Miyazawa et al. 2002). The FA, which has an auxin-like
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

750 P. B. N. V. et al.
activity, could have induced similar changes in AGPase gene expression andstarch levels.
Global Gene Expression Changes Upon FA Application with SpecificEmphasis on Potassium and Starch Metabolism
Transcriptome sequencing was carried out to elucidate the global geneexpression changes in FA- and NAA-treated leaf samples. High through-put RNA sequencing with Illumina HiSeq2000 generated 20 and 40 millionsingle-end reads for the control and treated samples, respectively, with morethan 90% being high-quality reads. These reads were assembled and thenaligned against contigs assembled from tobacco transcriptome data (D. S.Gurumurthy, personal comm. 2013) to obtain information on expression lev-els for specific transcripts. Contigs that showed differential expressions wereannotated against TAIR database (www.arabidopsis.org).
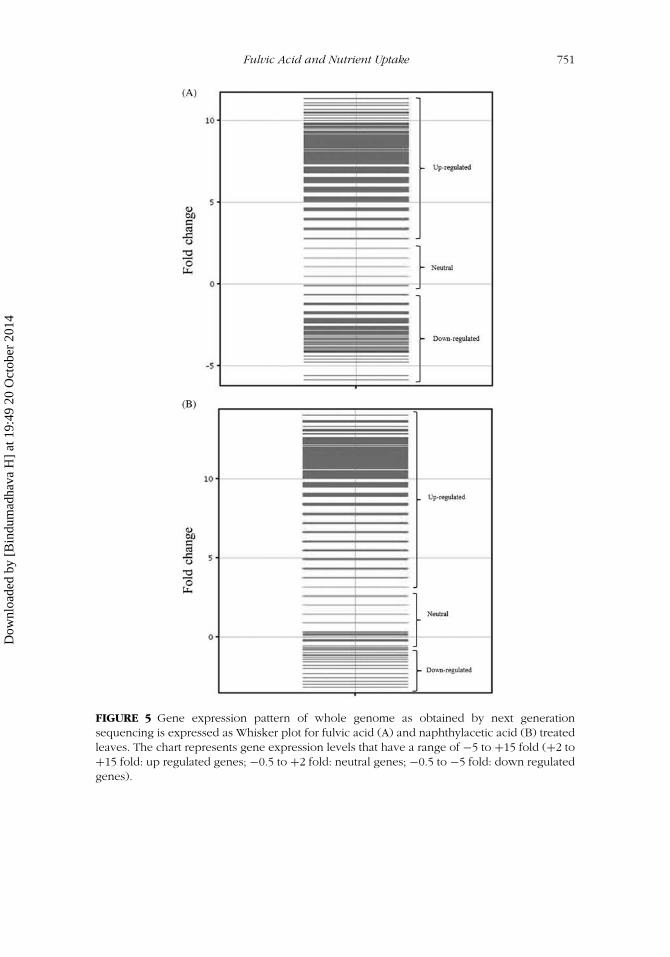
Transcriptome sequence of the treated plants revealed significant differ-ences in gene expression pattern relative to the control. Gene expressionchanges induced by FA application were similar to those induced by NAA,but the fold-change was much higher with NAA treatment. A few geneswere down regulated 7-fold and many others were up regulated 12-fold(Figure 5A and 5B). Expression of 5177 transcripts affected by foliar appli-cation of FA was annotated and categorized based on their GO-ontologyfunction (Figure 6A-D). Many auxin-related pathways were up regulated,including photosynthesis (Calvin cycle, light harvesting complexes), gly-colysis (pyruvate kinase), cell division (cyclins, cyclin-dependent kinases,mitogen-activated protein kinases [MAPK]), nitrogen fixation (nitrate trans-porters, nitrate reductase), starch (amylases, water dikinase, glucosidases),transcription factors, protein synthesis (translation initiation factors, ribo-somes), and translocation of sugars (plastidic glucose translocator [PGLCT],glucose phosphate translocator [GPT2]).
Annotated gene list was used to identify genes involved in K transport,sugar, and starch metabolism to deduce the effect of FA on expression ofenzymes/proteins involved in their respective pathways. Many transportersand channels involved in K uptake and mobilization were up regulated in FA-and NAA-treated samples (Table 2). Ashley et al. (2006) and Philippar et al.(1999) explained that auxin acted as a signal for low potassium and triggeredthe plant to express high affinity transporters/channels for its uptake. One ofthe reasons for enhanced K content in FA-treated leaves could be the auxin-like effect of FA. Auxin-like effect of humic substances is not uncommon, asreported by several workers (Nardi et al. 1994; Muscolo et al. 1998, 1999) onthe basis of analytical and biochemical assays. Our NGS data indicate thatmode of action of FA in facilitating K absorption and transport may be similarto that of auxin as it activates auxin-signaling pathway.
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014
Fulvic Acid and Nutrient Uptake 751
FIGURE 5 Gene expression pattern of whole genome as obtained by next generationsequencing is expressed as Whisker plot for fulvic acid (A) and naphthylacetic acid (B) treatedleaves. The chart represents gene expression levels that have a range of −5 to +15 fold (+2 to+15 fold: up regulated genes; −0.5 to +2 fold: neutral genes; −0.5 to −5 fold: down regulatedgenes).
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

752 P. B. N. V. et al.
FIGURE 6 Functional classification of transcripts (number indicated in parenthesis) of fulvicacid (A and B) and naphthylacetic acid (C and D) treated leaves. Most of the up-regulatedgenes are involved in photosynthesis and cell division followed by potassium transport andstarch degradation.
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

Fulvic Acid and Nutrient Uptake 753
TABLE 2 List of transporters/channels that get over-expressed because of fulvic acidapplication and their role in potassium mobilization (root to leaves)
S. No Transporters Function
1 KUP/AKT1/PIP Transporters involved in K uptake from root2 AKT 2 and 3 Phloem loading and transport of K to other parts3 CNGC Transport of K to other parts4 KCO/KAT Transporter involved in mesophyll movement (in leaves) of K ion
Global gene expression profile indicates that various enzymes involvedin degradation of large molecules (starch) to simple sugars were up regulatedupon FA or NAA application (Figures S-3a & b). Enzymes such as amy-lase, isoamylase, and glucan water dikinase, which catalyze initial reactionsof starch degradation (leads to glucans of different chain length), werehighly up regulated in FA-treated plants. The FA not only affects synthesis ofstarch but also activates enzymes that degrade stored starch (SupplementaryTable 1). Sugar metabolism in wheat seedlings implicates involvement of FAin up-regulation of starch-degrading enzymes (α-amylases) (Xu et al. 1998).Our results are in agreement with a similar report on application of humicacid in reducing starch synthesis in tobacco (Ye et al. 2009).
Thus, based on results obtained from biochemical and molecular stud-ies, we can conclude that foliar application of FA enhances K uptake andreduces starch in FCV tobacco of the SBCS region. Further, FA elicits almostsimilar gene expression changes as those caused by auxin. These findingsalso support the use of FA for crop species grown in soils where bioavail-ability and accessibility of nutrients is a constraint in achieving higher yields.
CONCLUSION
Foliar application of FA has a significant beneficial influence on general qual-ity of FCV tobacco of the SBCS region. Specifically, FA resulted in increasedleaf K and reduced starch contents. Molecular analysis revealed that FAactivated expression of high-affinity K transporters (enhanced K uptake),degraded stored starch, and energized plant metabolism (efflux of trios-PO4 for sucrose synthesis). We believe this is the first report on FCV tobaccogrown on black soils that provides encouraging and consistent results thatcomprehensively highlight inclusive benefits of FA both from agronomic andmolecular dimensions.
ACKNOWLEDGEMENTS
We thank Dr. C.C. Lakshmanan, Head, ITC-LSTC, ITC Ltd., Bangalore, India,for his continued support during the study. We appreciate Dr. M. Mani, Chief
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

754 P. B. N. V. et al.
Scientist, Research division, research & technical staff of ILTD, Rajahmundry,India, for supporting us to conduct field experiments and sample analysis.We also thank Dr. PRS Reddy, CTRI, Rajahmundry, India, for his crucial inputfor field experiment and Dr. Venkata Reddy (Agriscience) for useful sugges-tions during the preparation of the manuscript. Ms. Priya personally thanksSravanthi and Pavani for their help during starch analysis. Editorial and use-ful tips from the Corp R&D in-house manuscript committee are appreciated.Overall support from team Agriscience, LSTC, is gratefully acknowledged.
SUPPLEMENTAL MATERIAL
Supplemental data for this article can be accessed on the publisher’s website.
REFERENCES
Ashley, M. K., M. Grant, and A. Grabov. 2006. Plant responses to potassiumdeficiencies: A role for potassium transport proteins. J. Exp. Bot. 57:425–436.
Asik, B. B., M. A. Turan, H. Celik, and A. V. Katkat. 2009. Effects of humic substanceson plant growth and mineral nutrients uptake of wheat (Triticum durum cv.Salihli) under conditions of salinity. Asian J. Crop. Sci. 1:87–95.
Brady, N. C., and R. R. Weil. 1999. The nature and properties of soils, 9th ed. NewYork, NY: Macmillan.
Cakmak, I., C. Hengeler, and H. Marschner. 1994. Changes in phloem exportof sucrose in leaves in response to phosphorus, potassium and magnesiumdeficiency in bean plants. J. Exp. Bot. 45:1251–1257.
Cakmak, I. 2005. The role of potassium in alleviating detrimental effects of abioticstresses in plants. J. Plant Nutr. Soil Sci. 168:521–530.
Canellas, L. P., L. R. L. Teixeira Junior, L. B. Dobbss, C. A. Silva, L. O. Medici, D. B.Zandonadi, and A. R. Façanha. 2008. Humic acids cross interactions with rootand organic acids. Ann. Appl. Biol. 153:157–66.
Chen, Y., and T. Aviad. 1990. Effect of humic substances on plant growth. In Humicsubstances in soil and crop science, edited by P. McCarthy, C. E. Clapp, R. L.Malcolm, P. R. Bloom, 161–186. Madison, WI: American Society of Agronomy,
Demiral, M. A., and A. T. Koseoglu. 2005. Effect of potassium on yield, fruit quality,and chemical composition of greenhouse-grown Galia melon. J. Plant Nutr.28:93–100.
Ebrahim, A., K. M. Mohammad, M. Maral, R. Hamid, and Bozorgi. 2012. Effects ofbio, mineral nitrogen fertilizer management, under humic acid foliar spraying onfruit yield and several traits of eggplant (Solanum melongena L.). Afr. J. Agric.Res. 7:1104–1109.
Geraldson, C. M. 1985. Potassium nutrition of vegetable crops. In Potassium inagriculture, edited by R. S. Munson, 915–927. Madison, WI: ASA-CSSA-SSSA.
Havlin, J. L., J. D. Beaton, S. L. Tisdale, and W. L. Nelson. 1999. Soil fertility andfertilizers, 6th ed. Upper Saddle River, NJ: Prentice Hall.
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

Fulvic Acid and Nutrient Uptake 755
Huber, S. C. 1984. Biochemical basis for effects of K deficiency on assimilate exportrate and accumulation of soluble sugars in soybean leaves. Plant Physiol.76:424–430.
Jackson, M. L. 1967. Soil and plant chemical analysis. New Delhi, India: PrenticeHall of India, Pvt. Ltd.
Jifon, J. L., and G. E. Lester. 2009. Foliar potassium fertilization improves fruit qualityof field-grown muskmelon on calcareous soils in south Texas. J. Sci. Food Agric.89:2452–2460.
Juan, L., Z. Mingqing, and L. Qiong. 2005. Effects of interaction of potassium, calciumand magnesium on flue-cured tobacco growth and nutrient absorption. J. AnhuiAgric. Univ. 32:529–533.
Kanai, S., K. Ohkura, J. J.Adu-Gyamfi, P. K. Mohapatra, N. T. Nguyen, H. Saneoka,and K. Fujita. 2007. Depression of sink activity precedes the inhibition ofbiomass production in tomato plants subjected to potassium deficiency stress.J. Exp. Bot. 58:2917–2928.
Krishnamurthy, V., and B. V. Ramakrishnayya. 1997. Potassium fertility of flue-curedtobacco growing soils of India. J. Potassium Res. 13:159–169.
Krishnamurthy, V., B. V. Ramakrishnayya, and K. Deo Singh. 2001. Potassium man-agement for improving yield and quality of flue-cured tobacco. J. Potassium Res.17:231–235.
Kwak, M. S., S. R. Min, S. M. Lee, K. N. Kim, J. R. Liu, K. H. Paek, J. S. Shin,and J. M. Bae. 2007. A sepal-expressed ADP-glucose pyrophosphorylase gene(NtAGP) is required for petal expansion growth in ‘Xanthi’ tobacco. PlantPhysiol. 145:277–289.
Lester, G. E., J. L. Jifon, and G. Rogers. 2005. Supple-mental foliar potassium appli-cations during muskmelon fruit development can improve fruit quality, ascorbicacid, and 13-carotene contents. J. Amer. Soc. Hort. Sci. 130: 649–653.
Lester, G. E., J. L. Jifon, and D. J. Makus. 2006. Supplemental foliar potassium appli-cations with or without a surfactant can enhance netted muskmelon quality.Hort Sci. 41:741–744.
Lester, G. E., J. L. Jifon, and J. M. Donald. 2010. Impact of potassium nutrition onfood quality of fruits and vegetables: A condensed and concise review of theliterature. Better Crops. 94:1.
Mahaboob Basha, S. K., and G. RajeswaraRao. 1980. Effect of potassium deficiencyon growth & metabolism of peanut (Arachis hypogaea L.) plants. Proc. IndianAcad. Sci. (Plant. Sci.) 89:415–420.
Mahavishnan, K., V. N. V. Priya, A. P. Upadhyay, and N. Sharma. 2011. Patent onFulvic Acid; an effective foliar nutrition for enhancing the quality and yield ofSBCS tobacco. Ref No. 1210/Kol/2011.
Marschner, H. 1995. Functions of mineral nutrients: macronutirents. In Mineral nutri-tion of higher plants, 2nd ed., edited by H. Marschner, 299–312. New York, NY:Academic Press.
Marschner, H., E. A. Kirkby, and I. Cakmak. 1996. Effect of mineral nutritional statuson shoot-root partitioning of photo assimilates and cycling of mineral nutrients.J. Exp. Bot. 47:1255–1263.
McCready, R. M., J. Guggolz, V. Silviera, and H. S. Owens. 1950. Determination ofstarch & amylase in vegetables. Anal. Chem. 22:1156–1158.
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

756 P. B. N. V. et al.
Miller, A. J., and S. J. Smith. 1996. Nitrate transport and compartmentation in cerealroot cells. J. Exp. Bot. 300:843–54.
Miyazawa, Y., A. Sakai, S. Miyagishima, H. Takano, S. Kawano, and T. Kuroiwa.1999. Auxin &cytokinin have opposite effects on amyloplast development &the expression of starch synthesis genes in cultured bright yellow-2 tobaccocells. Plant Physiol. 121:461–470.
Miyazawa, Y., N. Kutsuna, H. Kuroiwa, T. Kuroiwa, N. Inada, and S. Yoshida.2002. Dedifferentiation of starch-storing cultured tobacco cells: effects of 2, 4-dichlorophenoxy acetic acid on multiplication, starch content, organellar DNAcontent, & starch synthesis gene expression. Plant Cell Rep. 21:289–295.
Muscolo, A., S. Cutrupi, and S. Nardi. 1998. IAA detection in humic substances. SoilBiol. Biochem. 30:1199–1201.
Muscolo, A., F. Bovalob, F. Gionfriddob, and S. Nardi. 1999. Earthworm humic matterproduces auxin-like effects on Daucuscarota cell growth & nitrate metabolism.Soil Biol. Biochem. 31:1303–1311.
Mylonas, V. A., and C. B. McCants. 1980. Effects of humic & fulvic acids on growthof tobacco. 2. Tobacco growth & ion uptake. J. Plant Nutr. 2:377–393.
Nardi, S., M. R. Panuccio, M. R. Abenavoli, and A. Muscolo. 1994. Auxin-like effect ofhumic substances extracted from faeces of Allolobophoracaliginosa & A. Rosea.Soil Biol. Biochem. 26:1341–1346.
Nardi, S., D. Pizzeghello, A. Muscolo, and A. Vianello. 2002. Physiological effects ofhumic substances on higher plants. Soil Biol. Biochem. 34:1527–1536.
Pascual, J. A., C. Garcia, and T. Hernandez. 1999. Comparison of fresh and com-posted organic waste in their efficacy for the improvement of arid soil quality.Bioresour. Technol. 68:255–264.
Paul, M. J., and T. K. Pellny. 2003. Carbon metabolite feedback regulation of leafphotosynthesis and development. J. Exp. Bot. 54:539–547.
Pettigrew, W. T. 1999. Potassium deficiency increases specific leaf weights and leafglucose levels in field-grown cotton. Agron. J . 91:962–968.
Philippar, K., I. Fuchs, H. Luthen, S. Hoth, and C. S. Bauer. 1999. Auxin-inducedK channel expression represents an essential step in coleoptile growth andgravitropism. Proc. Natl. Acad. Sci. U.S.A. 96:186–191.
Piccolo, A., G. Celano, and G. Pietramellara. 1993. Effects of fractions of coal-derivedhumic substances on seed germination and growth of seedlings (Lactuga sativaand Lycopersicumesculentum). Biol. Fertil. Soil. 16:11–15.
Prabhavathi, K., B. I. Bidari, G. B. Shashidhara, and J. C. Mathad. 2008. Influence ofsources and levels of potassium on quality attributes and nutrient compositionof red chilies. Karnataka J. Agric. Sci. 21:379–381.
Priya, V. N. V., H. Bindumadhava, K. Mahavishnan, A. P. Upadhyay, and N. Sharma.2011. Effect of fulvic acid in improving carbon (14C) uptake and translocation intobacco. J. Plant Biol. 38:67–70.
Quaggiotti, S., B. Ruperti, D. Pizzeghello, O. Francioso, V. Tugnoli, and S. Nardi.2004. Effect of low molecular size humic substances on nitrate uptake andexpression of genes involved in nitrate transport in maize (Zea mays L.). J. Exp.Bot. 55:803–813.
Rauthan, B. S., and M. Schnitzer. 1981. Effect of soil fulvic acid on the growth andnutrient content of cucumber plants. Plant Soil. 63:491–495.
Raschke, K. 1975. Stomatal action. Annu Rev Plant Physiol. 26:309–340.
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014

Fulvic Acid and Nutrient Uptake 757
Rufty, T. W., and S. C. Huber. 1983. Changes in starch formation and activities ofsucrose phosphate synthase and cytoplasmic fructose-1, 6-bisphosphatase inresponse to source-sink alterations. Plant Physiol. 72:474–480.
SanjayaDurett, T. P., S. E. Weise, and C. Benning. 2011. Increasing the energy den-sity of vegetative tissues by diverting carbon from starch to oil biosynthesis intransgenic Arabidopsis. Plant Biotechnol. J . 9:874–883.
Sheen, J. 1990. Metabolic repression of transcription in higher plants. Plant Cell.2:1027–1038.
Srinivas, D., and B. V. Seshaiah. 1993. Quantity-intensity parameters of potassium intobacco growing soils of and Andhra Pradesh [India]. Tob. Res. 19:37–40.
Stitt, M., J. Lunn, and B. Usadel. 2010. Arabidopsis and primary photosyntheticmetabolism: More than the icing on the cake. Plant J . 61:1067–1091.
Taiz, L., and E. Zeiger. 2010. Plant physiology, 5th ed. Sunderland, MA: SinauerAssociates.
Tisdale, S. L., W. L. Nelson, and J. D. Beaton. 1985. Soil and fertilizer potassium. InSoil Fertility and Fertilizers, 4th ed., edited by S. L. Tisdale, W. L. Nelson, and J.D. Beaton, 249–291. New York, NY: Macmillan.
Trevisan, S., D. Pizzeghello, B. Ruperti, O. Francioso, A. Sassi, and K. Palme. 2009.Humic substances induce lateral root formation and expression of the earlyauxin responsive IAA19 gene and DR5 synthetic element in Arabidopsis. PlantBiol. 12:604–614.
Trevisan, S., O. Francioso, S. Quaggiotti, and S. Nardi. 2010. Humic substancesbiological activity at the plant-soil interface. From environmental aspects tomolecular factors. Plant Signal Behav. 5:635–643.
Varanini, Z., and R. Pinton. 1995. Humic substances and plant nutrition. Prog. Bot.56:97–117.
White, H. L. 1936. The interaction of factors in the growth of Lemna.7. The effect ofpotassium on growth and multiplication. Ann. Bot. 5:175–196.
Wuzhong, N. 2002. Yield and quality of fruits of solanaceous crops as affected bypotassium fertilization. Better Crops Inter. 16:1.
Xu, Y., J.H. Liu, A. Peng, and Z. J. Wang. 1998. Effect of selenium and fulvic acidon the sugar metabolism in wheat seedling. Acta Scientiae Circumstantiae.18:401–405.
Xudan, X. 1986. The effect of foliar application of fulvic acid on water use, nutrientuptake & yield in wheat. Aust. J. Agric. Res. 37:343–350.
Yaofu, W. 2005. Effect of irrigation and rate of humic acid on concentration ofnutrients and yield and quality of flue-cured tobacco. J. Anhui Agric. Sci.33:96–97.
Ye, X., A. Ling, B. Zhang, X. Liu, Y. Li, and G.Liu. 2009. Effect of humic acid fertilizeron soil properties and leaf qualities of tobacco. Acta Agriculturae Boreali Sinica.24:170–173.
Dow
nloa
ded
by [
Bin
dum
adha
va H
] at
19:
49 2
0 O
ctob
er 2
014
View publication statsView publication stats