mouse beta-defensin-14, an antimicrobial ortholog of human beta-defensin-3
TRANSCRIPT

Published Ahead of Print 10 March 2008. 10.1128/AAC.01308-07.
2008, 52(5):1876. DOI:Antimicrob. Agents Chemother. Jens M. Schröder, Jürgen Harder and Ehrhardt ProkschKerstin Hinrichsen, Rainer Podschun, Sabine Schubert, Ortholog of Human Beta-Defensin-3Mouse Beta-Defensin-14, an Antimicrobial
http://aac.asm.org/content/52/5/1876Updated information and services can be found at:
These include:
REFERENCEShttp://aac.asm.org/content/52/5/1876#ref-list-1at:
This article cites 19 articles, 11 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on May 2, 2014 by guest
http://aac.asm.org/
Dow
nloaded from
on May 2, 2014 by guest
http://aac.asm.org/
Dow
nloaded from

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, May 2008, p. 1876–1879 Vol. 52, No. 50066-4804/08/$08.00�0 doi:10.1128/AAC.01308-07Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Mouse Beta-Defensin-14, an Antimicrobial Ortholog of HumanBeta-Defensin-3�
Kerstin Hinrichsen,1 Rainer Podschun,2 Sabine Schubert,2 Jens M. Schroder,1Jurgen Harder,1†* and Ehrhardt Proksch1†
Department of Dermatology1 and Institute for Infection Medicine,2 University Hospital Schleswig-Holstein, Campus Kiel,24105 Kiel, Germany
Received 10 October 2007/Returned for modification 6 November 2007/Accepted 27 February 2008
Searching the database for mouse homologs of the antimicrobial peptide human beta-defensin-3 (hBD-3)revealed highest identity (69%) to mouse beta-defensin-14 (mBD-14). Recombinant mBD-14 exhibited broad-spectrum, nanomolar microbicidal activity. Treatment of keratinocytes with gamma interferon or transforminggrowth factor alpha increased mBD-14 gene expression. These data suggest that mBD-14 is the functionalortholog of hBD-3.
Antimicrobial proteins are effector molecules of the innateimmune system and offer a fast response to invading microor-ganisms by exhibiting potent antimicrobial activity (19). A ma-jor family of antimicrobial proteins in mammals comprises thebeta-defensins. Beta-defensins are small (4 to 5 kDa), cationicproteins which exhibit a potent antimicrobial activity at micro-to nanomolar concentrations (12). Human beta-defensin-3(hBD-3) was originally isolated from lesional psoriatic skinextracts, and it is inducibly expressed in many epithelia, such asthe skin, respiratory tract, and gut (4, 5, 12). hBD-3 is charac-terized by its potent antibacterial activity against many bacte-ria, including multiresistant strains (5, 8, 15). A BLAST searchof the mouse protein database with the amino acid sequence ofhBD-3 revealed highest identity (69%) to mouse beta-defen-sin-14 (mBD-14, Defb14) (Fig. 1). The next-closest hit wasmBD-3 (Defb3), with only 44% identity, followed by mBD-6,with only 38% identity. Interestingly, a BLAST search with themature hBD-3 peptide revealed only one hit, mBD-14. Thissuggests that mBD-14 could be the mouse ortholog of hBD-3.However, no data exist regarding the biological function ofmBD-14, and its potential role as an antimicrobial protein hasnot yet been evaluated. Therefore, we decided to recombi-nantly express mBD-14 in Escherichia coli and to analyze itsantimicrobial activity.
We used the software program SignalP 3.0 (2) to determinethe putative cleavage site in the mBD-14 amino acid sequence
to generate the mature protein (Fig. 1). The correspondingDNA encoding mBD-14 was amplified from mouse keratino-cyte cDNA using the forward primer 5�-ATCCAGATCTGGGTACCGACGACGACGACAACTTCCTACCAAAAACCCTCC-3 and reverse primer 5�-ATTTGCGGGCGCCTACTTCTTCTTTCGGCAGC-3�. The resulting fragment was clonedinto the expression vector pET32a(�) (Novagen, Madison,WI) to generate a fusion protein containing an N-terminal Histag sequence allowing purification of the fusion protein by theuse of a nickel affinity column. After expression in E. coliBL21(DE3)pLysS (Novagen), the fusion protein was purifiedusing a nickel affinity column (Macherey-Nagel, Dueren, Ger-many), followed by preparative C8 reversed-phase high-pres-sure liquid chromatography as described previously for thepurification of human beta-defensin-3 (5). The N-terminal partof the purified fusion protein was cleaved off by incubation for1 h at 37°C with enterokinase (Invitrogen, Carlsbad, CA), andthe resulting mature mBD-14 protein was purified by C4 re-versed-phase high-pressure liquid chromatography, similar tothe method described previously (5). Mass analysis using elec-trospray ionization mass spectrometry (QTOF-II hybrid massspectrometer; Micromass, Manchester, United Kingdom)yielded a mass of 5,184.3 Da, which is 6 Da less than thetheoretical mass calculated from the deduced amino acid se-quence (5,190.3 Da), suggesting that the six cysteyl residues ofmBD-14 are connected through three disulfide bridges.
* Corresponding author. Mailing address: University HospitalSchleswig-Holstein, Department of Dermatology, Campus Kiel, Schit-tenhelmstr. 7, 24105 Kiel, Germany. Phone: 49 431 5971598. Fax: 49431 5975243. E-mail: [email protected].
† J.H. and E.P. share senior authorship of this article.� Published ahead of print on 10 March 2008.
FIG. 1. Amino acid sequence alignment of hBD-3 and mBD-14 (single-letter code). The signal peptide and the recombinantly expressed maturepeptide are indicated.
1876
on May 2, 2014 by guest
http://aac.asm.org/
Dow
nloaded from
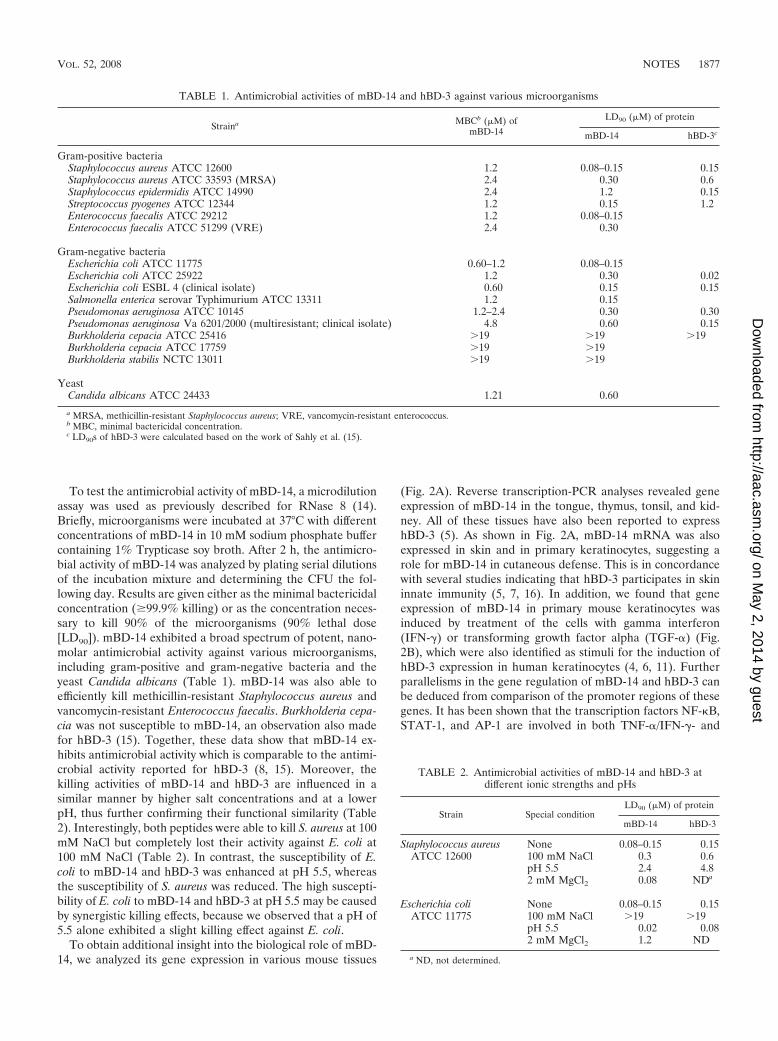
To test the antimicrobial activity of mBD-14, a microdilutionassay was used as previously described for RNase 8 (14).Briefly, microorganisms were incubated at 37°C with differentconcentrations of mBD-14 in 10 mM sodium phosphate buffercontaining 1% Trypticase soy broth. After 2 h, the antimicro-bial activity of mBD-14 was analyzed by plating serial dilutionsof the incubation mixture and determining the CFU the fol-lowing day. Results are given either as the minimal bactericidalconcentration (�99.9% killing) or as the concentration neces-sary to kill 90% of the microorganisms (90% lethal dose[LD90]). mBD-14 exhibited a broad spectrum of potent, nano-molar antimicrobial activity against various microorganisms,including gram-positive and gram-negative bacteria and theyeast Candida albicans (Table 1). mBD-14 was also able toefficiently kill methicillin-resistant Staphylococcus aureus andvancomycin-resistant Enterococcus faecalis. Burkholderia cepa-cia was not susceptible to mBD-14, an observation also madefor hBD-3 (15). Together, these data show that mBD-14 ex-hibits antimicrobial activity which is comparable to the antimi-crobial activity reported for hBD-3 (8, 15). Moreover, thekilling activities of mBD-14 and hBD-3 are influenced in asimilar manner by higher salt concentrations and at a lowerpH, thus further confirming their functional similarity (Table2). Interestingly, both peptides were able to kill S. aureus at 100mM NaCl but completely lost their activity against E. coli at100 mM NaCl (Table 2). In contrast, the susceptibility of E.coli to mBD-14 and hBD-3 was enhanced at pH 5.5, whereasthe susceptibility of S. aureus was reduced. The high suscepti-bility of E. coli to mBD-14 and hBD-3 at pH 5.5 may be causedby synergistic killing effects, because we observed that a pH of5.5 alone exhibited a slight killing effect against E. coli.
To obtain additional insight into the biological role of mBD-14, we analyzed its gene expression in various mouse tissues
(Fig. 2A). Reverse transcription-PCR analyses revealed geneexpression of mBD-14 in the tongue, thymus, tonsil, and kid-ney. All of these tissues have also been reported to expresshBD-3 (5). As shown in Fig. 2A, mBD-14 mRNA was alsoexpressed in skin and in primary keratinocytes, suggesting arole for mBD-14 in cutaneous defense. This is in concordancewith several studies indicating that hBD-3 participates in skininnate immunity (5, 7, 16). In addition, we found that geneexpression of mBD-14 in primary mouse keratinocytes wasinduced by treatment of the cells with gamma interferon(IFN-�) or transforming growth factor alpha (TGF-�) (Fig.2B), which were also identified as stimuli for the induction ofhBD-3 expression in human keratinocytes (4, 6, 11). Furtherparallelisms in the gene regulation of mBD-14 and hBD-3 canbe deduced from comparison of the promoter regions of thesegenes. It has been shown that the transcription factors NF-�B,STAT-1, and AP-1 are involved in both TNF-�/IFN-�- and
TABLE 1. Antimicrobial activities of mBD-14 and hBD-3 against various microorganisms
Straina MBCb (�M) ofmBD-14
LD90 (�M) of protein
mBD-14 hBD-3c
Gram-positive bacteriaStaphylococcus aureus ATCC 12600 1.2 0.08–0.15 0.15Staphylococcus aureus ATCC 33593 (MRSA) 2.4 0.30 0.6Staphylococcus epidermidis ATCC 14990 2.4 1.2 0.15Streptococcus pyogenes ATCC 12344 1.2 0.15 1.2Enterococcus faecalis ATCC 29212 1.2 0.08–0.15Enterococcus faecalis ATCC 51299 (VRE) 2.4 0.30
Gram-negative bacteriaEscherichia coli ATCC 11775 0.60–1.2 0.08–0.15Escherichia coli ATCC 25922 1.2 0.30 0.02Escherichia coli ESBL 4 (clinical isolate) 0.60 0.15 0.15Salmonella enterica serovar Typhimurium ATCC 13311 1.2 0.15Pseudomonas aeruginosa ATCC 10145 1.2–2.4 0.30 0.30Pseudomonas aeruginosa Va 6201/2000 (multiresistant; clinical isolate) 4.8 0.60 0.15Burkholderia cepacia ATCC 25416 �19 �19 �19Burkholderia cepacia ATCC 17759 �19 �19Burkholderia stabilis NCTC 13011 �19 �19
YeastCandida albicans ATCC 24433 1.21 0.60
a MRSA, methicillin-resistant Staphylococcus aureus; VRE, vancomycin-resistant enterococcus.b MBC, minimal bactericidal concentration.c LD90s of hBD-3 were calculated based on the work of Sahly et al. (15).
TABLE 2. Antimicrobial activities of mBD-14 and hBD-3 atdifferent ionic strengths and pHs
Strain Special conditionLD90 (�M) of protein
mBD-14 hBD-3
Staphylococcus aureus None 0.08–0.15 0.15ATCC 12600 100 mM NaCl 0.3 0.6
pH 5.5 2.4 4.82 mM MgCl2 0.08 NDa
Escherichia coli None 0.08–0.15 0.15ATCC 11775 100 mM NaCl �19 �19
pH 5.5 0.02 0.082 mM MgCl2 1.2 ND
a ND, not determined.
VOL. 52, 2008 NOTES 1877
on May 2, 2014 by guest
http://aac.asm.org/
Dow
nloaded from

bacterium-mediated hBD-3 gene induction (1, 9). Interest-ingly, analysis of the mBD-14 promoter region using the Geno-matix software program revealed that the promoter region ofmBD-14 also contains putative binding sites for NF-�B,STAT-1, and AP-1 (not shown).
In summary, this is the first report demonstrating thatmBD-14 exhibits potent antimicrobial activity, with an antimi-crobial spectrum similar to that reported for hBD-3. In addi-tion, as seen for hBD-3, the expression of mBD-14 in kerati-nocytes is induced by IFN-� and TGF-�. These results,together with the high sequence identity between mBD-14 andhBD-3, indicate that mBD-14 is the functional ortholog ofhBD-3.
Mouse models are useful for studying the in vivo relevanceof antimicrobial proteins. The knockout mouse for the humancathelicidin LL-37 ortholog CRAMP has been extensively usedto study the role of cathelicidins in innate immunity (3, 10).Another study showed that matrilysin-deficient mice lackedmature Paneth cell-derived cryptdins and were more suscepti-ble to orally administered bacteria (18). Therefore, studyingmBD-14 with mouse models could help us to further under-stand the role of antimicrobial peptides such as hBD-3 in hostdefense.
During revision of the manuscript, two other studies re-ported the identification of mBD-14 as the mouse ortholog ofhBD-3 (13, 17). The work of Taylor et al. (17) reported thathBD-3 and mBD-14 exhibit similar potent antimicrobial activ-ities against S. aureus and Pseudomonas aeruginosa which are
independent of intramolecular disulfide bonds. Rohrl et al.(13) reported that mBD-14 is active against E. coli but notagainst S. aureus, a finding that contrasts with our findings.However, it should be noted that whereas we used the maturepeptide for analysis of antimicrobial activity, Rohrl et al. usedan mBD-14–immunoglobulin fusion protein of approximately37 kDa. In addition, both reports showed that mBD-14 exhibitschemotactic activities similar to those of hBD-3. Taken to-gether, both reports confirmed that mBD-14 is the functionalortholog of hBD-3.
This work was supported by the Deutsche Forschungsgemeinschaft(grant SFB 617 and a Heisenberg fellowship to J.H.).
We thank Graziella Francesca Podda, Sylvia Voss, and AndreaHoelzgen for excellent technical assistance and Grace Chen for helpwith the manuscript.
REFERENCES
1. Albanesi, C., H. R. Fairchild, S. Madonna, C. Scarponi, O. De Pita, D. Y.Leung, and M. D. Howell. 2007. IL-4 and IL-13 negatively regulate TNF-alpha- and IFN-gamma-induced beta-defensin expression through STAT-6,suppressor of cytokine signaling (SOCS)-1, and SOCS-3. J. Immunol. 179:984–992.
2. Bendtsen, J. D., H. Nielsen, G. von Heijne, and S. Brunak. 2004. Improvedprediction of signal peptides: SignalP 3.0. J. Mol. Biol. 340:783–795.
3. Chromek, M., Z. Slamova, P. Bergman, L. Kovacs, L. Podracka, I. Ehren, T.Hokfelt, G. H. Gudmundsson, R. L. Gallo, B. Agerberth, and A. Brauner.2006. The antimicrobial peptide cathelicidin protects the urinary tractagainst invasive bacterial infection. Nat. Med. 12:636–641.
4. Garcia, J. R., F. Jaumann, S. Schulz, A. Krause, J. Rodriguez-Jimenez, U.Forssmann, K. Adermann, E. Kluver, C. Vogelmeier, D. Becker, R. Hedrich,W. G. Forssmann, and R. Bals. 2001. Identification of a novel, multifunc-tional beta-defensin (human beta-defensin 3) with specific antimicrobial
FIG. 2. Analysis of mBD-14 gene expression. (A) Reverse transcription-PCR analysis of mBD-14 and glycerolaldehyde-3-phosphate dehydro-genase (GAPDH) gene expression in various tissues. The following intron-spanning primers were used: mBD-14, forward primer, 5�-TCTTGTTCTTGGTGCCTGCT-3; reverse primer, 5�-CGACCGCTATTAGAACATCGAC-3�; GAPDH, forward primer, 5�-TGTTCCTACCCCCAATGTGT-3; reverse primer, 5�-TGTGAGGGAGATGCTCAGTG-3. (B) Mouse primary keratinocytes were stimulated with IFN-� or TGF-� (50ng/ml) for 24 h, and mBD-14 mRNA expression was analyzed by real-time PCR using the primers for mBD-14 and GAPDH as shown above. Barsrepresent the relative mBD-14 transcript levels normalized to GAPDH transcript levels. Results are from one representative experiment of threeindependent experiments and are presented as means � standard deviations.
1878 NOTES ANTIMICROB. AGENTS CHEMOTHER.
on May 2, 2014 by guest
http://aac.asm.org/
Dow
nloaded from

activity. Its interaction with plasma membranes of Xenopus oocytes and theinduction of macrophage chemoattraction. Cell Tissue Res. 306:257–264.
5. Harder, J., J. Bartels, E. Christophers, and J. M. Schroder. 2001. Isolationand characterization of human beta-defensin-3, a novel human induciblepeptide antibiotic. J. Biol. Chem. 276:5707–5713.
6. Harder, J., U. Meyer-Hoffert, K. Wehkamp, L. Schwichtenberg, and J. M.Schroder. 2004. Differential gene induction of human beta-defensins(hBD-1, -2, -3, and -4) in keratinocytes is inhibited by retinoic acid. J. In-vestig. Dermatol. 123:522–529.
7. Kisich, K. O., M. D. Howell, M. Boguniewicz, H. R. Heizer, N. U. Watson,and D. Y. Leung. 2007. The constitutive capacity of human keratinocytes tokill Staphylococcus aureus is dependent on beta-defensin 3. J. Investig.Dermatol. 127:2368–2380.
8. Maisetta, G., G. Batoni, S. Esin, W. Florio, D. Bottai, F. Favilli, and M.Campa. 2006. In vitro bactericidal activity of human beta-defensin 3 againstmultidrug-resistant nosocomial strains. Antimicrob. Agents Chemother. 50:806–809.
9. Menzies, B. E., and A. Kenoyer. 2006. Signal transduction and nuclearresponses in Staphylococcus aureus-induced expression of human beta-de-fensin 3 in skin keratinocytes. Infect. Immun. 74:6847–6854.
10. Nizet, V., T. Ohtake, X. Lauth, J. Trowbridge, J. Rudisill, R. A. Dorschner,V. Pestonjamasp, J. Piraino, K. Huttner, and R. L. Gallo. 2001. Innateantimicrobial peptide protects the skin from invasive bacterial infection.Nature 414:454–457.
11. Nomura, I., E. Goleva, M. D. Howell, Q. A. Hamid, P. Y. Ong, C. F. Hall,M. A. Darst, B. Gao, M. Boguniewicz, J. B. Travers, and D. Y. Leung. 2003.Cytokine milieu of atopic dermatitis, as compared to psoriasis, skin preventsinduction of innate immune response genes. J. Immunol. 171:3262–3269.
12. Pazgier, M., D. M. Hoover, D. Yang, W. Lu, and J. Lubkowski. 2006. Humanbeta-defensins. Cell Mol. Life Sci. 63:1294–1313.
13. Rohrl, J., D. Yang, J. J. Oppenheim, and T. Hehlgans. 2008. Identificationand biological characterization of mouse -defensin 14, the orthologue ofhuman -defensin 3. J. Biol. Chem. 283:5414–5419.
14. Rudolph, B., R. Podschun, H. Sahly, S. Schubert, J. M. Schroder, and J.Harder. 2006. Identification of RNase 8 as a novel human antimicrobialprotein. Antimicrob. Agents Chemother. 50:3194–3196.
15. Sahly, H., S. Schubert, J. Harder, P. Rautenberg, U. Ullmann, J. Schroder,and R. Podschun. 2003. Burkholderia is highly resistant to human beta-defensin 3. Antimicrob. Agents Chemother. 47:1739–1741.
16. Sorensen, O. E., D. R. Thapa, A. Rosenthal, L. Liu, A. A. Roberts, and T.Ganz. 2005. Differential regulation of -defensin expression in human skinby microbial stimuli. J. Immunol. 174:4870–4879.
17. Taylor, K., D. J. Clarke, B. McCullough, W. Chin, E. Seo, D. Yang, J.Oppenheim, D. Uhrin, J. R. Govan, D. J. Campopiano, D. Macmillan, P. E.Barran, and J. R. Dorin. 7 January 2008, posting date. Analysis and sepa-ration of residues important for the chemoattractant and antimicrobial ac-tivities of beta-defensin 3. J. Biol. Chem. [Epub ahead of print.] doi: 10.1074/jbc.M709238200.
18. Wilson, C. L., A. J. Ouellette, D. P. Satchell, T. Ayabe, Y. S. Lopez-Boado,J. L. Stratman, S. J. Hultgren, L. M. Matrisian, and W. C. Parks. 1999.Regulation of intestinal alpha-defensin activation by the metalloproteinasematrilysin in innate host defense. Science 286:113–117.
19. Zasloff, M. 2002. Antimicrobial peptides of multicellular organisms. Nature415:389–395.
VOL. 52, 2008 NOTES 1879
on May 2, 2014 by guest
http://aac.asm.org/
Dow
nloaded from