of fibrils methods
TRANSCRIPT

TIHE LIPID CONTENT OF AMYLOID FIBRILSPURIFIED BY A VARIETY OF METHODS
IN CHANG KIM, M.D., TSURANOBU SHIRAHAMA, M.D., and ALAN S. COHEN, M.D.
From the Robert Dawson Evans Department of ClinicalResearch, University Hospital, and Boston University
School ot Medicine, Boston University Medical Center, Boston, Mass.
It is generally agreed that the major component of amyloid is a pro-teinaceous material whose precise chemical characterization has beendifficult owing to problems in isolation and solubilization. However, it isof historical interest to note that, prior to the introduction of the name"amyloid" by Virchow in I854, it was classified as "lardaceous" or"waxy"' degeneration, suggesting that lipid was a major component ofamyloid.2 Even today the question of the presence or absence of lipid inamyloid as a minor but essential component continues to be a contro-versial point. Certainly both histochemical and biochemical studies ofamyloid-laden tissues, as reported, have given varying results.
For example, there are numerous histochemical studies which havesuggested the presence of lipid in amyloid tissue.," In a recent report,McAlpine and Fuller6 described positive reactions to fat stains in lo-calized laryngeal amyloidosis. Sorenson, Heefner, and Kirkpatrick7noted that oil red 0-positive lipid granules were rather a constant findingin experimental murine amyloidosis induced by subcutaneous injectionsof casein. On the other hand, there are equally numerous reports ofnegative results for lipid staining of amyloid tissue."13 The results ofbiochemical studies present similar problems. Calkins and Cohen,"4Letterer, Gerok, and Schneider,16 and Battaglia and Pernis,16 all re-ported the presence of varying amounts of lipid in crude amyloid prep-arations, whereas Wagner 17 commented on its absence.
These conflicting reports on the presence of lipid in amyloid are largelydue to the fact that most of the analyses were carried out on whole tissuesor extracts of crude amyloid preparations and the stains on tissue sec-tions. However, the ultrastructure of amyloid has been well delineatedin the recent years, and the fibrillar nature of amyloid is now generallywell established.2'18 19 There also has been the development of improved
Preliminary results of these studies were presented at the American Federation for Ex-perimental Pathology in April x965.1
Supported by Grants AM-o4S99 and Ti AM-5285 from the National Institute of Ar-thritis and Metabolic Diseases, U. S. Public Health Service, and grants from the Massa-chusetts Chapter of the Arthritis Foundation and from the Arthritis Foundation.
Accepted for publication Dec. I, 1966.869

KIM, SHIRAHAMA, AND COHEN
physical as well as chemical methods of separating amyloid fibrils inincreasingly purer states.20'21 These advances in isolation and identifi-cation of amyloid fibrils now make it possible to perform more definitivebiochemical analyses on these fibrils, which (except for ground sub-stance) appear to constitute the major part of amyloid deposits.
This paper describes: (i) a new method of purification devised in ourcontinuous effort to isolate amyloid fibrils in increasingly purer states;(2) the results of lipid analysis of isolated amyloid fibrils of varyingpurity, using the techniques of thin layer chromatography (TLC) andgas-liquid chromatography (GLC); and (3) the results of lipid analysisof an amyloid fibril hydrolysate by a proteolytic enzyme.
MATERIALSFor this study the spleens from i case of primary and I case of secondary human
amyloidosis as well as a normal control spleen were used in preference to the liverbecause of the known higher lipid content of the liver in certain diseases. The patientwith secondary amyloidosis had had rheumatoid arthritis for over 40 years and hadhepatosplenomegaly and a nephrotic syndrome. At necropsy, the patient was foundto have extensive amyloidosis involving the liver, spleen, and kidneys. The estimatedreplacement of the gross spleen with amyloid was 75%. The patient with primaryamyloidosis had developed a nephrotic syndrome and gastrointestinal hemorrhagewithout other evidence of predisposing disease. Necropsy disclosed massive amy-loidosis involving the spleen, liver, kidneys, heart, adrenal glands, gastrointestinaltract, lungs, and bone marrow. The large, 6oo-gm. spleen was most affected and wasalmost completely replaced by amyloid. The amyloid from both of these cases waspositive to Congo red staining and showed crystal violet metachromasia. The Congored-stained amyloid demonstrated unequivocal green birefringence on polarizationmicroscopy. On electron-microscopic examination, the amyloid in both cases wasfound to consist of fine fibrils.
METHODSSeparation Procedures
For lipid determinations, a series of increasingly pure harvests of amyloid fibrilswere prepared using the scheme as outlined in Text-fig. i. The purpose was to testthe hypothesis that if lipid were a contaminant, it would not only decrease but woulddo so proportionately in various lipid fractions in these preparations and that lipidextraction would cause no ultrastructural alterations in the amyloid fibrils.Specimen A. Our earliest and crudest preparation consisted of an amyloid-rich
"top layer" which was obtained from homogenized whole spleen as previously re-ported.20Specimen B. A purer preparation was obtained by subjecting Specimen A to
sucrose density gradient centrifugation as recently reported.21Specimen C. For the final and purest specimen, Specimen B was further purified
in the following manner:i. A ioo-mg. portion of Specimen B was suspended in I5 ml. of o.I M phosphate
buffer (pH 7.4). After 3 drops of toluene had been added as a preservative, themixture was kept in an incubator at 370 C. for 3 days with occasional shaking.The incubated material was centrifuged in a Servall RC-2 refrigerated centrifuge atIo,ooo rpm (I2,100 X g) at o0 C. for i hr. The supernatant was discarded and thesediment was washed with 20 ml. of demineralized distilled water and centrifuged
870 Vol. 50, No. 5

LIPID IN AMYLOID FIBRILS
WHOLE AMYILom SPrrxn
I HomogenizationCentrifugation
SPECIMEN A ("Top Layer")
Sucrose densityI Gradient centrifugation
SPECIMEN B
I Chemical & Enzymatic treatment
SPECIMEN CTEXT-FIG. Z. Scheme of separating amyloid fibrils, to purify specimens and remove
contaminants.
at IO,OO0 rpm (I2,IOO X g) at o0 C. for I hr. The supematant was discarded andthe sediment was washed and centrifuged as above in demineralized distilled wateronce again.
2. The sediment was suspended in io ml. of o.i M phosphate buffer (pH 7.4) inwhich collagenase (Worthington Biochemical Corporation, Freehold, N.J.), I% ofthe weight of the substrate, was added. The mixture was then incubated in anEberbach shaking water bath at 370 C. for 48 hr., with the enzyme added every 24hr. At the end of the incubation the specimen was centrifuged at io,ooo rpm (I2,IOOX g) at o0 C. for i hr., and the supernatant was discarded. The sediment was washedwith 20 ml. of demineralized distilled water and centrifuged at Io,ooo rpm (12,IOOX g) for i hr. at o0 C. The washing was repeated.
3. Next, the sediment was suspended in ioo ml. of a io% NaCl solution and leftat room temperature for 2 days with occasional shaking. The resultant suspensionwas then centrifuged at io,ooO rpm (12,IOO x g) for i hr. at o0 C. The sedimentwas washed with 30 ml. of demineralized distilled water twice and centrifuged asabove.
4. Finally, the sediment was suspended in IOO ml. of 3% ammonium hydroxidesolution and left at room temperature for 48 hr. with occasional shaking. The samplewas centrifuged at Io,ooo rpm (I2,IOO X g) for io min. at o0 C. The supernatantwas carefully decanted into another centrifuge tube and the sediment was put aside.Then the supernatant was centrifuged at i8,ooo rpm (39,ooo X g) for 5 hr. at o° C.A pellet was obtained which was dialyzed against demineralized distilled water for72 hr. and the supernatant discarded. After dialysis, the residue was centrifuged ati8,000 rpm (39,000 X g) for i hr. at o0 C. The final sediment was lyophilized priorto lipid extraction.Only the primary amyloid spleen was subjected to all 3 procedures to obtain
Specimens A, B, and C. Purification of the secondary amyloid spleen was carried tothe second procedure and yielded Specimens A and B. In the case of the normal con-trol spleen, only Specimen A could be obtained.
Enzymatic Degradation of Amyloid Fibrils by a Proteolytic EnzymeTo test for the presence or absence of lipid between the interstices of, or within,
the amyloid fibrils, a preparation of fibrils was digested by pronase in the followingmanner.
Twenty-five milligrams (dry weight) of Specimen B (sucrose separated amyloidfibrils) of the primary amyloid spleen, from which lipid had been extracted (asbelow, Folch method) was suspended in 4 ml. of O.I M phosphate buffer (pH 7.4).
871May z967

KIM, SHIRAHAMA, AND COHEN
To the suspension o.5 mg. of pronase (Calbiochem., Los Angeles, Calif.) was addedevery 24 hr. The incubation was carried out for 6 days in a shaking water bath at370 C. Following this, the incubation mixture was centrifuged at io,ooo rpm (I2,100X g) for i hr. at o0 C. The supernatant was saved and the residue was washedwith 2 ml. of O.I M phosphate buffer (pH 7.4). The supernatants were pooled forlipid extraction.
Electron MicroscopyAliquots of the original frozen spleen specimens were sliced to i-cu.-mm. portions,
fixed in osmium tetroxide, and embedded in Epon using standard techniques.22'23The blocks were sectioned with an LKB microtome, stained with lead 24 and examinedin a Siemens Elmiskop I electron microscope.The lyophilized material obtained from Specimens A, B, and C was studied by
preparing suspensions on grids which were then shadowed with platinum-palladium.In addition, negative staining techniques were used. For the latter, an aliquot ofthe sediment was suspended in distilled water or 0.4% sucrose, applied to aParlodion-coated, carbon-coated grid and air dried. Two per cent phosphotungsticacid (pH 7.0) or I% uranyl acetate (pH s.o) was applied to the grid for a moment,according to the technique of Brenner and Horne.25 The excess was drained off,the specimen air-dried and examined in the electron microscope.
Lipid AnalysisLipid Extraction. Lipid in the samples was extracted using the method of Folch.28
Specimens A and B, 6o mg. each, and 40 mg. of Specimen C were treated with 20ml. of chloroform: methanol (2:I, V/v) mixture and the extracts were concentratedto o.4-o.5 ml. using oxygen-free nitrogen gas so that the final concentration of themajor components was in the range of i.o-o.I%. The lipid extract of the supernatantof the amyloid fibril hydrolysate obtained by pronase treatment was concentratedto o.i ml. to obtain the maximum concentration of lipid which might be present inthe supernatant.
Qualitative Lipid Analysis by Thin Layer Chromatography (T.L.C.). Qualitativelipid analyses of the extracts were performed by T.L.C., using the method ofMangold.27 Layers of Silica gel G (Brinkmann Co., Inc., Great Neck, Long Island,N.Y.) 250 u thick were coated on 20 X 20 cm. glass plates by the method of Stahlas quoted by Mangold.27 The plates were dried at room temperature and activatedat ixo° C. for i hr. before use. Aliquots of the concentrted extracts were spottedon the plates using a Hamilton microsyringe (Hamilton Company Inc., Whittier,Calif.) and the lipids were separated by ascending elution with a solvent mixture ofpetroleum ether: diethylether: glacial acetic acid (85:15 :1, v :v :v). When the solventfront reached the predeternined point, the plates were dried at room temperature,and the spots were visualized by spraying with So% sulfuric acid solution and heat-ing at I100 C. for 5 min. and then at 1700 C. for io min. longer. The unknown spotswere identified by simultaneously running the known standards of lecithin, cholesterolpalmitate, tripalmitin, oleic acid (Applied Science Lab., Inc., State College, Pa.) andcholesterol (Fisher Scientific Co., Medford, Mass.).
Quantitative Lipid Analysis by T.L.C. Quantitative determinations of freecholesterol, cholesterol esters, free fatty acids, and triglycerides separated werecarried out by T.L.C. using the graphic method of Purdy and Truter.28 This is basedon the principle that in T.L.C. the square root of the spot area is a linear function ofthe logarithm of the weight present. Since each analysis requires its own calibration,known standard samples of cholesterol, tripalmitin, oleic acid, and cholesterolpalmitate were always chromatographed side by side with the unknown on eachplate. After separation and visualization of the spots as stated above, a photostaticcopy of the plate chromatogram was made, using a Copease photocopier, and thespot area determined by tracing the spots on transparent graph paper. The weight of
872 Vol.go, No. f

LIPID IN AMYLOID FIBRILS
a fraction was determined from its area by reference to a weight area curve whichwas determined by plotting the areas of a series of spots containing known weights.All determinations were done in duplicate and the results were converted to milli-grams per hundred milligrams of lypholized sample. The accuracy and reproducibilityof this method were checked using 2 compounds with different Rf values-namely,cholesterol, which has a relatively low Rf value, and triglyceride, which migratesfarther to the solvent front. For this purpose 70 ,ug. of cholesterol and tripalmitinwere applied as 0.7% solutions in chloroform: methanol (2:1, v/v) and were mea-sured as the unknowns. Quantitative T.L.C. was found to be not suitable for phos-pholipids since the latter remained at the point of origin on a T.L.C. plate withresultant poor correlation between the spot area and the weight applied.
Therefore, phospholipid determination was performed indirectly by measuring thephosphorus content in the aliquot of the chloroform-methanol extracts by the methodof Bartlett.29 Phosphorus content was multiplied by a factor of 25 to calculate thequantity of phospholipid. All determinations were done in duplicate and the resultswere converted to milligrams per hundred milligrams of lyophilized samples.Long-Chain Fatty-Acid Analysis by Gas Liquid Chromatography (G.L.C.) Long-
chain fatty-acid (C14-20) analyses were done on separate aliquots of the Folch ex-tract. After evaporating the extract to dryness under a stream of oxygen-free nitrogen,the residue was methylated using the method of Metcalfe and Schmitz.30 The fattyacids were analyzed as methyl esters by gas-liquid chromatography using an F-MModel 400 apparatus with a hydrogen flame ionization detector. The column used wasglass (6 ft., o.s in. outside diameter) packed with5% DEGS (diethylene glycol suc-cinate) coated on 80/IOO mesh Diatoport S (obtained from F & M Scientific Corpo-ration, Avondale, Pa.) at a column temperature of I80° C. Helium was used as thecarrier gas at the flow rate of 6o ml./min. Fatty acids were identified by comparisonof their retention times with those of commercially available pure standards (Ap-plied Science Labs., Inc., State College, Pa.). The areas under peaks were determinedby multiplying peak height by width at half height, as recommended by the AdvisoryCommittee, Lipid Standard Program, The National Heart Institute.81
RESULTSLipid staining of frozen sections of the amyloid-laden spleens demon-
strated abundant oil red 0-positive granules indicating the presenceof lipids in the whole amyloid tissue (Fig.I). Electron micrographs ofamyloid fibrils by the technique of shadow casting showed that our
specimens did become significantly purer after each purification pro-cedure. In Specimen A (Fig. 2A), amyloid fibrils were abundant in theoriginal tissue, but there was a considerable amount of surface con-taminant and amorphous debris which obscured the details of fibrilstructure. In Specimen B (Fig. 2B) the contaminant had largely, but notcompletely, been eliminated. The long, thin, nonbranching character ofthe fibril was more apparent. Occasional twists in fibrils were seen aswell as apparent lateral aggregation. Finally, Specimen C (Fig. 2C)showed almost pure amyloid fibrils and virtually no contaminant. Herethe suggestion of beading or periodicity in the fibrils is strongly suggestedas the surface structure is better visualized. Lateral packing of fibrilsagain may be seen.The results of thin layer chromatographic fractionation of lipid using
873May zo67

KIM, SHIRAHAMA, AND COHEN
SAWPLE STANDARD# . . ^ Chol. Ester
F.A. Methyl Ester
* * * Triglyceride
* 9 FPatty acid
' a a* Chol.
* P-lipid
TEXT-FiG. 2. Standard thin-layer chromatographic fractionation of lipids in silica gel.
Silica gel G and the solvent system of petroleum ether: diethylether:acetic acid (85:15:I, v:v:v) are illustrated in Text-fig. 2. With thesystem employed, separation of lipid into its components-phospho-lipids, cholesterol, fatty acids, triglycerides, fatty acid methyl esters,and cholesterol esters-was always complete without overlap betweenadjacent spots. When the lipid extracts of the Specimens A, B, and C ofthe amyloid fibrils were examined by T.L.C., varying amounts of choles-terol, cholesterol esters, phospholipids, free fatty acids and triglycerideswere present in all the samples.The accuracy and reproducibility of quantitative T.L.C. by the
graphic method of Purdy and Truter 28 were checked, using cholesteroland triglycerides, compounds with different Rf values. When 70 uag. ofcholesterol was applied as an unknown and chromatographed, the aver-age weight was 66.o + 3.9 Ag. on I4 determinations. The comparable
TABLE IANALYSIS OF THE ACCURACY AND REPRODUCIBILITY OF THE QUANTITATIVE THIN LAYER
CHROMATOGRAPHY METHOD
Actual wt. Ave. wt. found No. of(Itg.) (Ag.) S. D. observations
Tripalmitin 70.0og 72.0 ,ug ±5.1 ,ug 14Cholesterol 70.0og 66.o pAg ±3.9 pg 14
874 Vo.. 5o, No. 5

LIPID IN AMYLOID FIBRILS
figure for tripalmitin was 72.0 + 5.I ug. (Table I). An example of weightarea curve, which was constructed by plotting the areas of a series ofspots containing known amounts of tripalmitin and cholesterol (as wasused in determining the standard deviation shown in Table I) is illus-trated in Text-fig. 3.Lipid composition of the amyloid fibrils of varying purity, as deter-
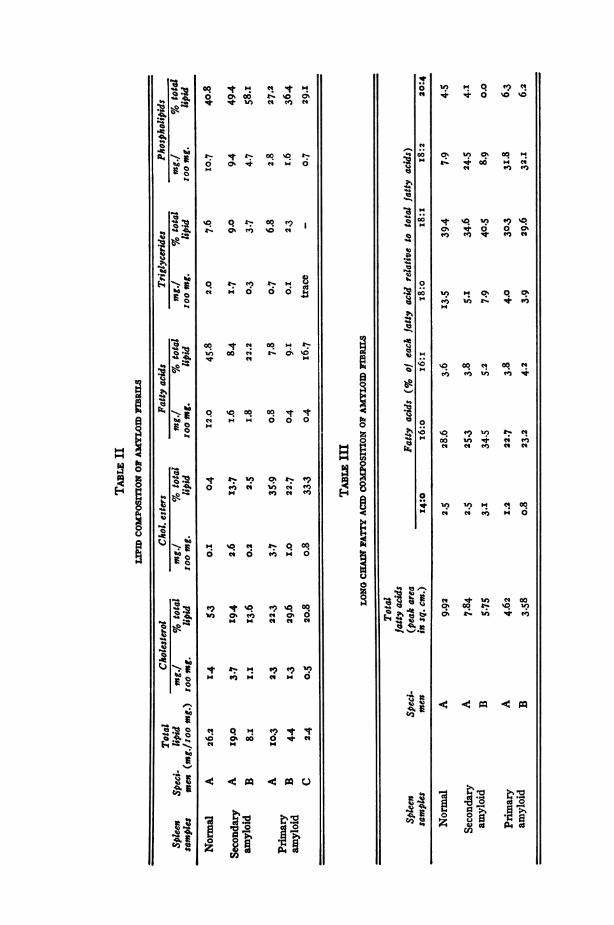
mined by qualitative T.L.C., exhibited a high cholesterol content(roughly 50% of the total lipids in the samples of the primary-amyloidspleen) and relative abundance of free fatty acids in all the samples incomparison to triglycerides in the comparable samples. As purificationproceeded from Specimen A to Specimen B, there was a decrease in thetotal lipid from I9.0 mg./ioo mg. of sample in the crudest preparation(Specimen A) to 8.i mg./ioo mg. of sample in the purer specimen(Specimen B) of the secondary-amyloid spleen. In the case of theprimary-amyloid spleen, the decrease was from I0.3 mg./Ioo mg. ofsample in the crudest specimen (Specimen A) to 2.4 mg./Ioo mg. ofsample in the purest preparation (Specimen C). At the same time, thedecrease in the individual lipid moiety was roughly proportionate exceptfor free fatty acids. In the secondary-amyloid spleen, free fatty acidschanged from i.6 mg./ioo mg. of sample in Specimen A to i.8 mg./Ioomg. of sample in Specimen B. In the primary amyloid spleen, however,free fatty acid did not decrease when Specimen B was further purified
4| Area
20
16
14TEXT-FIG. 3. Weight
area curve demonstratinglinear relationship existing 12between square root ofsample spot area on T.L.C. 10and logarithm of theweight. 8
6
4
2
Tripaimitin
0.5 1.0 1.5
log. weight
875May I967
0.5 1.0 1.5 2.0
0o It '!. q 'tI0 o%G t.. %O 0%. 4 in '4 ) N4
t-0 0%~4. " .4 0
%R O I- °° -? I4- C0% ) %O '4
4)
'4i .-406 00.4
00 It IlN 00 Il t-.) 00 4N C .40
'I N H
. 00 0
0 C) " in )
.4C40 '4 C
4 so , *' 0. 00
fo ct I6 ?C)cooin 0% V) ' oh 0
Ct ) .4 ' .? 0t4 ") m " m O
o. Il ? v v0% 00 0 -*
Ca *
0
0
0
809
*.0o4
00
0004
000
0
H0
a4tm
" ts1-
I %)E .
5'..._
C-I.5
UItS
%n m O. t? Ni
tsl. 4t00 .4"'4 to) C-1)
"It %q in 19 '
C% O. 0 0 0%mE CE 4. CE) '
inC)1j0% 0q Cin t-.. t CE)
'0 00 C4 00 4
CE 'E U; CE 4
%0q C9'in &.'400 in 4 '4 AE'4 '14 CE) '14
in in 4 '4 00
ea '-4 CE) 0
'4 4. in 'q 00
0s 00*' '0" )0% 4 in 4. in
11 1-- llc
19 ..4 ..F7 0f Vq al X Cd~P-
0.4N
0
0
0*"5'I
--
SX
--b
'q%0'4
1-4**
Mo

LIPID IN AMYLOID FIBRILS
to Specimen C after an initial 50%0 decrease between Specimens A andB. Qualitatively, lipids in the amyloid fibril samples were not differentfrom those in the normal spleen used as a control (Table II).
In contrast to the presence of the small but not insignificant quantityof lipids (2.4%), even in the purest preparation (Specimen C) of theprimary-amyloid spleen, no detectable lipids were found in the amyloidfibril hydrolysate by pronase when lipids had been extracted from thefibrils prior to the enzymatic digestion. Under the conditions employed,micro-Kjeldahl determinations of nitrogen in the amyloid fibrils and thehydrolysate indicate that approximately 85-90% of the amyloid fibrilswere digested by pronase.32 Thus it would appear that no detectablequantity of lipid exists in the interstices of amyloid fibrils or within theirsubstance.The long-chain fatty-acid profile of the amyloid fibrils by G.L.C.
showed satisfactory separation of both the saturated and unsaturatedfatty-acid methyl esters with a chain length of C-I4 through C-20.Palmitic (i6:o), oleic (i8: i), and linoleic (I8:2) acids were the majorcomponents and myristic (I4:o), palmitoleic (i6:i), stearic (i8:o) andarachidonic (20:4) acids were present in minor quantities. Here again,the fatty-acid composition of the amyloid preparations showed no sig-nificant qualitative difference from the normal spleen used as a control(Table III).
High-resolution electron-microscopic study by the techniques of nega-tive staining of amyloid fibrils before and after lipid extraction disclosedno change in the ultrastructure of amyloid after lipid extraction (Fig. 3Aand B). Characteristic 75 A diameter and subfibrillar structure19 33 werefound to be unchanged in the post-lipid-extraction samples.
DIscussIoNIn interpreting the published data in biochemical studies of amyloid,
the degree of purity of the amyloid samples used is of utmost importance.The fact that the majority of the reported chemical analyses were doneon whole amyloid tissues or only partially purified amyloid is largelyresponsible for the conflicting and varying results that exist in the litera-ture. Limitations inherent in analyzing partially purified amyloid areparticularly serious for those substances which are usually found insmall quantities, such as carbohydrates. To overcome such difficulties,fractionation schemes were devised to yield progressively purer amyloidfibrils for biochemical analysis of lipids and simultaneous examination ofparallel specimens in the electron microscope. At the end of fractiona-tion, it was possible to harvest amyloid fibrils which appeared almostcompletely pure as judged by electron microscopy. Currently, the ultra-structure is the only definite means for identifying and estimating the
877May I967

KIM, SHIRAHAMA, AND COHEN
purity of amyloid since the fibrillar nature of amyloid is well established,while the exact chemical composition is as yet undetermined. It must beadmitted, however, that sole reliance on electron microscopy for estima-tion of the purity of amyloid fibrils is not completely satisfactory sincethe highest resolution possible may not detect slight ultrastructural al-terations or the presence of minute amounts of contaminants.
In this study, 3 fractionation procedures for amyloid fibrils were em-ployed. The first 2 methods utilized mild physical procedures which weredesigned to effect separation of amyloid fibrils with minimum alterationof the physicochemical structure of the fibrils. The first procedure, whichwas used to obtain the Specimen A, consists of homogenizing amyloid-laden tissue, centrifuging, and harvesting the top layer which has beenfound to be rich with amyloid fibrils.20 Electron micrographs of theSpecimen A showed moderately pure amyloid fibrils but some gross con-tamination was also evident. The second method consisted of a 3-stagesucrose gradient centrifugation procedure which gave a yield of purerfibrils (Specimen B). The contaminant present in the Specimen A waslargely eliminated, but the details of amyloid fibrils were not well de-lineated in areas where amyloid fibrils were packed close together.Whether this represents incomplete elimination of contaminants pos-sibly present between the closely adjacent fibrils or merely tight packingof fibrils is not known. Collagen fibers were also occasionally found.Furthermore, in these methods one could not completely exclude thepossibility of minute contaminating substances closely adsorbed on thefibrils themselves.The third fractionation procedure was devised to eliminate possible
contaminants which the mild physical methods failed to do. The pro-cedure utilized the principle of enzymatic degradation and chemicalwashings of the contaminants. First, the Specimen B was incubated ino.i M phosphate buffer (pH 7.4) at 370 C. to allow autolysis of possiblecontaminating tissue protein to take place. Uncontrolled overgrowth ofcontaminating bacteria was prevented by toluene. The sediment wasthen incubated with collagenase to digest collagen fibrils, and washedwith a io% NaCl solution for the purpose of removing mucopolysac-charides, nucleoproteins and other contaminating proteins which mightbe present. Lastly, the sediment was washed with 3%o ammonium hy-droxide solution for more complete elimination of any remaining con-taminating proteins. After centrifuging the supernatant at very highspeed, the sediment was dialyzed against demineralized distilled waterand lyophilized. Between each of the different steps, the sample wascarefully washed and centrifuged. Extensive electron-microscopic ex-aminations of the amyloid fibrils before and after each step failed toreveal any alterations in their ultrastructure. Solubility of amyloid is
878 Vol. 50, No. f

LIPID IN AMYLOID FIBRILS
increased in alkaline pH,34 and a considerable amount of amyloid is ex-pected to have been lost during washing with 3%So ammonium hydroxidesolution (pH I I.5). The remaining amyloid fibrils which were harvested,however, were identical in ultrastructure to the original amyloid fibrils.The resultant final sample, the Specimen C, was almost completely pureamyloid fibrils, free of any visible contaminants.
While the fractionation procedure outlined above was highly success-ful in producing the desired pure amyloid fibrils, its yield was very smalland amounted to I0-20% of the starting material. Despite the limitedquantity of samples for analysis, the amounts were easily enough formicrolipid analysis by qualitative and quantitative thin layer chro-matography.When frozen sections of the amyloid-laden spleens were stained for
lipids, there were abundant oil red 0-positive granules confirming thepresence of lipids in the whole amyloid tissue as reported by others.3-7Also T.L.C. examinations of the lipid extracts of the amyloid fibrils indifferent stages of fractionation demonstrated the presence of varyingamounts of cholesterol, cholesterol esters, phospholipids, free fattyacids, and triglycerides in all the samples examined. The quantity oftotal lipids present in our crude amyloid fibril preparations, Specimen A,was considerable (i9.0% in the secondary-amyloid spleen and I0.3%in the primary-amyloid spleen). Similar quantities of lipids have beenreported to be present in crude amyloid preparations by others. Forexample, Calkins and Cohen observed in whole homogenized amyloid aI % ether-soluble fraction which was not subjected to further analysis.14Battaglia and Pernis reported a total lipid content of I4.3 gm./IOO gm.of protein in amyloid.'6 The latter authors stated in the same report thatamyloid had a cholesterol content of 3.6 gm./ioo gm. of protein. Accord-ing to Letterer and co-workers, I.6-5o% of amyloid was cholesterol.15Our analyses for total cholesterol in Specimen A showed 6.3% in thesecondary amyloid spleen and 6.o%o in the primary amyloid spleen.The above data alone may be taken to indicate that lipid is a signifi-
cant component of amyloid. However, quantitative measurements oflipids in purer amyloid preparations demonstrated progressive decreasein the total lipid as well as in each lipid moiety as the samples becameprogressively purer by electron-microscopic observations. For example,the total lipid decreased from 10.3%o in Specimen A, the crudest speci-men, to 2.4%o in Specimen C, the purest specimen, in the case of primaryhuman amyloid spleen. Thus, the data from lipid analyses correlatedwell with the electron-microscopic observations that our amyloid fibrilpreparations became progressively and significantly purer following eachfractionation procedure.
879May I967

KIM, SHIRAHAMA, AND COHEN
The one exception to this general trend was the free fatty-acid moietywhich either remained unchanged or decreased relatively less (TableII). This does not necessarily imply that free fatty acids are a moreintegral part of amyloid fibrils than other lipid moieties. A more likelyexplanation for this unexpected finding is that it probably represents anartifact caused by degradation of triglycerides and other fatty-acid-containing lipid moieties (i.e., phospholipids) during tissue homogeniza-tion and fractionation procedures (even though every precaution wastaken to minimize such degradation), for we have observed in our labora-tory progressive increase in free fatty acids and decrease in triglycerideswhen homogenates were spot checked by T.L.C. during tissue homogeni-zation at o0 C. This is quite evident in Specimen A of the normal spleenwhich contained I2.0 mg. of free fatty acids per ioo-mg. sample in com-parison to only 2.0 mg. of triglycerides per ioo-mg. sample. It is dis-tinctly unusual to see so much free fatty acid in a tissue sample.
Despite the progressive decrease in the total lipids as amyloid fibrilsbecame increasingly purer, the persistence of lipids even in the purestpreparation raised the possibility that lipids indeed are an integral com-ponent of amyloid. In addition, the possibility of the presence of lipidsin the interstices of amyloid fibrils or within amyloid fibrils, which mightmake conventional lipid extraction procedures relatively ineffective, hadto be considered. In an attempt to provide answers to these questions,sucrose-separated amyloid fibrils were digested using a potent proteoly-tic enzyme, pronase, which is capable of hydrolyzing 85-90o% of amyloidfibrils. The absence of detectable amounts of lipids in the hydrolysateindicates that the lipid extraction procedure previously used was effec-tive and that no significant quantity of lipid was present within amyloidfibrils or as integral components of them.An answer to the important question of whether minute quantities of
lipid not detectable by the present techniques exist in amyloid fibrilsmust await the establishment of chemical criteria for estimating thepurity of amyloid. However, we are inclined at the moment to acceptthe conclusion that lipid is present in amyloid only as a contaminant.This is based on the observations that (i) there were no fine structuralalterations in the post-lipid-extraction samples of amyloid, (2) therewas a progressive decrease in the total lipid as the samples became pro-gressively purer, and (3) no detectable lipid was present in the enzymatichydrolysate of post-lipid-extraction amyloid fibrils.
SUMMARYi. A new purification procedure for isolating amyloid fibrils was de-
scribed. The procedure utilizing the principle of enzymatic degradation
880 Vol. 50, No. 5

May I967 LIPID IN AMYLOID FIBRILS 88i
and chemical treatment to wash out tissue protein contaminants yieldedalmost pure amyloid fibrils whose ultrastructure was virtually free ofvisible contaminants.
2. Lipid analyses demonstrated the presence of considerable quanti-ties of lipid in the crude amyloid preparations. However, total lipidswere found to decrease proportionately as the samples became progres-sively purer.
3. No detectable lipid was found in the enzymatic hydrolysate of post-lipid-extracted amyloid fibrils.
4. High-resolution electron microscopy revealed no alterations inthe ultrastructure of the post-lipid-extraction samples of the amyloidfibrils.
5. On the basis of these observations, lipid would appear to be inamyloid only as a contaminant.
REFERENCESi. KIM, I. C., and COHEN, A. S. An analysis of isolated amyloid fibrils for lipids
(abst.). Fed Proc 24:366, I965.2. COHEN, A. S. Constitution and genesis of amyloid. Int Rev Exp Path 4:I59-
243, I965.3. WEISMANN, R. E., CLAGETT, 0. T., and McDONALD, J. R. Amyloid disease of
the lung treated by pneumonectomy: Report of a case. J Thor Surg. z6:269-28I, I947.
4. STARX, D. B., and McDONALD, J. R. Amyloid tumors of the larynx, trachea andbronchi: A histologic study of fifteen cases. Amer J CGin Path I8:778-788,1948.
5. SYMMERS, W. ST. C. Primary amyloidosis: A review. J Clin Path 9:I87-2II,I956.
6. McALPINE, J. C., and FULLER, A. P. Histochemistry of fresh frozen material inlocalized laryngeal amyloidosis. J Path Bact 88:455-462, I964.
7. SORENSON, G. D., HEEFNER, W. A., and KRKPATRICK, J. B. Experimentalamyloidosis. II. Light and electron microscopic observations of liver. AmerJ Path 44:629-644, i964.
8. WINDRUM, G. M., and KRAMER, H. Some observations on the histochemical re-action of amyloid. Arch Path (Chicago) 63:373-378, I957.
9. ARVY, L., and SORS, C. ttude histochimique de la substance amyloide. Actahistochem (Jena) 6:77, I958.
IO. PFEIFFER, H. H. Zur topochemischen Analyse von Amyloidablagerungen aufGlycoproteide. Acta Histochem (Jena) 8:97-IO4, I959.
II. BRAUNSTEIN, H., and BUERGER, L. A study of the histochemical and stainingcharacteristics of amyloid. Amer J Path 35:79I-800, I959.
I12. DIEZEL, P. B., and PFLEIDERER, A., JR. Histochemische und polarisationsoptische Untersuchungen am Amyloid. Virchow Arch Path Anat 332:552-567, 1959.
I3. PEARSE, A. G. E. Histochemistry, Theoretical and Applied (ed. 2). Little, Bos-ton, I960.
14. CALKINS, E., and COHEN, A. S. Chemical composition of amyloid (abst.). JClin Invest 37:882-883, I958.

882 KIM, SHIRAHAMA, AND COHEN Vol. 50, No. s
I5. LETTERER, E., GEROR, W., and ScHNEInRE, G. Vergleichende Untersuchungenfiber den Aminos'aurenbestand von Serum-Einweiss, Lebereiweiss, Amyloid,Hyalin und Kollagen. Virchow Arch Path Anat 327:327-342, 1955.
i6. BATTAGLIA, S., and PPERNis, B. Sulla composizione chimica della sostanza amiloidein un caso di amiloidosi primitiva (Paramiloidosi neoplastiforme). InstitutoLombardo di Scienze e Lettere 92:45-54, I957.
I 7. WAGNER, B. M. Histochemical studies of fibrinoid substances and other abnormaltissue proteins; protein character of amyloid. Arch Path (Chicago) 60:22I-229, 1955.
I8. COHEN, A. S., and CALKINS, E. Electron microscopic observations on a fibrouscomponent in amyloid of diverse origins. Nature (London) Z83:1 202-I203,I959.
I9. SHIRAHAMA, T., and COHEN, A. S. The structure of amyloid fibriIs after negativestaining and high resolution electron microscopy. Nature (London) 206:737-738, I965.
20. COHEN, A. S., and CALKINS, E. The isolation of amyloid fibrils and a study ofthe effect of collagenase and hyaluronidase. J Cell BiOl 21:48I-486, 1964.
2I. COHEN, A. S. Preliminary chemical analyses of partially purified amyloid fibrils.Lab Invest z5(i, Part i):66-83, I966.
22. LUFT, J. H. Improvements in epoxy resin embedding methods. J Biophys Bio-chem Cytol 9:409-4I4, ig6i.
23. MILLONIG, G. "Further Observations on a Phosphate Buffer for Osmium Solu-tions in Fixation." In Electron Microscopy; Fifth International Congress forElectron Microscopy held in Philadelphia, Pennsylvania, August 29th to Sep-tember Sth, z962, Vol. 2, BREESE, S. S., Ed. Acad Press, New York, I962, P.P-8.
24. KARNOVSKY, M. J. Simple methods for "staining with lead" at high pH in elec-tron microscopy. J Biophys Biochem Cytol XI:729-732, I96I.
25. BRENNER, S., and HORNE, R. W. A negative staining method for high resolu-tion electron microscopy of viruses. Biochim Biophys Acta 34:103-IIO, I959.
26. FOLCH, J., LEES, M., and SLOANE STANLEY, G. H. A simple method for theisolation and purification of total lipides from animal tissues. J Biol Chem226:497-509, I957.
27. MANGOLD, H. K. Thin layer chromatography of lipids. I Amer Oil Chem Soc38:708-727, I96I.
28. PURDY, S. J. and TRUTER, E. V. Quantitative analysis by thin-film chromatog-raphy. Analyst 87:802-80g, I962.
29. BARTLETT, G. R. Phosphorus assay in column chromatography. J Biol Chem234:466-468, I959.
30. METCALFE, L. D., and SCHMITZ, A. A. The rapid preparation of fatty acid estersfor gas chromatographic analyses. Anal Chem 33:363-364, 196I.
3I. HORNING, E. C., AHRENS, E. H., JR., LIPsKY, S. R., MATTASON, F. H., MEAD,J. F., TURNER, D. A., and GOLDWATER, W. H. Quantitative analysis of fattyacids by gas-liquid chromatography. J Lipid Res 5:20-27, I964.
32. KIM, I. C., FRANzBLAu, C., and COHEN, A. S, Unpublished data.33. SHIHAMA, T., and COHEN, A. S. High resolution electron microscopic analysis
of the amyloid fibril. J Cell Biol (In press)34. NEWCOMBE, D. S., and COHEN, A. S. Solubility characteristics of isolated
amyloid fibrils. Biochim Biophys Acta x04:480-486, I965.The authors would like to acknowledge the expert technical assistance of Orville
Rodgers, the critical review of the manuscript and phosphorus determinations by Dr. AramChobanian, and photographic assistance of David Feigenbaum.

LIPID IN AMYLOID FIBRILS
L...._9 ifi 2 . r qi|| 0! r 4 # 9 >A
FIG I. Frozen section of amyloid-laden spleen stained with oil red 0. Dull graybackground (orange-brown in original) is nonspecific staining of connective tis-sue and amyloid, while dark black globules at arrows (bright orange in original)represent lipid droplets. X 400.
May I_967 883
I

KIM, SHIRAHAMA, AND COHEN
FIG. 2. Electron micrographs of amyloid fibrils obtained from Specimens A, B, andC. All preparations shadow-casted with platinum-palladium and photographedin Siemens Elmiskop I electron microscope. X 40,000. A. Specimen A. Top-layer separation technique provides abundant amyloid fibrils with some amor-phous debris and surface contamination. B. Specimen B. Sucrose-separatedpreparation demonstrates less contamination but more closely packed amyloidfibrils, with occasional fuzziness of surface structure. C. Specimen C. Fibrilspurified by chemical and enzymatic means appear virtually free of contamination.All preparations indicate that amyloid fibrils are thin, long, nonbranching, rigidrods. There are occasional twists and lateral aggregation occurs often.
884 Vol. so, No. 5

LIPID IN AMYLOID FIBRILS
2A
2BK'':o;]M
.s .2i .r..3, . .
2C
885May I967
.f w.
ew9 ...:.: .:. kO-s,.*: .. 8::. -1-
." o
,;! --.:, ...i6 o' .:rlrk. .'
-'s'-tA.,,- F'.q " .
1. f-0

kIM, SHIRAHAMA, AND COHEN
3A
3B
FIG. 3. Electron micrographs of amyloid fibrils before and after lipid extraction. Fibrils were pre-pared by technique of negative staining25 and photographed in Siemens Elmiskop I elec-tron microscope. X 320,000. A. Amyloid fibrils before lipid extraction, demonstrating fila-mentous and protofibrillar ultrastructure 33 at high resolution. B. Amyloid fibrils after lipidextraction, demonstrating no detectable change in high-resolution ultrastructure.
886 Vol. 50, Vo.. 5