new sporulation loci in streptomyces coelicolor a3(2)jb.asm.org/content/181/17/5419.full.pdf · new...
TRANSCRIPT

JOURNAL OF BACTERIOLOGY,0021-9193/99/$04.0010
Sept. 1999, p. 5419–5425 Vol. 181, No. 17
Copyright © 1999, American Society for Microbiology. All Rights Reserved.
New Sporulation Loci in Streptomyces coelicolor A3(2)N. JAMIE RYDING,† MAUREEN J. BIBB, VIRGINIE MOLLE, KIM C. FINDLAY, KEITH F. CHATER,
AND MARK J. BUTTNER*
John Innes Centre, Colney, Norwich NR4 7UH, United Kingdom
Received 1 April 1999/Accepted 24 June 1999
Sporulation mutants of Streptomyces coelicolor appear white because they are defective in the synthesis of thegrey polyketide spore pigment, and such white (whi) mutants had been used to define eight sporulation loci,whiA, whiB, whiD, whiE, whiG, whiH, whiI, and whiJ (K. F. Chater, J. Gen. Microbiol. 72:9–28, 1972; N. J.Ryding, Ph.D. thesis, University of East Anglia, 1995). In an attempt to identify new whi loci, we mutagenizedS. coelicolor M145 spores with nitrosoguanidine and identified 770 mutants with colonies ranging from whiteto medium grey. After excluding unstable strains, we examined the isolates by phase-contrast microscopy andchose 115 whi mutants with clear morphological phenotypes for further study. To exclude mutants representingcloned whi genes, self-transmissible SCP2*-derived plasmids carrying whiA, whiB, whiG, whiH, or whiJ (but notwhiD, whiE, or whiI) were introduced into each mutant by conjugation, and strains in which the wild-typephenotype was restored either partially or completely by any of these plasmids were excluded from furtheranalysis. In an attempt to complement some of the remaining 31 whi mutants, an SCP2* library of wild-typeS. coelicolor chromosomal DNA was introduced into 19 of the mutants by conjugation. Clones restoring thewild-type phenotype to 12 of the 19 strains were isolated and found to represent five distinct loci, designatedwhiK, whiL, whiM, whiN, and whiO. Each of the five loci was located on the ordered cosmid library: whiL, whiM,whiN, and whiO occupied positions distinct from previously cloned whi genes; whiK was located on the samecosmid overlap as whiD, but the two loci were shown by complementation to be distinct. The phenotypesresulting from mutations at each of these new loci are described.
In the filamentous, gram-positive bacterium Streptomycescoelicolor, dispersal is achieved through a simple differentia-tion process that results in the release of exospores (7). Duringthis process, multigenomic aerial hyphae divide into unig-enomic prespore compartments by synchronized multiple sep-tation at regular intervals along their length. These cylindricalprespore compartments subsequently mature to give rise tochains of 50 to 100 ovoid, thick-walled spores. During thismaturation phase, colonies develop a grey color, due to thesynthesis of a polyketide spore pigment.
This pigmentation has been exploited to isolate develop-mental mutants; all of the original sporulation-deficient mu-tants were identified by virtue of their inability to synthesizewild-type levels of the spore pigment, resulting in colonies thatremained white, even on prolonged incubation (18). Fifty ofthese white (whi) mutants were analyzed genetically, and cur-rent information suggests that they represent eight separateloci, whiA, whiB, whiD, whiE, whiG, whiH, whiI, and whiJ (6, 9,31). Of these, whiE is a complex locus which encodes theenzymes that synthesize the spore pigment itself, and whiEmutants do not appear to be morphologically defective (6, 12,25). whiA, whiB, and whiG mutants are completely blocked insporulation septation and are white in appearance. whiH andwhiI mutants produce some sporulation septa; while whiI mu-tants are completely white, whiH mutants are pale grey andmake low but detectable levels of the whiE transcripts thatspecify the spore pigment (6, 7, 9, 22, 25, 31, 32). whiJ mutantsproduce low numbers of apparently normal spore chains, arepale grey, and again make low but detectable levels of the whiE
transcripts (22, 31). Mutation in the remaining locus, whiD,causes the formation of spores of highly irregular size andshape which are defective in wall thickening and lyse exten-sively (6, 25, 28). Two other loci, whiC and whiF, are no longerincluded: the only known whiC mutant has been lost, andwhiF99 was shown to be an unusual allele of whiG (5, 31). Theeight known whi loci have all been cloned and sequenced (1,11–13, 27, 31, 32).
Chater (6) suggested that the genetic map of whi loci mightnot be saturated on the grounds that several loci are poorlyrepresented in the original collection of whi mutants analyzed;examples are whiD (one allele), whiB (two alleles), whiE (twoalleles) and whiJ (two alleles). The subsequent discovery byreverse genetics of the involvement of the sigF locus in sporu-lation supported this viewpoint (21, 22, 29). As a consequence,we have attempted to identify novel whi loci through the iso-lation of new whi mutants and their complementation, takingadvantage of previously cloned whi genes to exclude knownloci from the screen. We report the identification, phenotypiccharacterization, mapping, and cloning of five new whi loci.
MATERIALS AND METHODS
Bacterial strains, plasmids, growth conditions, protoplast transformation,and chemical mutagenesis. S. coelicolor A3(2) strains used were the wild-typestrain 1147 (prototrophic, SCP11 SCP21 Pgl1 [19]), M145 (prototrophic, SCP12
SCP22 Pgl1 [19]), J1501 (hisA1 uraA1 strA1 SCP12 SCP22 Pgl2 [10]), 1258(proA1 hisC9 argA1 cysD18 uraA1 strA1 SCP1NF [SCP2 status uncertain] Pgl1
[19]), and J243 (uraA1 strA1 whiD16 SCP11 [SCP2 status uncertain] Pgl1 [8]). S.coelicolor strains were cultured on minimal medium MM (19) containing 0.5%(wt/vol) mannitol as the carbon source or on MS (mannitol plus soya flour) agar(17). Protoplasts were made and transformed as described by Hopwood et al.(19). S. coelicolor M145 spores were mutagenized with nitrosoguanidine (NTG)as described previously (14, 18). Plasmids used were pIJ698 (23) and pSET152(4).
Construction of a genomic library and complementation of mutants. Totalchromosomal DNA from wild-type S. coelicolor was partially digested withSau3AI and size fractionated on a sucrose gradient, and fragments in the sizerange 15 to 22 kb were treated with calf intestinal alkaline phosphatase andligated with the self-transmissible, single-copy SCP2*-derived plasmid, pIJ698,
* Corresponding author. Mailing address: John Innes Centre, Col-ney, Norwich NR4 7UH, United Kingdom. Phone (44) 1603 452571.Fax: (44) 1603 456844. E-mail [email protected].
† Present address: Department of Microbiology, Michigan StateUniversity, East Lansing, MI 48824-1101.
5419
on May 19, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

cut with BglII. The ligation mix was introduced into the histidine and uracilauxotroph S. coelicolor J1501 by protoplast transformation, and 4,000 of theresulting transformants were arrayed on master plates (MM containing histidine,uracil, and 50 mg of hygromycin per ml), with 100 patches per plate.
For preparation of an inoculum for mating, developmental mutants weregrown on cellophane discs on MM agar, scraped into 3 ml of 10.3% (wt/vol)sucrose, and homogenized in a manual glass homogenizer. This suspension wasdiluted with 10.3% (wt/vol) sucrose to 9 ml of final volume, and 200 ml wasspread and dried on each mating plate (MM containing histidine and uracil).Genomic library master plates were replicated onto the mating plates, and afterincubation to allow mating and growth, exconjugants of the developmental mu-tant carrying the plasmid were selected by replica plating to MM lacking auxo-trophic supplements but containing 50 mg of hygromycin per ml.
Complementation of mutants with known whi genes. Self-transmissibleSCP2*-derived plasmids carrying whiA (pIJ6204 [31]), whiB (pIJ2157 [13]), whiG(pIJ597 [31]), whiH (pIJ6201 [32]), or whiJ (pIJ6205 [31]) were transferred byconjugation from S. coelicolor J1501 into new whi mutants by replica plating andsubsequent counterselection of the auxotrophic donor strain as described for thegenomic library.
Conjugation from Escherichia coli into Streptomyces developmental mutants.Because many whi mutants do not sporulate, the method of Flett et al. (16) wasadapted to promote conjugal transfer from E. coli into Streptomyces by usingmycelial fragments, rather than spores, as an inoculum; high frequencies ofexconjugants were obtained without difficulty. pSET152 and its derivatives wereintroduced by transformation or electroporation into the dam dcm hsdS E. colistrain ET12567 containing the RK2 derivative pUZ8002 (35). pUZ8002 suppliestransfer functions to oriT-carrying plasmids, such as pSET152, but is not effi-ciently transferred itself because of a mutation in its own oriT. E. coli containingpSET152 or its derivatives was grown in L broth to an A600 of 0.4, washed twicewith an equal volume of fresh medium, and resuspended in 1/10 the volume ofL broth. Four-day-old lawns of the whi mutants, grown on MS agar, wereharvested by pipetting 3 to 4 ml of 20% (vol/vol) glycerol onto the surface andgently dislodging the aerial mycelium with a sterile loop. The resulting suspen-sion of mycelial fragments was vortexed for 1 min, and 0.5 ml was mixed with 0.5ml of washed E. coli cells. After harvesting by centrifugation, the pellet, contain-ing Streptomyces mycelial fragments and E. coli cells, was resuspended in theresidual medium and plated on MS agar containing 10 mM MgCl2. Followingincubation for 16 to 20 h at 30°C, each plate was overlaid with 1 ml of watercontaining 0.5 mg of nalidixic acid (to kill E. coli) and 1 mg of apramycin (toselect Streptomyces exconjugants).
Physical mapping of cloned DNA. Inserts from complementing clones were gelisolated, 32P radiolabelled by random priming, and used to probe Qiabranemembranes on which the entire minimal, ordered cosmid library of Redenbachet al. (30) had been arrayed.
Genetic mapping of whiL mutants. A suspension of mycelial fragments wasprepared from each of the mutants R214, R349, and R491 in the same way as theinoculum for the library matings. Each suspension was mixed with spores of S.coelicolor 1258 and plated on appropriately supplemented MM. After incubationto allow mating and growth, serial dilutions of harvested mycelium and sporeswere plated on MM containing uracil, proline, arginine, cystine, and 10 mg ofstreptomycin per ml, selecting for His1 streptomycin-resistant recombinants.One hundred recombinants from each cross were arrayed on master plates of thesame medium, incubated for several days, and then replicated to MM lacking oneof the auxotrophic supplements (uracil, proline, arginine, or cystine). Recombi-nants were scored for the whiL phenotype by phase-contrast microscopy and forgrowth on each medium.
Scanning electron microscopy. For scanning electron microscopy, colonieswere mounted on the surface of an aluminum stub with O.C.T. compound (BDHLaboratory Supplies, Poole, United Kingdom), plunged into liquid nitrogen slushat approximately 2210°C to cryopreserve the material, and transferred to thecryostage of a CT1500HF cryotransfer system (Oxford Instruments, Oxford,United Kingdom) attached to a Philips XL30 FEG scanning electron microscope(Philips Electron Optics, FEI UK Ltd., Cambridge, United Kingdom). Surfacefrost was sublimated at 295°C for 3 min before sputter coating the sample withplatinum for 2 min at 10 mA, at below 2110°C. Finally, the sample was movedonto the cryostage in the main chamber of the microscope, held at approximately2140°C, and viewed at 1.2 to 5.0kV. Photographs were taken with Ilford FP4 120roll film in a Linhof camera.
RESULTS
Isolation of new whi mutants. S. coelicolor M145 spores weremutagenized with NTG and plated directly on MM at a densityof approximately 200 colonies per plate; 1,500 mutant colonieswith aerial surface color varying from white to medium greywere picked from a total population of approximately 30,000.From the 1,500 patches, 770 strains were streaked for singlecolonies, and those that showed unstable phenotypes (approx-imately one-third of the total) were discarded. A large majority
of the unstable mutants exhibited a hypervariable phenotypesimilar to that described for strains of Streptomyces ambofa-ciens that have undergone large chromosomal rearrangements(24, 34). Coverslip impressions of the remaining strains wereexamined, and 431 isolates with stable phenotypes distinguish-able from that of the wild type were identified. These strainscould be divided into three groups: 71 that failed to produceabundant aerial hyphae, 144 that produced abundant aerialhyphae but showed clear defects in sporulation; and 216 mu-tants that sporulated abundantly but produced either sporeswith aberrant size or shape or spores of normal appearance butwith reduced pigmentation. Primary interest was in the secondgroup (144 strains).
Complementation with known whi genes. In an attempt toexclude mutants that represented whi genes that had alreadybeen cloned, self-transmissible SCP2*-derived plasmids carry-ing whiA, whiB, whiG, whiH, or whiJ (but not whiD, whiE, orwhiI) were introduced into 115 of the 144 whi mutants bymating. The morphological defects shown by the mutantsmade it unlikely that they were caused by mutations in thewhiE spore pigment biosynthesis cluster, and self-transmissibleclones were not available for whiD or whiI. Complementa-tion was judged initially by looking for the restoration of greypigmentation to the aerial surface of exconjugants and subse-quently by inspection of coverslip impressions in a phase-con-trast microscope. Of the 115 strains, 6 were fully comple-mented by whiA, 15 were fully complemented by whiB, 19 werefully complemented by whiG, 16 were fully complemented bywhiH, and 3 were fully complemented by whiJ. A striking ob-servation was that in two of the strains complemented by whiB,five of the strains complemented by whiG, and seven of thestrains complemented by whiH, one or more of the other whigenes also increased the level of grey pigmentation. In somecases, there was also some degree of morphological correctionas well. In a further 25 strains, no fully complementing whigene was identified, but one or more of the five whi genesincreased the level of grey pigmentation.
All strains in which the wild-type phenotype was restored,either partially or completely, by whiA, whiB, whiG, whiH, orwhiJ were excluded from further analysis. This left 31 whimutants.
Complementation and phenotypes of novel whi mutants.Attempts were made to complement 19 of the remaining 31whi mutants. An SCP2* library of chromosomal DNA isolatedfrom wild-type S. coelicolor was constructed in the auxotrophicstrain J1501. Four thousand clones from this library were ar-rayed on master plates and mated with each mutant in turn byreplica plating, and transconjugants were isolated by a secondround of replica plating to a selective medium as described inMaterials and Methods. Potentially complementing cloneswere identified initially by looking for grey patches and subse-quently by inspection of coverslip impressions in a phase-con-trast microscope. In each case, we confirmed plasmid linkageof the phenotype by reintroduction of the cloned DNA into thecorresponding whi mutant(s), either by protoplast transforma-tion using the primary SCP2* clone itself or by mating from E.coli, having first subcloned the insert from SCP2* into theintergeneric, conjugative vector pSET152, which integrates sitespecifically into the S. coelicolor chromosome at the phagefC31 attB site (4). In all, complementing clones were found for12 of the 19 strains, and the mutations were allocated to fiveloci, according to their complementation groups. A summaryof information on these 12 strains is given in Table 1.
whiK. One clone, pIJ6700, complemented three of the newwhi mutants, R273, R318, and R655. Microscopic examinationshowed that the plasmid restored wild-type levels of sporula-
5420 RYDING ET AL. J. BACTERIOL.
on May 19, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

tion to all three strains. Hybridization, using the insert frompIJ6700 as a probe, localized the complementing DNA to theoverlap between cosmids 6G4 and D63, in the 7 o’clock regionof the chromosome (Fig. 1). whiD also maps to this overlap(27, 30), but further analysis showed that whiD did not com-plement R273, R318, or R655 and that whiK did not comple-ment the whiD strain J243 (26). Accordingly, the new locus wasdesignated whiK.
The phenotypes of R273 and R318 were indistinguishable.Both mutants produced white colonies with long, tightly coiledaerial hyphae with frequent septation (Fig. 2B). In contrast,the third whiK mutant, R655, showed a more severe pheno-type; although very occasional short chains of spores weredetected, R655 produced, almost entirely, straight, undifferen-tiated aerial hyphae (Fig. 2C).
whiL. Three clones, pIJ6701, pIJ6702, and pIJ6703, comple-mented five of the new whi mutants, R139, R214, R349, R491,and R514. Southern hybridization showed that these threeclones overlapped and that they mapped to the unique regionof cosmid 4A7, in the 10 o’clock region of the chromosome(Fig. 1). This locus was designated whiL.
There was significant phenotypic variation among the whiLmutants. R139, R491, and R514 had pale grey colonies thatproduced long, curved and straight spores (Fig. 2D). R214 wasmorphologically indistinguishable from these three strains butproduced less spore pigment. The most striking phenotypeamong the whiL mutants was that of R349. R349 producedmedium grey colonies with tightly coiled aerial hyphae that hadsepta at long intervals, giving rise to corkscrew-like fragmentsof irregular length, often several times longer than a wild-typeprespore compartment (Fig. 2E). Although the cloned DNArestored the wild-type phenotype to four of the whiL strains, itdid not completely complement R349; introduction of thecloned DNA restored wild-type levels of colony pigmentationand regular septation, but the spores produced were still no-ticeably longer than those of the wild type (Fig. 2F). Onepossible explanation for this observation is that R349 carries asecond mutation that contributes to its morphological pheno-type, in addition to the mutation in whiL.
The variation in the phenotypes of the strains apparentlycomplemented by whiL, and the fact that whiL clones did notrestore a full wild-type phenotype to R349 raised the possibilitythat not all of these strains carried allelic mutations and thatsome of the effects observed might represent suppression. Inan attempt to address this question, we determined approxi-mate genetic map locations for the whi mutations carried byR214, R349, and R491, three strains with distinguishable phe-notypes, to see if the genetic map locations of these mutationswere compatible with the physical map location of the com-plementing DNA. In all three cases, the mutation mappedbetween proA and hisC but closer to proA (e.g., Fig. 3). proAhas been mapped physically to the unique region of cosmidC123 (30). Cosmid 4A7, which carries whiL, lies between proAand hisC, 10 cosmids distant from proA and 18 cosmids distantfrom hisC. The approximate genetic map locations of the mu-tations in R214, R349, and R491 are therefore consistent withthe physical map location of the DNA that complements thesestrains.
whiM. One clone, pIJ6704, complemented R432, and hybrid-ization showed that the cloned DNA mapped to the unique
FIG. 1. Locations of developmental genes on the combined physical andgenetic map of S. coelicolor. Positions of the five new whi loci are shown on theoutside of the circle, and positions of previously identified developmental genesare shown on the inside (adapted from reference 30). Sizes of the AseI fragmentsare in kilobases. }, oriC; F, telomeres.
TABLE 1. Characteristics of the novel S. coelicolor whi mutants
Locus Cosmid Allele Aerial surfacecolor Phenotype
whiK D63 R273 White Long tight coils but with septationR318 White Long tight coils but with septationR655 White Undifferentiated aerial hyphae with very rare chains of curved spores
whiL 4A7 R139 Pale grey Long curved and straight sporesR214 Very pale grey Long curved and straight sporesR349 Medium grey Short coils and long and short fragmentsR491 Pale grey Long curved and straight sporesR514 Pale grey Long curved and straight spores
whiM I51 R432 White Undifferentiated hyphae with occasional spore chains of normal appearance; aerial hyphaelyse rapidly
whiN E68 R112 White Undifferentiated hyphae with rare spore chains sometimes showing irregular septumplacement
R650 Medium grey Slightly oligosporogenous; spores longer than those of the wild typewhiO 6G4 R589 Medium grey Undifferentiated hyphae with frequent spore chains of normal appearance
VOL. 181, 1999 NEW SPORULATION LOCI IN S. COELICOLOR 5421
on May 19, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
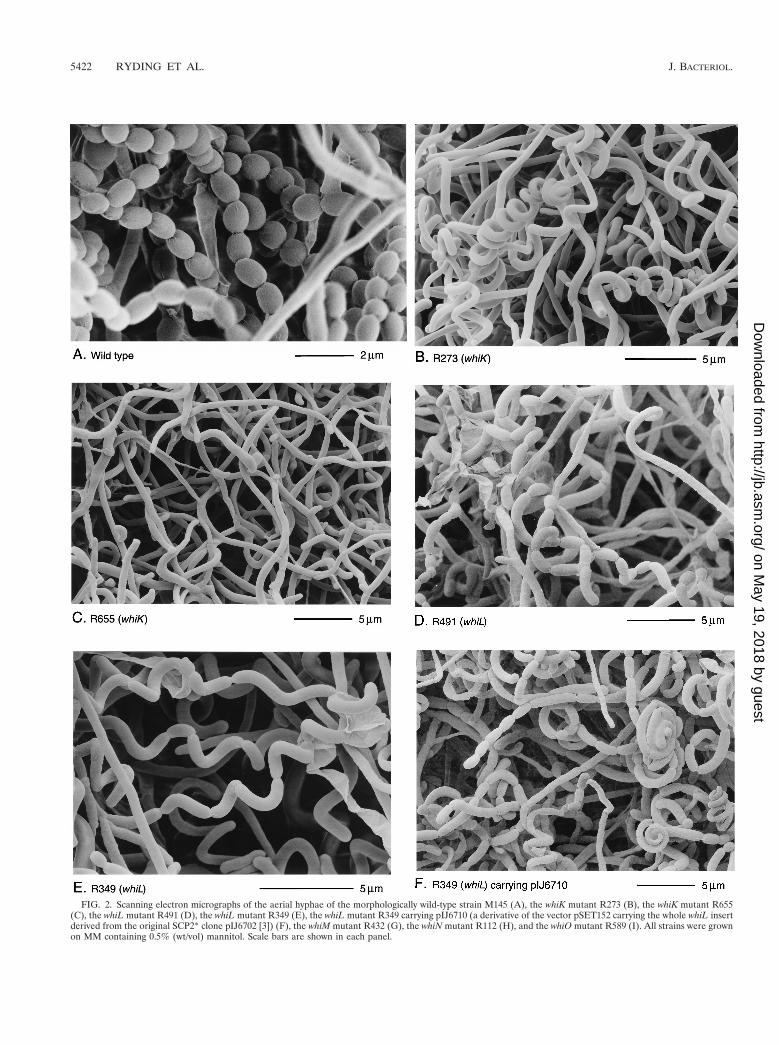
FIG. 2. Scanning electron micrographs of the aerial hyphae of the morphologically wild-type strain M145 (A), the whiK mutant R273 (B), the whiK mutant R655(C), the whiL mutant R491 (D), the whiL mutant R349 (E), the whiL mutant R349 carrying pIJ6710 (a derivative of the vector pSET152 carrying the whole whiL insertderived from the original SCP2* clone pIJ6702 [3]) (F), the whiM mutant R432 (G), the whiN mutant R112 (H), and the whiO mutant R589 (I). All strains were grownon MM containing 0.5% (wt/vol) mannitol. Scale bars are shown in each panel.
5422 RYDING ET AL. J. BACTERIOL.
on May 19, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
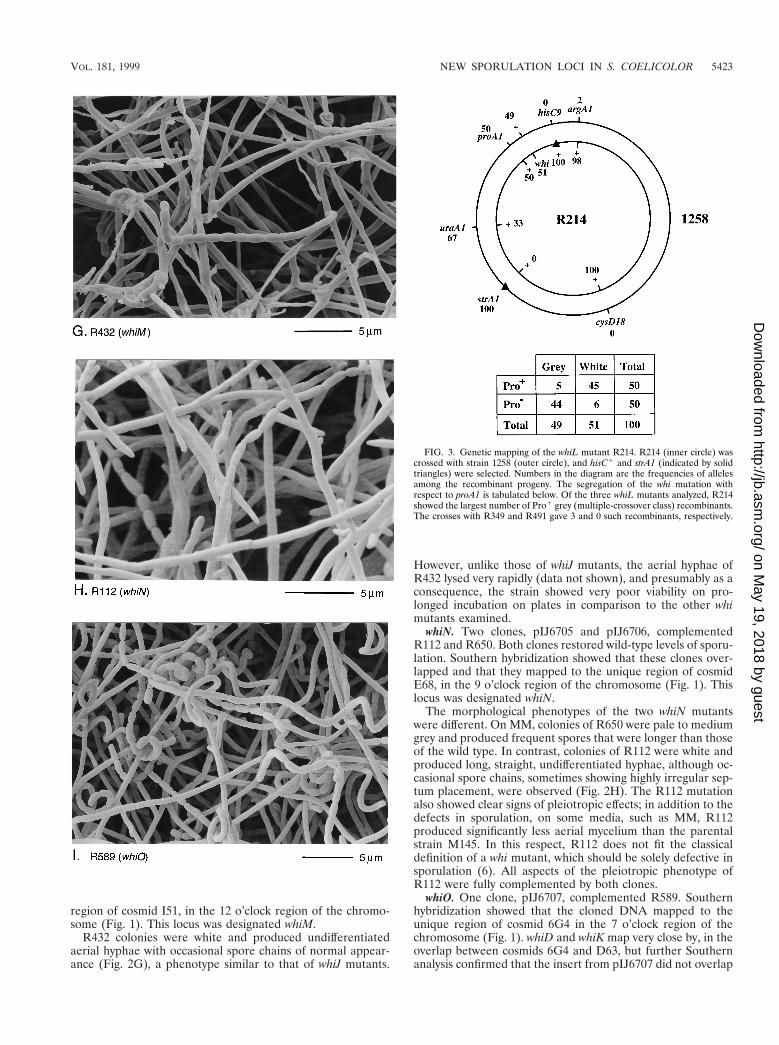
region of cosmid I51, in the 12 o’clock region of the chromo-some (Fig. 1). This locus was designated whiM.
R432 colonies were white and produced undifferentiatedaerial hyphae with occasional spore chains of normal appear-ance (Fig. 2G), a phenotype similar to that of whiJ mutants.
However, unlike those of whiJ mutants, the aerial hyphae ofR432 lysed very rapidly (data not shown), and presumably as aconsequence, the strain showed very poor viability on pro-longed incubation on plates in comparison to the other whimutants examined.
whiN. Two clones, pIJ6705 and pIJ6706, complementedR112 and R650. Both clones restored wild-type levels of sporu-lation. Southern hybridization showed that these clones over-lapped and that they mapped to the unique region of cosmidE68, in the 9 o’clock region of the chromosome (Fig. 1). Thislocus was designated whiN.
The morphological phenotypes of the two whiN mutantswere different. On MM, colonies of R650 were pale to mediumgrey and produced frequent spores that were longer than thoseof the wild type. In contrast, colonies of R112 were white andproduced long, straight, undifferentiated hyphae, although oc-casional spore chains, sometimes showing highly irregular sep-tum placement, were observed (Fig. 2H). The R112 mutationalso showed clear signs of pleiotropic effects; in addition to thedefects in sporulation, on some media, such as MM, R112produced significantly less aerial mycelium than the parentalstrain M145. In this respect, R112 does not fit the classicaldefinition of a whi mutant, which should be solely defective insporulation (6). All aspects of the pleiotropic phenotype ofR112 were fully complemented by both clones.
whiO. One clone, pIJ6707, complemented R589. Southernhybridization showed that the cloned DNA mapped to theunique region of cosmid 6G4 in the 7 o’clock region of thechromosome (Fig. 1). whiD and whiK map very close by, in theoverlap between cosmids 6G4 and D63, but further Southernanalysis confirmed that the insert from pIJ6707 did not overlap
FIG. 3. Genetic mapping of the whiL mutant R214. R214 (inner circle) wascrossed with strain 1258 (outer circle), and hisC1 and strA1 (indicated by solidtriangles) were selected. Numbers in the diagram are the frequencies of allelesamong the recombinant progeny. The segregation of the whi mutation withrespect to proA1 is tabulated below. Of the three whiL mutants analyzed, R214showed the largest number of Pro1 grey (multiple-crossover class) recombinants.The crosses with R349 and R491 gave 3 and 0 such recombinants, respectively.
VOL. 181, 1999 NEW SPORULATION LOCI IN S. COELICOLOR 5423
on May 19, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

either of these genes. Therefore, this locus was designatedwhiO.
R589 colonies were medium grey in color and had an oligo-sporogenous phenotype; by scanning electron microscopy,some undifferentiated aerial hyphae, but with fairly frequentspore chains of normal appearance, were observed (Fig. 2I).
DISCUSSION
The poor representation of several loci among the originalcollection of 50 whi mutants analyzed suggested that the ge-netic map of whi loci was not saturated (6, 31), a conclusionborne out by the results presented here. The primary criterionfor the selection of sporulation mutants in the new screenremained the loss of production of the grey spore pigment,because, in contrast to the endospores of Bacillus and Clostrid-ium, the exospores of S. coelicolor are not particularly resistantto high temperatures or chemical attack, and screens for sen-sitivity to these kinds of stresses therefore could not be used.However, we recognized that some developmental mutantscould have severe morphological defects but be only partiallyblocked in production of spore pigment (as is true for whiHand whiJ mutants), and so during the selection process weattempted to include all mutants in which spore pigmentationwas reduced to a noticeable extent. Two important variablesbetween the first and second screens whose influence cannotreadily be assessed are that (i) in the first screen (18) a richmedium was used, whereas in this screen MM was usedthroughout for the assessment of phenotype; and (ii) some ofthe mutations in the earlier screen were induced by UV light(18), whereas all the mutations in this screen were induced byNTG. Here, genetic mapping was not relied on heavily as ameans of determining the positions of mutations, because thelibrary-based complementation required less time and effortand ultimately led to the physical mapping of the new loci. Likethe old whi loci, the new loci are located in the 6-Mb centralregion of the linear S. coelicolor chromosome where virtuallyall genes identified by classical genetics have mapped (Fig. 1),in contrast to the two ;1-Mb regions at the chromosome endsthat are almost devoid of classical markers (30).
From the 770 mutants that were streaked for single colonies,431 were stable and could be distinguished from the wild typeby spore pigmentation, and the vast majority of these alsoshowed morphological defects. As in the first whi mutant anal-ysis (6), emphasis was placed on mutants showing clear mor-phological defects that could easily be distinguished from thewild type by phase-contrast microscopy, thereby making thescoring of complementation relatively straightforward. How-ever, this left as subjects for further study a large number ofmore subtle mutants that showed variation in spore size orshape. This group contained strains that showed phenotypessimilar to those of sigF (29) and whiD (6) and could be used toexpand the number of genes known to be involved in the laterstages of spore development. Both whiL (Fig. 2D and E) andwhiN (Fig. 2H) mutants showed unusual intervals betweensporulation septa. The control of septal placement in bacteriais mostly studied in rod-shaped unicellular bacteria, but furtherstudies of these Streptomyces mutants may provide distinctiveand novel insights into this process.
Of the 19 whi strains that received the library, 12 werecomplemented. The 12 mutants were grouped by complemen-tation into five loci, named whiK to whiO. The failure to com-plement the other seven strains, and the small numbers ofcomplementing clones isolated for each of the new loci (whiK,one; whiL, three; whiM, one; whiN, two; and whiO, one) indi-cated that the library was not fully representative. An attractive
alternative strategy to complement the remaining strains wouldbe to derive an approximate genetic map position for eachmutation and then to use the minimal, ordered cosmid libraryof Redenbach et al. (30) to “walk” across the correspondinginterval of the combined physical and genetic map. Althoughthese cosmids cannot replicate autonomously in S. coelicolor,selection for kanamycin resistance after protoplast transforma-tion results in the recovery of isolates in which the cosmid hasintegrated into the chromosome via insert-directed homolo-gous recombination. The cosmids can therefore be used toclone genes by complementation (30), an approach that hasbeen used to isolate whiD (27, 28), whiI (1), and bldC (20).
The new screen showed a wide variation in the number ofmutations isolated at each whi locus (whiA, 6; whiB, 15; whiG,19; whiH, 16; whiJ, 3; whiK, 3; whiL, 5; whiM, 1; whiN, 2; whiO,1), as was found in the previous screen (6). Part of this disparitycan be explained by the fact that some of the mutants (partic-ularly whiA, whiB, and whiG) yield colonies that are snowywhite and have a raised aerial mycelium, but others (especiallysome of those representing the new loci described here) pro-duce some spore pigment and so were less likely to be pickedout from a field of colonies.
Of the 115 mutants that were checked for complementationby clones of whiA, whiB, whiG, whiH, and whiJ carried onself-transmissible plasmids, 59 showed full complementationby one of the clones, 25 showed partial complementation byone or more of the clones, and 31 strains were unaffected,although many of the strains that were fully complemented byone of the existing clones were also partially complemented byanother. The 25 mutants that were only partially comple-mented by one of the five clones could, in principle, representpartially dominant alleles that do not allow full restoration ofthe wild-type phenotype, but it seems much more likely thatthey contain mutations in genes other than whiA, whiB, whiG,whiH, and whiJ which are partially suppressed by the introduc-tion of the cloned gene on the SCP2* low-copy-number plas-mid. In cases where more than one of these five genes wereable partially to restore the wild-type phenotype, there can beno doubt that suppression is involved. Suppression effects havenot previously been reported in developmental work in Strep-tomyces, although the ability of an additional copy of whiGpartially to suppress the spore pigment defect of whiH mutantswithout affecting their morphological phenotype has recentlybeen noted (15). Suppression effects caused by additional cop-ies of genes have been both informative and problematic in theanalysis of the regulation of antibiotic biosynthesis in Strepto-myces (2).
In the first genetic screen for whi loci, most of the mutantsassigned to a given locus were similar in phenotype (6), asmight be expected if most of the alleles were null or close tonull. The one notable exception was a mild allele of whiG thatcaused the formation of long spores, rather than the morecharacteristic block in sporulation septum formation, and in-deed this mutant was originally designated whiF because of thiswide phenotypic difference (5, 6, 31). In contrast, in the workdescribed here, there was wide phenotypic variation withineach of the three loci represented by multiple alleles, whiK,whiL, and whiN. This again raised the possibility that some ofthese effects might represent suppression rather than truecomplementation (although the three clones complementingwhiL mutants overlapped, as did the two clones complement-ing whiN mutants). However, in genetic mapping experimentsusing three whiL strains each having a distinct phenotype(R214, R349, and R491), the genetic map locations of thethree mutations were consistent with the physical map locationof the complementing DNA.
5424 RYDING ET AL. J. BACTERIOL.
on May 19, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

The approach taken here has enabled us simultaneously toidentify, map, and clone five new whi loci and should facilitatefurther rapid progress. Our prospects in these endeavors willundoubtedly be enhanced by the current S. coelicolor genomesequencing project (33).
ACKNOWLEDGMENTS
We thank Gabriella Kelemen, David Hopwood, and Mervyn Bibbfor helpful comments on the manuscript, Sue Bunnewell for hand-printing the scanning electron micrographs, and Tobias Kieser for helpin preparing Fig. 1.
This work was funded by BBSRC grants CAD 04380 (to K.F.C.) and83/P07658 (to M. J. Buttner), by a Lister Institute research fellowship(to M. J. Buttner), by a John Innes Foundation studentship (to V.M.),and by a grant-in-aid to the John Innes Centre from the BBSRC.
REFERENCES
1. Ainsa, J., H. D. Parry, and K. F. Chater. Unpublished data.2. Bibb, M. J. 1996. The regulation of antibiotic production in Streptomyces
coelicolor A3(2). Microbiology 142:1335–1344.3. Bibb, M. J., and M. J. Buttner. Unpublished data.4. Bierman, M., R. Logan, K. O’Brien, E. T. Seno, R. N. Rao, and B. E.
Schoner. 1992. Plasmid cloning vectors for the conjugal transfer of DNAfrom Escherichia coli to Streptomyces spp. Gene 116:43–49.
5. Bruton, C. J. Personal communication.6. Chater, K. F. 1972. A morphological and genetic mapping study of white
colony mutants of Streptomyces coelicolor. J. Gen. Microbiol. 72:9–28.7. Chater, K. F. 1998. Taking a genetic scalpel to the Streptomyces colony.
Microbiology 144:1465–1478.8. Chater, K. F. Unpublished data.9. Chater, K. F., and M. J. Merrick. 1976. Approaches to the study of differ-
entiation in Streptomyces coelicolor A3(2), p. 583–593. In K. D. MacDonald(ed.), Second International Symposium on the Genetics of Industrial Micro-Organisms. Academic Press, London, England.
10. Chater, K. F., C. J. Bruton, A. A. King, and J. E. Suarez. 1982. The expres-sion of Streptomyces and Escherichia coli drug resistance determinants clonedinto the Streptomyces phage fC31. Gene 19:21–32.
11. Chater, K. F., C. J. Bruton, K. A. Plaskitt, M. J. Buttner, C. Mendez, andJ. D. Helmann. 1989. The developmental fate of Streptomyces coelicolorhyphae depends upon a gene product homologous with the motility sigmafactor of Bacillus subtilis. Cell 59:133–143.
12. Davis, N. K., and K. F. Chater. 1990. Spore colour in Streptomyces coelicolorA3(2) involves the developmentally regulated synthesis of a compound bio-synthetically related to polyketide antibiotics. Mol. Microbiol. 4:1679–1691.
13. Davis, N. K., and K. F. Chater. 1992. The Streptomyces coelicolor whiB geneencodes a small transcription factor-like protein dispensable for growth butessential for sporulation. Mol. Gen. Genet. 232:351–358.
14. Delic, V., D. A. Hopwood, and E. J. Friend. 1970. Mutagenesis by N-methyl-N9-nitro-N-nitrosoguanidine (NTG) in Streptomyces coelicolor. Mutat. Res.9:167–182.
15. Flardh, K. Personal communication.16. Flett, F., V. Mersinias, and C. P. Smith. 1997. High efficiency intergeneric
conjugal transfer of plasmid DNA from Escherichia coli to methyl DNA-restricting streptomycetes. FEMS Microbiol. Lett. 155:223–229.
17. Hobbs, G., C. Frazer, D. C. J. Gardner, J. Cullum, and S. G. Oliver. 1989.Dispersed growth of Streptomyces in liquid culture. Appl. Microbiol. Bio-technol. 31:272–277.
18. Hopwood, D. A., H. Wildermuth, and H. M. Palmer. 1970. Mutants ofStreptomyces coelicolor defective in sporulation. J. Gen. Microbiol. 61:397–408.
19. Hopwood, D. A., M. J. Bibb, K. F. Chater, T. Kieser, C. J. Bruton, H. M.Kieser, D. J. Lydiate, C. P. Smith, J. M. Ward, and H. Schrempf. 1985.Genetic manipulation of Streptomyces: a laboratory manual. The John InnesFoundation, Norwich, England.
20. Kelemen, G. H. Personal communication.21. Kelemen, G. H., G. L. Brown, J. Kormanec, L. Potuckova, K. F. Chater, and
M. J. Buttner. 1996. The positions of the sigma factor genes, whiG and sigF,in the hierarchy controlling the development of spore chains in the aerialhyphae of Streptomyces coelicolor A3(2). Mol. Microbiol. 21:593–603.
22. Kelemen, G. H., P. Brian, K. Flardh, L. Chamberlin, K. F. Chater, and M. J.Buttner. 1998. Developmental regulation of transcription of whiE, a locusspecifying the polyketide spore pigment in Streptomyces coelicolor A3(2). J.Bacteriol. 180:2515–2521.
23. Kieser, T., and R. E. Melton. 1988. Plasmid pIJ699, a multi-copy positive-selection vector for Streptomyces. Gene 65:83–91.
24. Leblond, P., and B. Decaris. 1994. New insights into the genetic instability ofStreptomyces. FEMS Microbiol. Lett. 123:225–232.
25. McVittie, A. 1974. Ultrastructural studies on sporulation in wild-type andwhite colony mutants of Streptomyces coelicolor. J. Gen. Microbiol. 81:291–302.
26. Molle, V., and M. J. Buttner. Unpublished data.27. Molle, V., W. J. Palframan, and M. J. Buttner. Unpublished data.28. Palframan, W. J. 1998. The whiD locus of Streptomyces coelicolor A3(2).
Ph.D. thesis. University of East Anglia, Norwich, England.29. Potuckova, L., G. H. Kelemen, K. C. Findlay, M. A. Lonetto, M. J. Buttner,
and J. Kormanec. 1995. A new RNA polymerase sigma factor, sF, is requiredfor the late stages of morphological differentiation in Streptomyces sp. Mol.Microbiol. 17:37–48.
30. Redenbach, M., H. M. Kieser, D. Denapaite, A. Eichner, J. Cullum, H.Kinashi, and D. A. Hopwood. 1996. A set of ordered cosmids and a detailedgenetic and physical map for the 8Mb Streptomyces coelicolor A3(2) chro-mosome. Mol. Microbiol. 21:77–96.
31. Ryding, N. J. 1995. Analysis of sporulation genes in Streptomyces coelicolorA3(2). Ph.D. thesis. University of East Anglia, Norwich, England.
32. Ryding, N. J., G. H. Kelemen, C. A. Whatling, K. Flardh, M. J. Buttner, andK. F. Chater. 1998. A developmentally regulated gene encoding a repressor-like protein is essential for sporulation in Streptomyces coelicolor A3(2). Mol.Microbiol. 29:343–357.
33. Streptomyces coelicolor Genome Project Web Site. [Online.] http://www.sanger.ac.uk/Projects/S_coelicolor/. Sanger Centre, Wellcome Trust Genome Cam-pus, Hinxton, Cambridge, United Kingdom.
34. Volff, J.-N., and J. Altenbuchner. 1998. Genetic instability of the Streptomy-ces chromosome. Mol. Microbiol. 27:239–246.
35. Wilson, J., and D. H. Figurski. Personal communication.
VOL. 181, 1999 NEW SPORULATION LOCI IN S. COELICOLOR 5425
on May 19, 2018 by guest
http://jb.asm.org/
Dow
nloaded from