morphology and distribution of catecholaminergic amacrine cells in the cone-dominated tree shrew...
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 308:91-102 (1991)
Morphology and Distribution of Catecholaminergic Amacrine Cells in the
Cone-Dominated Tree Shrew Retina
BRIGITTE MULLER AND LEO PEICHL Max-Planck-Institut fur Hirnforschung, D-6000 Frankfurtm. 71, Germany
ABSTRACT The tree shrew (Tupaia belangeri) has a cone-dominated retina with a rod proportion of
only 5%. This is in contrast to the usual mammalian pattern of rod-dominated retinae. Rod bipolar cells are present at relatively low densities in the tree shrew retina, suggesting that a reduced, but normal, rod pathway might be preserved. The present study investigated another common constituent of the rod pathway, the dopaminergic amacrine cells, and analysed their morphology and distribution by light and electron microscopy.
Catecholaminergic (presumed dopaminergic) amacrine cells were labelled with an antibody against tyrosine hydroxylase (TH). Intense TH-immunoreactivity was found in perikarya and dendrites of a uniform amacrine cell population. TH-immunoreactive amacrine cell density varies across the retina from 10 cells/mm2 in the periphery to 40 cellsimm' in more central regions (mean cell density about 25 cells/mm2). The relatively large cell bodies are located exclusively in the innermost part of the inner nuclear layer. The dendrites form a dense plexus at the border between the inner plexiform layer and the inner nuclear layer. The finer dendritic processes contain many varicosities and form characteristic dendritic "rings" like those seen in other mammals. TH-immunoreactive processes also run between cell bodies in the vitread inner nuclear layer; a few extend into the sclerad inner nuclear layer and occasionally reach the outer plexiform layer (possible interplexiform cells). A few TH-immunoreactive processes are seen in the middle of the inner plexiform layer. Electron microscopy of TH-immunoreactive processes revealed conventional synapses onto somata and processes of unlabelled amacrine cells.
Key words: Tupuiu, dopamine, tyrosine hydroxylase immunocytochemistry, rod pathway, interplexiform cells
Most mammalian retinae studied so far are rod-domi- nated. In contrast, tree shrews (Tupaia belangeri) are diurnal omnivores with a highly cone-dominated retina (95% cones and 5% rods, Muller and Peichl, '89). A number of studies in rod-dominated mammalian species have dem- onstrated that rod and cone signals are processed in two separate pathways, which involve distinct populations of bipolar and amacrine cells. The cone signal passes through cone bipolar cells, which are presynaptic to ganglion cells and type I amacrine cells. The rod signal passes through the rod bipolar cells, which are presynaptic only to type I and type I1 (AII) amacrine cells but not to ganglion cells directly (Kolb and Famiglietti, '74; Kolb and Nelson, '83; reviewed by Daw et al., '90). In a recent paper we introduced the tree shrew retina as an appropriate test system for the phyloge- netic flexibility of the rod-associated circuitry. We have shown that rod bipolar cells exist but have a lower density and a different branching pattern than those of rod domi- nated retinae (Muller and Peichl, '91).
In the present work, we describe the morphology and distribution of the dopaminergic amacrine cells in the tree shrew retina. In other mammals, dopaminergic amacrine cells predominantly innervate the AII amacrine cells (cat: Pourcho, '82; cat, rat, and rabbit: Voigt and Wassle, '871, suggesting that they are an important interneuron in the rod pathway of mammalian retinae.
Dopamine is the predominant catecholaminergic neuro- transmitter in the retina. Significant amounts have been detected in the retinae of all vertebrate species studied so far, whereas noradrenaline and epinephrine have been reported as being undetectable or present only as traces (for review see Nguyen-Legros, '88). Antibodies directed against tyrosine hydroxylase (TH), the rate-limiting enzyme of catecholamine synthesis, have been used to label the dopa-
Accepted February 15,1991. Address reprint requests to Brigitte Muller, Max-Planck-Institut fur
Hirnforschung, Deutschordenstr. 46, D-6000 FrankfurtiM. 71, Germany.
O 1991 WILEY-LISS. INC.

92 B. MULLER AND L. PEICHL
minergic neurons in a variety of mammalian retinae (re- viewed by Nguyen-Legros, ’88). It has since been confirmed by the use of an antibody against dopamine that strongly TH-immunoreactive (TH-IR) cells are dopaminergic (Ver- saux-Botteri et al., ’86).
Here we have used a TH-antibody to identify the dopa- minergic amacrine cell population with both light micro- scopic and electron microscopic immunocytochemistry. Our aim was to see whether there were any features in the dopaminergic neurons of the tree shrew retina that could be related to its rod paucity. The somata of dopaminergic cells in other mammalian retinae are located mainly in the innermost row of the inner nuclear layer (INL) with only a few (2-5%) displaced to the inner plexiform layer (IPL) or the ganglion cell layer (GCL). Dopaminergic cells have the largest perikarya in the amacrine cell layer and wide-field, mainly unistratified dendritic fields. They either have all of their processes in the IPL (amacrine cells) or have addi- tional processes projecting to the outer plexiform layer (interplexiform cells). TH-IR cells represent 2-5% of the total amacrine cell population and their mean density is about 20-25 cells/mm2 in mammals (for review see Nguyen- Legros, ’88).
METHODS Tissue preparation
Twelve eyes from 11 adult tree shrews (T. belangeri) of either sex were used. All animals were obtained from breeding colonies at our institute and at the Deutsches Primatenzentrum, Gottingen. The animals were sacrificed by an overdose of pentobarbital given intraperitoneally. For light microscopy the eyes were enucleated directly postmor- tem. The eyeball was cut open around the equator, the anterior part with lens and vitreous was removed, and the posterior eyecup with the retina attached was immersed for a total of 2 hours at 4°C in the fixative (4% paraformalde- hyde (PA) in phosphate buffer (PB 0.1 M, pH 7.4)). After at least 1.5 hours in the fixative, the retina was removed from the eyecup. The pigment epithelium adhering to the retina was carefully brushed off and four to six radial cuts were made to flatten the retina. After fixation the retina was rinsed in PB for several hours. The tissue was cryopro- tected by immersion in a series of 10,20, and 30% sucrose in PB overnight and repeatedly shock-frozen and thawed in liquid nitrogen to improve penetration of the antibodies.
Immunocytochemistry was performed both on retinal wholemounts and sections. The retinae were sectioned
ac
GCL C
I INL IPL IS ONL OPL m N nuc PE S T TH TH-IR
Abbreviations
amacrine cell capillary ganglion cell layer inferior inner nuclear layer inner plexiform layer inner segments outer nuclear layer outer plexiform layer mitochondria nasal nucleus pigment epithelium superior temporal tyrosine hydroxylase TH-immunoreactive
either parallel to the vitreal surface (tangential sections), or perpendicular to it (transverse sections). To cut tangential sections the retina was cut into two pieces, frozen onto a mounting block, and cut into 40-pm-thick sections on a sliding microtome. For transverse sections, small tissue pieces were frozen in embedding medium (tissue tec, Reichert-Jung) and 10-pm thick sections were cut on a cryostat and collected on gelatin-coated slides. For vi- bratome sectioning the retinal pieces were embedded in 4% Agar Agar (Merck) in phosphate-buffered saline (PBS) and transversely sectioned with a vibratome at 70-100 ym. The vibratome sections were transferred to PBS and processed free floating, as were the retinal wholemounts and the tangential sections.
For electron microscopy three animals were perfusion fixed before enucleation with phosphate-buffered 0.1% glu- taraldehyde in 4% PA for 2.5 minutes. After enucleation the eyes were postfixed in 4% PA for 1 hour at 4°C. Immunocy- tochemistry was performed on vibratome sections.
Immunocytochemistry A mouse monoclonal antibody against tyrosine hydroxy-
lase (TH) from rat pheochromocytoma (Boehringer Man- nheim: clone 2/40/15; Cat. No. 1017 381) was used. This has been shown to react with samples of rat, chicken, quail, and bovine origin (Boehringer Mannheim data sheet) and recog- nizes a single protein band in the molecular weight range of 56,000-60,000 Dalton.
Immunoreactivity against tyrosine hydroxylase was dem- onstrated by the avidin-biotin peroxidase complex (ABC) method according to Hsu et al. (’81). All incubation steps were carried out at 4°C except for the cryostat sections, which were incubated at room temperature. The sections and retinal wholemounts were preincubated for 1 hour in 10% normal goat serum (NGS), 1% TritonX-100,1% bovine serum albumin, 0.1 M DL-Lysine in PB or PBS. Triton X-100 was omitted when vibratome sections were intended for electron microscopy. The primary antibody was diluted 1 : l O O for cryostat sections, 1 5 0 for the tangential and vibratome sections, and 1 : l O for wholemounts in a medium containing 3% NGS, 0.5% Triton X-100, 0.01% sodium azide in PB or PBS. Incubation time varied between 1 day (cryostat sections) and 5 days (wholemounts) a t 4°C.
After several rinses in PB or PBS, the sections and wholemounts were incubated in biotinylated goat anti- mouse IgG (Sigma B-7265) diluted 1:50 in 3% NGS, 0.5% Triton X-100 in PB or PBS for 2 days, or 2 hours for cryostat sections. For some cryostat sections the secondary antibody used was an FITC-conjugated anti-mouse IgG
Fig. 1. Micrographs of TH-immunoreactivity in transverse cryostat sections of the tree shrew retina, visualized by the indirect-immunoflu- orescent method with a fluorescein (F1TC)-conjugated secondary anti- body. In A, an immunoreactive cell body is present in the innermost INL and processes arborize in the outermost stratum of the IPL and the vitread INL. In B and C, retinal layers between the OPL and IPL of the transverse sections are shown. TH-IR processes form a prominent plexus in the outermost IPL and a three dimensional meshwork in the INL. At the IPLDNL border occasional dendritic “rings” are visible (big arrowheads in B and C). In the vitread INL the fluorescent processes form some pericellular baskets (arrows) around unlabelled amacrine cell bodies. A few processes reach the OPL (double arrowheads) but have not been observed to ramify there. Only few TH-IR processes are present deeper in the IPL at about 40% depth. The fluorescent spot in the lower right corner of A is a dirt particle. Scale bar is 50 km for A-C.
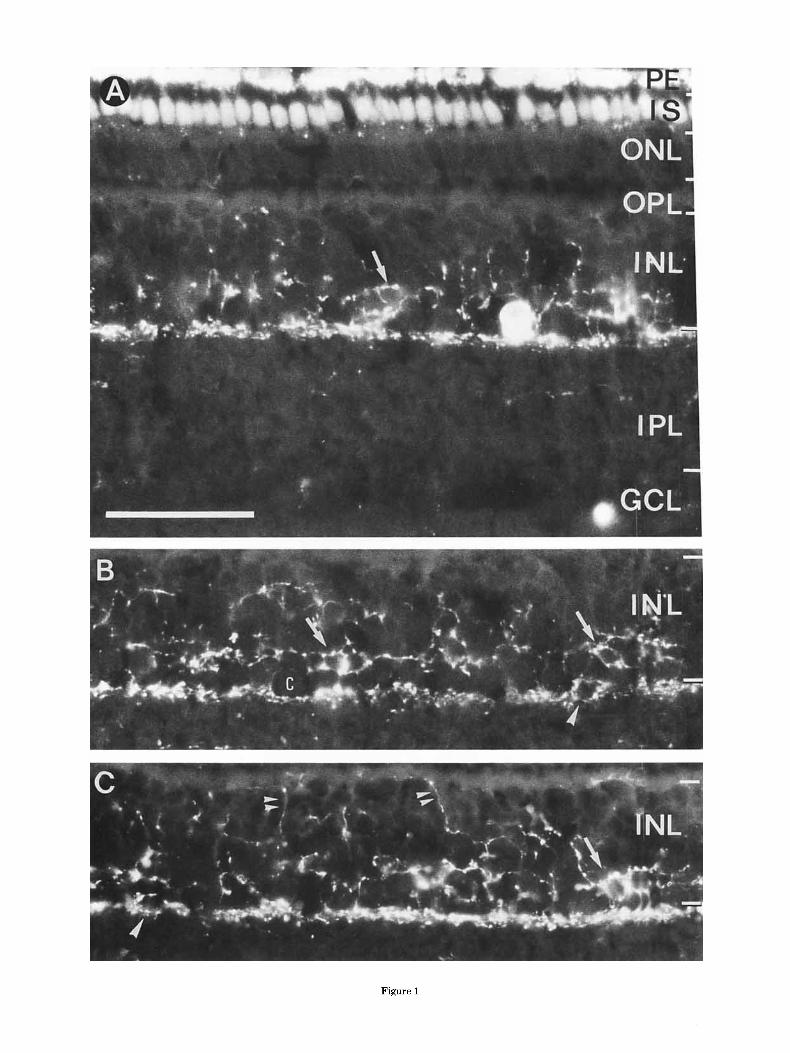
Figure 1
94 B. MULLER AND L. PEICHL
Figure 2

CATECHOLAMINERGIC AMACRINES IN TREE SHREW RETINA 95
(Sigma, diluted 1:200). After rinsing in PB or PBS, fluores- cent sections were coverslipped with glycerol. All other preparations were incubated in avidin peroxidase (Sigma A-3151) diluted 1:400 in PB or PBS for 1 day, or 1.5 hours for cryostat sections. The peroxidase was visualized with 3’,3-diaminobenzidine tetrahydrochloride (DAB, 0.05%; Sigma) and hydrogen peroxide (0.01%) in 0.05 M tris HC1 buffer (pH 7.6). The cryostat sections were rinsed in PB, dehydrated, and coverslipped with Permount (Fisher). Af- ter several washes the tangential sections and the whole- mounts were mounted, vitread side up, on gelatinized slides, dehydrated, cleared, and coverslipped with Per- mount.
The vibratome sections were postfixed in 2.5% glutaralde- hyde in 0.1 M cacodylate buffer (pH 7.4) for 2 hours. Sections intended for electron microscopy were osmicated (1% osmium tetroxide in 0.2 M cacodylate buffer for 1 hour). Finally, all vibratome sections were dehydrated, infiltrated in propylene oxide, and then flat-embedded in Epon 812 (SERVA). During the dehydration procedure the sections intended for electron microscopy were en bloc stained with 1% uranyl acetate in 70% ethanol for 1 hour. After curing for 2 days at 59”C, ultrathin sections (ca. 70 nm silver-gold interference colors) were cut on an ultrami- crotome (C. Reichert) for electron microscopy. They were mounted on single-slot grids coated with Formvar film and stained with lead citrate.
For light microscopy, serial semithin transverse sections (1 to 2 pm) were taken from the embedded vibratome sections, collected on gelatin-coated slides, and counter- stained with toluidine blue.
Quantitative evaluation Density maps of the labelled amacrine cells were obtained
from retinal wholemounts and a series of tangential sec- tions. Maps of wholemounted retinae were drawn with a low power objective (x1.25 or x2.5) and a Zeiss drawing tube. The retinal blood vessel pattern and the position of the optic disc were used to orient the retinae and mark the positions of areas to be analyzed. Drawings of tangential sections were obtained in the same way and the map of the entire retina was reconstructed from these drawings. Each section was examined, and in each area containing the innermost INL, counts of the labelled somata were made. Sample fields were analysed as in wholemounts with a ~1610.40 objective (Zeiss Neofluar; sample field size 610 x 610 Fm); 97 sample fields were analysed for the density map in Figure 7. Shrinkage was assessed by compar- ing retinal size before histological treatment with that after mounting, and areal shrinkage was about 10%. Density values given in the results are not corrected for this shrinkage.
The terms temporal, nasal, superior, inferior, and central are used here with respect to the optic disc, not with respect to the central area, which therefore is located in temporal retina.
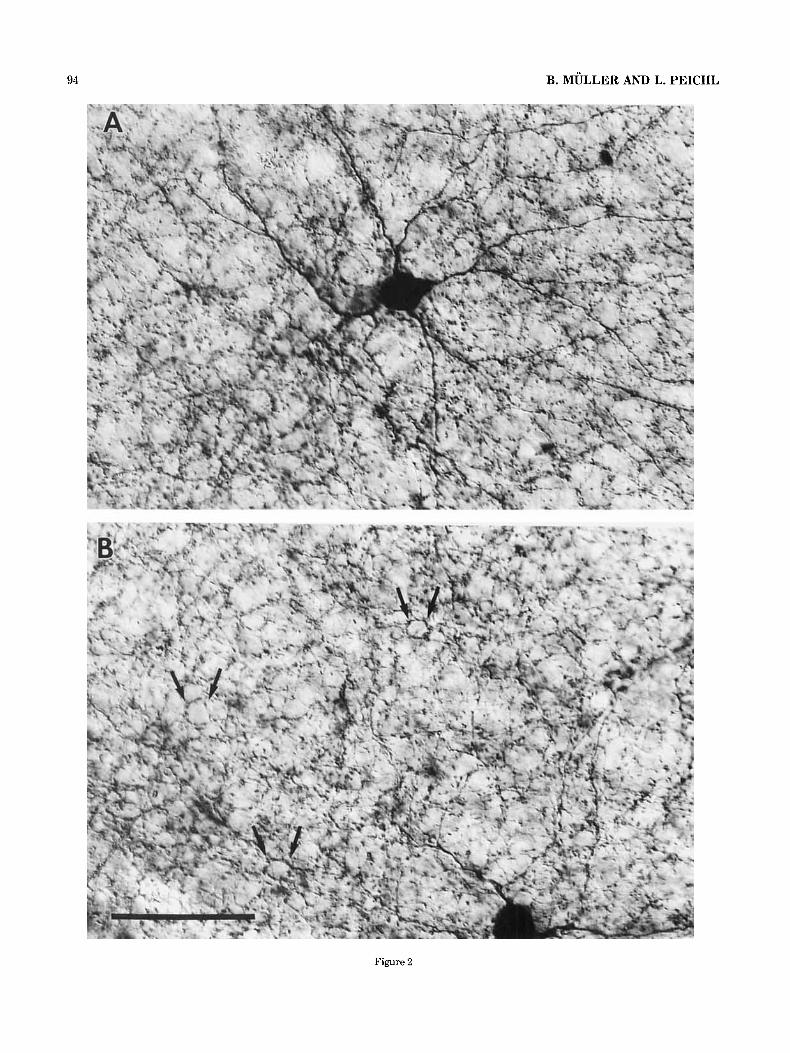
Fig. 2. TH-IR amacrine cells in tangential sections of the tree shrew retina. TH-immunoreactivity was visualized by the ABC-method. La- belled cell bodies and dendrites at the INLDPL border are in focus. After branching the main dendrites establish a rather dense plexus of varicose fibers. In this plexus occasional circular holes, the “dendritic rings,” are visible (arrows in B). They are formed by several varicose dendrites. Scale bar = 50 km for A and B.
RESULTS Morphology of TH-immunoreactive cells
Tyrosine hydroxylase was detected in a single morphological class of amacrine cells. Intense TH-immunoreactivity (TH-IR) was located in perikarya and varicose processes (Fig. 1). The large cell bodies were found exclusively in the innermost INL. The dendrites formed a prominent, continuous plexus at the INL/IPL border. In the INL, fine TH-IR fibers occured in a loosely meshed network between the amacrine cell somata, which form the two vitread rows of cell bodies in the INL of the tree shrew retina. Baskets of TH-IR processes were ob- served around some unlabelled somata of the vitread INL (arrows in Fig. 11, occasionally also in more sclerad regions. A few TH-IR processes extended to the sclerad INL, sometimes reaching the OPL. They were often observed to run laterally through the OPL (Fig. lC, arrowheads). The TH-IR fibers that coursed laterally or radially in the inner nuclear layer originated most commonly from TH-IR pro- cesses in the outermost stratum of the IPL, and less frequently from TH-IR somata. Additional immunopositive processes were found at the 40% level of the IPL (INL/IPL border: 0%, IPL/GCL border: 100%). These are also illus- trated in Figure 4F.
Evaluation of tangential, 40-pm-thick sections revealed the morphology of the TH-IR cells (Figs. 2A, 3). They were characterized by relatively large somata and processes with extensive fields. The somata were about 15 pm in diameter. Processes from neighbouring cells showed considerable overlap, so that any point of the retina was covered by the fields of at least three TH-IR cells. Two to four primary dendrites emerged from each soma and after branching established a rather dense plexus of varicose fibers at the INL/IPL margin, so that labelled somata and the main dendrites were observed in the same plane of focus (Fig. 2). Individual processes could usually be only followed for 250-300 pm from the soma before they merged in the dense plexus. The actual dendritic field of individual cells must be much larger and have many more branches than shown in Figure 3 to produce such a dense plexus.
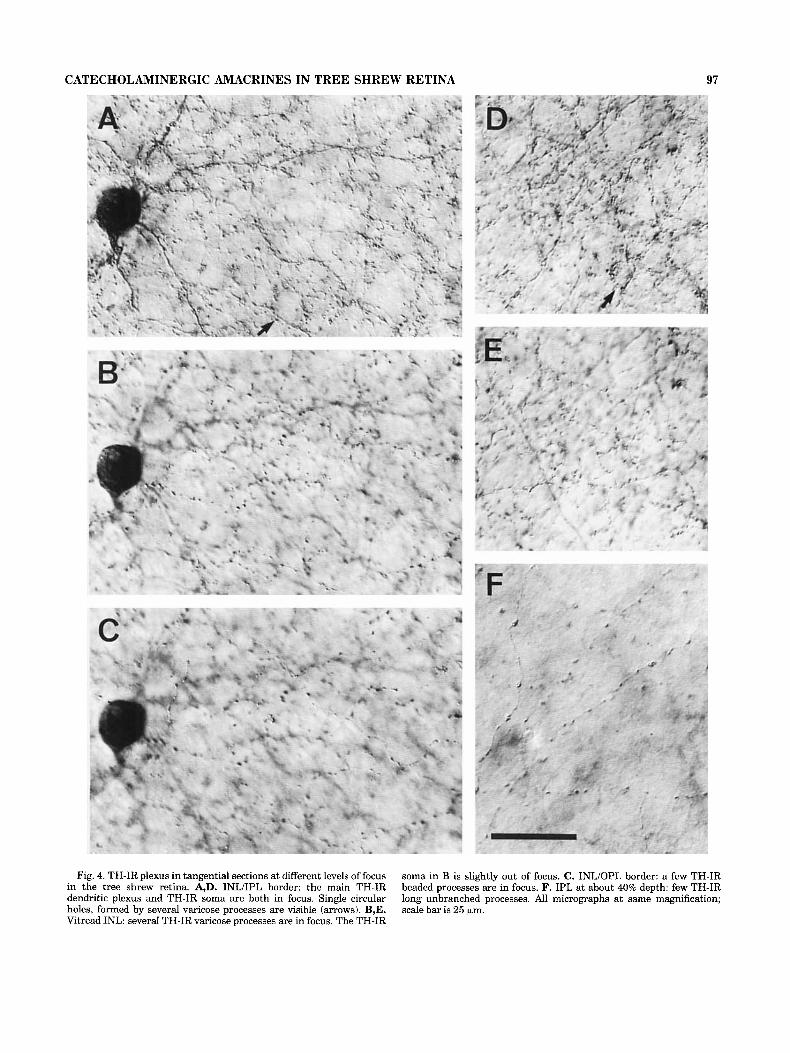
In tangential sections it became obvious that the den- dritic network in the outermost IPL was not as homoge- neous as it appeared in the transverse sections. More or less distinct circular holes surrounded by varicose TH-IR pro- cesses could be observed (Fig. 2B, arrows). The occurrence of such “ring-like’’ structures in the main catecholaminer- gic plexus is well known from other mammalian retinae. In the tree shrew retina, however, the distinct dendritic “rings” in the main TH-IR plexus were not as regularly distributed and not as numerous as in other mammals such as the cat (Fig. 4A,D). Besides the dendritic “rings” of the main plexus, there were additional “rings” in the second amacrine cell row of the INL (Fig. 4B,E), which consisted of single TH-IR processes (also obvious in transverse sections, c.f. Fig. 1). The tangential sections also show the sparse plexus of processes in the middle and outer parts of the INL (Fig. 4B,C). Some processes could be followed over a considerable distance through the OPL before they ended there or turned back to the INL or IPL.
Electron micrographs of the im- munocytochemically treated tree shrew retina revealed that the TH-IR processes of the main plexus at the IPL/INL border made synapses onto the cell bodies they approached (Fig. 5A-D). Tissue preservation, as judged from electron
Light microscopy.
Electron microscopy.

96 B. MULLER AND L. PEICHL
Fig. 3. Camera lucida drawing of the dendritic trees of TH-IR amacrine cells in a sample field. Only dendrites of somata contained in the field are drawn. TH-IR cell density is 28 cells/mm2 here. Individual processes were drawn until they merged in the dense plexus (dotted ends); the complete dendritic fields presumably are much larger than illustrated here.
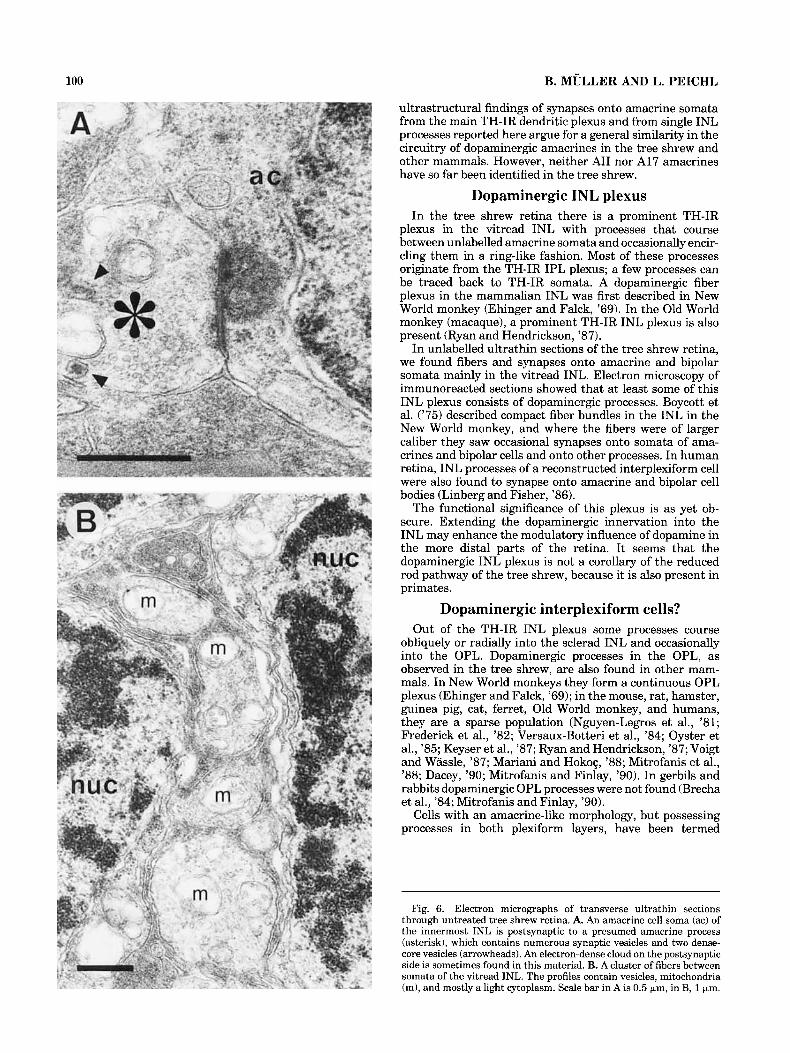
microscopy, was not optimal, since the TH-antiserum re- quired a low glutaraldehyde concentration in the fixative. Therefore samples of unlabelled, better preserved tissue were used in addition to assess the ultrastructure of synapses onto cell bodies in the amacrine cell layer. Several amacrine cell somata with conventional input synapses from vesicle-filled processes could be observed (Fig. 6A). Some of the presynaptic elements contained dense-core vesicles (arrowheads in Fig. 6A), which have also been found in identified dopaminergic processes (Frederick et al., '82; Kolb et al., '90). These conventional synapses were characterized by an aggregation of synaptic vesicles clus- tered close to the presynaptic membrane and electron- dense material associated with both the pre- and postsynap- tic membranes. In the immuno-stained material the DAB reaction product completely obscured cytoplasmic or- ganelles, and only the electron-dense postsynaptic mem- brane and the width of the synaptic cleft between the parallel pre- and postsynaptic membranes could be used as identification criteria of synapses.
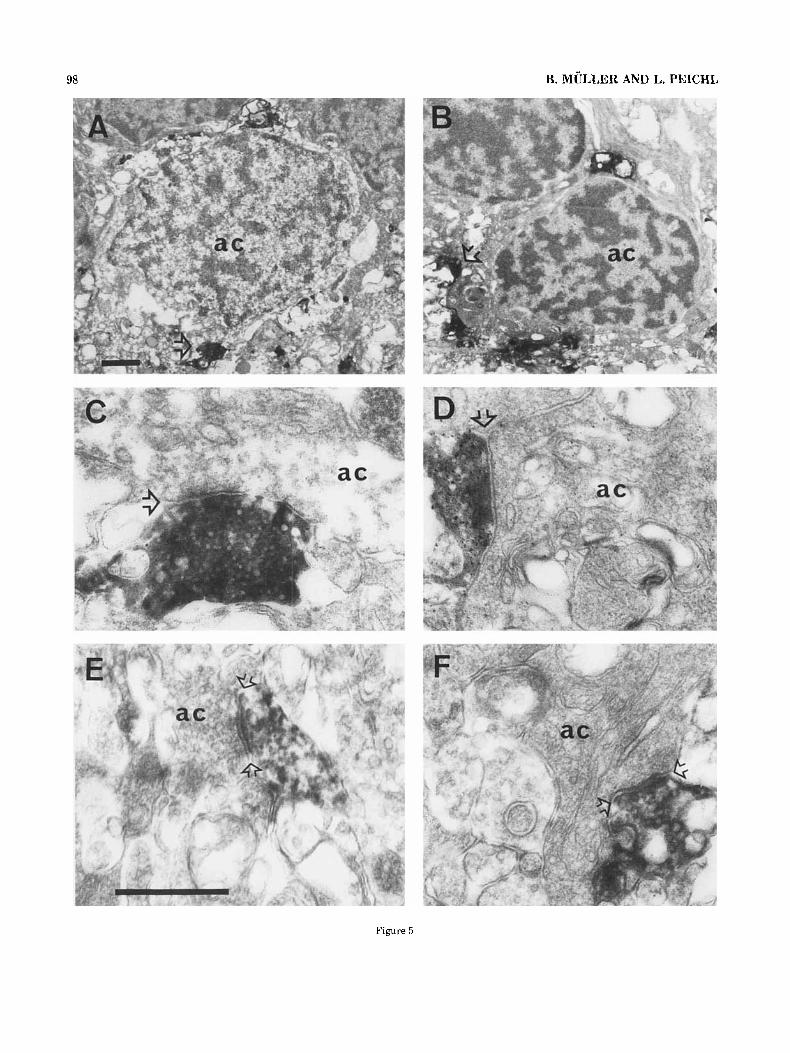
Figure 5A,B shows transverse sections of the vitread INL and the IPL below. In both micrographs an amacrine cell body is surrounded by the labelled TH-IR processes and receives synapses from them. The TH-IR process, which synapses on the cell body at the bottom of Figure 5A, is shown at higher magnification in Figure 5C; the TH-IR process that synapses on the cell body in the lower left corner of Figure 5B is magnified in Figure 5D. Presumed synaptic profiles were up to 0.7 pm in length and could be followed over three to six consecutive ultrathin sections. Figure 5C,D shows the synaptic sites optimally some sections away from those of Figure 5A,B.
The amacrine cell bodies illustrated in Figure 5A,B represent two types that were found to receive synaptic input from the TH-IR processes. They can be distinguished by the appearance of their nucleus. The type in Figure 5A contains many scattered chromatin granules besides some chromatin clusters embedded in an electron light matrix. The matrix of the other nuclear type in Figure 5B is relatively dark and contains large clusters of chromatin, the
CATECHOLAMINERGIC AMACRINES IN TREE SHREW RETINA 97
Fig. 4. TH-IR plexus in tangential sections at different levels of focus in the tree shrew retina. A,D. INL/IPL border: the main TH-IR dendritic plexus and TH-IR soma are both in focus. Single circular holes, formed by several varicose processes are visible (arrows). B,E. Vitread INL: several TH-IR varicose processes are in focus. The TH-IR
soma in B is slightly out of focus. C. INLiOPL border: a few TH-IR beaded processes are in focus. F. IPL at about 40% depth: few TH-IR long unbranched processes. All micrographs at same magnification; scale bar is 25 km.
98 B. MULLER AND L. PEICHL
Figure 5

CATECHOLAMINERGIC AMACRINES IN TREE SHREW RETINA 99
nucleus hence appears darker all together. Amacrine cells receiving synaptic input of TH-IR profiles are not restricted to the innermost row of the INL but are also located in the second row. Interestingly, all amacrine cells of the second row that received TH-IR contacts had a dark nucleus. Cell bodies were identified as amacrines by their location in the innermost INL and by often having an indented nucleus and containing more cytoplasm than bipolar cell bodies (Dowling and Boycott, ’66).
In addition to providing extensive input to amacrine cell somata (Fig. 5A-D), the TH-IR processes were seen to be presynaptic to the dendritic stalk of amacrine cells and unlabelled amacrine cell processes in the outermost IPL (Fig. 5E,F). No TH-IR processes were found to be presynap- tic to bipolar axon terminals or postsynaptic at ribbon synapses. Identification criteria for amacrine and bipolar cell processes were adopted from Dowling and Boycott (’66). The most conspicuous property of amacrine cell processes is their content of unevenly scattered vesicles, whereas bipo- lar cell axon terminals are characterized by numerous, evenly distributed vesicles, a few mitochondria, and synap- tic ribbons.
The fiber plexus in the INL (c.f. Figs. 1, 4) was also studied electron microscopically. Figure 6B shows an exam- ple of a cluster of fibers between somata in the vitread INL of unlabelled tissue. The profiles contain vesicles, mitochon- dria, and mostly a light cytoplasm, the latter simplifying their discrimination from Muller cell processes. Electron microscopy of TH-immunoreacted sections showed similar INL fiber clusters, containing labelled fibers, confirming the light microscopic observations. Thus there is a neuronal fiber plexus in the INL, which is at least partly composed of dopaminergic processes. We have not been able to deter- mine whether the few TH-IR processes detectable in the light microscope in the sclerad half of the INL and in the OPL make synapses. Only a few processes could be traced at the electron microscope, and no unequivocal synapses were found on them.
Distribution of TH-immunoreactive cells In both sectioned and whole retina preparations, TH-IR
cells were observed in all retinal regions. No TH-IR cells were found in the ganglion cell layer. The density and distribution of TH-IR amacrine cells were determined mainly from serial tangential 40-pm sections of several retinae. Because the tree shrew has a thick optic nerve fiber layer, penetration of the antibodies in wholemount prepara- tions was not sufficiently consistent across the retina for a complete topographic analysis.
Figure 7 illustrates the distribution of the TH-IR ama- crine cells in one retina. Figure 7A gives the numerical
Fig. 5 . Electron micrographs of transverse ultrathin sections through tree shrew retina after preembedding TH-immunocytochemis- try. A,B. TH-IR processes encircle amacrine cell bodies (ac) in the innermost INL. The nuclei of the contacted cell bodies differ in their appearance. The chromatin of the nucleus in B contains a more clumped chromatin, embedded in a more electron-dense matrix. Two presumed synapses of TH-IR processes (open arrows in A,B) are shown at higher magnification in C and D, respectively, from a few sections farther than A,B. E. TH-IR profile in the outermost IPL making a conventional synapse on an unlabelled, vesicle-filled ac process. F. TH-IR varicosity synapsing on the dendritic stalk of an ac at the INLiIPL border. The open arrows in C-F point to the synaptic cleft. Scale bars: in (A), 1 pm for A,B, in E, 0.5 p,m for C-F.
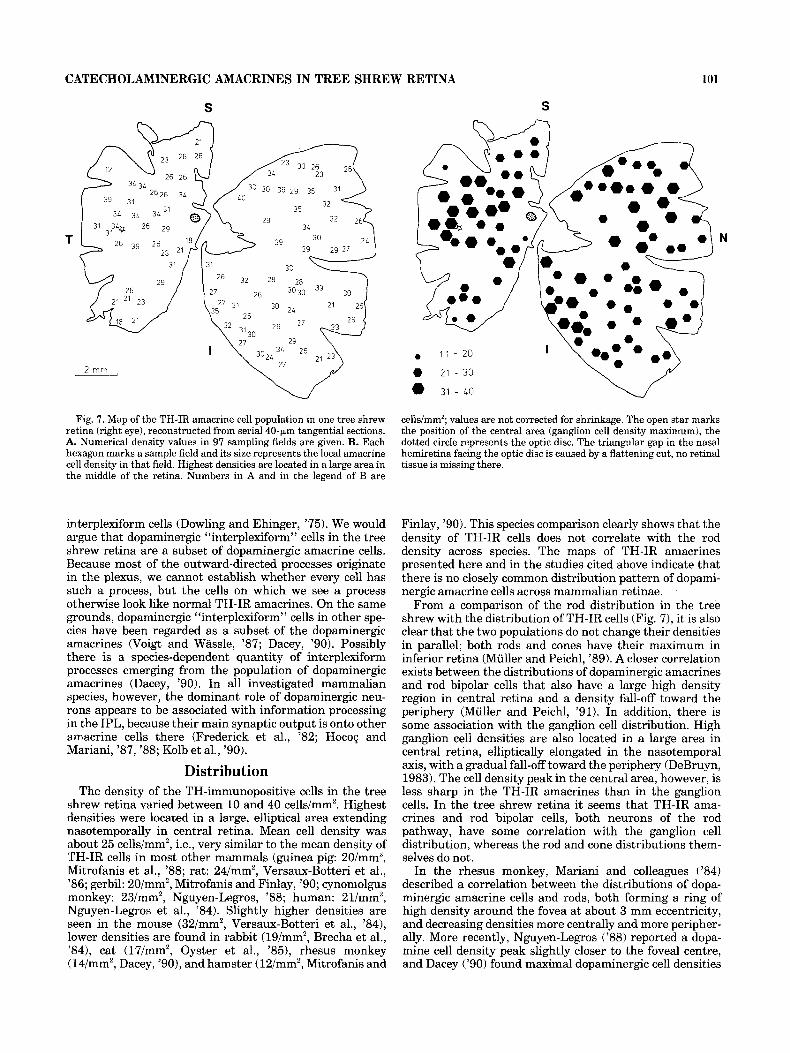
density values in the sampling fields, and Figure 7B pro- vides a more visual impression of the density changes, with each hexagon representing a sample field and its area being proportional to the cell density. TH-IR cell density varies between 10 and 40 cells/mm2. High densities are located in a large area in central retina, elliptically elongated in the nasotemporal axis, and densities decrease toward the reti- nal margins. The mean cell density is about 25 cells/mm2.
DISCUSSION Morphology
In the tree shrew retina, tyrosine hydroxylase immunore- activity (and therefore presumably dopamine) was located in one distinct type of stellate wide-field amacrine cell. The cell bodies were relatively large and exclusively located in the innermost row of the INL. The main dendritic plexus was narrowly stratified at the INL/IPL border and con- tained some ring-like structures. Occasionally, TH-IR vari- cose processes were found deeper in the inner IPL. In addition, fine laterally and radially oriented varicose pro- cesses coursed in the INL, forming an inner nuclear layer plexus. A few processes reached the OPL but were not observed to arborize there.
The TH-IR amacrine cells in the tree shrew retina resemble those of other mammals (Nguyen-Legros et al., ’81, ’84; Brecha, ’83; Brecha et al., ’84; Versaux-Botteri et al., ’84, ’86; Oyster et al., ’85; Keyser et al., ’87; Mariani and Hokog, ’88; Mitrofanis et al., ’88; Mitrofanis and Finlay, ’90). In all studied species the gross morphology of the TH-IR amacrine cells is very similar with stellate wide-field, more or less monostratified dendritic fields. Their cell bodies are large compared to those of other cells in the amacrine cell layer (15 pm versus 8-10 pm). Soma size is a specific characteristic of dopaminergic cells and rather homogeneous among mammalian species (for review see Nguyen-Legros, ’88).
The ring-like structures in the dendritic plexus at the IPL/INL border are a further characteristic of mammalian dopaminergic neurons. They were first described in the cat retina by Tork and Stone (’79). An electron microscopic analysis revealed that in the cat 3H-dopamine accumulating processes encircle the somata of A11 amacrine cells close to their primary dendritic stalk, contributing the majority of synaptic inputs to those somata as well as to the close-by “lobular appendages” of A11 dendrites (Pourcho, ’82). In the rat, rabbit, and cat, Voigt and Wassle (’87) provided direct evidence that AII amacrine somata occupy the TH-IR dendritic “rings,” and in the rat they showed synaptic contacts between those cells. These observations strongly suggested that dopamine exerts a modulatory influence on AII amacrines, one crucial interneuron in the retinal rod pathway. In the cat there may also be dopaminergic input to the somata of A17 amacrines, another neuron in the rod pathway (Kolb et al., ’90).
The relatively low density of dendritic “rings” in the tree shrew retina could indicate that the density of AII cells is lower than in rat, rabbit, and cat. Rod bipolar cell densities in the tree shrew are also up to a factor of 50 lower than in the cat, consistent with the low rod density (Muller and Peichl, ’91). Another explanation for the paucity of den- dritic “rings” in the TH-IR plexus of the tree shrew could be a different position of presumed A11 somata in the INL. If they did not protrude into the IPL as in other species, they would not form “holes” within the plexus. The
100 B. MULLER AND L. PEICHL
ultrastructural findings of synapses onto amacrine somata from the main TH-IR dendritic plexus and from single INL processes reported here argue for a general similarity in the circuitry of dopaminergic amacrines in the tree shrew and other mammals. However, neither AII nor A17 amacrines have so far been identified in the tree shrew.
Dopaminergic INL plexus In the tree shrew retina there is a prominent TH-IR
plexus in the vitread INL with processes that course between unlabelled amacrine somata and occasionally encir- cling them in a ring-like fashion. Most of these processes originate from the TH-IR IPL plexus; a few processes can be traced back to TH-IR somata. A dopaminergic fiber plexus in the mammalian INL was first described in New World monkey (Ehinger and Falck, '69). In the Old World monkey (macaque), a prominent TH-IR INL plexus is also present (Ryan and Hendrickson, '87).
In unlabelled ultrathin sections of the tree shrew retina, we found fibers and synapses onto amacrine and bipolar somata mainly in the vitread INL. Electron microscopy of immunoreacted sections showed that at least some of this INL plexus consists of dopaminergic processes. Boycott et al. ('75) described compact fiber bundles in the INL in the New World monkey, and where the fibers were of larger caliber they saw occasional synapses onto somata of ama- crines and bipolar cells and onto other processes. In human retina, INL processes of a reconstructed interplexiform cell were also found to synapse onto amacrine and bipolar cell bodies (Linberg and Fisher, '86).
The functional significance of this plexus is as yet ob- scure. Extending the dopaminergic innervation into the INL may enhance the modulatory influence of dopamine in the more distal parts of the retina. It seems that the dopaminergic INL plexus is not a corollary of the reduced rod pathway of the tree shrew, because it is also present in primates.
Dopaminergic interplexiform cells? Out of the TH-IR INL plexus some processes course
obliquely or radially into the sclerad INL and occasionally into the OPL. Dopaminergic processes in the OPL, as observed in the tree shrew, are also found in other mam- mals. In New World monkeys they form a continuous OPL plexus (Ehinger and Falck, '69); in the mouse, rat, hamster, guinea pig, cat, ferret, Old World monkey, and humans, they are a sparse population (Nguyen-Legros et al., '81; Frederick et al., '82; Versaux-Botteri et al., '84; Oyster et al., '85; Keyser et al., '87; Ryan and Hendrickson, '87; Voigt and Wassle, '87; Mariani and Hokog, '88; Mitrofanis et al., '88; Dacey, '90; Mitrofanis and Finlay, '90). In gerbils and rabbits dopaminergic OPL processes were not found (Brecha et al., '84; Mitrofanis and Finlay, '90).
Cells with an amacrine-like morphology, but possessing processes in both plexiform layers, have been termed
Fig. 6. Electron micrographs of transverse ultrathin sections through untreated tree shrew retina. A. An amacrine cell soma (ac) of the innermost INL is postsynaptic to a presumed amacrine process (asterisk), which contains numerous synaptic vesicles and two dense- core vesicles (arrowheads). An electron-dense cloud on the postsynaptic side is sometimes found in this material. B. A cluster of fibers between somata of the vitread INL. The profiles contain vesicles, mitochondria (m), and mostly a light cytoplasm. Scale bar in A is 0.5 pm, in B, 1 pm.
CATECHOLAMINERGIC AMACRINES IN TREE SHREW RETINA 101
S
Fig. 7. Map of the TH-IR amacrine cell population in one tree shrew retina (right eye), reconstructed from serial 40-pm tangential sections. A. Numerical density values in 97 sampling fields are given. B. Each hexagon marks a sample field and its size represents the local amacrine cell density in that field. Highest densities are located in a large area in the middle of the retina. Numbers in A and in the legend of B are
interplexiform cells (Dowling and Ehinger, ’75). We would argue that dopaminergic “interplexiform” cells in the tree shrew retina are a subset of dopaminergic amacrine cells. Because most of the outward-directed processes originate in the plexus, we cannot establish whether every cell has such a process, but the cells on which we see a process otherwise look like normal TH-IR amacrines. On the same grounds, dopaminergic “interplexiform” cells in other spe- cies have been regarded as a subset of the dopaminergic amacrines (Voigt and Wassle, ’87; Dacey, ’90). Possibly there is a species-dependent quantity of interplexiform processes emerging from the population of dopaminergic amacrines (Dacey, ’90). In all investigated mammalian species, however, the dominant role of dopaminergic neu- rons appears to be associated with information processing in the IPL, because their main synaptic output is onto other amacrine cells there (Frederick et al., ’82; Hocog and Mariani, ’87, ’88; Kolb et al., ’90).
Distribution The density of the TH-immunopositive cells in the tree
shrew retina varied between 10 and 40 cells/mm2. Highest densities were located in a large, elliptical area extending nasotemporally in central retina. Mean cell density was about 25 cells/mm2, i.e., very similar to the mean density of TH-IR cells in most other mammals (guinea pig: 20/mmz, Mitrofanis et al., ’88; rat: 24/mm2, Versaux-Botteri et al., ’86; gerbil: 20/mm2, Mitrofanis and Finlay, ’90; cynomolgus monkey: 23/mm2, Nguyen-Legros, ’88; human: 21/mm2, Nguyen-Legros et al., ’84). Slightly higher densities are seen in the mouse (32/mm2, Versaux-Botteri et al., ’84), lower densities are found in rabbit (19/mm2, Brecha et al., ’84), cat (17/mm2, Oyster et al., ’85), rhesus monkey (14/mm2, Dacey, ’go), and hamster (12/mm2, Mitrofanis and
S
. 1 ; - 20
0 21 - 30
0 31 - LO
N
cellslmm’; values are not corrected for shrinkage. The open star marks the position of the central area (ganglion cell density maximum), the dotted circle represents the optic disc. The triangular gap in the nasal hemiretina facing the optic disc is caused by a flattening cut, no retinal tissue is missing there.
Finlay, ’90). This species comparison clearly shows that the density of TH-IR cells does not correlate with the rod density across species. The maps of TH-IR amacrines presented here and in the studies cited above indicate that there is no closely common distribution pattern of dopami- nergic amacrine cells across mammalian retinae.
From a comparison of the rod distribution in the tree shrew with the distribution of TH-IR cells (Fig. 7), it is also clear that the two populations do not change their densities in parallel; both rods and cones have their maximum in inferior retina (Muller and Peichl, ’89). A closer correlation exists between the distributions of dopaminergic amacrines and rod bipolar cells that also have a large high density region in central retina and a density fall-off toward the periphery (Muller and Peichl, ’91). In addition, there is some association with the ganglion cell distribution. High ganglion cell densities are also located in a large area in central retina, elliptically elongated in the nasotemporal axis, with a gradual fall-off toward the periphery (DeBruyn, 1983). The cell density peak in the central area, however, is less sharp in the TH-IR amacrines than in the ganglion cells. In the tree shrew retina it seems that TH-IR ama- crines and rod bipolar cells, both neurons of the rod pathway, have some correlation with the ganglion cell distribution, whereas the rod and cone distributions them- selves do not.
In the rhesus monkey, Mariani and colleagues (’84) described a correlation between the distributions of dopa- minergic amacrine cells and rods, both forming a ring of high density around the fovea at about 3 mm eccentricity, and decreasing densities more centrally and more peripher- ally. More recently, Nguyen-Legros (’88) reported a dopa- mine cell density peak slightly closer to the foveal centre, and Dacey (’90) found maximal dopaminergic cell densities

102 B. MULLER AND L. PEICHL
around the foveal pit at about 0.5 mm eccentricity, i.e., central to the rod density peak. Thus it is at present not clear whether a correlation between the dopaminergic amacrine cell and rod distributions in rhesus monkey exists.
With respect to ganglion cells, Mitrofanis and coworkers (’88, ’90) conclude from a comparative survey of the gerbil, rat, hamster, guinea pig, rabbit, and cat that the distribu- tion of dopaminergic amacrines is more or less independent of the ganglion cell distribution.
There is a possible explanation for this lack of close correlations. It could be that for the dopaminergic amacrine cells associations with other neuron populations are not manifest in the cell density but in the density of the plexus and synapses. With the proposed modulatory role of dopa- mine, the low absolute density of the population, and the large dendritic fields, it is conceivable that the functional unit here is not the individual cell but parts of dendrites that could act locally. This idea has been put forward by Masland (’88) for the function of densely overlapping and long-range amacrine cell types. The situation might be even more complicated should Dacey’s (’90) hypothesis of func- tionally separate dendritic and axonal processes be con- firmed experimentally. Quantitative data on the density of the dopaminergic plexus and its synapses are, however, as yet not available in any retina to test these hypotheses.
The “normal” density of dopaminergic amacrine cells in the rod-depleted tree shrew retina might also indicate that they are not only integrated in the rod pathway but also in the cone pathway.
ACKNOWLEDGMENTS The skillful technical assistance of Heide Ahmed, Felici-
tas Boij, and Gong-Sun Nam is gratefully acknowledged. We thank Paul Martin for comments and for improving the English manuscript. This study was supported by a grant from Deutsche Forschungsgemeinschaft (DFG-SFB 451C7).
LITERATURE CITED Boycott, B.B., J.E. Dowling, S.K. Fisher, H. Kolb, and A.M. Laties (1975)
Interplexiform cells of the mammalian retina and their comparison with catecholamine-containing retinal cells. Proc. R. SOC. Lond. B 191:353- 368.
Brecha, N. (1983) Retinal neurotransmitters: Histochemical and biochemi- cal studies. In P.C. Emson (ed): Chemical Neuroanatomy. New York: Raven, pp. 85-129.
Brecha, N.C., C.W. Oyster, and E.S. Takahashi (1984) Identification and characterization of tyrosine hydroxylase immunoreactive amacrine cells. Invest. Ophthalmol. & Vis. Sci. 25:66-70.
Dacey, D.M. (1990) The dopaminergic amacrine cell. J. Comp. Neurol. 301t461-489.
Daw, N.W., R.J. Jensen, and W.J. Brunken (1990) Rod pathways in mammalian retinae. Trends Neurosci. 13:llO-115.
DeBruyn, E.J. (1983) The organization and central terminations of retinal ganglion cells in the tree shrew (Tupaia glss). PhD thesis, University of Nashville.
Dowling, J.E., and B.B. Boycott (1966) Organization of the primate retina: electron microscopy. Proc. R. Soc. Lond. B 255230-111.
Dowling, J.E., and B. Ehinger (1975) Synaptic organization of the amine- containing interplexiform cells of the goldfish and cebus monkey retinas. Science 188270-273.
Ehinger, B., and B. Falck (1969) Adrenergic retinal neurons of some New World monkeys. Z. Zellforsch. 100:364-375.
Frederick, J.M., M.E. Rayborn, A.M. Laties, D.M.K. Lam, and J.G. Hollyfield (1982) Dopaminergic neurons in the human retina. J. Comp. Neurol. 210:65-79.
Hokog, J.N. and A.P. Mariani (1987) Tyrosine hydroxylase immunoreactiv- ity in the rhesus monkey retina reveals synapses from bipolar cells to dopaminergic amacrine cells. J. Neurosci. 79785-2793.
Hokog, J.N., and A.P. Mariani (1988) Synapses from bipolar cells onto dopaminergic amacrine cells in cat and rabbit retinas. Brain Res. 461: 17-26.
Hsu, S.M., L. Raine, and H. Fanger (1981) Use of avidin-biotin peroxidase complex (ABC) in immunoperoxidase techniques. J. Histochem. Cy- tochem. 29:577-580.
Keyser, K.T., H.J. Karten, B. Katz, andM.C. Bohn (1987) Catecholaminergic horizontal and amacrine cells in the ferret retina. J. Neurosci. 7:3996- 4004.
Kolb, H., and E.V. Famiglietti (1974) Rod and cone pathways in the inner plexiform layer of cat retina. Science 186:47-49.
Kolb, H., and R. Nelson (1983) Rod pathways in the retina of the cat. Vision Res. 23:301-312.
Kolb, H., N. Cuenca, H.-H. Wang, and L. Dekorver (1990) The synaptic organization of the dopaminergic amacrine cell in the cat retina. J. Neurocytol. 19:343-366.
Linberg, K.A., and S.K. Fisher (1986) An ultrastructural study of interplexi- form cell synapses in the human retina. J. Comp. Neurol. 243:561-576.
Mariani, A.P., and J.N. Hokog (1988) Two types of tyrosine hydroxylase- immunoreactive amacrine cell in the rhesus monkey retina. J. Comp. Neurol. 276:8 1-91.
Mariani, A.P., H. Kolb, and R. Nelson (1984) Dopamine-containing amacrine cells of rhesus monkey retina parallel rods in spatial distribution. Brain Res. 322:l-7.
Masland, R.H. (1988) Amacrine cells. Trends Neurosci. 11:405410. Mitrofanis, J., and B.L. Finlay (1990) Developmental changes in the
distribution of retinal catecholaminergic neurons in hamsters and gerbils. J. Comp. Neurol. 292t480494.
Mitrofanis, J., A. Vigny, and J. Stone (1988) Distribution of catecholaminer- gic cells in the retina of the rat, guinea pig, cat, and rabbit: Independence from ganglion cell distribution. J. Comp. Neurol. 267:l-14.
Miiller, B., and L. Peichl (1989) Topography of cones and rods in the tree shrew retina. J. Comp. Neurol. 282:581-594.
Miiller, B., and L. Peichl (1991) Rod bipolar cells in the cone-dominated retina of the tree shrew Tupaia belangeri. Vis. Neurosci. (in press, Vol. 7).
Nguyen-Legros, J. (1988) Morphology and distribution of catecholamine- neurons in mammalian retina. In N. Osborne and G. Chader (eds): Progress in Retinal Research, Vol. 7. Oxford: Pergamon Press, pp. 113-147.
Nguyen-Legros, J., B. Berger, A. Vigny, and C. Alvarez (1981) Tyrosine hydroxylase-like immunoreactive interplexiform cells in the rat retina. Neurosci. Lett. 27:255-259.
Nguyen-Legros, J., C. Botteri, P. Le Hoang, A. Vigny, and M. Gay (1984) Morphology of primate’s dopaminergic amacrine cells as revealed by TH-like immunoreactivity on retinal flat-mounts. Brain Res. 295r145- 153.
Oyster, C.W., E.S. Takahashi, M. Cilluffo, and N.C. Brecha (1985) Morphol- ogy and distribution of tyrosine hydroxylase-like immunoreactive neu- rons in the cat retina. Proc. Natl. Acad. Sci. USA 82:6335-6339.
Pourcho, R.G. (1982) Dopaminergic amacrine cells in the cat retina. Brain Res. 252:lOl-109.
Ryan, M.K., and A.E. Hendrickson (1987) Interplexiform cells in macaque monkey retina. Exp. Eye Res. 45:57-66.
Tork, I., and J. Stone (1979) Morphology of catecholamine-containing amacrine cells in the cat’s retina, as seen in retinal whole mounts. Brain Res. 169:261-273.
Versaux-Botteri, C., J. Nguyen-Legros, A. Vigny, and N. Raoux (1984) Morphology, density and distribution of tyrosine hydroxylase-like immu- noreactive cells in the retina of mice. Brain Res. 301t192-197.
Versaux-Botteri, C., E. Martin-Martinelli, J. Nguyen-Legros, M. Geffard, A. Vigny, and L. Denoroy (1986) Regional specialization of the rat retina: Catecholamine-containing amacrine cell characterization and distribu- tion. J. Comp. Neurol. 243:422433.
Voigt, T., and H. Wassle (1987) Dopaminergic innervation of A11 amacrine cells in mammalian retina. J. Neurosci. 7:4115-4128.