neurones from older neonates (p5-p7) were hyperpolarized by
TRANSCRIPT

Journal of Physiology (1991), 440, pp. 67-83 67With 10 figuresPrinted in Great Britain
STRYCHNINE-SENSITIVE GLYCINE RESPONSES OF NEONATAL RATHIPPOCAMPAL NEURONES
BY SUSUMU ITO* AND ENRICO CHERUBINItFrom the Unite 029, INSERM, 123 Boulevard de Port-Royal, 75014 Paris, France
(Received 5 November 1990)
SUMMARY
1. Intracellular recordings employing current and voltage clamp techniques wereused to study the effects of glycine on rat CA3 hippocampal neurones during the first3 weeks of postnatal (P) life.
2. Glycine (0 3-1 mM) depolarized neurones from rats less than 4 days old (P4).Neurones from older neonates (P5-P7) were hyperpolarized by glycine, whereasadult neurones were unaffected.
3. Both depolarizing and hyperpolarizing responses were associated with largeconductance increases; they reversed polarity at a potential which changed with theextracellular chloride concentration. The responses persisted in tetrodotoxin (1 gM)or in a solution with a much reduced calcium concentration.
4. Strychnine (1 #M) but not bicuculline (10-50 JIM) antagonized the effects ofglycine. The action of strychnine was apparently competitive with a dissociationconstant of 350 nm.
5. In voltage clamp experiments, glycine elicited a non-desensitizing outwardcurrent at -60 mV. When a maximal concentration of glycine was applied at thesame time as y-aminobutyric acid (GABA), the conductance increase induced by thetwo agonists was additive, suggesting the activation of different populations ofchannels.
6. Concentrations of glycine lower than 100 JM did not affect membrane potential.However, at 30-50#M glycine increased the frequency of spontaneous GABA-mediated synaptic responses; this action was not blocked by strychnine.
7. It is concluded that during the first 2 weeks of life glycine acts at strychnine-sensitive receptors to open chloride channels.
INTRODUCTION
Glycine is the main inhibitory transmitter in hindbrain areas (Aprison & Daly,1978). Its effect is mediated through the activation of high-affinity receptorslocalized on neurones of the spinal cord, medulla and pons (Young & Snyder, 1973).The interaction of glycine with its receptor results in an increase in chloride
* Present address: Department of Neurophysiology, Medical Research Institute, Tokyo Medicaland Dental University, Kanda-Surugadai Chiyoda-ku, Tokyo, Japan.
t Present address and to whom correspondence should be sent: International School forAdvanced Studies, Strada Costiera 11, 34014 Trieste, Italy.MS 8914 3-2

S. ITO AND E. CHERUBINI
conductance (Coombs, Eccles & Fatt, 1955; Araki, Ito & Oscarsson, 1961; Eccles,1964) an effect specifically blocked by the convulsant alkaloid strychnine (Curtis,Hosli & Johnston, 1968). The neurodepressant potency of glycine is related to theuneven distribution of this endogenous amino acid, being greatest in the spinal cordand brain stem level and progressively decreasing in more rostral areas (Kishimoto,Simon & Aprison, 1981). Electrophysiological experiments on cat and rat corticalneurones in vivo have shown that on these cells glycine does not play a major role insynaptic inhibition (Kelly & Krnjevic, 1969; Levi, Bernardi, Cherubini, Gallo,Marciani & Stanzione, 1982). Furthermore, glycine fails to change membranepotential or conductance of adult rat hippocampal neurones in vivo (Ben-Ari,Krnjevic, Reiffenstein & Reinhardt, 1981).
In addition to its classical inhibitory action, glycine serves also as an allostericregulator of excitatory neurotransmission mediated by the N-methyl-D-aspartate(NMDA) receptors (Johnson & Ascher, 1987). This action involves distinct glycinebinding sites, which are insensitive to strychnine (Bristow, Bowery & Woodruff,1986) and are mainly localized to rostral brain areas, including cortex andhippocampus (Kessler, Terramani, Lynch & Baudry, 1989). We have recentlyreported that, in hippocampal neurones from neonatal (but not adult) rats,micromolar concentrations of glycine enhance the frequency of y-aminobutyric acid(GABA)-mediated synaptic potentials and potentiate NMDA responses (Gaiarsa,Corradetti, Cherubini & Ben-Ari, 1990). This effect, which is strychnine insensitive,is clearly mediated by the allosteric site of the NMDA receptor located onGABAergic interneurones (Ben-Ari, Cherubini, Corradetti & Gaiarsa, 1989) since itcan be prevented by the specific NMDA receptor antagonist D-(-)-2-amino-5-phosphonovalerate (AP-5; Gaiarsa et al. 1990).We now report that, in addition to its facilitatory action on NMDA-mediated
events, glycine induced strychnine-sensitive, chloride-dependent responses detect-able only during the first 2 weeks of postnatal life.
Part of this work has been reported in a preliminary form (Ito & Cherubini, 1990).
METHODS
Experiments were performed on CA3 hippocampal neurones from slices obtained from 2- to 21-day-old (P2-P21 days; 0 is taken as the day of birth) and adult Wistar rats. The methods forpreparing and maintaining the slices have been extensively reported (Ben-Ari et al. 1989). Ratswere decapitated under ether anaesthesia and their brains quickly removed from the skulls.Hippocampi were dissected free; transverse 600 ,am thick slices were cut and immediatelyincubated at room temperature (20-22 °C) in artificial cerebrospinal fluid (ACSF) of the followingcomposition (mm): NaCl, 126; KCl, 3-5; CaCl2, 2; NaH2PO4, 1-2; MgCl2, 13; NaHCO3, 25; glucose,11. Equilibrating the ACSF with 95% 02 and 5% CO2 gave a pH of 7 3-7A4. The slices were allowedto recover for at least 1 h before being transferred to a recording chamber in which they werecontinuously superfused at 33-34 °C with oxygenated ACSF at a rate of 25-3 ml min-'.Intracellular recordings were made with microelectrodes filled with 2 M-K-methylsulphate(resistance 50-100 MQ). In a few experiments CA3 neurones were impaled with microelectrodescontaining 3 M-KCl or 3 M-CsCl (resistance 40-60 MQ). Current was injected through the recordingelectrode by means of an Axoclamp 2A amplifier. Bridge balance was checked repeatedly duringthe experiments and capacitative transients (with the electrode tip outside the neurone) werereduced to a minimum by negative capacity compensation. Stabilization of intracellular recordingwas helped by the injection of small hyperpolarizing currents (0 1-0 3 nA) but in cells from slicesobtained from the youngest animals it often was necessary to maintain such a small current
68

GLYCINE ON NEONATAL HIPPOCAMPAL NEURONESindefinitely (Schwartzkroin & Kunkel, 1982). Membrane potential was estimated from thepotential observed upon withdrawal of the electrode from the cell. Membrane input resistance wasmeasured from the amplitude of small hyperpolarizations (200-300 ms duration) evoked by passingknown currents across the cell membrane. The conductance increment induced by glycine(fractional conductance increase, Ag, Morita, North & Tokimasa, 1982) was calculated from(R/R') -1, where R is the input resistance at the resting potential and R' is the input resistanceduring glycine response.
In voltage clamp experiments, membrane currents were recorded via a single-electrode voltaaeclamp amplifier (Axoclamp 2A, Axon Instruments, Inc.), switching between voltage recording andcurrent injection at 3-4 kHz (30% duty cycle). The voltage signal at the head stage amplifier wascontinuously monitored on a separate oscilloscope to ensure optimal performance of the voltageclamp system. Responses were digitized and displayed on a Nicolet digital oscilloscope and on acomputer-driven chart-recorder. Some experiments were performed in a low-calcium-high-magnesium ACSF (0-2 mM-Ca2+ and 6 mM-Mg2+ instead of 2 and 1-3 mM) or in a low-chloride ACSFwhere 126 mM-NaCl was substituted by 126 mM-sodium isethionate, corresponding to 92-6%chloride substitution. Drugs were dissolved in ACSF and applied through a three-way tap systemby changing the superfusing solution to one which differed only in its content of drug(s). There wasa delay of 20-30 s between turning the tap and the first arrival at the tissue of the changed solution.The ratio of flow rate to bath volume ensured complete exchange within 1 min. Drugs used were:glycine, GABA, strychnine, bicuculline, all purchased from Sigma; D-(-)-2-amino-5-phos-phonovalerate (AP-5) and (± )3-(2-carboxy-piperazin-4-yl)-propyl-1-phosphonic acid (CPP)purchased from CRB or Tocris. Unless otherwise stated, all data are expressed as means+ S.D.
RESULTS
Stable intracellular recordings were obtained from sixty-four pyramidal neuronesin slices from 2- to 21-day-old rats or from three adult rats. In agreement with aprevious study (Ben-Ari et al. 1989), all neurones recorded during the first postnatalweek with potassium methylsulphate electrodes (n = 42) had a resting membranepotential of - 65-2 + 78 mV (ranging from -55 to -75 mV, see Fig. 5A), restinginput resistance ranging from 80 to 130 MQ and action potentials greater than55 mV.
Membrane potential changes induced by glycineDays PO-P4
Superfusion with glycine (0-3 mm), caused at resting membrane potential amembrane depolarization (4 ± P17 mV, n = 7), which was associated with anincrease in membrane input conductance. The depolarization had a latency of20-30 s after glycine reached the tissue and achieved its peak amplitude at 1 min.This effect washed out in a few minutes. A typical example from a 3-day-old rat isshown in Fig. 1A. The sensitivity to glycine varied from cell to cell, but in a givenneurone the response varied with the concentration applied. The minimum dose ofglycine required to produce membrane potential changes was 100 ,LM. Increasing theconcentration of glycine increased the peak amplitude, particularly the rate of riseof the depolarization.As already reported (Ben-Ari et al. 1989), > 90% of the neurones at P0-P4 exhibit
spontaneous GABA-mediated giant depolarizing potentials (GDPs). These consistedof brief, high-frequency bursts of spikes riding on large depolarizing potentials.Concentrations of glycine lower than those required to cause membrane potentialchanges (30-50 JtM) increased the frequency of GDPs. This effect, which wasinsensitive to strychnine (1 /M), was blocked by the NMDA receptor antagonists AP-
69

S. ITO AND E CHERUBINI
5 or CPP (50 JLM), suggesting that it was mediated by the allosteric modulatory siteof the NMDA receptors located on GABAergic interneurones (Gaiarsa et al. 1990). Asimilar effect was seen with application of the higher glycine concentrations, but itoccurred transiently and before the membrane potential changed, presumably a
AGly (300 gM, P3) Gly (1 mM, P6).=. =
-61 mV' -64 MV II
~I10 mVttt ~~~1min
B
Control (P7) TTX (1 gM) Low calcium, high magnesium
-63 mV 4p ft4V
10 mV
1 minFig. 1. Glycine depolarized or hyperpolarized neonatal neurones. A, at P3, bathapplication of glycine (Gly, open bars) induced a membrane depolarization associatedwith an increase in input conductance. Note that glycine first increased and thensuppressed (for 5 min) spontaneously occurring GABA-mediated giant depolarizingpotentials. At P6, glycine induced a membrane hyperpolarization and an increase in inputconductance. The effect of glycine was associated with an increase in frequency ofspontaneous GABA-mediated hyperpolarizing potentials (arrows) followed by theirsuppression. Upward deflections on the left panel are action potentials riding on slowdepolarizations; on the right panel are single spikes. B, the responses to glycine (1 mM,open bars) were unchanged in the presence of TTX or in a low-calcium (0-2 mM), high-magnesium (6 mM) solution. Downward deflections in this and following figures areelectrotonic potentials resulting from injection of a fixed current pulse through therecording electrode.
reflection of the slowly increasing concentration of glycine in the tissue slice when itwas applied by superfusion.
Days P4-P7Towards the end of the first postnatal week, during a transitional period
characterized by the progressive disappearance of GDPs and their replacement bylarge hyperpolarizing potentials (responses to GABA also changed from thedepolarizing to the hyperpolarizing ones; Ben-Ari et al. 1989), glycine inducedmembrane hyperpolarization and increased input conductance (Fig. 1A).
Like the depolarizing response, the hyperpolarizing response was also dosedependent. Increasing the concentration of glycine caused an increase in the peak
70

GLYCINE ON NEONATAL HIPPOCAMPAL NEURONES
amplitude of the membrane hyperpolarization, particularly in the rise of thehyperpolarization. At P7, concentrations of glycine of 0 3 and 1 mm caused (atresting membrane potential) a hyperpolarization of 2-2 + 0-6 (n = 9) and 4-3 + 1-5 mV(n = 7) respectively.The depolarizing or hyperpolarizing response to glycine was unchanged by
tetrodotoxin (TTX, 1 ,tM) or by a low-calcium (0-2 mM), high-magnesium (6 mm)solution, implying that it probably did not result from action potential generationand release of transmitter(s) from surrounding cells (Fig. 1B).As for GDPs, the hyperpolarizing effect of glycine was associated with a transient
increase in frequency of spontaneous GABA-mediated large hyperpolarizingpotentials which, at this stage of development, replace GDPs (Gaiarsa et al. 1990).The spontaneous firing was also completely and reversibly blocked by glycine (Fig.1A).
Days P7-adultAfter the first postnatal week, the sensitivity to glycine decreased gradually and
completely disappeared after P14. At the end of the second postnatal week and inadult neurones, glycine at concentrations of 3-10 mm failed to induce any change inmembrane potential, input conductance or synaptic noise.
Conductance changes elicited by glycineDays PO-P4
Glycine-induced depolarizations were accompanied by an increase in cell inputconductance, measured from the amplitude of the hyperpolarizing electrotonicpotentials (Fig. 2A). The conductance increase persisted after repolarizing themembrane potential to its initial value at rest. The conductance increase reached itsmaximum within 20-30 s after the beginning of glycine application. Applications of1-2 min were usually enough to obtain steady-state responses; prolongation of thedrug application did not show any further significant change in conductance.Although the sensitivity to glycine varied from cell to cell, the conductance changebecame detectable at a concentration of 100 #M (Fig. 2A). The conductance changesincreased with higher concentrations and reached the maximum at concentrations of3-10 mm. With these glycine concentrations, the amplitude of the response andassociated conductance increase, after an initial peak, declined to reach a steady-state value; this was also seen when the application of glycine was maintained forseveral minutes.
Days P4-P7Glycine-induced hyperpolarizations were associated with an increase in cell input
conductance. Figure 2B shows the mean fractional conductance increase induced byincreasing concentration of glycine during this period of postnatal life. The half-maximal conductance change was obtained with a glycine concentration of 1H1 mm.The Hill coefficient, calculated in two neurones from the conductance changesinduced by increasing concentrations of glycine, was 1-6 and 1-8 respectively.
71
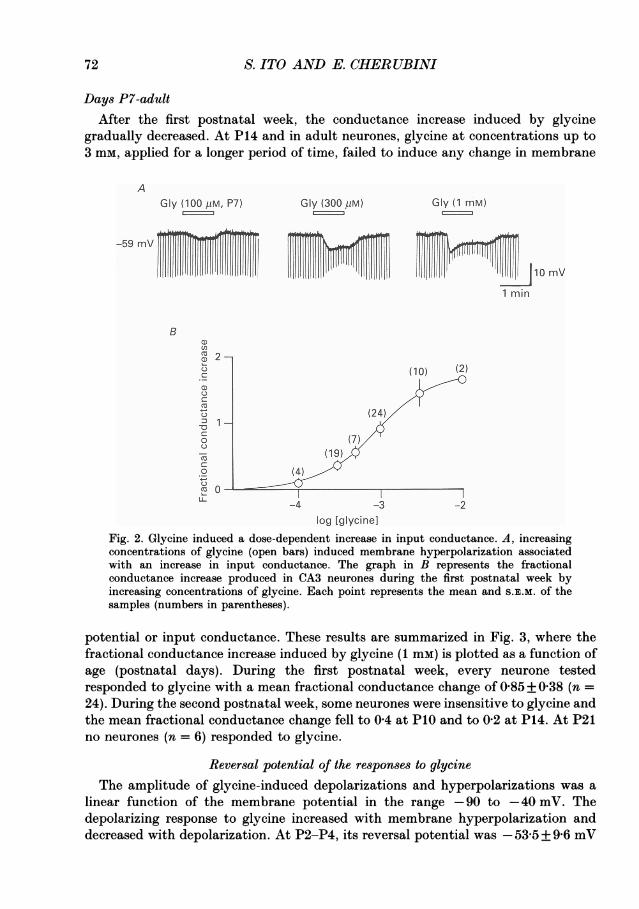
S. ITO AND E. CHERUBINI
Days P7-adultAfter the first postnatal week, the conductance increase induced by glycine
gradually decreased. At P14 and in adult neurones, glycine at concentrations up to3 mm, applied for a longer period of time, failed to induce any change in membrane
AGly (100 kiM, P7) Gly (300 PM) Gly (1 mM)
-59 mV TiI W 10m1 min
Bu)
(10) (2)
(24)
o (7)
o ~~~~~(4)/0-
-4 -3 -2
log lglycinelFig. 2. Glycine induced a dose-dependent increase in input conductance. A, increasingconcentrations of glycine (open bars) induced membrane hyperpolarization associatedwith an increase in input conductance. The graph in B represents the fractionalconductance increase produced in CA3 neurones during the first postnatal week byincreasing concentrations of glycine. Each point represents the mean and S.E.M. of thesamples (numbers in parentheses).
potential or input conductance. These results are summarized in Fig. 3, where thefractional conductance increase induced by glycine (1 mM) is plotted as a function ofage (postnatal days). During the first postnatal week, every neurone testedresponded to glycine with a mean fractional conductance change of 0-85 + 0-38 (n =24). During the second postnatal week, some neurones were insensitive to glycine andthe mean fractional conductance change fell to 0 4 at PIO and to 0-2 at P14. At P21no neurones (n = 6) responded to glycine.
Reversal potential of the responses to glycineThe amplitude of glycine-induced depolarizations and hyperpolarizations was a
linear function of the membrane potential in the range -90 to -40 mV. Thedepolarizing response to glycine increased with membrane hyperpolarization anddecreased with depolarization. At P2-P4, its reversal potential was - 53-5 + 9-6 mV
72

GLYCINE ON NEONATAL HIPPOCAMPAL NEURONES
(Fig. 4, n = 10) with potassium methylsulphate-containing electrodes and-23 + 7-2 mV (n = 5) with KCl- or CsCl-filled electrodes. After P4, the responses toglycine shifted from the depolarizing to the hyperpolarizing direction. Thus theamplitude of glycine-induced hyperpolarization increased with depolarization and
A
Gly (300 pm, P3) Gly (300 pM, P6) Gly (3 mM, P14)
-60 .... -68-63 mV mV mV
jiQ mVB ° 1 min
a,
CD)1 (9)
C4 (9)() (6)
o (6 )
0C (6)C.) 5_ (; ) 6
.L 1 7 14 21Age (days)
Fig. 3. Glycine induced a conductance increase only during the first 2 weeks of postnatallife. A, glycine (open bars) caused membrane depolarization or hyperpolarizationassociated with an increase in input conductance at P3 and P6 respectively. At P14, ahigher concentration of glycine, applied for 2 min, failed to induce any change inmembrane potential or input conductance. B, fractional conductance increase induced byglycine (1 mM) as a function of age (postnatal days). Each point represents the mean andS.E.M. (numbers in parentheses). TTX (1 /iM) present throughout the experiment.
decreased with hyperpolarization. The reversal potential recorded between P5 andP7 was - 63-2 + 11-6 mV with potassium methylsulphate-containing electrodes (Fig.4, n = 20) and -40X2 + 10-6 mV with KCl-filled electrodes (n = 5).
Figure 5 shows the distribution values of the resting membrane potential and thereversal potential of the responses to glycine in neurones recorded at P2-P7 withpotassium methylsulphate-containing electrodes. In agreement with a previousreport (Ben-Ari et al. 1989), we observed that, in contrast to the reversal potentialof the responses to glycine, which showed a significant difference (P < 0-01,rank-sum test) between the P2-P4 and P5-P7 groups, the resting membranepotential of cells at P2-P4 was not significantly different from that recorded atP5-P7. The mean resting membrane potential at P2-P4 was - 66-6+ 7 mV (n = 11)and at P5-P7 was - 64-5+ 8 mV (n = 31). The regression line for reversal potentialscrossed that of resting membrane potentials between P5 and P6.
73

S. ITO AND E. CHERUBINI
Gly(1 mM, P2,A)lZZ=
Gly (300 piM, P7, 0)
-38 mVi-68 mv
-78
-68 -88r8 "
..;. :, .~~ ~~~~~~~~~ .,...
10 mv1 min
B
-4
lJjI10m\1 min
E
-10 vCD0A O.
0 A CD
90 -80 -70 A -50 -40 0-6 'A '° 60
>
.
°O -5 0)0 0
Membrane potential (rmV) -1 0 V
v ~~~~~~~EQ
Fig. 4. Reversal of glycine responses. A, changing the membrane potential by passing asteady current through the recording electrode changed the amplitude of thedepolarization or hyperpolarization induced by glycine (open bars) at P2 (A) and P7 (0)respectively. The resting membrane potential of both cells was -68 mV. B, relationshipbetween the membrane potential and the amplitude of glycine responses for the cellsshown in A. Note the shift in the reversal potential from -55 mV (at P2) to -80 mV (atP7). TTX (1 /sM) present throughout the experiment.
A B
-30 -
0
*0 M9. . t. s
E
-zo._
0)
a
U)
cc
-50 -
-70 -
_1nn
0~~~
c I I -II I I I I -
1 2 3 4 5 6 7 8 1 2 3 4 5 6 7 1Age (days) Age (days)
Fig. 5. Changes in reversal potential for glycine are independent from the restingmembrane potential. Plot of the distribution values of the resting membrane potentials(A) and the reversal potentials of glycine responses (B) as a function of age (postnataldays). The regression line for reversal potentials crossed that of resting membranepotentials between P5 and P6.
8
Ionic mechanismsChanging the extracellular chloride concentrations [CI-]0 from 136-1 to 101 mm
(isethionate substitution) increased the amplitude of the depolarizing response toglycine. In a low-chloride medium the hyperpolarizing response to glycine shifted to
74
A
g -30-
4-C2 -50-0a0C
. -70-a) .E0)E
C -
0) -inn -a
-48 .p
I,! ';il, ii, "!

GLYCINE ON NEONATAL HIPPOCAMPAL NEURONES
the depolarizing direction. Once it reached the threshold, the depolarization gave riseto action potentials (Fig. 6). A 13-5-fold change in [Cl-]0 yielded a change in glycinereversal potential, Egly, of 40+3-6 mV (n = 4). As shown in Fig. 6B, the slightdeviation of Egly from chloride reversal potential (Ec1), calculated from the Nernst
A Control, Gly (1 mM, P6) Low Cl-, Gly (1 mM) Wash, Gly (1 mM)
-70~mV1'/ |tX 0 T | -25 mV
1 min
B Eci0
E-50 - )_( (3)
(7)
-100- r10 30 100
[CI-]o (mM)Fig. 6. Chloride dependency of the response to glycine. A, the response to glycine (openbar) recorded in the presence of TTX (1 uM) shifted to the depolarizing direction andincreased in amplitude during superfusion with low-chloride (10 mM) medium (isethionatesubstitution). The depolarization gave rise to calcium action potentials. B, relationbetween the reversal potential for glycine responses obtained at P6-P7 (Eg,y) and theextracellular chloride concentration [Cl-]0. Each point represents the mean and S.D. forthe samples (numbers in parentheses). The dashed line represents the change in 67-8 mVfor 13-5-fold change of [Cl-]0, predicted from the Nernst equation.
equation, might reflect either a small permeability of isethionate through chloridechannels or slow changes in driving forces concomitant with movements of chlorideions in and out of the cells (Adams, Constanti & Banks, 1981).
Reduction of calcium concentration from 2 to 0-2 mm changed neither theamplitude (Fig. IB) nor the reversal potential of the responses to glycine.The role of potassium ions in glycine response was assessed by recording with CsCl-
filled electrodes. Glycine depolarizations were similar whether Cs+ or K+ was used inthe recording electrode. In three cells recorded with electrodes containing CsCl, atresting membrane potential, glycine (0 3 ,tM) caused a membrane depolarization of5-2 + 2 mV.
75

S. ITO AND E. CHERUBINI
Strychnine selectively antagonizes glycine responsesBath application of strychnine (0 3-1 ,tM) reversibly depressed or abolished glycine
responses. Usually strychnine (1 ,UM) completely antagonized the responses to glycine(300-500 /tM). The depression of glycine response was maximal within 3-5 min from
A Control
Gly (1 mM) Gly (3 mM)B
-59 mV (
-59m V_ TT u 5Control
-l X ~~~StrychnineStrychnine (1 pM) ' 4 S ry pM
X1I 03 X / 1 M3pU,Mg30
Strychnine (3pM) , 10
i. 00 300 1 3 10 3011,1 '""jJlOmV PM PM mMM M MMm1 min [Glycine]
C2-
0)0
1 Onm 1pm 3pm 10pmStrychnine
Fig. 7. Strychnine antagonizes the responses to glycine in an apparently competitivemanner. A, responses obtained at P7 to two different concentrations of glycine (open bars)in control conditions and in the presence of 1 or 3 /tM-strychnine (in the presence of TTX,1 /IM). B, dose-response curves for glycine before and during superfusion with strychninein the concentrations indicated (/uM). Strychnine caused a parallel shift of the glycineconcentration-response curve. C, Schild plot of results shown in B. Linear regression line(slope 1) yielded an estimated pA2 (-log Kd) value of 6-57 for strychnine. DR, dose ratio.
the beginning of strychnine superfusion and a complete recovery was obtained20-30 min after wash. Figure 7 shows the parallel shift to the right of the glycineconcentration-response curve by increasing doses of strychnine. The dose ratio was30 + 6 at 10 /tM-strychnine and the apparent dissociation constant value (apparentKd) for strychnine was 350+ 76 nM (n = 4).To see whether the depressant effect of strychnine was selective for glycine, both
strychnine and bicuculline were tested on the responses induced in the same neuronesby superfusion of glycine and GABA (n = 6). Strychnine (1 ,UM) antagonized theresponse to glycine but had little effect on the GABA response. In contrast
76

GLYCINE ON NEONATAL HIPPOCAMPAL NEURONES
bicuculline (10-50 /uM) depressed the GABA response without affecting the responseto glycine (Fig. 8).
Comparison of the responses to glycine and GABAIn neonatal hippocampal neurones at PO-P4, GABA, acting via GABAA receptors,
causes a bicuculline-sensitive depolarization and an increase in chloride conductance
Control
Gly (1 mM) GABA (300 AiM)
PII
-62 mVlg I<I
Bicuculline (50 AiM)
Strychnine (1 AM)
1!il. 1ii il!iiEll1!lil!i.l! 1| li 10 mV1 min
Fig. 8. Strychnine antagonized glycine but not GABA responses. Responses to glycine(open bar) and GABA (filled bar) obtained at P6 in a neurone recorded with a potassiummethylsulphate-containing electrode in the presence of TTX (1 /uM). Bicuculline (50 /M)blocked GABA but had no effects on glycine response. Strychnine (1 /tM) blocked glycinebut had no effects on GABA response.
(Ben-Ari et al. 1989). At P5-P7, GABA responses shift from the depolarizing to thehyperpolarizing direction (Ben-Ari et al. 1989). It seems therefore of interest tocompare, on the same cells, responses to GABA and glycine in order to find outwhether they both followed the same temporal changes in polarity. Comparableresponse amplitudes were usually obtained with concentrations of glycine 2-3 timeshigher than those ofGABA. When applied to the same neurones (n = 8), the reversalpotential for GABA was exactly the same as that for glycine. In fact, the difference
77

78 S. ITO AND E. CHERUBINI
in the reversal potential of GABA and glycine (recorded with electrodes containingpotassium methylsulphate) was 0-33 + 0 5 mV at P2-P4 (n = 3) and 0-2 + 1 1 mV atP5-P7 (n = 5).
In voltage clamp experiments with KCl electrodes, superfusion ofGABA (0 3 mM)or glycine (1 mM) caused inward currents (in three neurones held at -60 mV these
A
Gly (500 gM) GABA (1 mM)
Vh -87 mV
0.5 nA1 min
B
Potential (mV) 0.4 Potential (mV) 0.4
02< 0.2<-80 -60 -40 -20 -80 -60 -40 -20 0
- -0)Corntrol Contro
-.2 o - -0.2 uGlycine GABA --
-0.4 GAA-0.4
Fig. 9. Glycine and GABA induced comparable inward currents and reversed at the samepotential. A, inward currents induced by bath application of glycine (open bar) andGABA (filled bar) to a P6 neurone held at -87 mV (recorded with a KCl-containingmicroelectrode in the presence of TTX, 1 ,UM). Note the difference in the baseline noisebetween glycine and GABA. Vh, holding potential. B, steady-state I-V plots obtained fromslow depolarizing ramps before and after application of glycine (300 1uM, left) and GABA(300 /SM, right) in a P4 neurone recorded with a potassium methylsulphate-containingelectrode in the presence of TTX (1 M).
were 347 + 22 and 362 + 32 pA for glycine and GABA, respectively). In theseconditions, the increase in the baseline noise due to GABA was much smaller thanthat due to glycine (Fig. 9A). This probably reflects differences in single-channelconductance between GABA and glycine (see also Gold & Martin, 1984). Withpotassium methylsulphate electrodes, at a holding potential of about -60 mV,glycine or GABA induced inward currents at P2-P4, whereas, after P4, in five outof six neurones, outward currents were induced. The currents usually reached thepeak after 40-60 s, then slightly declined to a plateau, which was maintainedthroughout GABA or glycine superfusion. The decline was not associated withchanges in leak conductance, making the possibility of receptor desensitizationunlikely (see Fig. 10). A full recovery was obtained a few minutes after wash-out. Thevoltage dependence of GABA- and glycine-induced currents was studied bydetermining the steady-state current-voltage plots obtained from slow depolarizingramps in the absence and presence of the agonists (Fig. 9B). GABA- and glycine-

GLYCINE ON NEONATAL HIPPOCAMPAL NEURONES 79
induced currents were almost linearly related to the membrane potential in the range-80 to -40 mV; in both cases the reversal potential occurred at -63 mV. Asalready mentioned, GABA or glycine responses were selectively blocked bybicuculline and strychnine respectively, suggesting the involvement of two different
AGly (3mM)
2010]3 P ooooooooooooooo10 0
GABA (300 jiM) 3020
0 ~0040-30 -
2010Gly GABA 0 A I
0 1 2 3 4Time (min)~~~~~~ ~~~~~10.5 nA
Vh -57 mV 1 min
Fig. 10. Glycine and GABA activated different populations of channels. A, outwardcurrents induced in a P6 neurone held at -57 mV by application of a saturatingconcentration of glycine (3 mm, open bar), GABA (300 #M, filled bar) and by combinedapplication of glycine and GABA (bottom trace). When applied together, the conductanceincrease produced by glycine summed to that produced by GABA. Hyperpolarizingvoltage steps of 10 mV (2 s duration) were applied throughout the experiment. Note thatthe conductance increase induced by glycine and GABA was maintained throughoutapplication of the agonists, showing only little desensitization. TTX (1 /tM) presentthroughout the experiment. B, time course of the conductance changes induced by glycine(0), GABA (@) and combination of glycine and GABA (A). The conductance changescaused by concomitant application of glycine and GABA (A) were identical to thoseobtained by the sum of the two individual conductances (A).
receptors. It was therefore interesting to see whether these amino acids in turnactivated separate channels having different properties. If we assume that GABA orglycine activate different conductances, a saturating dose of glycine should give aresponse not occluded by co-application ofGABA. A typical experiment is illustratedin Fig. 10 where the conductance increase produced by a saturating concentration ofglycine and the conductance change induced by GABA when the agonists wereapplied together were additive. Similar results were obtained in three other neurones.This suggests that GABA and glycine activate different populations of channels.
DISCUSSION
The major finding of the present investigation is the novel observation of chloride-dependent, strychnine-sensitive responses to glycine only during the first 2 weeks ofpostnatal life and not after P15.

S. ITO AND E. CHERUBINI
Glycine responses represent a characteristic feature of postnatal developmentIn adult neurones, binding sites for strychnine, which represent postsynaptic
glycine receptors associated with chloride channels (Young & Snyder, 1973), aremostly concentrated in the spinal cord and brain stem and less abundant in higherbrain areas (Kishimoto et al. 1981). At birth, strychnine binding sites are weaklyexpressed in the spinal cord (Benavides, Lopez-Lahoya, Valdivieso & Ugarte, 1981;Becker, Hoch & Betz, 1988; Bruning, Bauer & Baumgarten, 1990); they increaseprogressively thereafter to reach adult levels at P15. This is different from what isfound in the brain, where a transient increase in glycine receptors with a peak aroundP5, followed by a progressive decrease to a plateau level at P15, has been reported(Benavides et al. 1981). The extremely low concentration of glycine receptors presentin the adult brain may explain the lack of effect of glycine in adult hippocampalneurones recorded in situ (Ben-Ari et al. 1981) or in vitro (present study).
Glycine activates a chloride conductanceAs suggested by the experiments with manipulations of chloride concentration, a
chloride permeability increase was probably responsible for the effects of glycine. Asin the case of GABA (Ben-Ari et al. 1989), glycine-mediated responses shifted fromthe depolarizing to the hyperpolarizing direction during the first postnatal days,without any significant change in resting membrane potential. As proposed forGABA-mediated responses (Ben-Ari et al. 1989), this change may be due tomodifications in the chloride gradient, resulting from the reversed operation of thechloride membrane pump (Misgeld, Deisz, Dodt & Lux, 1986) during early postnataldays. Alternatively, if glycine-activated channels are also permeable to bicarbonateions (HC03-), the reversal potential for glycine responses will deviate significantlyfrom the chloride equilibrium potential towards the bicarbonate equilibriumpotential (Kaila, Pasternack, Saarikoski & Voipio, 1989). Since the intracellular levelof HC03- is determined by the pH regulatory mechanism of the cell (Thomas, 1984),any developmental change in cell metabolism might shift the HC03- equilibriumpotential and therefore the glycine equilibrium potential.
In agreement with other studies on cultured mouse spinal neurones (Barker,McBurney & Macdonald, 1982) and acutely dissociated hippocampal and medullaoblongata neurones (Krishtal, Osipchuk & Vrublevsky, 1988), the glycine-gatedcurrents were voltage independent over the range -80 to -20 mV. The Hillcoefficient of 1'7 is consistent with two molecules of glycine binding to one receptorsite to open the associated chloride channel. This value is similar to that found forglycine-activated currents of acutely isolated hypothalamic neurones (Akaike &Kaneda, 1989).As in spinal cord neurones (Curtis et al. 1968), the activation of the chloride
conductance by glycine was selectively blocked by strychnine in a competitivemanner. The Kd value (350 nM) was about 20-fold higher than that found in spinalcord preparation using membrane fractions (for a review see Betz & Becker, 1988).The antagonism of glycine response by strychnine and not by bicuculline indicates
that glycine was acting via a receptor distinct from GABAA. Based on cross-desensitization experiments and analysis of channel closure times, one common class
80

GLYCINE ON NEONATAL HIPPOCAMPAL NEURONES
of multistate chloride channels has been proposed for the glycine and GABAAreceptor (Barker & McBurney, 1979; Hamill, Bormann & Sakmann, 1983).Nevertheless, other results, based on differences in permeabilities to organic anionsbetween GABA and glycine receptors (Kelly, Krnjevic, Morris & Yim, 1969;Bormann, Hamill & Sakmann, 1987) and on occlu.ion experiments (Gold & Martin,1984), argue in favour of two distinct chloride channel entities. Since in the presentstudy a saturating response to glycine was not occluded by simultaneous applicationof GABA, it seems that at least in neonatal hippocampal neurones, GABA andglycine receptors were probably coupled to separate chloride channels.
In recent years, a considerable amount of work has been devoted to thecharacterization of the molecular structure of the glycine receptor (Betz & Becker,1988) which comprises three separate subunits having strong similarities in theiramino acid sequences with the GABA or nicotinic acetylcholine receptors(Grenningloh, Rienitz, Schmitt, Methfessel, Zensen, Beyreuther, Gundelfinger &Betz, 1987). During postnatal development, a different form of the inhibitory glycinereceptor has been identified in the rat spinal cord. This is characterized by lowstrychnine binding affinity, altered antigenicity and abundance of the 49 kDasubunit which is not present in the adult form (Becker et al. 1988). Furthermore, twodifferent classes of glycine receptors with distinct electrophysiological characteristicscan be expressed in Xenopus oocytes injected with mRNA isolated from neonatal oradult spinal cord (Akagi & Miledi, 1988). The neonatal form differs from that of theadult in the rate of desensitization (faster) and sensitivity to glycine (lower). Theresponses to glycine obtained in neonatal hippocampal neurones probably reflect theactivation of a slightly different receptor type since they were quite sensitive toglycine and showed little desensitization.
Physiological significanceWe have already reported that glycine in low concentrations enhances the
frequency of spontaneously occurring GABAergic synaptic potentials during the first2 weeks of postnatal life (Gaiarsa et al. 1990). This effect is strychnine insensitive andis exerted on NMDA receptors located on GABAergic interneurones, since it wasblocked by NMDA receptor antagonists or by bicuculline. In addition to such afacilitatory effect, the present study shows that glycine can also modulate cellexcitability by exerting an inhibitory action mediated through strychnine-sensitivereceptors coupled to chloride channels present only during a restricted period ofpostnatal life.
The authors wish to thank Drs Y. Ben-Ari and A. Nistri for helpful comments on the manuscriptand Mrs S. Guidasci for photographic assistance.
REFERENCES
ADAMS, P. R., CONSTANTI, A. & BANKS, F. W. (1981). Voltage clamp analysis of inhibitory synapticaction in crayfish stretch receptor neurons. Federation Proceedings 40, 2637-2641.
AKAGI, H. & MILEDI, R. (1988). Heterogeneity of glycine receptors and their messenger RNAs inrat brain and spinal cord. Science 242, 270-273.
81

S. ITO AND E. CHER UBINI
AKAIKE, N. & KANEDA, M. (1989). Glycine-gated chloride current in acutely isolated rathypothalamic neurons. Journal ofNVeurophysiology 62, 1400-1409.
APRISON, M. H. & DALY, E. C. (1978). Biochemical aspects of transmission at inhibitory synapses:The role of glycine. In Advances in Neurochemistry 3, ed. AGRANOFF, B. WV. & APRISON, M. H., pp.203-294. Plenum Publishing Corporation, New York.
ARAKI, T., ITO, M. & OSCARSSON, 0. (1961). Anion permeability of the synaptic and non-synapticmotoneurone membrane. Journal of Physiology 159, 410-435.
BARKER, J. L. & McBURNEY, R. N. (1979). GABA and glycine may share the same conductancechannel on cultured mammalian neurones. Nature 277, 234-236.
BARKER, J. L., McBURNEY, R. N. & MACDONALD, J. F. (1982). Fluctuation analysis of neutralamino acid responses in cultured mouse spinal neurones. Journal of Physiology 322, 365-387.
BECKER, C. M., HoC, W. & BETZ, H. (1988). Glycine receptor heterogeneity in rat spinal cordduring postnatal development. EMBO Journal 7, 3717-3726.
BEN-ARI, Y., CHERUBINI, E., CORRADETTI, R. & GAIARSA, J.-L. (1989). Giant synaptic potentialsin immature rat CA3 hippocampal neurones. Journal of Physiology 416, 303-325.
BEN-ARI, Y., KRNJEVIC, K., REIFFENSTEIN, R. J. & REINHARDT, WV. (1981). Inhibitoryconductance changes and action of y-aminobutyrate in rat hippocampus. Neuroscience 6,2445-2463.
BENAVIDES, J., LOPEZ-LAHOYA, J., VALDIVIESo, F. &IUGARTE, M. (1981). Postnatal developmentof synaptic receptors in normal and hyperglycemic rats. Journal of Neurochemistry 37, 315-320.
BETZ, H. & BECKER, C. M. (1988). The mammalian glycine receptor: biology and structure of aneuronal chloride channel protein. Neurochemistry International 13, 137-146.
BORMANN, J., HAMILL, 0. P. & SAKMANN, B. (1987). Mechanisms of anion permeation throughchannels gated by glycine and y-aminobutyric acid in mouse cultured spinal neurones. Journalof Physiology 385, 243-286.
BRISTOW, J). R., BOWERY, N. G. & WOODRUFF, G. N. (1986). Light microscopic autoradiographiclocalisation of [3H]glycine and [3H]strychnine binding sites in rat brain. European Journal ofPharmacology 126, 303-307.
BRUNING, G., BAUER, R. & BAUMGARTEN, H. G. (1990). Postnatal development of [3H]fu-nitrazepam and [3H]strychnine binding sites in rat spinal cord localized by quantitativeautoradiography. Neuroscience Letters 110, 6-10.
COOMBS, J. S., ECCLES, J. C. & FATT, P. (1955). The specific ionic conductances and the ionicmovements across the motoneuronal membrane that produce the inhibitory postsynapticpotential. Journal of Physiology 130, 326-373.
CURTIS, D. R., HOSLI, L., JOHNSTON, G. A. R. & JOHNSTON, I. H. (1968). The hyperpolarization ofspinal motoneurones by glycine and related amino acids. Experimental Brain Research 5,235-258.
ECCLES, J. C. (1964). The ionic mechanism of postsynaptic inhibition. In Les Prix Nobel en 1963,pp. 261-283. The Nobel Foundation, Stockholm.
GAIARSA, J.-L., CORRADETTI, R., CHERUBINI, E. & BEN-ARI, Y. (1990). The allosteric glycine siteof the N-methyl-D-aspartate receptor modulates GABAergic-mediated synaptic events inneonatal rat CA3 hippocampal neurons. Proceedings of the National Academy of Sciences of theUSA 87, 343-346.
GOLD, M. R. & MARTIN, A. R. (1984). y-Aminobutyric acid and glycine activate Cl- channelshaving different characteristics in CNS neurones.,NVature 308, 639-641.
GRENNINGLOH, G., RIENITZ, A., SCHMITT, B., METHFESSEL, C., ZENSEN, M., BEYREUTHER, K.,GUNDELFINGER, E. D. & BETZ, H. (1987). The strychnine-binding subunit of the glycine receptorshows homology with nicotinic acetylcholine receptors. Nature 328, 215-220.
HAMILL, 0. P., BORMANN, J. & SAKMANN, B. (1983). Activation of multiple-conductance statechloride channels in spinal neurones by glycine and GABA. Nature 305, 805-808.
ITO, S. & CHERUBINI, E. (1990). Strychnine sensitive glycine responses in immature CA3hippocampal neurones. European Journal of Neuroscience, suppl. 3. 23.
JOHNSON, J. W. & ASCHER, P. (1987). Glycine potentiates the NMDA response in cultured mousebrain neurons. Nature 325, 529-531.
KAILA, K., PASTERNACK, M., SAARIKOSKI, J. & VOIPIO, J. (1989). Influence of GABA-gatedbicarbonate conductance on potential, current and intracellular chloride in crayfish musclefibres. Journal of Physiology 416, 161-181.
82

GLYCINE ON NEONATAL HIPPOCAMPAL NEURONES
KELLY, J. S. & KRNJEVId, K. (1969). The action of glycine on cortical neurons. Experimental BrainResearch 9, 155-163.
KELLY, J. S., KRNJEVI6, K., MoRRIs, M. E. & YIM, G. K. W. (1969). Anionic permeability ofcortical neurones. Experimental Brain Research 7, 11-31.
KESSLER, M., TERRAMANI, T., LYNCH, G. & BAUDRY, M. (1989). A glycine site associated with N-methyl-D-aspartic acid receptors: characterization and identification of a new class ofantagonists. Journal of Neurochemistry 52, 13 19-1328.
KISHIMOTO, H., SIMON, J. R. & APRISON, M. H. (1981). Determination of the equilibriumdissociation constants and number of glycine binding sites in several areas of the rat centralnervous system, using a sodium-independent system. Journal of Neurochemistry 37, 1015-1024.
KRISHTAL, 0. A., OSIPCHUK, Y. V. & VRUBLEVSKY, S. V. (1988). Properties of glycine-activatedconductances in rat brain neurones. Neuroscience Letters 84, 271-276.
LEVI, G., BERNARDI, G., CHERUBINI, E., GALLO, V., MARCIANI, M. G. & STANZIONE, P. (1982).Evidence in favour of a neurotransmitter role of glycine in the rat cerebral cortex. Brain Research236, 121-131.
MISGELD, U., DEISZ, R. A., DODT, H. U. & Lux, H. D. (1986). The role of chloride transport inpostsynaptic inhibition of hippocampal neurons. Science 232, 1413-1415.
MORITA, K., NORTH, R. A. & TOKIMASA, T. (1982). The calcium-activated potassium conductancein guinea-pig myenteric neurones. Journal of Physiology 329, 341-354.
SCHWARTZKROIN, P. A. & KUNKEL, D. D. (1982). Electrophysiology and morphology of thedeveloping hippocampus of fetal rabbits. Journal of Neuroscience 2, 448-462.
THOMAS, R. C. (1984). Experimental displacement of intracellular pH and the mechanism of itssubsequent recovery. Journal of Physiology 354, 3-22P.
YOUNG. A. B. & SNYDER, S. H. (1973). Strychnine binding associated with glycine receptors of thecentral nervous system. Proceedings of the National Academy of Sciences of the USA 70, 2832-2836.
83