novel mechanism for nisin resistance via proteolytic ... agents and... · protein (nsr) (10)....
TRANSCRIPT

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, May 2009, p. 1964–1973 Vol. 53, No. 50066-4804/09/$08.00�0 doi:10.1128/AAC.01382-08Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Novel Mechanism for Nisin Resistance via Proteolytic Degradation ofNisin by the Nisin Resistance Protein NSR�
Zhizeng Sun,1,2 Jin Zhong,1* Xiaobo Liang,1† Jiale Liu,1 Xiuzhu Chen,1 and Liandong Huan1*State Key Laboratory of Microbial Resources, Institute of Microbiology, Chinese Academy of Sciences, Beijing 100101,
China,1 and Graduate University, Chinese Academy of Sciences, Beijing 100039, China2
Received 15 October 2008/Returned for modification 8 January 2009/Accepted 27 February 2009
Nisin is a 34-residue antibacterial peptide produced by Lactococcus lactis that is active against a wide rangeof gram-positive bacteria. In non-nisin-producing L. lactis, nisin resistance could be conferred by a specificnisin resistance gene (nsr), which encodes a 35-kDa nisin resistance protein (NSR). However, the mechanismunderlying NSR-mediated nisin resistance is poorly understood. Here we demonstrated that the proteinwithout the predicted N-terminal signal peptide sequence, i.e., NSRSD, could proteolytically inactivate nisin invitro by removing six amino acids from the carboxyl “tail” of nisin. The truncated nisin (nisin1–28) displayeda markedly reduced affinity for the cell membrane and showed significantly diminished pore-forming potencyin the membrane. A 100-fold reduction of bactericidal activity was detected for nisin1–28 in comparison to thatfor the intact nisin. In vivo analysis indicated that NSR localized on the cell membrane and endowed hoststrains with nisin resistance by degrading nisin as NSRSD did in vitro, whereas NSRSD failed to conferresistance upon the host strain. In conclusion, we showed that NSR is a nisin-degrading protease. ThisNSR-mediated proteolytic cleavage represents a novel mechanism for nisin resistance in non-nisin-producingL. lactis.
Nisin is a 34-residue antibacterial peptide and a widely usedfood preservative because of its potent bactericidal activityagainst a variety of food spoilage bacteria. It is produced bysome strains of Lactococcus lactis and contains three dehy-drated amino acids and five lanthionine (Lan) or �-methyllan-thionine (MeLan) rings, for which reason the term lantibiotics(lanthionine-containing antibiotics) was coined (30). Nisin Aand nisin Z are two well-studied natural variants of nisin,differing with a residue at position 27 (His for nisin A and Asnfor nisin Z) but showing no difference in antibacterial activity.Studies have shown that nisin exerts its bactericidal activity bypore formation and disruption of cell wall synthesis by specificbinding to lipid II, an essential intermediate in peptidoglycansynthesis (4, 14, 16, 46). In addition, nisin can induce cellautolysis (2) and inhibit the outgrowth of bacterial spores(13, 32).
However, the bactericidal efficacy of nisin has been compro-mised by the occurrence of nisin resistance in various bacteria.It could be acquired by repeated exposure of sensitive strainsto increasing concentrations of nisin through alterations in theexpression of genes involved in cell wall and cytoplasmic mem-brane biosynthesis, which is referred to as a physiological ad-aptation (12, 31, 36). In L. lactis with acquired nisin resistance,the ABC transporter might also be involved in removing nisinfrom the cytoplasmic membrane (23). Furthermore, a nisin-inactivating enzyme was found in several nisin-resistant strains
of Bacillus subsp. (17). It was later identified as a dehydroala-nine (Dha) reductase, reducing the carboxyl Dha to Ala toinactivate nisin as well as subtilin, a close relative of nisin inBacillus subtilis (18). Nisin resistance in the nisin nonproducerL. lactis subsp. diacetylactis DRC3 was reported to be con-ferred by a specific nisin resistance gene (nsr), which is locatedon a 60-kb plasmid and encodes a 35-kDa nisin resistanceprotein (NSR) (10). Thereafter, several groups have isolatednisin-resistant lactococcal strains with nsr on the plasmid (28,48). From fresh milk samples, our lab isolated several nisin-insensitive lactococcal strains, one of which was L. lactis subsp.lactis TS1640 (43). In the strain, a 47-kb plasmid, designatedpTS50, was identified which encodes nisin resistance followingtransformation into L. lactis MG1363, a highly nisin-sensitivestrain. A nisin resistance determinant was localized on a 1.9-kbEcoRI fragment by Southern blotting hybridization probedwith a reported nsr sequence. The fragment was subcloned andsequenced. Its open reading frame encoding sequence is 99%identical to that reported previously (10), suggesting a com-mon origin.
Recent studies of nsr have been focused on the constructionof food-grade vectors utilizing its nisin resistance phenotype(29), whereas virtually nothing is known about the underlyingmechanism. It has been proposed that NSR-mediated nisinresistance might occur via proteolytic degradation of nisin forthe presence of a C-terminal conserved tail-specific protease(TSPc) domain at the C terminus of NSR (7, 33). Tail-specificproteases are a group of endoproteases involved in processingtheir polypeptide substrates with nonpolar C termini. So far,the well-characterized tail-specific proteases include Tsp inEscherichia coli (19, 21, 39), CT441 in Chlamydia trachomatis(25, 26), and D1P in green alga (27) and in spinach (35). All ofthese proteases bear a conserved PDZ domain adjacent tothe TSPc domain (Fig. 1A). Studies have shown that the
* Corresponding author. Mailing address: Institute of Microbiology,Chinese Academy of Sciences, A3 Datun Road, Chaoyang District,Beijing 100101, China. Phone: (86) 10 6480 7439. Fax: (86) 10 64807401. E-mail for Jin Zhong: [email protected]. E-mail for LiandongHuan: [email protected].
† Present address: Department of Pediatric Dentistry, University ofAlabama at Birmingham, Birmingham, AL 35294.
� Published ahead of print on 9 March 2009.
1964
at INS
TIT
UT
E O
F M
ICR
OB
IOLO
GY
CA
S on A
pril 25, 2009 aac.asm
.orgD
ownloaded from

PDZ domain is indispensable for binding of Tsp with non-polar C termini of its peptide substrates and thus for thecatalytic activity (1, 40). However, the domain is absent inNSR, the unique protein with a TSPc domain identified in L.lactis. In addition, nisin harbors charged and polar residues(Lys, His, and Ser) at the C terminus, which is also differentfrom the sequence determinants of C-terminal substratesrecognized by Tsp (20). Therefore, it is necessary to eluci-date whether NSR could actually cause proteolysis of nisinand, if not, how NSR-mediated nisin resistance occurs in L.lactis.
In this study, we demonstrate that the purified NSRSD(NSR without the predicted N-terminal signal peptide se-quence) could proteolytically inactivate nisin in vitro by cleav-ing the peptide bond between MeLan28 and Ser29. The trun-cated nisin (nisin1-28), compared with intact nisin, showed amarkedly reduced affinity for the lactococcal membrane, asignificantly diminished pore-forming potency in the targetmembrane, and a 100-fold-lower bactericidal activity against L.lactis MG1363. Western blotting hybridization revealed thatboth NSR and NSRSD were localized on the cell membraneof L. lactis. However, unlike the former, the anchoredNSRSD had no ability to degrade nisin and therefore failedto endow the host strain with nisin resistance. Thus, wepresent a novel mechanism for nisin resistance in non-nisin-producing L. lactis.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions. Bacterial strains and plas-mids used in this study are listed in Table 1. Molecular cloning experiments wereconducted using E. coli strain JM109. The bacteria were cultured in LB brothplus 100 �g/ml ampicillin (Amp) or erythromycin (Em) when necessary withaeration at 37°C. E. coli BL21(DE3) was used as the host strain for the expres-sion of glutathione-S-transferase (GST)-fused proteins under the same growthconditions as for E. coli JM109. L. lactis strains were cultivated at 30°C in M17medium (44) supplemented with 0.5% (wt/vol) glucose (GM17 medium), and 5�g/ml Em was added when needed. In addition, unless otherwise indicated, 5�g/ml and 1.25 �g/ml nisin A (Sigma) were used for selection of L. lactis TS1640and L. lactis MG1363/pTS50, respectively. To produce solid media, agar wasadded into different media at a final concentration of 1.2% (wt/vol).
Manipulation of DNA. E. coli plasmid DNA was isolated by the alkali lysismethod (38). Plasmids of L. lactis were isolated according to the method de-scribed previously by O’Sullivan and Klaenhammer (34). Restriction endonucle-ases, ligases, and polymerases were used according to the recommendations ofthe manufacturers. Linear DNA fragments were electrophoretically purifiedusing a DEAE-cellulose membrane. Competent E. coli cells were prepared andtransformed by standard techniques (38). L. lactis was transformed by electro-poration as described previously (15). All other procedures were conductedaccording to the standard protocols.
Overexpression and purification of NSRSD in E. coli. To obtain a DNAfragment without the signal peptide (SP) coding sequence, i.e., nsrSD, pMG36e-PnsrM was used as the template for PCR with the primer pairs Nsr F1 and NsrF2 (Table 2). The purified fragment was digested with EcoRI and XhoI andligated into the same sites of pGEX-6P-1 (GE Healthcare) encoding for Pre-Scission protease cleavable N-terminal GST-tagged fusion proteins. The result-ing plasmid, designated pSZ311 (Table 1), was transformed into E. coliBL21(DE3) in which the expression of NSRSD was fused with GST and under
FIG. 1. Functional analysis of purified NSRSD. (A) Schematic structures of NSR and NSRSD in comparison with well-characterized tail-specific proteases. Striped blocks represent the C-terminal conserved tail-specific protease (TSPc) domain. Dotted blocks stand for the PDZdomain. Gray blocks in NSR and NSRSD represent hydrophobic regions. The N-terminal 27 amino acids of NSR including one hydrophobic regionthat constitute a predicted SP sequence have been deleted in NSRSD. (B) SDS-PAGE analysis of purified NSRSD. Lane 1, molecular size markerwith masses in kDa indicated on the left; lane 2, noninduced bacterial lysate; lane 3, supernatant of induced bacterial lysate after sonication andcentrifugation; lane 4, precipitate of induced bacterial lysate after sonication and centrifugation; lane 5, purified NSRSD. The positions ofGST-NSRSD and NSRSD are indicated by arrows. (C) Determination of antibacterial activities of control (left) or NSRSD-digested nisin (right)against L. lactis MG1363 by the agar diffusion assay.
VOL. 53, 2009 PROTEOLYTIC MECHANISM FOR NISIN RESISTANCE 1965
at INS
TIT
UT
E O
F M
ICR
OB
IOLO
GY
CA
S on A
pril 25, 2009 aac.asm
.orgD
ownloaded from

the control of an isopropyl 1-thio-�-D-galactopyranoside (IPTG)-inducible pro-moter.
For the expression of GST-NSRSD, pSZ311-transformed E. coli BL21(DE3)was grown in LB broth containing 100 �g/ml Amp at 37°C until the opticaldensity at 600 nm (OD600) reached 0.6. Next, IPTG was added to a final con-centration of 0.6 mM and the cultures were incubated at 16°C for 6 h undergentle shaking. The bacterial cells were collected by centrifugation, washed withice-cold phosphate-buffered saline (PBS), and resuspended in 50 ml of PBS perliter of culture. The cells were disrupted by sonication, and the unbroken cellswere removed by centrifugation at 10,000 � g for 10 min at 4°C. Next, GST-NSRSD in the supernatant was purified by column chromatography with gluta-thione-agarose beads (GE Healthcare), and the GST tag was cleaved on acolumn with PreScission Protease (GE Healthcare), following the manufactur-er’s instructions. Additional gel filtration chromatography was carried out on aSuperdex 75 10/300 GL column (GE Healthcare) to remove traces of unspecificproteins in NSRSD. Finally, the concentration of NSRSD was determined usingthe bicinchoninic acid protein assay reagent (Pierce) with bovine serum albuminas a standard.
Assays for NSRSD activity in vitro. Fifty micrograms of pure nisin Z (agenerous gift from Zhejiang Silver-Elephant Bio-engineering Co., Ltd., China)were incubated with 2 �g purified NSRSD in a 50-�l reaction buffer (50 mMTris-HCl, pH 6.0) at 30°C for 6 h. The incubation without NSRSD was used asthe control. After the reaction, 1 �g of the digested nisin Z or control sample ina volume of 20 �l was transferred into the wells lawned with the indicator strainL. lactis MG1363. The plate was kept at 4°C for 2 h for diffusion and incubatedat 30°C overnight. Additionally, the two samples were subjected to the analysesof matrix-assisted laser desorption/ionization time-of-flight mass spectrometry(MALDI-TOF MS) and electrospray ionization mass spectrometry (ESI MS).
Characterization of proteolytic activity of NSRSD against nisin Z. For quan-titative analyses of nisin Z and its digested products, 20 �l of the aliquots were
injected onto a SinoChrom ODS-BP 5-�m, 4.6-mm by 250-mm reverse-phasecolumn (Elitehplc, Dalian, China) in a Waters 600 high-performance liquidchromatography (HPLC) system. Peptides were eluted at a flow rate of 1 ml/minusing a linear gradient of 10 to 50% acetonitrile containing 0.1% trifluoroaceticacid (TFA) (vol/vol/vol) for 15 min and then 90% acetonitrile in 0.1% TFA for5 min. The eluates were monitored with a fluorescence detector at 220 nm, andthe amount of nisin Z was calculated using a calibration curve made by injecting20 �l of pure nisin Z at concentrations ranging from 18 to 600 �M.
Kinetic constants for cleavage of nisin Z were determined by mixing 43 nMNSRSD with 15 to 600 �M nisin Z in reaction buffer (50 mM Tris-HCl, pH 6.0)at 30°C for 2 min. The reaction mixture was quenched with 0.1% TFA andanalyzed by reverse-phase HPLC (RP-HPLC) as described above. The unit ofcleavage was determined in �M/min by recording the rate of appearance of onedigested product. By fitting the data to a Lineweaver-Burk double-reciprocalplot, the apparent kinetic parameters Km and kcat were obtained.
Overexpression of NSR and NSRSD in L. lactis. The plasmid pMG36e-PnsrMwas used as the template to amplify the P59 promoter with the primer pair P59F and P59 R (Table 2). The purified product was digested with EcoRI and SacIand cloned into pUC18 for the plasmid pUC18-P59 (Table 1). Then, nsr wasamplified from pMG36e-PnsrM using the primer pair Nsr F2 and Nsr R2 (Table2) and cloned downstream of P59 in pUC18-P59 to generate the plasmid pUC18-P59-nsr (Table 1). An EcoRI- and PstI-digested fragment containing P59-nsr wassubcloned into pMG36e restricted by the same enzymes, yielding the lactococcalexpression plasmid pSZ531 (Table 1). Similarly, nsrSD was amplified frompMG36e-PnsrM by PCR with the oligonucleotides Nsr F3 and Nsr R2 (Table 2)and cloned downstream of P59 in pUC18-P59 to produce the plasmid pUC18-P59-nsrSD (Table 1). Then, an EcoRI- and PstI-digested fragment containingP59-nsrSD was ligated into the corresponding sites in pMG36e, resulting in annsrSD expression vector designated pSZ537 (Table 1).
The constitutive expression vectors pSZ531 and pSZ537 were separately trans-
TABLE 1. Bacterial strains and plasmids used in this study
Strain or plasmid Relevant genotype/description Reference or source
StrainsE. coli JM109 recA supE44 endA1 hsdR17 gyrA96 relA1 thi �(lac-proAB) F��traD36 proAB�
lacIq lacZ�M15�38
E. coli BL21(DE3) F ompT hsdSB (rB mB
) gal dcm (DE3) InvitrogenL. lactis MG1363 Plasmid-free derivative of NCDO712 11L. lactis MG1363/pTS50 L. lactis MG1363 transformed by pTS50, nisin resistant 43L. lactis TS1640 A naturally occurring nisin-resistant strain 43
PlasmidspUC18 2.7 kb, Amp resistant, cloning vector for E. coli 38pGEX-6P-1 4.9 kb, Amp resistant, expression vector for E. coli GE HealthcarepMG36e 3.6 kb, Em resistant, expression vector for L. lactis 45pMG36e-PnsrM 4.9 kb, Em resistant, pMG36e containing P59-nsr in which SacI restriction
site in nsr was mutated, template for amplification of P59 and nsr by PCROur lab
pUC18-P59 2.9 kb, Amp resistant, pUC18 containing the P59 promoter This studypUC18-P59-nsr 3.9 kb, Amp resistant, pUC18 containing P59-nsr This studypUC18-P59-nsrSD 3.8 kb, Amp resistant, pUC18 containing P59-nsrSD This studypSZ311 5.9 kb, Amp resistant, pGEX-6P-1 containing nsrSD This studypSZ531 4.6 kb, Em resistant, the 1.2-kb EcoRI/PstI P59-nsr fragment from pUC18-
P59-nsr subcloned into the same sites of pMG36eThis study
pSZ537 4.5 kb, Em resistant, the 1.1-kb EcoRI/PstI P59-nsrSD fragment frompUC18-P59-nsrSD subcloned into the same sites of pMG36e
This study
TABLE 2. Primers used in this study
Primer Sequence (5� to 3�)a Use
Nsr F1 CGGAATTCAAATTCAACATATATTTAGTAC NSRSD expression in E. coliNsr R1 CCGCTCGAGTTACTTTATTTGAGATTTTAT NSRSD expression in E. coliNsr F2 CGGAGCTCATGAAAATAGGTAAGCGAATTT NSR expression in L. lactisNsr F3 GCGAGCTCATGAAATTCAACATATATTTAGTAC NSRSD expression in L. lactisNsr R2 CGCTGCAGTAAATAACGTTAAGTTTTG NSR or NSRSD expression in L. lactisP59 F CCGAATTCTAACCAAAGAAGCGCGTAATATC Amplification of P59P59 R CGGAGCTCAATAAATATTCTCCTTCTTTATTCAGTTTTCAATGGTC Amplification of P59
a Relevant restriction sites are underlined.
1966 SUN ET AL. ANTIMICROB. AGENTS CHEMOTHER.
at INS
TIT
UT
E O
F M
ICR
OB
IOLO
GY
CA
S on A
pril 25, 2009 aac.asm
.orgD
ownloaded from

formed into L. lactis MG1363 through electroporation. For nsr or nsrSD expres-sion, overnight cultures of L. lactis MG1363 carrying the respective plasmidswere diluted 100-fold in GM17 broth containing 5 �g/ml Em and incubated at30°C. Cells were harvested at the late exponential phase of growth.
Localization analysis of NSR and NSRSD in L. lactis. Total protein extractsfrom cell cultures (containing supernatants) were fractionated following aslightly modified procedure of Piard et al. (37). Briefly, 100 ml of late-exponen-tial-phase culture was centrifuged at 8,000 � g for 5 min at 4°C. The supernatantand the cells were processed separately. The supernatant was filtered through0.2-�m-pore-size filters to remove bacteria, and proteins from 4 ml of the filtratewere precipitated with trichloroacetic acid at a final concentration of 16% (wt/vol). The resulting pellet was washed twice with 2 ml of acetone and dissolved in100 �l of 50 mM NaOH. The cell pellets were washed once with ice-cold PBS andresuspended in 5 ml PBS. The cells were disrupted by sonication, and intact cellswere removed by centrifugation at 6,000 � g for 20 min. Soluble and insolubleproteins were separated by ultracentrifugation at 120,000 � g for 2 h. Thesupernatant (cytoplasmic fraction) and the viscous pellet (cell membrane frac-tion) were separated by sodium dodecyl sulfate-polyacrylamide gel electrophore-sis (SDS-PAGE) and subjected to Western blot analysis using mouse antiserumagainst NSRSD. The binding of the antiserum was probed with a secondary goatanti-mouse immunoglobulin G conjugated with horseradish peroxidase (Sino-American Biotechnology, Henan, China) and visualized using enhanced chemi-luminescence (Applygen, Beijing, China).
Assays for nisin cleavage activity of NSR or NSRSD expressed in L. lactis. Toinvestigate whether NSR or NSRSD could degrade nisin in vivo, a series ofpeptide release assays were performed according to the method described byStein et al. (42). Flasks containing 50 ml of GM17 medium (with 5 �g/ml Emadded when needed; no nisin A was added for L. lactis TS1640 and L. lactisMG1363/pTS50) were inoculated with a 1/100 volume of overnight preculturesand incubated at 30°C until the OD600 reached 0.6. Cells were harvested bycentrifugation at 4,000 � g for 10 min, washed twice with 50 mM Tris-HCl (pH6.0), centrifuged again, and resuspended in an incubation buffer (50 mM sodiumphosphate buffer [pH 6.0], 1 M NaCl, and 1% [wt/vol] glucose) at 1-ml aliquotsin microcentrifuge tubes. Nisin Z (100 �g) was added, and the aliquots wereincubated for 30 min at 30°C with gentle shaking. After incubation, the aliquotswere centrifuged for 10 min at full speed in a microcentrifuge, and the super-natants were collected. The harvested cell pellets were washed with the incuba-tion buffer, gently mixed with 1 ml of 20% acetonitrile in water containing 0.1%TFA, and incubated with gentle shaking at 30°C for 5 min. The cells wereremoved by centrifugation at 12,000 � g for 10 min, and the supernatants werecollected. The collected supernatants (200 �l each) were subjected to RP-HPLCanalysis as described above.
Additionally, overnight precultures of recombinant L. lactis MG1363 strainswere diluted to 1/100 in 10 ml of fresh GM17 medium containing 5 �g/ml Em andincubated at 30°C until the OD600 reached 1.0. Cells were harvested by centrif-ugation at 8,000 � g for 10 min, washed once with 50 mM Tris-HCl (pH 6.0), andresuspended in 0.5 ml wash buffer. The cells were disrupted by sonication, andintact cells were removed by centrifugation at 6,000 � g for 20 min. Next, nisinZ was added to the suspension to a final concentration of 50 �g/ml. Afterincubation at 30°C for 30 min, 20 �l of the aliquots were tested for antibacterialactivity against L. lactis MG1363 by agar diffusion assay as described above.
Purification of nisin Z-derivatized fragments. Nisin Z (10 mg) was incubatedwith 20 �g NSRSD in 10 ml buffer (50 mM Tris-HCl, pH 6.0) at 30°C for 24 h.After the reaction, the mixture was lyophilized and reconstituted with 2 mldeionized water. The desired fragments, nisin Z1–28 and nisin Z29–34, werepurified using semipreparative RP-HPLC with a C18 SinoChrom ODS-BP 10-�m, 10-mm by 250-mm column (Elitehplc, Dalian, China). The column waseluted with a linear gradient of 10 to 50% acetonitrile in 0.1% TFA for 15 minand then 90% acetonitrile containing 0.1% TFA for 10 min at the flow rate of 2ml/min. Most of the solvent was removed by rotative evaporation under vacuum,followed by lyophilization. Then, nisin Z1–28 was dissolved in 0.02 M HCl, whilenisin Z29–34 was dissolved in deionized water, which was subjected to Edmandegradation sequencing using an Applied Biosystems 491 automatic sequencer.
Assays for antibacterial activities of nisin Z and nisin Z1–28. Antibacterialactivities of nisin Z and nisin Z1–28 were tested against various lactococcal strainsby a method slightly modified from that of Kuipers et al. (24). Briefly, overnightcultures of indicator organisms were diluted in GM17 broth (with 5 �g/ml Emadded when needed; no nisin A was added for L. lactis TS1640 and L. lactisMG1363/pTS50) to an OD600 of 0.025 and were divided into aliquots of 5 ml.These tubes were inoculated with different amounts of nisin Z or nisin Z1–28 andwere incubated at 30°C without shaking. Outgrowth was measured when theculture without nisin Z or nisin Z1–28 had reached an OD600 of 0.8. The lowest
concentration of nisin Z or nisin Z1–28 that resulted in no obvious growth of thetested strains (OD600 0.04) was taken as the MIC.
Determination of potassium efflux. Potassium efflux was determined accordingto the method of Xiao et al. (47) with slight modification. L. lactis MG1363 wascultured in GM17 medium plus 2.5 mM KCl at 30°C to mid-exponential phase(OD600 � 0.6 to 0.7) for cell dry weight (CDW) and potassium efflux determi-nations. Cells were harvested by centrifugation at 10,000 � g at 4°C for 10 min,washed with 0.1 M morpholine ethanesulfonic acid buffer at pH 6.3 containing0.2% (wt/vol) glucose and 0.6 mM KCl, and resuspended in the same volume of
FIG. 2. Mass spectrometric analyses of nisin and its degraded prod-ucts after cleavage by NSRSD. (A) MALDI-TOF MS analysis of nisin.(B) MALDI-TOF MS analysis of NSRSD-cleaved nisin. (C) ESI-MSanalysis of NSRSD-cleaved nisin. The peak with a molecular mass of326.9 Da represents the doubly protonated ion of the peak with amolecular mass of 652.6 Da (M1). In addition, the peak with a molec-ular mass of 1,349.8 Da is equivalent to the doubly protonated ion ofthe peak with a molecular mass of 2,697.0 Da (M2) detected byMALDI-TOF MS in panel B.
VOL. 53, 2009 PROTEOLYTIC MECHANISM FOR NISIN RESISTANCE 1967
at INS
TIT
UT
E O
F M
ICR
OB
IOLO
GY
CA
S on A
pril 25, 2009 aac.asm
.orgD
ownloaded from

the washing buffer. Then, purified nisin Z or nisin Z1–28 was added to the cellsuspension at different concentrations. After treatment for 1 h at room temper-ature, the suspension was centrifuged at 10,000 � g for 10 min. The collectedsupernatants were applied to an atomic absorption spectrometer (5100PC; Per-kin-Elmer, Ueberlingen, Germany) for determination of the potassium content.Potassium efflux was expressed in �mol/mg CDW, and each treatment wasperformed in triplicate.
Nucleotide sequence accession number. The subcloned and sequenced EcoRIfragment was assigned GenBank accesion no. AY219902.
RESULTS
Expression of NSRSD in E. coli. To obtain a sufficientamount of NSR for biochemical characterization, it was fusedwith GST and expressed in E. coli. However, the expressionlevel and solubility of GST-NSR were too low for it to bepurified (data not shown), which was probably due to theadverse effect of a putative SP sequence composed of 27 aminoacid residues at the N terminus (predicted by the softwareprogram SignalP 3.0). To test the expression without the SPregion (the product designated NSRSD), E. coli BL21(DE3)was transformed by the expression vector pSZ311, in whichexpression of NSRSD was fused with GST. After inductionwith IPTG, GST-NSRSD was detected mainly in the solublefraction of cell lysate by SDS-PAGE (Fig. 1B, lane 3). NSRSDwas purified to homogeneity by GST affinity purification, Pre-Scission protease cleavage, and gel filtration chromatography(Fig. 1B, lane 5).
In vitro degradation of nisin by NSRSD and identification ofthe cleavage site. To test whether purified NSRSD has proteo-lytic activity against nisin, it was incubated with purified nisin
and then tested against L. lactis MG1363. As shown in Fig. 1C,antibacterial activity of treated nisin decreased by a large mar-gin compared with that of the control. Subsequent MALDI-TOF MS analyses revealed the absence of nisin (3,330.6 Da)but the presence of one molecule with a smaller molecularmass of 2,697.0 Da in the reaction buffer (Fig. 2A and B). Withthe help of ESI MS, another molecule was detected, with amolecular mass of 652.6 Da (Fig. 2C). Meanwhile, doublyprotonated molecular ions of the above molecules were furtheranalyzed with ESI MS, resulting in two molecular masses of1,349.8 Da and 326.9 Da (Fig. 2C), respectively. These twofragments were then purified using semipreparative RP-HPLC, and the small one was sequenced by Edman degrada-tion analysis. Five amino acids of its N terminus were identifiedas Ser-Ile-His-Val-X (X represents a modified amino acid), anexact match to the residues from Ser29 to Dha33 of nisin. Thedata indicated that NSRSD can degrade nisin by hydrolyzingthe peptide bond between MeLan28 and Ser29, thus resulting inthe loss of antibacterial activity of nisin. Hence, NSRSD-cata-lyzed proteolysis of nisin is shown in Fig. 3.
Characterization of proteolytic activity of NSRSD againstnisin. RP-HPLC was used to analyze nisin and its digestedproducts. As shown in Fig. 4, nisin and its two digested frag-ments were completely separated, with a retention time of 16.5min for nisin and 9.8 min and 17.3 min for the two cleavedproducts, respectively. This facilitated the integration of eachpeak and the quantification of corresponding peptides.
Kinetic parameters for NSRSD cleavage of nisin were de-termined by using a reaction (conducted at 30°C and pH 6.0)
FIG. 3. Proteolysis of nisin catalyzed by NSRSD. The shadowed residues indicate amino acids which have undergone posttranslationalmodifications. A-S-A, lanthionine; Abu-S-A, �-methyllanthionine; Dha, 2,3-didehydroalanine; Dhb, 2,3-didehydrobutyrine. Molecular masses ofintact and NSRSD-cleaved nisin are indicated in parentheses to the right of the corresponding peptides.
1968 SUN ET AL. ANTIMICROB. AGENTS CHEMOTHER.
at INS
TIT
UT
E O
F M
ICR
OB
IOLO
GY
CA
S on A
pril 25, 2009 aac.asm
.orgD
ownloaded from
with nisin at concentrations ranged from 15 �M to 600 �M.Fitting the data to a Lineweaver-Burk double-reciprocal plotyields kinetic parameters, i.e., Km � 290 (�19) �M; kcat � 6.05(�0.02) s1; and kcat/Km � 20.85 (�1.34) � 103 M1 s1.
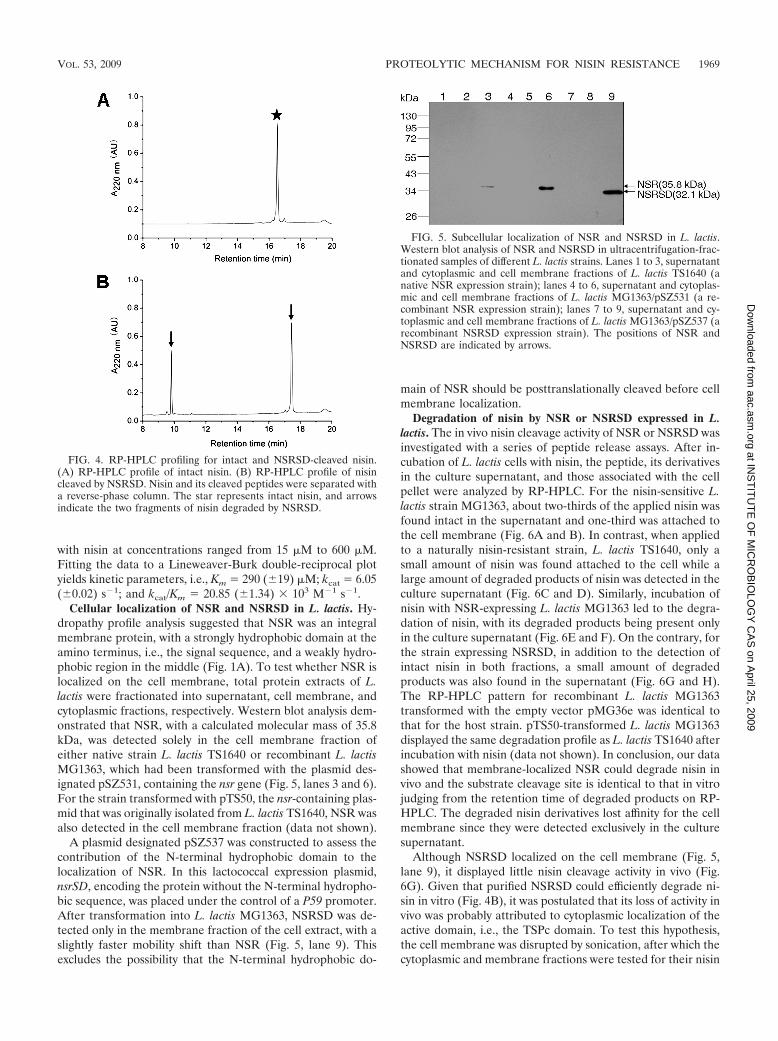
Cellular localization of NSR and NSRSD in L. lactis. Hy-dropathy profile analysis suggested that NSR was an integralmembrane protein, with a strongly hydrophobic domain at theamino terminus, i.e., the signal sequence, and a weakly hydro-phobic region in the middle (Fig. 1A). To test whether NSR islocalized on the cell membrane, total protein extracts of L.lactis were fractionated into supernatant, cell membrane, andcytoplasmic fractions, respectively. Western blot analysis dem-onstrated that NSR, with a calculated molecular mass of 35.8kDa, was detected solely in the cell membrane fraction ofeither native strain L. lactis TS1640 or recombinant L. lactisMG1363, which had been transformed with the plasmid des-ignated pSZ531, containing the nsr gene (Fig. 5, lanes 3 and 6).For the strain transformed with pTS50, the nsr-containing plas-mid that was originally isolated from L. lactis TS1640, NSR wasalso detected in the cell membrane fraction (data not shown).
A plasmid designated pSZ537 was constructed to assess thecontribution of the N-terminal hydrophobic domain to thelocalization of NSR. In this lactococcal expression plasmid,nsrSD, encoding the protein without the N-terminal hydropho-bic sequence, was placed under the control of a P59 promoter.After transformation into L. lactis MG1363, NSRSD was de-tected only in the membrane fraction of the cell extract, with aslightly faster mobility shift than NSR (Fig. 5, lane 9). Thisexcludes the possibility that the N-terminal hydrophobic do-
main of NSR should be posttranslationally cleaved before cellmembrane localization.
Degradation of nisin by NSR or NSRSD expressed in L.lactis. The in vivo nisin cleavage activity of NSR or NSRSD wasinvestigated with a series of peptide release assays. After in-cubation of L. lactis cells with nisin, the peptide, its derivativesin the culture supernatant, and those associated with the cellpellet were analyzed by RP-HPLC. For the nisin-sensitive L.lactis strain MG1363, about two-thirds of the applied nisin wasfound intact in the supernatant and one-third was attached tothe cell membrane (Fig. 6A and B). In contrast, when appliedto a naturally nisin-resistant strain, L. lactis TS1640, only asmall amount of nisin was found attached to the cell while alarge amount of degraded products of nisin was detected in theculture supernatant (Fig. 6C and D). Similarly, incubation ofnisin with NSR-expressing L. lactis MG1363 led to the degra-dation of nisin, with its degraded products being present onlyin the culture supernatant (Fig. 6E and F). On the contrary, forthe strain expressing NSRSD, in addition to the detection ofintact nisin in both fractions, a small amount of degradedproducts was also found in the supernatant (Fig. 6G and H).The RP-HPLC pattern for recombinant L. lactis MG1363transformed with the empty vector pMG36e was identical tothat for the host strain. pTS50-transformed L. lactis MG1363displayed the same degradation profile as L. lactis TS1640 afterincubation with nisin (data not shown). In conclusion, our datashowed that membrane-localized NSR could degrade nisin invivo and the substrate cleavage site is identical to that in vitrojudging from the retention time of degraded products on RP-HPLC. The degraded nisin derivatives lost affinity for the cellmembrane since they were detected exclusively in the culturesupernatant.
Although NSRSD localized on the cell membrane (Fig. 5,lane 9), it displayed little nisin cleavage activity in vivo (Fig.6G). Given that purified NSRSD could efficiently degrade ni-sin in vitro (Fig. 4B), it was postulated that its loss of activity invivo was probably attributed to cytoplasmic localization of theactive domain, i.e., the TSPc domain. To test this hypothesis,the cell membrane was disrupted by sonication, after which thecytoplasmic and membrane fractions were tested for their nisin
FIG. 4. RP-HPLC profiling for intact and NSRSD-cleaved nisin.(A) RP-HPLC profile of intact nisin. (B) RP-HPLC profile of nisincleaved by NSRSD. Nisin and its cleaved peptides were separated witha reverse-phase column. The star represents intact nisin, and arrowsindicate the two fragments of nisin degraded by NSRSD.
FIG. 5. Subcellular localization of NSR and NSRSD in L. lactis.Western blot analysis of NSR and NSRSD in ultracentrifugation-frac-tionated samples of different L. lactis strains. Lanes 1 to 3, supernatantand cytoplasmic and cell membrane fractions of L. lactis TS1640 (anative NSR expression strain); lanes 4 to 6, supernatant and cytoplas-mic and cell membrane fractions of L. lactis MG1363/pSZ531 (a re-combinant NSR expression strain); lanes 7 to 9, supernatant and cy-toplasmic and cell membrane fractions of L. lactis MG1363/pSZ537 (arecombinant NSRSD expression strain). The positions of NSR andNSRSD are indicated by arrows.
VOL. 53, 2009 PROTEOLYTIC MECHANISM FOR NISIN RESISTANCE 1969
at INS
TIT
UT
E O
F M
ICR
OB
IOLO
GY
CA
S on A
pril 25, 2009 aac.asm
.orgD
ownloaded from

FIG. 6. RP-HPLC profiles of nisin and its cleaved products by membrane-localized NSR or NSRSD in vivo. Different L. lactis strains were incubated withnisin. After centrifugation, nisin or its degraded fragments in the supernatant or attached to the cell pellet were analyzed by RP-HPLC. Asterisks represent intactnisin, and arrows indicate the two fragments of nisin degraded by membrane-localized NSR or NSRSD. Panels A, C, E, and G show peptides detected in thesupernatants of L. lactis MG1363, L. lactis TS1640, L. lactis MG1363/pSZ531, and L. lactis MG1363/pSZ537, respectively. Panels B, D, F, and H show peptidesattached to the cell pellets of L. lactis MG1363, L. lactis TS1640, L. lactis MG1363/pSZ531, and L. lactis MG1363/pSZ537, respectively.
1970
at INS
TIT
UT
E O
F M
ICR
OB
IOLO
GY
CA
S on A
pril 25, 2009 aac.asm
.orgD
ownloaded from

cleavage activities. As shown in Fig. 7 (sample 4 and sample 6),NSRSD displayed nisin cleavage activity similar to that ofNSR, which was reflected by the reduced antibacterial activityof nisin after incubation with lactococcal sonicates containingNSR or NSRSD.
Growth inhibitory activities of nisin and nisin1–28 against L.lactis. To further establish the connection between the abilityof NSR to degrade nisin and its contribution to nisin resistancein L. lactis, the MIC of nisin was determined by growth inhib-itory assays. In general, the increase of the MIC is positivelycorrelated with nisin-degrading activity (Table 3). Therefore, itcan be confirmed that NSR contributes to nisin resistance in L.lactis. The exception of L. lactis TS1640 would imply that thereare additional mechanisms conferring nisin resistance (Table3). However, when the strain was incubated overnight in GM17medium without nisin, it became less resistant to the antibac-terial peptide, with an MIC of 3 �g/ml, though expression ofNSR was not influenced (data not shown). Therefore, afterrepeated exposure to nisin during screening, L. lactis TS1640evolved unstable physiological adaptations, which were lostonce the nisin pressure was removed (3, 22).
The inhibitory activity of nisin1-28 was assayed and resultedin an MIC of 6 �g/ml against L. lactis MG1363, 100-fold higherthan that of the intact nisin molecule (0.06 �g/ml). This impliesthat the C-terminal “tail” of nisin plays an essential role in itsantibacterial activity.
Potassium efflux induced by nisin and nisin1–28. To evaluatethe pore-forming potency of nisin and nisin1–28, potassiumefflux assays were carried out. As shown in Fig. 8A, nisincaused potassium efflux from L. lactis MG1363 in a concentra-tion-dependent manner. A significant efflux of potassium (0.54�mol) per mg of CDW was induced by using nisin at a con-centration (0.1 �g/ml) slightly in excess of the MIC (0.06 �g/ml). When a higher concentration of nisin (0.5 �g/ml) wasapplied, the efflux reached a maximum of 1.4 �mol/mg CDW,
indicating a total intracellular potassium efflux. In contrast,nisin1–28 induced less than 40% efflux (0.5 �mol/mg CDW) ofthe intracellular potassium even when the applied peptidereached 100 �g/ml (Fig. 8B), 16 times higher than the MIC (6�g/ml). Increasing the concentration of nisin1–28 up to 200�g/ml did not result in further-activated potassium efflux (datanot shown). Therefore, removal of the C-terminal “tail” ofnisin resulted in a remarkably decreased potency in formingpores in the membrane.
DISCUSSION
There are overall two mechanisms by which bacteria canbecome resistant to nisin: a shielding mechanism to prevent itfrom reaching the target membrane and an enzymatic inacti-vation mechanism by modification or destruction of the anti-biotics (3). All data published so far indicate that the former isthe main mechanism, with the involvement of a number ofgenes, whereas the latter involves a Dha reductase, found inseveral strains of Bacillus subsp., that reduces Dha to Ala (18).However, the present study unveiled a lactococcal tail-specificprotease, NSR, responsible for nisin resistance through pro-teolytic degradation of nisin.
It was demonstrated that the purified NSRSD inactivatednisin by hydrolytically removing its C-terminal “tail,” whichis consistent with the presence of a conserved TSPc domainat the C terminus of NSRSD. Prolonged incubation withNSRSD did not lead to further degradation of nisin beyondits cleavage into two fragments. Kinetic analysis of nisincleavage by NSRSD yielded a Km value of 290 �M, which ishigher than that for substrate cleavage by Tsp (35 to 50 �M)(21). It is probably attributed to the fact that NSRSD, unlikeTsp, lacks a PDZ domain, which facilitates binding of pro-teases with their cognate substrates (40). However, thecleavage rate of NSRSD (kcat/Km � 20.85 � 103 M1 s1)was much higher than those of Tsp (3.8 � 103 M1 s1) orchymotrypsin (4.4 � 103 M1 s1) for their substrate, avariant of Arc repressor (21). Additionally, in contrast to thepreference of Tsp for substrates with small, uncharged ornonpolar C-terminal residues (20), nisin harbors large, pos-itively charged and polar residues (His, Lys, and Ser) at theC terminus, reflecting the difference in substrate specificitybetween NSRSD and Tsp.
Western blot analysis showed that NSR localized specificallyon the cell membrane of native and nsr recombinant lactococ-cal strains. It is conceivable that the TSPc domain of NSR
FIG. 7. Antibacterial activity of nisin against L. lactis MG1363 afterincubation with lactococcal sonicates. Con, nisin (1 �g) incubated in 50mM Tris-HCl (pH 6.0); 1, 3, and 5, sonicates of L. lactis MG1363/pMG36e, L. lactis MG1363/pSZ531, and L. lactis MG1363/pSZ537,respectively; 2, 4, and 6, nisin (1 �g) incubated with sonicates of L.lactis MG1363/pMG36e, L. lactis MG1363/pSZ531, and L. lactisMG1363/pSZ537, respectively.
TABLE 3. Inhibition of bacterial growth by nisin Z
Indicator strain Expressedproteina
Nisin degradingactivity (%)b
MIC(�g/ml)
L. lactis MG1363 None 0 0.06L. lactis MG1363/pMG36e None 0 0.06L. lactis MG1363/pSZ537 NSRSD 8 0.07L. lactis MG1363/pTS50 NSR 89 4L. lactis MG1363/pSZ531 NSR 100 5L. lactis TS1640 NSR 89 10
a None, neither NSR nor NSRSD was expressed in the strain.b Nisin degrading activity is expressed in relative activity determined by com-
paring the amount of digested products quantified by RP-HPLC with that of L.lactis MG1363/pSZ531, which is normalized to 100%.
VOL. 53, 2009 PROTEOLYTIC MECHANISM FOR NISIN RESISTANCE 1971
at INS
TIT
UT
E O
F M
ICR
OB
IOLO
GY
CA
S on A
pril 25, 2009 aac.asm
.orgD
ownloaded from

resides on the extracellular side of the membrane, where itcould contact and degrade nisin. The fact that NSRSD was alsodetected on the membrane fraction of the recombinant strainwould suggest that the predicted N-terminal signal peptideregion, if any, has no effect on protein-targeted localization.However, the evidence of NSRSD almost completely losingnisin cleavage activity in vivo (Fig. 6G) would mean that theN-terminal 27 amino acid residues of NSR are functionallynondisposable. Given that purified NSRSD could degrade ni-sin in vitro, it is speculated that the loss of the N-terminalregion would change the structure/topology of NSR, probablywith the TSPc domain of NSRSD redirecting to the cytoplas-mic side of the membrane. And when the membrane barrierwas removed, NSRSD in the lactococcal sonicates recoveredits nisin cleavage activity (Fig. 7).
It has been reported that nisin acts on a model membrane ata micromolar concentration level whereas it exerts bactericidalactivity at a nanomolar level in vivo, where lipid II is availableas a specific docking molecule for binding (4). Studies so farhave indicated that the N terminus of nisin is involved inbinding with lipid II (5, 16, 46). The removal of the C-terminalfive residues or a further nine residues from nisin to producenisin1-29 or nisin1-20, respectively, leads to a ca. 16-fold or110-fold decrease in bactericidal potency compared with thatof intact nisin (6). In this study, we found that nisin1-28, pro-duced from the cleavage of nisin by NSR, also showed a 100-fold-reduced inhibitory activity against L. lactis MG1363. Allthese data indicate that the C-terminal “tail” of nisin plays asignificant role in its antibacterial activity. Furthermore, Ser29,as the sole hydroxyl residue that escapes dehydration duringmaturation of nisin, might be vital for bactericidal activity ofnisin since its absence resulted in a marked decrease in bacte-ricidal potency. However, there are instances indicating thatunmodified Ser/Thr is not critical for biological activities ofsome other lantibiotics (8, 9). In this sense, the presence of anegatively charged carboxylate on the C terminus of MeLan28
might be detrimental for interaction between nisin1-28 and thenegatively charged membrane.
Compared with intact nisin, nisin1-28 displays a markedlydecreased affinity for the cell membrane (Fig. 6D and F) and asignificantly reduced capacity for potassium efflux induction
(Fig. 8A and B). Presently it is unknown whether the former isattributed to the decreased affinity of nisin1-28 for lipid II.Nevertheless, the latter is an indicator of its abolished pore-forming potency, which might rationalize its significantly re-duced antibacterial activity.
In short, for the first time, we provide direct evidence for anovel nisin resistance mechanism conferred by NSR: proteo-lytically cleaving the antibacterial peptide by removal of theC-terminal “tail.” Furthermore, it is worthwhile to elucidatewhether this proteolytic mechanism could be applied to theoccurrence of other structure-related lantibiotic resistancemechanisms, such as that for subtilin, though either of theircognate immunity systems provides no cross-tolerance againstthe other lantibiotic (41, 42). Further studies are still requiredto decipher the active pocket, i.e., the TSPc of NSR, and howto engineer nisin to counteract bacterial resistance.
ACKNOWLEDGMENTS
We thank Chengshu Wang (Shanghai Institutes for Biological Sci-ences, Chinese Academy of Sciences) for critical reading of the manu-script. We greatly appreciate the gift of pure nisin Z from ZhejiangSilver-Elephant Bio-engineering Co., Ltd., China.
This work was supported by grants from the National Program forHigh Technology Research and Development of China (2006AA10Z319) and the Key Project of the Chinese Academy of Sciences(KSCXZ-YW-G-016).
REFERENCES
1. Beebe, K. D., J. Shin, J. Peng, C. Chaudhury, J. Khera, and D. Pei. 2000.Substrate recognition through a PDZ domain in tail-specific protease. Bio-chemistry 39:3149–3155.
2. Bierbaum, G., and H. G. Sahl. 1985. Induction of autolysis of staphylococciby the basic peptide antibiotics Pep 5 and nisin and their influence on theactivity of autolytic enzymes. Arch. Microbiol. 141:249–254.
3. Breukink, E., and B. de Kruijff. 2006. Lipid II as a target for antibiotics. Nat.Rev. Drug Discov. 5:321–332.
4. Breukink, E., I. Wiedemann, C. van Kraaij, O. P. Kuipers, H. Sahl, and B.de Kruijff. 1999. Use of the cell wall precursor lipid II by a pore-formingpeptide antibiotic. Science 286:2361–2364.
5. Brotz, H., M. Josten, I. Wiedemann, U. Schneider, F. Gotz, G. Bierbaum,and H. G. Sahl. 1998. Role of lipid-bound peptidoglycan precursors in theformation of pores by nisin, epidermin and other lantibiotics. Mol. Micro-biol. 30:317–327.
6. Chan, W. C., M. Leyland, J. Clark, H. M. Dodd, L. Y. Lian, M. J. Gasson,B. W. Bycroft, and G. C. Roberts. 1996. Structure-activity relationships in thepeptide antibiotic nisin: antibacterial activity of fragments of nisin. FEBSLett. 390:129–132.
FIG. 8. Potassium efflux assays of L. lactis MG1363 induced by nisin (A) or nisin1–28 (B). The presented values represent the means � standarddeviations of three independent determinations.
1972 SUN ET AL. ANTIMICROB. AGENTS CHEMOTHER.
at INS
TIT
UT
E O
F M
ICR
OB
IOLO
GY
CA
S on A
pril 25, 2009 aac.asm
.orgD
ownloaded from

7. Chatterjee, C., M. Paul, L. Xie, and W. A. van der Donk. 2005. Biosynthesisand mode of action of lantibiotics. Chem. Rev. 105:633–684.
8. Cooper, L. E., A. L. McClerren, A. Chary, and W. A. van der Donk. 2008.Structure-activity relationship studies of the two-component lantibiotic halo-duracin. Chem. Biol. 15:1035–1045.
9. Cotter, P. D., L. H. Deegan, E. M. Lawton, L. A. Draper, P. M. O’Connor, C.Hill, and R. P. Ross. 2006. Complete alanine scanning of the two-componentlantibiotic lacticin 3147: generating a blueprint for rational drug design. Mol.Microbiol. 62:735–747.
10. Froseth, B. R., and L. L. McKay. 1991. Molecular characterization of thenisin resistance region of Lactococcus lactis subsp. lactis biovar diacetylactisDRC3. Appl. Environ. Microbiol. 57:804–811.
11. Gasson, M. J. 1983. Plasmid complements of Streptococcus lactis NCDO 712and other lactic streptococci after protoplast-induced curing. J. Bacteriol.154:1–9.
12. Gravesen, A., K. Sorensen, F. M. Aarestrup, and S. Knochel. 2001. Sponta-neous nisin-resistant Listeria monocytogenes mutants with increased expres-sion of a putative penicillin-binding protein and their sensitivity to variousantibiotics. Microb. Drug Resist. 7:127–135.
13. Gut, I. M., A. M. Prouty, J. D. Ballard, W. A. van der Donk, and S. R. Blanke.2008. Inhibition of Bacillus anthracis spore outgrowth by nisin. Antimicrob.Agents Chemother. 52:4281–4288.
14. Hasper, H. E., N. E. Kramer, J. L. Smith, J. D. Hillman, C. Zachariah, O. P.Kuipers, B. de Kruijff, and E. Breukink. 2006. An alternative bactericidalmechanism of action for lantibiotic peptides that target lipid II. Science313:1636–1637.
15. Holo, H., and I. F. Nes. 1989. High-frequency transformation, by electropo-ration, of Lactococcus lactis subsp. cremoris grown with glycine in osmoticallystabilized media. Appl. Environ. Microbiol. 55:3119–3123.
16. Hsu, S. T., E. Breukink, E. Tischenko, M. A. Lutters, B. de Kruijff, R.Kaptein, A. M. Bonvin, and N. A. van Nuland. 2004. The nisin-lipid IIcomplex reveals a pyrophosphate cage that provides a blueprint for novelantibiotics. Nat. Struct. Mol. Biol. 11:963–967.
17. Jarvis, B. 1967. Resistance to nisin and production of nisin-inactivatingenzymes by several Bacillus species. J. Gen. Microbiol. 47:33–48.
18. Jarvis, B., and J. Farr. 1971. Partial purification, specificity and mechanismof action of the nisin-inactivating enzyme from Bacillus cereus. Biochim.Biophys. Acta 227:232–240.
19. Keiler, K. C., and R. T. Sauer. 1995. Identification of active site residues ofthe Tsp protease. J. Biol. Chem. 270:28864–28868.
20. Keiler, K. C., and R. T. Sauer. 1996. Sequence determinants of C-terminalsubstrate recognition by the Tsp protease. J. Biol. Chem. 271:2589–2593.
21. Keiler, K. C., K. R. Silber, K. M. Downard, I. A. Papayannopoulos, K.Biemann, and R. T. Sauer. 1995. C-terminal specific protein degradation:activity and substrate specificity of the Tsp protease. Protein Sci. 4:1507–1515.
22. Kramer, N. E., E. J. Smid, J. Kok, B. de Kruijff, O. P. Kuipers, and E.Breukink. 2004. Resistance of Gram-positive bacteria to nisin is not deter-mined by lipid II levels. FEMS Microbiol. Lett. 239:157–161.
23. Kramer, N. E., S. A. van Hijum, J. Knol, J. Kok, and O. P. Kuipers. 2006.Transcriptome analysis reveals mechanisms by which Lactococcus lactisacquires nisin resistance. Antimicrob. Agents Chemother. 50:1753–1761.
24. Kuipers, O. P., H. S. Rollema, W. M. Yap, H. J. Boot, R. J. Siezen, and W. M.de Vos. 1992. Engineering dehydrated amino acid residues in the antimicro-bial peptide nisin. J. Biol. Chem. 267:24340–24346.
25. Lad, S. P., J. Li, J. da Silva Correia, Q. Pan, S. Gadwal, R. J. Ulevitch, andE. Li. 2007. Cleavage of p65/RelA of the NF-kappaB pathway by Chlamydia.Proc. Natl. Acad. Sci. USA 104:2933–2938.
26. Lad, S. P., G. Yang, D. A. Scott, G. Wang, P. Nair, J. Mathison, V. S. Reddy,and E. Li. 2007. Chlamydial CT441 is a PDZ domain-containing tail-specificprotease that interferes with the NF- B pathway of immune response. J.Bacteriol. 189:6619–6625.
27. Liao, D. I., J. Qian, D. A. Chisholm, D. B. Jordan, and B. A. Diner. 2000.Crystal structures of the photosystem II D1 C-terminal processing protease.Nat. Struct. Biol. 7:749–753.
28. Liu, C. Q., M. L. Harvey, and N. W. Dunn. 1997. Cloning of a gene encodingnisin resistance from Lactococcus lactis subsp. lactis M189 which is tran-scribed from an extended 210 promoter. J. Gen. Appl. Microbiol. 43:67–73.
29. Liu, C. Q., P. Su, N. Khunajakr, Y. M. Deng, S. Sumual, W. S. Kim, J. E.
Tandianus, and N. W. Dunn. 2005. Development of food-grade cloning andexpression vectors for Lactococcus lactis. J. Appl. Microbiol. 98:127–135.
30. Lubelski, J., R. Rink, R. Khusainov, G. N. Moll, and O. P. Kuipers. 2008.Biosynthesis, immunity, regulation, mode of action and engineering of themodel lantibiotic nisin. Cell Mol. Life Sci. 65:455–476.
31. Mantovani, H. C., and J. B. Russell. 2001. Nisin resistance of Streptococcusbovis. Appl. Environ. Microbiol. 67:808–813.
32. Montville, T. J., T. De Siano, A. Nock, S. Padhi, and D. Wade. 2006. Inhi-bition of Bacillus anthracis and potential surrogate bacilli growth from sporeinocula by nisin and other antimicrobial peptides. J. Food Prot. 69:2529–2533.
33. O’Driscoll, J., F. Glynn, G. F. Fitzgerald, and D. van Sinderen. 2006. Se-quence analysis of the lactococcal plasmid pNP40: a mobile replicon forcoping with environmental hazards. J. Bacteriol. 188:6629–6639.
34. O’Sullivan, D. J., and T. R. Klaenhammer. 1993. Rapid mini-prep isolationof high-quality plasmid DNA from Lactococcus and Lactobacillus spp. Appl.Environ. Microbiol. 59:2730–2733.
35. Oelmuller, R., R. G. Herrmann, and H. B. Pakrasi. 1996. Molecular studiesof CtpA, the carboxyl-terminal processing protease for the D1 protein of thephotosystem II reaction center in higher plants. J. Biol. Chem. 271:21848–21852.
36. Peschel, A., M. Otto, R. W. Jack, H. Kalbacher, G. Jung, and F. Gotz. 1999.Inactivation of the dlt operon in Staphylococcus aureus confers sensitivity todefensins, protegrins, and other antimicrobial peptides. J. Biol. Chem. 274:8405–8410.
37. Piard, J. C., I. Hautefort, V. A. Fischetti, S. D. Ehrlich, M. Fons, and A.Gruss. 1997. Cell wall anchoring of the Streptococcus pyogenes M6 protein invarious lactic acid bacteria. J. Bacteriol. 179:3068–3072.
38. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, NY.
39. Silber, K. R., K. C. Keiler, and R. T. Sauer. 1992. Tsp: a tail-specific proteasethat selectively degrades proteins with nonpolar C termini. Proc. Natl. Acad.Sci. USA 89:295–299.
40. Spiers, A., H. K. Lamb, S. Cocklin, K. A. Wheeler, J. Budworth, A. L. Dodds,M. J. Pallen, D. J. Maskell, I. G. Charles, and A. R. Hawkins. 2002. PDZdomains facilitate binding of high temperature requirement protease A(HtrA) and tail-specific protease (Tsp) to heterologous substrates throughrecognition of the small stable RNA A (ssrA)-encoded peptide. J. Biol.Chem. 277:39443–39449.
41. Stein, T., S. Heinzmann, S. Dusterhus, S. Borchert, and K. D. Entian. 2005.Expression and functional analysis of the subtilin immunity genes spaIFEGin the subtilin-sensitive host Bacillus subtilis MO1099. J. Bacteriol. 187:822–828.
42. Stein, T., S. Heinzmann, I. Solovieva, and K. D. Entian. 2003. Functionof Lactococcus lactis nisin immunity genes nisI and nisFEG after coordi-nated expression in the surrogate host Bacillus subtilis. J. Biol. Chem.278:89–94.
43. Tang, S., X. Chen, W. Yang, M. Chen, and L. Huan. 2001. Isolation andcharacterization of a plasmid pTS50, which encodes nisin resistance de-terminant in Lactococcus lactis TS1640. Wei Sheng Wu Xue Bao 41:536–541.
44. Terzaghi, B. E., and W. E. Sandine. 1975. Improved medium for lacticstreptococci and their bacteriophages. Appl. Microbiol. 29:807–813.
45. van de Guchte, M., J. M. van der Vossen, J. Kok, and G. Venema. 1989.Construction of a lactococcal expression vector: expression of hen egg whitelysozyme in Lactococcus lactis subsp. lactis. Appl. Environ. Microbiol. 55:224–228.
46. Wiedemann, I., E. Breukink, C. van Kraaij, O. P. Kuipers, G. Bierbaum, B.de Kruijff, and H. G. Sahl. 2001. Specific binding of nisin to the peptidogly-can precursor lipid II combines pore formation and inhibition of cell wallbiosynthesis for potent antibiotic activity. J. Biol. Chem. 276:1772–1779.
47. Xiao, H., X. Chen, M. Chen, S. Tang, X. Zhao, and L. Huan. 2004. BovicinHJ50, a novel lantibiotic produced by Streptococcus bovis HJ50. Microbiol-ogy 150:103–108.
48. Yun, X. X., J. Hu, and Q. Chen. 2006. Screening, isolation and identificationof nisin resistance determinant gene in strains of Lactococcus lactis. NanFang Yi Ke Da Xue Xue Bao 26:839–842.
VOL. 53, 2009 PROTEOLYTIC MECHANISM FOR NISIN RESISTANCE 1973
at INS
TIT
UT
E O
F M
ICR
OB
IOLO
GY
CA
S on A
pril 25, 2009 aac.asm
.orgD
ownloaded from