nitrogen mineralization and soil fertility in the dry chaco ecosystem (argentina)
TRANSCRIPT

Nitrogen Mineralization and Soil Fertility in the Dry Chaco Ecosystem (Argentina)M. J. Mazzarino,* L. Oliva, A. Nunez, G. Nunez, and E. Buffa
ABSTRACTGrassland productivity of Dry Chaco ecosystems in Argentina is
often decreased by the invasion of shrubs (Larrea spp.). To betterunderstand the reasons for this degradation and how to reverse oravoid it, we studied soil fertility and N-mineralization rates underfour vegetation types predominant in the region: trees, shrubs, in-terspaces between trees and shrubs, and pure grassland. Two typesof trees were studied: leguminous (Prosopis flexuosa DC) and non-leguminous (Aspidosperma quebracho-blanco Schlecht.). Net N-min-eralization rates were significantly higher under Prosopis (ISO mgN kg-' yr1) than under Larrea (95 mg N kg-1 yr1), the interspaces(66 mg N kg"1 yr1) and the pure grassland (68 mg N kg"1 yr1).Mineralization rates under Aspidosperma (115 mg N kg ' yr1) werenot significantly different from Prosopis. Nitrogen produced in thewet season (spring-summer months) represented 90% of the accu-mulated N for 1 yr. Nitrogen mineralization was positively correlated(P < 0.05) with both soil moisture and temperature. Nitrates com-prised the highest proportion of mineralized N, with a trend to someNHj accumulation in the dry season. At the 0- to 10-cm depth,organic C and total N were significantly higher under Prosopis thanunder Aspidosperma, Larrea, or the interspaces. At deeper soildepths, organic C and extractable P were higher under and betweenLarrea and under the pure grassland than under and between treecanopies. Results indicate that the highest soil fertility and N-min-eralization rates were under leguminous trees and the lowest in theinterspaces and under Larrea.
THE EXPANSION of beef production into semiaridand arid regions of Argentina has lead to in-
creased clear-cutting of natural forests to enhance de-velopment of pure grasslands. This practice hasresulted in a loss of grassland productivity followedby shrub invasion in many areas (Anderson, 1980;Karlin and Diaz, 1984).
Some authors (Felker, 1979; Oldeman, 1983; Karlinand Diaz, 1984; Miehe, 1986) indicate a need for anagroforestry management of arid regions with partic-ular emphasis on leguminous trees, like Acacia spp.or Prosopis spp. It has been suggested that these speciesare phreatophitic and capable of fixing N, which couldfacilitate the establishment of grasses under their can-opies. This hypothesis is based on the fact that themain determinants of soil fertility in arid regions arewater and N (Tiedemann and Klemmedson, 1973;Charley and West, 1977; Rundel et al., 1982; Bern-hard-Reversat, 1982; Virginia, 1986).
It is possible, however, that the effects attributed toleguminous trees are not necessarily specific to them,M.J. Mazzarino, Nitrogen Fixing Trees Project, P.O. Box 2, TropicalAgronomic Center for Research and Training (CATIE), Turrialba,Costa Rica; L. Oliva and E. Buffa, Soil Microbiology, Fac. of Ag-ricultural Sciences, Univ. of Cordoba, P.O. Box 509, 5000 Cordoba,Argentina; and A. Nuflez and G. Nunez, Analytical Chemistry, Fac.of Agronomy, Univ. of Buenos Aires, Av. San Martin 4453, 1417Buenos Aires, Argentina. Contribution from the Fac. of Agronomy,Univ. of Buenos Aires, the Fac. of Agricultural Sciences, Univ. ofCordoba, and the National Council of Research and Technology(CONICET), Argentina. Received 21 Aug. 1989. 'Correspondingauthor.
Published in Soil Sci. Soc. Am. J. 55:515-522 (1991).
but due to a tree effect common to all tree species. Atthe same time, the observed shrub invasion, domi-nated by Larrea spp., could lead to a recovery in soilfertility, since nutrients have been reported to accu-mulate under this shrub (Garcia Moya and McKell,1970; Cox et al., 1984).
To help clarify some of these issues, a study wasconducted to quantify and compare soil fertility andN mineralization under: (i) two dominant trees of thenatural forest, one leguminous (Prosopis) and one non-leguminous (Aspidosperma), (ii) a shrub (Larrea spp.)apparently dominant in overgrazed pastures, (iii) theinterspaces between trees and shrubs, and (iv) a puregrassland of perennial grasses.
MATERIALS AND METHODSTwo adjacent study areas were sampled in the Los Pocitos
Forest Reserve, west of Sierra de Pocho, near Chancani,Province of Cordoba, Argentina. This site has been describedas a Dry Chaco plain with high summer temperatures andmoderate winters. Mean annual precipitation ranges be-tween 300 and 500 mm and is concentrated in the summer.The potential evapotranspiration index is < —20 (Karlinand Diaz, 1984). During our study, precipitation was aboveaverage, reaching 790 mm, but always concentrated in thesummer. The soils of the site are generally Entisols of alluvialorigin.
The areas studied were (i) a natural forest where Aspidos-perma and Prosopis are the dominant trees, (ii) an area in-vaded by Larrea for approximately 15 yr, and (iii) an areakept clear of shrubs and trees for 15 yr and dominated byperennial grasses (Papophorum spp.). None of the sites hadbeen grazed for a period of at least 10 yr. Soils in the firsttwo areas are classified as Ustorthents, those of the third siteas Ustifluvents. The Ustifluvents have a finer texture thanthe Orthents, which translates to higher cation-exchange ca-pacity (CEC) and water-holding capacity. Sampling was con-ducted under three plants of each species and at threerandomly selected points in the pure grassland. Samples be-neath tree and shrubs were taken about 50 cm from the treebase; the interspace samples were taken about 1 to 1.5 mfrom the canopy border of each tree or shrub. Aspidospermatrees had a single stem with a basal girth (BG) of 60 to 100cm and an extended vertical canopy. Prosopis trees forkedat the base, had a BG of 80 to 100 cm, and an extendedhorizontal canopy.
Two sampling sequences were carried out: (i) in September1986 soil samples were collected from the 0- to 10-, 10- to25-, 25- to 45-, and 45- to 75-cm depths for chemical andphysico-chemical characterization; and (ii) monthly betweenJune 1987 and June 1988 surface soil was collected for Nmineralization. Sampling was done using a modified Eno(1960) technique. Tin cans, 13 cm high and 7 cm in diameter,which were sealed with a polyethylene film, were insertedinto the soil and incubated in situ for 28 d. An adjacent soilcore was collected at the same depth, from which two sub-samples were removed: a 10-g subsample was used to es-tablish the initial concentrations of inorganic N (N at ta) anda 50- to 100-g sample for determining soil water content.Subsamples of 10 g were also taken in July (dry season) andJanuary (wet season) for in vitro mineralization assays. Theywere incubated for 28 d under controlled temperature (20-25 °C in July and 25-30 °C in January) and moisture con-
515
516 SOIL SCI. SOC. AM. J., VOL. 55, MARCH-APRIL 1991
ditions (field capacity in July and at the field moisture of tain January). Water losses were determined gravimetricallyeach week and adjusted with distilled water.
Soil samples (10 g) were extracted in 100 mL of 2 MKC1,left to equilibrate for 12 h, and then filtered. Extraction ofin situ incubation samples was done in the field within 3 hof collection. Extracts were analyzed colorimetrically by theindofenol blue method for NH4-N (Keeney and Nelson,1982) and by the Griess-Ilosvay method for NO3-N (Kamp-shake et al., 1967). Net N mineralization was estimated asthe final amount of NH4-N plus NO3-N after the 28-d in-cubation (tu) minus the initial amount of NH4-N plus NO3-N (t0). A detailed description of the N-mineralization meth-od is presented in Matson et al. (1987).
Soil samples were analyzed for pH in a soil/water sus-pension having a 1:2.5 volumetric ratio, electrical conduc-tivity in a saturated paste, organic C by the wet method ofWalicley and Black, total N by the micro-Kjeldahl technique,extractable P by NaHCO3 extraction (Olsen and Dean,1965), and CEC for samples having a pH >7.5 with car-bonates present by the NaOAc-NH4OAc method and insamples with a pH <7.5 by summing exchangeable basesobtained by an NH4OAc extraction. Samples analyzed forpH, electrical conductivity, CEC, and extractable P were airdried and ground to pass through a 2-mm sieve, while thoseanalyzed for organic C and total N were further ground topass through a 0.2-mm sieve.
Data were statistically analyzed using the SPSS/PC com-puter program (Norusis, 1984). Analysis of variance (A-NOVA) and Duncan analysis of significance of P < 0.05
were conducted for the following treatments: Prosopis (underand between canopies), Aspidosperma (under and betweencanopies), Larrea (under and between canopies), and puregrassland. Means were compared among treatments at thesame depth or among depths for each treatment. Correlationcoefficients were also calculated to evaluate soil moisture andtemperature effects on N mineralization and NO3-N pools.
RESULTS AND DISCUSSIONSoil Properties
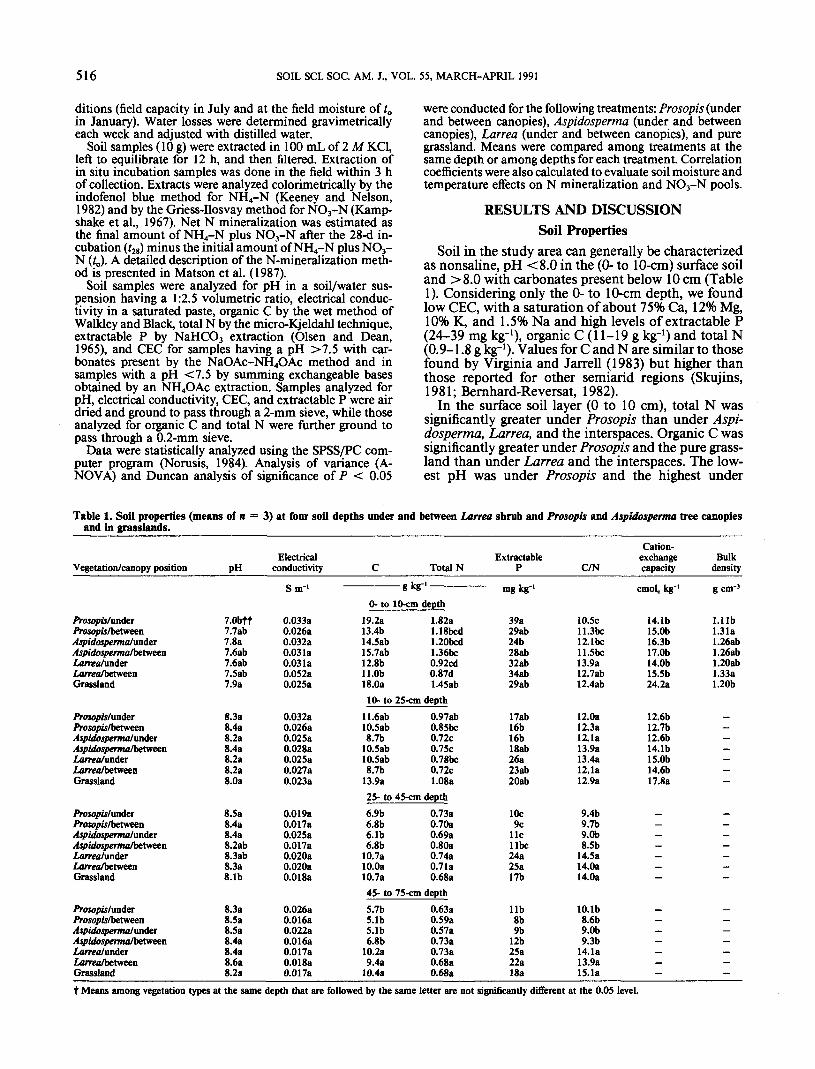
Soil in the study area can generally be characterizedas nonsaline, pH <8.0 in the (0- to 10-cm) surface soiland >8.0 with carbonates present below 10 cm (Table1). Considering only the 0- to 10-cm depth, we foundlow CEC, with a saturation of about 75% Ca, 12% Mg,10% K, and 1.5% Na and high levels of extractable P(24-39 mg kg-1), organic C (11-19 g kg-1) and total N(0.9-1.8 g kg-1). Values for C and N are similar to thosefound by Virginia and Jarrell (1983) but higher thanthose reported for other semiarid regions (Skujins,1981; Bernhard-Reversat, 1982).
In the surface soil layer (0 to 10 cm), total N wassignificantly greater under Prosopis than under Aspi-dosperma, Larrea, and the interspaces. Organic C wassignificantly greater under Prosopis and the pure grass-land than under Larrea and the interspaces. The low-est pH was under Prosopis and the highest under
Table 1. Soil properties (means of nand in grasslands.
3) at four soil depths under and between Larrea shrub and Prosopis and Aspidosperma tree canopies
Vegetation/canopy position PHElectrical
conductivityExtractable
C Total N P C/N
Cation-exchangecapacity
Bulkdensity
Prosopis/underProsopis/betweenAspidosperma/underAspidosperma/betv/eenLarrea/underLarrea/betweenGrassland
Prosopis/underProsopis/betweenAspidosperma/underAspidosperma/betvteenLarrea/underLarrea/betv/eenGrassland
7.0btt7.7ab7.8a7.6ab7.6ab7.5ab7.9a
8.3a8.4a8.2a8.4a8.2a8.2a8.0a
Sm-'
0.033a0.026a0.032a0.031a0.031a0.052a0.025a
0.032a0.026a0.02Sa0.028a0.025a0.027a0.023a
0- to 10-cm depth19.2a13.4b14.5ab15.7ab12.8b11. Ob18.0a
1.82al.lgbcd1.20bcd1.36bc0.92cd0.87d1.45ab
10- to 25-cm depth11.6ablO.Sab8.7blO.SablO.Sab8.7b13.9a
0.97ab0.8Sbc0.72c0.75c0.78bc0.72cl.OSa
mgkg-'
39a29ab24b28ab32ab34ab29ab
17ab16b16b18ab26a23ab20ab
25- to 4S-cm depth
10.5c11.3bc12.1bcll.Sbc13.9a12.7ab12.4ab
12.0a12.3a12.1a13.9a13.4a12.1a12.9a
cmol,; kg-'
14.1bIS.Ob16.3b17.0b14.0b15.5b24.2a
12.6b12.7b12.6b14.1bIS.Ob14.6b17.8a
g cm-
l.llb1.31a1.26ab1.26ab1.20ab1.33a1.20b
Prosopis/underProsopis/betvteenAspidosperma/underAspidosperma/betv/eenLarrea/underLarrea/betv/eenGrassland
8.5a8.4a8.4a8.2ab8.3ab8.3a8.1b
0.019a0.017a0.025a0.017a0.020a0.020aO.OlSa
6.9b6.8b6.1b6.8b
10.7alO.Oa10.7a
0.73a0.70a0.69aO.SOa0.74a0.71a0.68a
lOc9c
liellbc24a25a17b
9.4b9.7b9.0b8.5b
14.5a14.0a -14.0a
-—————-
45- to 75-cm depthProsopis/underProsopis/betv/eenAspidosperma/underAspidosperma/betv/eenLarrea/underLarrea/betv/eenGrassland
8.3a8.5a8.5a8.4a8.4a8.6a8.2a
0.026a0.016a0.022a0.016a0.017aO.OlSa0.017a
5.7bS.lbS.lb6.8b
10.2a9.4a
10.4a
0.63a0.59a0.57a0.73a0.73a0.68a0.68a
lib8b9b
12b25a22a18a
lO.lb8.6b9.0b9.3b
14.1a13.9aIS.la
———————
t Means among vegetation types at the same depth that are followed by the same letter are not significantly different at the O.OS level.

MAZZARINO ET AL.: NITROGEN MINERALIZATION AND FERTILITY IN DRY CHACO 517
Aspidosperma and the pure grassland. Lowest concen-trations of P were found under Aspidosperma and theinterspaces but, overall, they were quite high, sug-gesting that this nutrient is not limiting in thisecosystem.
At lower soil depths (>25 cm) extractable P andorganic C were significantly higher under and betweenLarrea and in the pure grassland than under and be-tween tree canopies.
A comparison of soil properties among depths forthe same vegetation type indicated that concentrationsof organic C, total N, and extractable P and CEC werehigher in the 0- to 10-cm surface soil and declinedsharply under trees (by about 40-45%) at depths of 10to 25 cm (Fig. 1). A more gradual decline of thesenutrients occurred in the interspaces between trees.These findings are consistent with other observationsfor semiarid and arid regions (Tiedemann andKlemmedson, 1973; Skujins and West, 1974; Bern-hard-Reversat, 1982; Virginia and Jarrell, 1983). Un-
LARREA10 20
40
60
u 20
40
60
; -jbo
bo,
• ,a1 1f ifb
>b
b
ORGANICCARBON(g kg")
40
PROSOPIS10 _ap
601 b]u
20
40
601
o~c
Ab b
b Jib-
TOTALNITROGEN(g kg")
20
40
40 60
EXTRACTABLEPHOSPHORUSt mg kg'") ~
20
40
a, ib
ob
40
bc.'c
60L c *c
Fig. 1. Organic C, total N, and extractable P distributions with depthunder (dashed line) and between (solid line) the canopies of Pro-sopis trees and Lama shrubs. Values (means of « = 3) werecompared among depths for each vegetation type. Common lettersindicate means are not significantly different at P < 0.05.
der and outside Larrea canopies, the decline was evenless. In the pure grassland, nutrients were highthroughout the entire profile.
The high organic-C and extractable-P levels at thedeeper soil depths under and between Larrea and inthe pure grassland may be caused by the vertical move-ment of organic matter, which is low in N and highin organic P, and which affects extractable P (Hang,1988, personal communication). The organic matterin these soils may be dominated by substances hayinglow molecular weight and high mobility. Organic Passociated with this type of organic matter is, there-fore, more susceptible to deep leaching than is N(Schoenau and Bettany, 1987). Alternatively, thetransfer of nutrients by trees (extractable P) from deep-er soil layers either from below their canopies or fromadjacent openings (Charley and Cowling, 1968; Tie-demann and Klemmedson, 1973; Virginia and Jarrell,1983) and organic-matter decomposition at depth bythe action of tree root exudates may explain the lowernutrient levels with increased depth. In contrast, grass-es and shrubs, because of their more surficial root dis-tribution, would recycle nutrients mainly from surfacesoil (Oldeman, 1983; Karlin and Diaz, 1984), thereforeleaving lower depth nutrients unaffected.
Nitrogen MineralizationEffect of Soil Moisture and Temperature
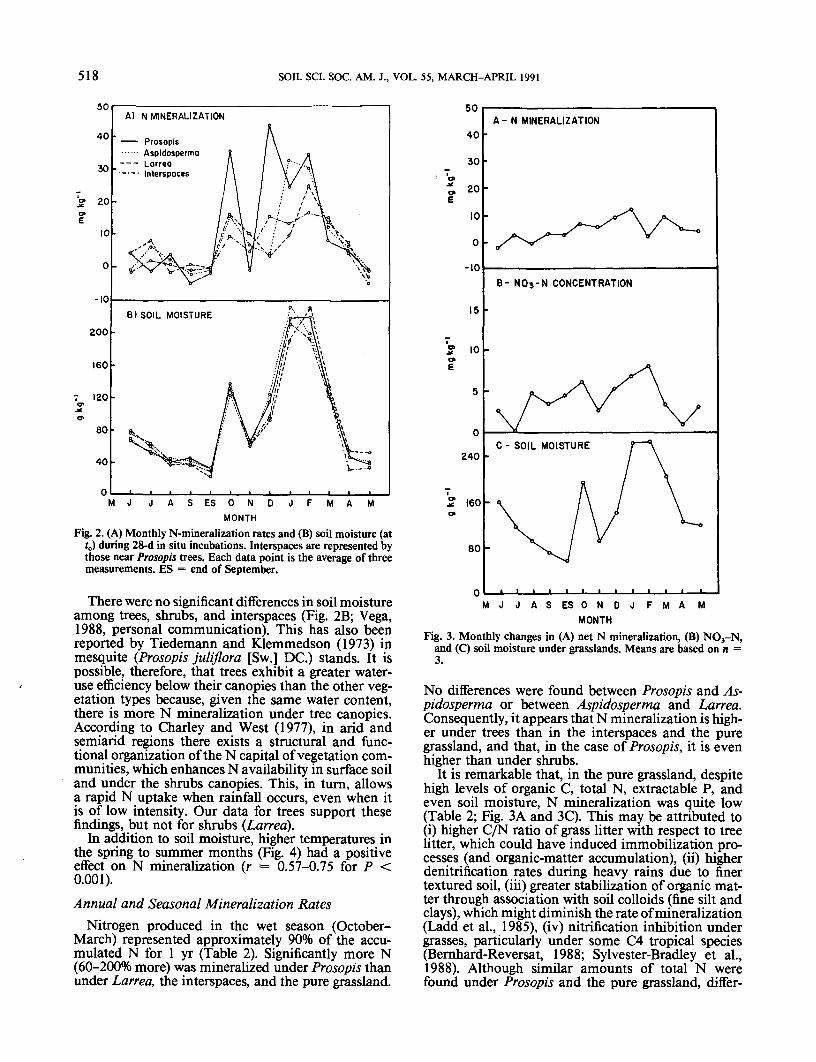
Nitrogen mineralization was similar among the dif-ferent vegetation types, including the grasslands, dur-ing the dry months (April through September) and wasbelow 10 mg kg"1 (Fig. 2A and 3A) when soil moisturewas lowest (Fig. 2B and 3C). During these dry periods,negative mineralization rates occurred, which corre-spond to a net immobilization described by Raison etal. (1987). After the beginning of the wet season, Nmineralization increased rapidly and peaked twice(Fig. 2A) in synchronization with high soil moisture(Fig. 2B). Other authors have reported that N min-eralization is high at the beginning of wet seasons insemiarid savannas, but decreases before the wet seasonends because mineralizable N has been depleted (Bern-hard-Reversat, 1977, 1982; Singh and Balasubraman-ian, 1980). In our study, however, N mineralizationwas strongly correlated with soil moisture. Significantcorrelations were found between net N mineralizationand soil moisture: r = 0.60 to 0.75, P < 0.001 fortrees, shrubs and interspaces; r = 0.40, P < 0.05 forthe pure grassland. The trends in N mineralization donot reflect the effect of total monthly rainfall (Fig. 4),but instead the proximity of the rainfall event to thesampling date. For example, although total precipi-tation in October and November were similar (Fig. 4),N mineralization was much more rapid in Octoberthan in November (Fig. 2A). This probably occurredbecause the October sample was taken 2 d after a rainwhereas the November sample was taken 11 d after arain when the soil was much drier. At the onset of thewet season, N-mineralization peaks were significantlyhighest under Prosopis alone. Towards the end of thewet season, however, both Prosopis and Aspidospermaexhibited similar (nonsignificantly different) N-min-eralization peaks. The lowest rates were observed un-der Larrea, the interspaces, and in the pure grassland.
518
50
40
30
£ 20o>
10
0
-10
SOIL SCI. SOC. AM. J.:
200 •
160
7 120o>jf0>
80
40
A) N MINERALIZATION
—— Prosopis• • • • • • Aspidospermu--- Larreo— • - • Interspaces
B) SOIL MOISTURE
VOL. 55, MARCH-APRIL 1991
M J A S ES M A M0 N D JMONTH
Fig. 2. (A) Monthly N-mineralization rates and (B) soil moisture (atO during 28-d in situ incubations. Interspaces are represented bythose near Prosopis trees. Each data point is the average of threemeasurements. ES = end of September.
There were no significant differences in soil moistureamong trees, shrubs, and interspaces (Fig. 2B; Vega,1988, personal communication). This has also beenreported by Tiedemann and Klemmedson (1973) inmesquite (Prosopis juliflora [Sw.] DC.) stands. It ispossible, therefore, that trees exhibit a greater water-use efficiency below their canopies than the other veg-etation types because, given the same water content,there is more N mineralization under tree canopies.According to Charley and West (1977), in arid andsemiarid regions there exists a structural and func-tional organization of the N capital of vegetation com-munities, which enhances N availability in surface soiland under the shrubs canopies. This, in turn, allowsa rapid N uptake when rainfall occurs, even when itis of low intensity. Our data for trees support thesefindings, but not for shrubs (Larrea).
In addition to soil moisture, higher temperatures inthe spring to summer months (Fig. 4) had a positiveeffect on N mineralization (r = 0.57-0.75 for P <0.001).Annual and Seasonal Mineralization Rates
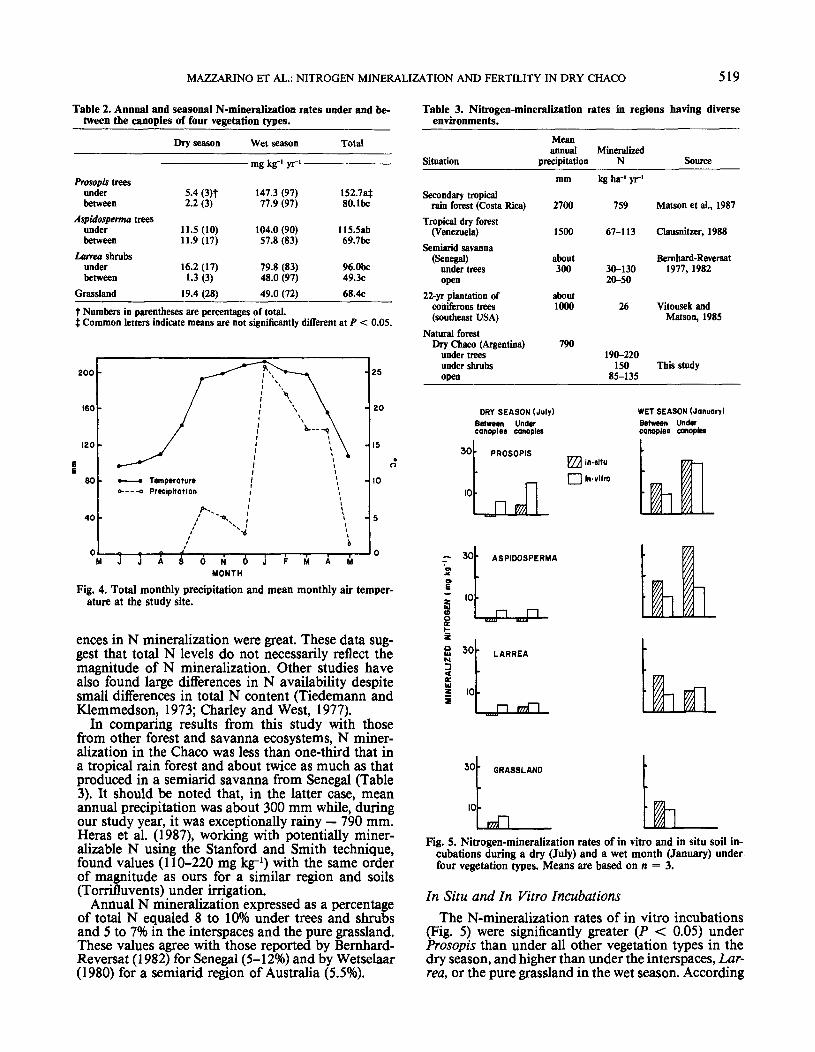
Nitrogen produced in the wet season (October-March) represented approximately 90% of the accu-mulated N for 1 yr (Table 2). Significantly more N(60-200% more) was mineralized under Prosopis thanunder Larrea, the interspaces, and the pure grassland.
is
10
0
240
160
80
B- N03-N CONCENTRATION
C - SOIL MOISTURE
M J J A S ES 0 N DMONTH
J F M A M
Fig. 3. Monthly changes in (A) net N mineralization, (B) NO3-N,and (C) soil moisture under grasslands. Means are based on n —3.
No differences were found between Prosopis and As-pidosperma or between Aspidosperma and Larrea.Consequently, it appears that N mineralization is high-er under trees than in the interspaces and the puregrassland, and that, in the case of Prosopis, it is evenhigher than under shrubs.
It is remarkable that, in the pure grassland, despitehigh levels of organic C, total N, extractable P, andeven soil moisture, N mineralization was quite low(Table 2; Fig. 3A and 3C). This may be attributed to(i) higher C/N ratio of grass litter with respect to treelitter, which could have induced immobilization pro-cesses (and organic-matter accumulation), (ii) higherdenitrification rates during heavy rains due to finertextured soil, (iii) greater stabilization of organic mat-ter through association with soil colloids (fine silt andclays), which might diminish the rate of mineralization(Ladd et al., 1985), (iv) nitrification inhibition undergrasses, particularly under some C4 tropical species(Bernhard-Reversat, 1988; Sylvester-Bradley et al.,1988). Although similar amounts of total N werefound under Prosopis and the pure grassland, differ-
MAZZARINO ET AL.: NITROGEN MINERALIZATION AND FERTILITY IN DRY CHACO 519
Table 2. Annual and seasonal N-mineralization rates under and be-tween the canopies of four vegetation types.
Dry season Wet season Total
- mg kg'1 yr1 -Prosopis trees
underbetween
Aspidosperma treesunderbetween
Larrea shrubsunderbetween
Grassland
5.4 (3)f2.2 (3)
11.5(10)11.9(17)
16.2 (17)1.3 (3)
19.4 (28)
147.3 (97)77.9 (97)
104.0 (90)57.8 (83)
79.8 (83)48.0 (97)49.0 (72)
152.7at80.1 be
115.5ab69.7bc
96.0bc49.3c68.4c
f Numbers in parentheses are percentages of total.f Common letters indicate means are not significantly different at P < 0.05.
200
M J
Fig. 4. Total monthly precipitation and mean monthly air temper-ature at the study site.
ences in N mineralization were great. These data sug-gest that total N levels dp not necessarily reflect themagnitude of N mineralization. Other studies havealso found large differences in N availability despitesmall differences in total N content (Tiedemann andKlemmedson, 1973; Charley and West, 1977).
In comparing results from this study with thosefrom other forest and savanna ecosystems, N miner-alization in the Chaco was less than one-third that ina tropical rain forest and about twice as much as thatproduced in a semiarid savanna from Senegal (Table3). It should be noted that, in the latter case, meanannual precipitation was about 300 mm while, duringour study year, it was exceptionally rainy — 790 mm.Heras et al. (1987), working with potentially miner-alizable N using the Stanford and Smith technique,found values (110-220 mg kg-1) with the same orderof magnitude as ours for a similar region and soils(Torrifluvents) under irrigation.
Annual N mineralization expressed as a percentageof total N equaled 8 to 10% under trees and shrubsand 5 to 7% in the interspaces and the pure grassland.These values agree with those reported by Bernhard-Reversat (1982) for Senegal (5-12%) and by Wetselaar(1980) for a semiarid region of Australia (5.5%).
Table 3. Nitrogen-mineralization rates in regions having diverseenvironments.
Situation
Meanannual Mineralized
precipitation N Source
Secondary tropicalrain forest (Costa Rica) 2700
Tropical dry forest(Venezuela) 1500
Semiarid savanna(Senegal) about
under trees 300open
22-yr plantation of aboutconiferous trees 1000(southeast USA)
Natural forestDry Chaco (Argentina) 790
under treesunder shrubsopen
kg haM yr1
759 Matson et al., 1987
67-113 Clausnitzer, 1988
30-13020-50
26
Bemhard-Reversat1977, 1982
Vitousek andMatson, 1985
190-220150 This study
85-135
DRY SEASON (July) WET SEASON (January)Betweencanopies
30
10
Under Between Undercanopies canopies canopies
• PROSOPIS
n
K/y| in-situ
[[] In-vltro
m \ •fln\,
~ 30
3I^ 10
ASPIDOSPERMA Ig 30N
sz 10
LARREA
30 GRASSLAND
Fig. 5. Nitrogen-mineralization rates of in vitro and in situ soil in-cubations during a dry (July) and a wet month (January) underfour vegetation types. Means are based on n = 3.
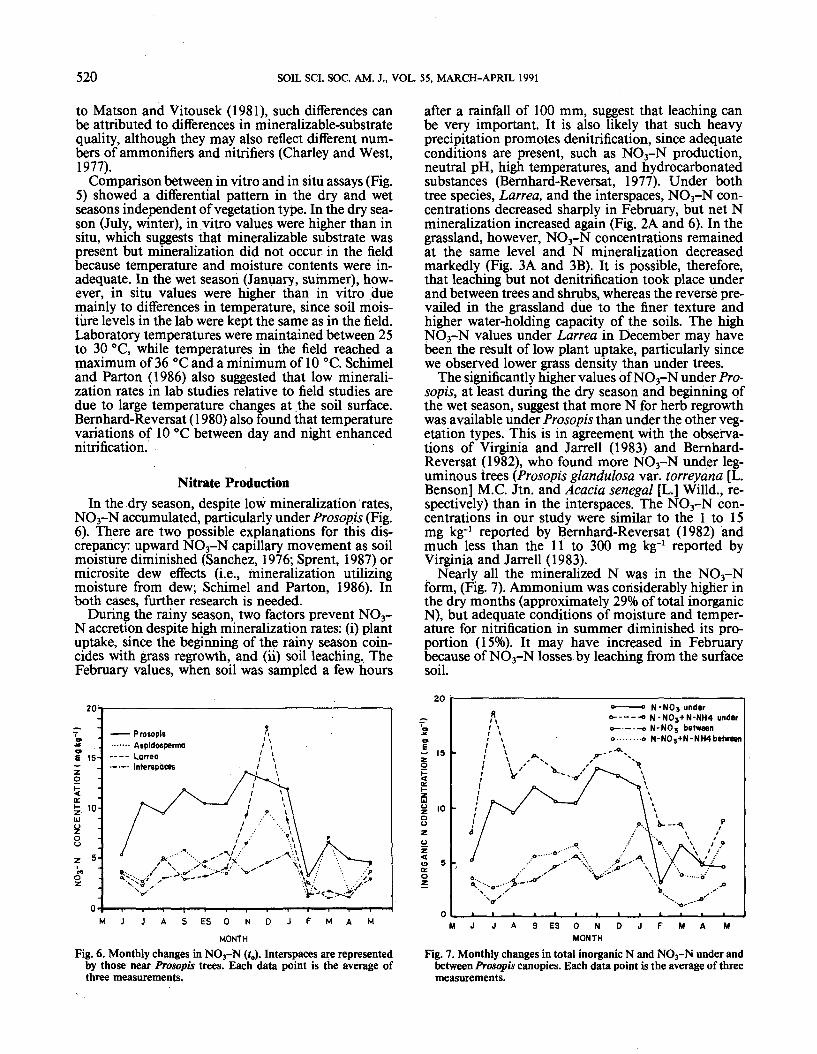
In Situ and In Vitro IncubationsThe N-mineralization rates of in vitro incubations
(Fig. 5) were significantly greater (P < 0.05) underProsopis than under all other vegetation types in thedry season, and higher than under the interspaces, Lar-rea, or the pure grassland in the wet season. According
520 SOIL SCI. SOC. AM. J., VOL. 55, MARCH-APRIL 1991
to Matson and Vitousek (1981), such differences canbe attributed to differences in mineralizable-substratequality, although they may also reflect different num-bers of ammonifiers and nitrifiers (Charley and West,1977).
Comparison between in vitro and in situ assays (Fig.5) showed a differential pattern in the dry and wetseasons independent of vegetation type. In the dry sea-son (July, winter), in vitro values were higher than insitu, which suggests that mineralizable substrate waspresent but mineralization did not occur, in the fieldbecause temperature and moisture contents were in-adequate. In the wet season (January, summer), how-ever, in situ values were higher than in vitro duemainly to differences in temperature, since soil mois-ture levels in the lab were kept the same as in the field.Laboratory temperatures were maintained between 25to 30 °C, while temperatures in the field reached amaximum of 36 °C and a minimum of 10 °C. Schimeland Parton (1986) also suggested that low minerali-zation rates in lab studies relative to field studies aredue to large temperature changes at the soil surface.Bernhard-Reversat (1980) also found that temperaturevariations of 10 °C between day and night enhancednitrification.
Nitrate ProductionIn the dry season, despite low mineralization rates,
NO3-N accumulated, particularly under Prosopis (Fig.6). There are two possible explanations for this dis-crepancy: upward NO3-N capillary movement as soilmoisture diminished (Sanchez, 1976; Sprent, 1987) ormicrosite dew effects (i.e., mineralization utilizingmoisture from dew; Schimel and Parton, 1986). Inboth cases, further research is needed.
During the rainy season, two factors prevent NO3-N accretion despite high mineralization rates: (i) plantuptake, since the beginning of the rainy season coin-cides with grass regrowth, and (ii) soil leaching. TheFebruary values, when soil was sampled a few hours
after a rainfall of 100 mm, suggest that leaching canbe very important. It is also likely that such heavyprecipitation promotes denitrification, since adequateconditions are present, such as NO3-N production,neutral pH, high temperatures, and hydrocarbonatedsubstances (Bernhard-Reversat, 1977). Under bothtree species, Larrea, and the interspaces, NO3-N con-centrations decreased sharply in February, but net Nmineralization increased again (Fig. 2A and 6). In thegrassland, however, NO3-N concentrations remainedat the same level and N mineralization decreasedmarkedly (Fig. 3A and 3B). It is possible, therefore,that leaching but not denitrification took place underand between trees and shrubs, whereas the reverse pre-vailed in the grassland due to the finer texture andhigher water-holding capacity of the soils. The highNO3-N values under Larrea in December may havebeen the result of low plant uptake, particularly sincewe observed lower grass density than under trees.
The significantly higher values of NO3^N under Pro-sopis, at least during the dry season and beginning ofthe wet season, suggest that more N for herb regrowthwas available under Prosopis than under the other veg-etation types. This is in agreement with the observa-tions of Virginia and Jarrell (1983) and Bernhard-Reversat (1982), who found more NO3-N under leg-uminous trees (Prosopis glandulosa var. torreyana [L.Benspn] M.C. Jtn. and Acacia Senegal [L.] Willd., re-spectively) than in the interspaces. The NO3-N con-centrations in our study were similar to the 1 to 15mg kg"1 reported by Bernhard-Reversat (1982) andmuch less than the 11 to 300 mg kg'1 reported byVirginia and Jarrell (1983).
Nearly all the mineralized N was in the NO3-Nform, (Fig. 7). Ammonium was considerably higher inthe dry months (approximately 29% of total inorganicN), but adequate conditions of moisture and temper-ature for nitrification in summer diminished its pro-portion (15%). It may have increased in Februarybecause of NO3-N losses by leaching from the surfacesoil.
20
E 15-zo
oz
10
5-
—— Prosopis• • • • • • Aspldosperma--- Larrea•— Interspaces
M J J A S E S O N D J F M A M
MONTH
Fig. 6. Monthly changes in NO3-N (Q. Interspaces are representedby those near Prosopis trees. Each data point is the average ofthree measurements.
20
15
10
o 5a:O
N-N03 underN-N03+N-NH4 underN-N03 betweenN-N03+N-NH4between
M J J A S E S O N D J F M A MMONTH
Fig. 7. Monthly changes in total inorganic N and NO3-N under andbetween Prosopis canopies. Each data point is the average of threemeasurements.

MAZZARINO ET AL: NITROGEN MINERALIZATION AND FERTILITY IN DRY CHACO 521
CONCLUSIONSIn summary, the vegetation types studied affected
N dynamics differently, despite similar soil water con-tents. Trees transferred nutrients from deeper in thesoil profile and from lateral interspaces (P) and ac-cumulated organic C and total N in the surface soil.A large fraction of this total N is easily mineralizable,producing higher inorganic N than found for the othervegetation types. Despite high N-mineralization rates,total N pools remained high, implying N accumula-tion. Leguminous trees seemed to be more effectivethan nonleguminous ones in maintaining N availa-bility, since higher total N and inorganic N concen-trations were found under Prosopis than underAspidosperma. In addition, Prosopis experienced twosignificant monthly peaks in N mineralization. Theseevents could influence substantially N availability forassociated pasture species.
The tree interspaces showed greatest soil impover-ishment: total-N, mineralizable-N, and inorganic-Nproduction were very low. Interspace nutrient levelsdeeper in the soil profile were also low, suggesting ef-fective competition by tree roots for lateral nutrientpools.
The pure grassland showed high values of organicC and total N and P throughout the entire profile. Suchaccretion is probably a consequence of low minerali-zation rates (low mineralizable-N pool) in the surface,and vertical movement of organic matter. High P val-ues at greater soil depths are related to either surficialroot distribution or to organic-matter movement.
The contribution of Larrea to soil fertility was min-imal. Organic C and total N and P were low in thesurface, in addition to a small mineralizable N pooland low mineralization rates. However, deeper soilshowed accretion of organic C and P. Two possibleexplanations include vertical movement of organicmatter, and invasion by Larrea of overgrazed grass-lands with surface soils already impoverished butoverlying a relatively nutrient-rich subsoil.
From a livestock-management perspective, grazingpure grasslands translates to continual nutrient export,resulting, eventually, in impoverished soil fertility. Ifthe grassland is left to Larrea invasion, such impov-erishment will be sustained for at least the first 15 yr,as was the case in this study.
In the case of pasture with leguminous trees, the N-availability scenario may be quite different. We wouldexpect that, because of high-N-content residues, min-eralization rates should remain high even under graz-ing. This, in turn, should sustain inorganic-N poolsmore effectively than grasslands without trees. Al-though such systems appear promising, we still needto investigate the contributions to total N accumula-tion from both the tree and grass components undergrazing. We also need to explore possible solutions tonegative effects in tree interspaces resulting from tree/pasture competition for nutrients.
ACKNOWLEDGMENTSWe wish to thank O. Bachmeier, O.T. Karlin, and C. Ca-
rranza for the discussions in the field and Chacho, Jose, and
Sandra Gonzalez for assisting in sample collection. We alsothank Leonard DeBano, Joann Roskoski, Leslie Cooper-band, and Tomas Schlichter for their helpful comments ondrafts of this manuscript. Financial support for this researchwas provided by the U.S. National Academy of Sciences/National Research Council by means of a grant from theU.S. Agency for International Development to the Catedrade Manejo de Areas Marginales (Facultad de Ciencias Agro-pecuarias, Universidad Nacional de Cordoba) directed byO.T. Karlin.

522 SOIL SCI. SOC. AM. J., VOL. 55, MARCH-APRIL 1991