molecular cloning and expression of cdna for rat pancreatic cholesterol esterase
TRANSCRIPT

Biochimica et Biophysica Acta, 1006 (1989) 227-236 227 Elsevier
BBALIP 53258
Molecular cloning and expression of cDNA for rat pancreatic cholesterol esterase
James A. Kissel ~, Robert N. Fontaine ~, Christoph W. Turck 2, Howard L. Brockman 3 and David Y. Hui
Departmou of Pathology and Laboratory Medicine, University of Cincinnati. College of Medicine, Cincinnati. OH. 2 The Howard Hughes Medical Institute, University of California, San Francisco, CA
and ~ The Hormel Institute, University of M#mesota, Austin. MN (U.S.A.)
(Received 18 April 1989)
Key words: Cholesterol esterase: cDNA
A full-length eDNA complementaD *o the rat pancreatic cholesterol esterase mRNA was isolated by screening a rat pancreatic eDNA expression library in ?~g~II vector with antibodies against the porcine pan teatic cholesterol esterase. The isolated cholesterol esterase eDNA is 2050 bp in lengfll and contains an open reading frame coding for a protein of 612 amino acids. A 20-amino acid hydrophobic leader sequence is predicted, based on the position of the first AT(; initiation codon upstream from the sequenced amino terminus of the isolated cholesterol esterase. The cholesterol esterase eDNA was subcloned into a mammalian expression vector, pSVL, for transfecfion stud|us. Expression of the eDNA in COS cells resulted in the production of bile salt-stimulated cholesterol estera~e. Comparison of the cholesterol esterase eDNA sequence with other proteins revealed that the pancreatic cholesterol esterase is identical to rat pancreatic iysophospholipase. The primary structure of cholesterol esterase displayed no significant homology with other lipeses, although the putative lipid interracial recognition site of G-X-S-X-G is present in the cholesterol esterase sequence. However, the cholesterol esterase sequence revealed a 63.amino-acid domaln which is highly homologous to the active site domain of other set |he esterases. These data suggest that cholesterul esterase may be a member of the serine esterase superge~le family. Analysis of the cholesterol esterase structure also revealed a repetitive sequence enriched with Pro, Asp, Gin, Set, and Thr residues at the C.terminal end of the protein. This sequence is reminiscent of the PEST-rich sequences in short-lived proteins, suggesting that cholesterol esterase may have a short half.life in vivo. Northern blot hybridization showed that the bile salt-stimulated cholesterol esterase mRNA is present in liver suggesting that th|s protein may also be synthesized by liver cells.
Introduction
The cholesterol esterase of the pancreas, also called carboxylester lipase or bile salt-stimulated lipase, plays an essential role in cholesterol absorption (see gef. 1 for revic.w). The cholesterol esterase is synthesized in the acinar cells and is stored in zymogen granules. The enzyme can be released into the intestinal lumen as a component of the pancreatic juice. In the lumen of the small intestine, and in the presence of bile salt, the cholesterol esterase aggregates and forms the active
The sequence data in this paper have been submitted to the EMBL/Genbank Data Libraries under accession nmnber X16054.
Correspondence: D.Y. Hui, Departmem of Pathology and Laboratory Medicine, University of Cincinnati, College of Medicine, 231 Be- thesda Ave, Cincinnati, OH 45267-0529, U.S.A.
enzyme which hydrolyzes dietary cholesteryl and fat- soluble vitamin esters before free cholesterol and other vitamins can be passed into the mucosa cells. The pancreatic cholesterol esterase is also found attached to the surface of intestinal mucosa and has been suggested to play a role in the re-esterification of cholesterol before secretion into the lymph [2]. The role of the pancreatic cholesterol esterase in these processes was demonstrated by observations that without pancreas or its secreted products, intestinal esterase activity and rate of cholesterol and vitamin absorption were dramatically reduced [2-4]. Immunological comparison of the cholesterol esterases has demonstrated the identity of the pancreatic cholesterol esterase with the enzyme found in the intestine, thus supporting the concept that this protein may be involved in both the hy~r,qy,is and re-esterification of dietary cholesteryl ester for absc~,,~- tion and transport [5].
0005-2760/89/$03.50 © 1989 Elsevier Science Publishers B.V. (Biomedical Division)

228
In addition to the cholesterol esterase, the pancreatic juice contains a variety of lipolytic enzymes. Several of the pancreatic lipases display similar substrate specifici- ties and co-factor requirements, suggesting a relation- ship between these proteins. For example, the pan- creatic cholesterol esterase can utilize phospholipids as substratet [6,7]. However, the enzyme is obviously dif- ferent from phospholipase A2, because the two proteins differ in suhstrate specificity and in molecular weight. Likewise, triacylglycerol lipase can be readily differenti- ated from cholesterol esterase, even though both en- zymes can hydrolyze short, and long-chain triacyl- 81yeerols [L7]. A pronounced difference in the posi- tional specificity of the enzymes for the substrate dis- tinguishes between the two proteins. Other pancreatic enzymes include the non-specific lipase [8], retinyl ester hydrolase [91, and lysophospholipase [10]. These en- zymes have characteristics similar to those of pancreatic cholesterol esterase. Whether the measured activities are due to the same enzyme or to related proteins remains to be determined. Understanding the structure of the cholesterol esterase will be helpful in delineating the structural and functional relationship of this protein, as well as its similarities to other lipases.
Purified pancreatic cholesterol esterase has been ob- tained from several mammalian sources. Most of the preparations show similar specificity and enzyme kinet- ics [1]. The purified protein from each species is a single polypeptide, but its extent of giycosylation is ~fferent between the different species. In all the species tested, the molecular weights of the protein moiety are similar, ranging from 69000 for the rat enzyme to 76000 for the human protein. This communication reports on the molecular cloning of the eDNA for rat pancreatic cholesterol esterase. The primary structure of the pro- tein, as deduced from sequencing of the eDNA, shows that the cholesterol esterase is identical to rat pancreatic lysophospholipase [11], but it is quite different from the other lipases. Results also show that the pancreatic cholesterol esterase may be a member of the serine esterase supergene family.
gxpenma
Preparation and sc~ening of rat pancreatic eDNA library The cold 8uanidine isothiocyanate procedure of Hart
et el, [II] was used to obtain RNA from f~_shly isolated rat pancreas. The mgNA was purified by two rounds of davmatography on oligo(dT)-cellulose column and used as template for oligo(dT) primed synthesis of the com- plimentary DNA. The second strand eDNA was synthe- sized using the Klenow fragment of DNA polymerase I after RHAase H digestion of the RNA. The double- stranded eDNA was ligated to EcoRI linkers and sub- cloned into EcoRl-digested ~,gtll vector. The eDNA was packaged into lambda phages using a commercially
available larabda packaging kit (Promega). Over 500000 recombinaat phages were screened with cholesterol esterase antiserum at a dilution of 1 : 5000 [12]. Positive reactions were identified by reaction with horseradish peroxidase-labeled anti-rabbit IgG.
Restriction mapping and nucleotide sequencing of the eDNA
The cholesterol esterase eDNA was isolated from positive lambda phages by digestion with EcoRI. The eDNA inserts were separated from the ~,gtll vectors by electrophoresis on 1~ low melting Agarose gels and were subeloned into pUC-13 plasmid for restriction mapping analysis. The plasmids containing the cholesterol esterase eDNA were digested with various restriction enzymes and the digests were analyzed on 6% polyacrylamide gels. Various ethidium bromide.stained bands corresponding to the digested eDNA were ex- cised from the gel. The eDNA fragments were isolated by electr ~qution and ligated to M13mpl8 or M13mpl9 vectors by standard procedures [13i. The recombinant plasmids were used to transform Escherichia coil DHSaF'/cells, single-stranded DNA was prepared, and sequencing was carried out by the didcoxy chain termination method [14] using the modified bacterio- phage T7 DNA polymerase-Sequenase (United States Biochemical Corp.). Sequencing overlaps were obtained from the various clones as indicated in Fig. 1. Sequence data analysis was performed using the BIONET System of Intelligenetics, Inc. (Palo Alto, CA).
Preparation of cholesterol esterase peptides and protein sequencing
Porcine pancreatic cholesterol esterase was purified as described previously [15]. For cyanogen bromide cleavage, 1 mg of the purified protein was dissolved in 705 formic acid and treated with 300 nag of cyanogen bromide for 24 h at ambient temperature. Tl'.e digested peptides were separated by C-18 reversed-phase high- pressure liquid chromatography and the eluted frag- ments were detected by the pest-column fluorescamine derivatization technique as previously described [16]. Protein sequences were determined from the amino terminus of the native cholesterol esterase and from three CNBr fragments (approximately 200 pmol each) using a gas-phase microsequenator.
Preparation of oligonucleotide probes and Southern Blot- ting of the cDNA
The peptide sequences were used to design oligo- nucleotide probes according to the codon preference table as described by Lathe [17]. Two probes of 57 nucleotides each, corresponding to the N-terminal se- quence of the native protein and one CNBr peptide, were synthesized using an Applied Biosystems 380A synthesizer. The oligonucleofides were purified by elec-

trophoresis through 10% polyaerylamide gels containing 7 M urea. The probes were labeled at the 5'-end with [~,-32p]ATP (Amersham) and T4 polynudeotide kinase (New England BioLabs). The radiolabeled oligonueleo- tide probes were separated from unicorporated nudeo= tides by Elu-tip columns (Schleicher and Sehudl) and were used for hybridization with EcoRI-digested k eDNA previously immobilized on nitroeelllulose paper following electrophoresis in 1% Ag:aose gels. The condi- tions of hybridization were 20% formamide/5 × concentrated SSC (1 x SSC is 150 mM NaC1/15 mM sodium citrate)/5 × Denhardt's solution/0.1% SDS/2 mg/ml of denatured salmon sperm DNA at 42°C. Filters were washed at increasing stringency with the final washes performed with 1 × SSC and 0.1% SDS at 42"C. The filters were exposed to Kodak XAR-2 films for 18 h to identify positive reactions.
Expression of cholesterol esterase cDNA The 2.1-kb insert of cholesterol esterase eDNA was
isolated from the 2~ clones by digestion with EcoR1. The isolated eDNA was made blunt-ended by a filled-in reaction with the Klenow fragment of DNA polymerase I m the presence of all four deoxynudeotides. The t~tunt-ended eDNA was ligated to the Sinai site of the ~aammalian cell expression vector pSVL (Pharmacia Fine Chemicals). The ligated plasmid was used to trans- feet E. coli DH5ot cells for propagation. Recombinant plasmids were purified by centrifugation on a CsC1 gradient. The recombination pSVL plasmid was analyzed by digestion with the restriction endonudease Sad. Both the pSVL vector and the cholesterol esterase eDNA displayed a single restriction site for this en- zyme. Therefore, electrophoresis of the Sacl-digested recombinant plasmid yielded infor~auon concerning the orientation of the eDNA insert in the vector. The plasmid containing the eDNA in the proper orientation, as well as the native pSVL plasmid, were used to transfect COS cells by the calcium phosphate precipit~,- tion method [18]. After 48 h of incubatl,3a, cell,., were washed with saline.EDTA and were harvested with buffer containing 20 mM Tris-HC1 (pH 7.5), 5 mM glutathione and 250 mM sucrose. The cell suspension was sonicated with a Branson sonifier, and cell debris was removed by centrifugation at 10000 x g for 10 min at 40C. The supernatant fraction was assayed im- mediately for cholesterol esterase activity as described [19]. In a typical experiment, 50 #1 of the sample was added to 150 lal of a solution contairdng 50 mM Tris-HCl (pH 7.5), 0.8~ tauroeholate, and 10 #M cholesteryl [~4C]oleate. After incubation for 1 h at 37°C, the reaction was terminated by adding 1.05 ml of 50 mM sodium carbonate and 50 mM sodium borate (pH 10). The reaction product, [~4C]oleate, was extracted as the free fatty acid with 3.25 ml of methanol /chloroform/ heptane (1.41:1.25:1.0) and was quantitated by liquid
229
scintillation. An aliquot of the cell extract was used for precipitation with trichloroacetie acid and the protein concentration was estimated by the Lowry method [20].
Northern blot ai~alysis of RNA from d~fferent rat tissues Total cellular RNA was isolated from freshly pre-
pared pancreas, intestine, and liver of a male Sprague- Dawley rat using the cold guartidine isothJoeyanate method [11]. Then, 20 #g of the cellular RNA was dectrophoresed in 1% Agarose gels in the presence of formaldehyde as described [13]. The RNA was s t ~ e d with ethidium bromide and the integrity of the isolated RNA was verified by the presence of 28S and 18S rRNA bands in the ratio of 2 : 1. The RNA was trans- ferred to nitrocellulose paper and hybridized with 32p. labeled cholesterol esterase eDNA. Hybridization was carried out for 18 h at 42°C as described above for Southern blotting of the eDNA, except that 50% for- mamide was used. Non-specific reaction was removed by washing four times at 50 °C with a buffer containing 0.1 × SSC and 0.1% SDS. Positive hybridization was identified l~y exposure to Kodak XAR-2 films at - 70 °C for 18 h.
Results
A rat pancreatic eDNA library, constructed in the kg t i l expression vector [21], was screened with an antibody prepared against porcine pancreatic cholesterol esterase. This antibody cross-reacted with cholesterol esterase in the rat pancreas and showed a single band on immunoblot analysis with total rat pancreatic pro- teins. The speciI.ic antibody was used at 1 : 5000 dilution t~ screen the eDNA library, lmmunoscreening of ap- proximately 500000 recombinant bacteriophages re- suited in identification of over 100 positive clones. The abundancy of positive reactions was consistent with the abundancy of cholesterol esterase in this tissue [22] Twenty-five positive clones were selected at random for characterization. Digestion of the phage DNA with EcoRI showed that the clones contained inserts ranging in size from 800 to 2000 bp. The eDNAs were suo- cloned into pUC-19 vector for further analysis. The relatedness of the cDNAs was investigated by using the largest eDNA (k done 5) as probe in hybridization experiments with the various cDNAs. Results showed that the 2.1 kb insert of k clone 5 hybridized to all the cDNAs (data not shown). Thus, these cDNAs were likely to derive from the same mRNA. Results obtained from restriction mapping of the cDNAs were also con- sistent with this conclusion. The relationship and partial restriction maps of the cDNAs are shown in Fig. 1.
In an effort to confirm the authenticity of the iso- lated eDNA for rat pancreatic cholesterol esterase, par- tial protein sequencing information was obtained from purified porcine pancreatic cholesterol esterase. The

230
k . C l e a e . I t 4 . . . . . . . . . . . . . . 4
• t k - C ~ e ' 2 ~. . . . . . . . . . . . . . . ~, ,~ ........... -4
~.c~., : p ,~ K, s + ....... :...~
B P BH g S k -CIo~ l .3 ,5 I I I I I I I I [ e
+....~:--~-t:~. ..... +~ ......... '"'~:::::::::L i ..... ,I
i | T"" i
' ' 400 800 1 ; 0 0 1~)0
Fig. 1. Restriction m p and :~iuencing strategy for the rat pancreatic ¢hoksterol estera~ eDNA. Five recombinant bacteriophages contain- ing the eholmteml atorau ~luence were isolated. The eDNA inserts were isolated by dile~tion ~th EcoRl. Sequencing of the eDNA, denoted as X ¢loNt 1-$, were performed in the directions and with appro~mate lengths as indicated by the arrows. The partial restriction map ~hows the overleq~ping t~ons of the various eDNA and the sites used for subcloning into MI3 vector for sequencing analysis. B, Ball;
H, Hlncll~ g. Kpnl; P, Pstl; [L SacI.
cholesterol esterase was subjected to cyanogen bromide hydrolysis and the digested peptides were separated by high.pressure liquid chromatography as described in Experimental Procedures. Three individual polypeptides were isolated for micro,sequencing analysis. The results of this experiment are shown in Table I. The sequence from one of the peptides (Ix'pride 31) agreed favorably with the N-terminal sequence of the mature protein (Table I, and Refs. 15 and 23) while the other two sequences (peptides 29 and 33) were presumably inter- ned sequences of cholesterol esterase, The partial protein sequencing data was used to design oligonucleotide probes for hybridization with the various cDNAs (Table I). The probes were synthesized based on codon usage frequency table [17], Results of the hybridization experi- ments indicated that all the eDNA insem reacted with the probes corresponding to the internal peptide se- quence. Therefore, the isolated cDNAs were likely to contain sequences for the pancreatic cholesterol esterase. Furthermore, the 2,1-kb inserts from clones 3 and 5 hybridized to the probe made according to the N-fermi- hal sequence of the protein suggesting that the 2,1-kb insert may represent a full-length cDNA.
The complete structure of the cholesterol esterase mRNA was determined from nucleotide sequencing of the eDNA. The strategy for sequencing various restric- tion fragments of the eDNA is shown in Fig. 1. The complete nucleotide sequences of both strands were obtained by sequencing the cDNAs obtained from the different clones. In all instances, the sequences of over- lapping regions of the different clones were identical. Overlapping sequence information was also obtained with restriction fragments obtaiued from the same eDNA (X clone 5). Results, as shown in Fig. 2, indi- cated that the eDNA was 2057 nucleotides in length and contained 17 nucleotides upstream from first ATG codon. The nucleotide sequence surrounding this ATG cedon fits the optimal consensus sequence, AX- XATG@, for initiation of translation by eukaryotic ribosomes as described by Kozak [24]. The polyadenyla- teen signal, AATAAA, is located at 13 nucleotide up- stream from the poly(A) tail. The primary structure of the cholesterol esterase was deduced from the nucleo- tide sequence. Beginning with the consensus ATG ini- tiation codon, a long open reading frame coding for a protein with 612 amino acid residues was observed. The termination codon TAA was found at nucleotide 1854 with a 3'.untranslated region of 203 bp. A 20-amino-acid hydrophobic leader sequence was predicted based on the position of the first ATG initiation codon upstream from the sequenced amino terminus of cholesterol esterase from rat [23] and porcine pancreas (Table I). The sequences corresponding to the two CNBr peptides of porcine cholesterol esterase could be identified within the coding region and were both preceded by methionine residues. The predicted molecular weight of the trans- lated nonglycosylated protein is 67048. A possible ?7- 81ycosylation site was located at residues 187-189. The rat pancreatic cholesterol esterase is known to be a glycoprotein [1]. Whether the post-translational modifi- cation occurs at this N-linked site remains to be de- termined.
The authenticity of the 2.1-kb cholesterol esterase
eDNA was further verified by expression studies in tissue cultured cells. The full-length cholesterol esterase
TABLE !
Amino ectd ~i~eeces of ~ estcea~ peMides and their ¢o~lementar~ oligonucleotide probes
N-terminallcquence Ate Lys Leu Gty Set Vet Tyr Thr Gtu SLy Gty CNSrpeptide31 ALe Lys Leu GLF Set Vet Tyr Thr Gtu Sty GLy C~ngDNA~lU~Cice 5'-GCC AA6 CTG GGC TCC GTG TAC ACA GAG GGC GGC Olil~li~¢lh~be $'-CGG TTC GAC CCG AGG CAC ATG TGT CTC CCG CCG
CNBrpepfide29 (eet )Pro Vat Tyr Pro Ser Trp Vat GLy Ats Asp C-~pomli~ONA~xluen~ 5*-ATG CCT GTC TAC CeA TCI TG6 GTG GGC GCT GAC OlilpxldeolidePmbe 3'-TAC GG& CAG ATG GGT AGI ACC CAC CCG CGA CTG
CNBrpeplide33" Ate l i e Ate Leu Vat ILe Thr Asn ILe ALe Ata
Phe Vat Gtu Phe VsL Gtu TTT GTG GAG AAA CAC CTC
His Ate Asp CAT GCT GAT GTA CGA CTA
Phe SLy GLy
ELy VaL Asn Lys Lys GLy Vet Ash Lys Lys GGC GTG AAC AAG AAG CCG CA¢ TTG TTC TTC
Asp Leu GLn Tyr VaL GAC CTI CAG TAT GTT CTG GAI GTC ATA CAA
Asp Pro bys Leu
a No o~o~.~ec~de probe was made with this pepfidc sequence.

1 -20
81 2
159 28
237 54
315 80
393 106
471 132
549 158
627 184
705 210
783 ~36
262
939 288
1017 314
1095 340
1173 ~66
1251 392
1329 418
1407 444
14~5 47O
1563 496
1641 522
1719 548
AGAGGCAGAC ACTCACT
AAG TTG GGT GCT GTG L_M s Leu GIy Ala Val
GTT GAC ATC TTC AAG Val Asp Ile Phe Lys
ACA CTG AAG GCT ACA Thr Leu Lys Ala Thr
TGC CTC TAT CTC AAC Cys Leu Tyr Leu ASh
GGA GGT GCC TTC CTC Gly Gly Ala Phe Leu
ACT AGA GGC AAT GTC Thr Arg Gly Ash Val
CTT CCA GGT AAC TTT Leu Pro Gly Asn Phe
GAC CCC GAT AAC ATC Asp Pr___oo Asp Ash ~le
AAG GGC CT~ ATC CGG LMs Oly Leu Xle Arg
TGG GCC AAA ACG ATC Trp Ala Lys Thr Tie
GAT CCC CGA GCC TTG Asp Pro Arg Ala Leu
ATC CCT GTC GTC GAT Ile Pro Val Val Asp
GCG GGT ATT AAT GAC Ala GIy Ile Asn Asp
ACA GAG GAG GAC TTC Thr Glu Glu Asp Phe
ATC TAC ACT GAG TCC Ile TyE ThE GIu Set
CTC TTC CTG ATC CCC Leu Phe Leu Ile Pro
TTT TCC CAC CCT TCA Phe Set His Pro Ser
GGG AAG CCC TTT GCC Gly Lys Pro Phe Ala
AAC 'FI~ GCC AAG AGT Asn Phe Ala Lys Ser
GGT AAC TAC CTG GAC GIy Asn Tyr Leu Asp
TGG GCT GTG ACA TTC Trp Ala Val Thr Phe
GTC CCA CCT ACA GAC Val Pro Pro Thr Asp
ATG GGG CGC CTG GAG GTT CTG MET Gly Arg Leu Glu Val Leu
TAC ACA GAA GGC GOT TTT GTG TMr Thr Ol_ u Gly Gky_phe Va!_
GGC ATC CCC TTT GCT ACC GCC Oly Zle Pro Phe A1a Thr Ala
GAC TTC AAG AAA CGA TGT CTA Asp Phe Lys Lys Arg Cys Leu
ATC TGG GTC CCT CAG GGC AGG ~le Tzp Val Pro Gln GIy Arg
ATG GGG TCT GGC CAG GGA GCC Met GIy Set GIy Gln Gly Ala
ATT GTG GTC ACC TTC AAC TAC Ile Val Val Thr Phe ASh Tyr
GGA CTT CGA GAT CAG CAC ATG GIy Leu Azg Asp Gln His Met
ACC ATC TTT GGG GAA TCT GCT Thr Zle Phe Gly Glu Set Ala
CGA GCC ATC AGT CAG AGT GGT Arg Ala Ile Ser Gln Set GIy
GCT AAG AAG GTG GGA TGC CCC Ala Lys Lys Val Gly Cys Pro
ACA CTG GCC TAC AGG TTG CCC Thr Leu Ala Ty~ Arg Lcu Pro
GGT ~AC TTC ATT CCT GAT GAT Gly Asp Phe Ile Pro Asp Asp
ATG GAT GGC CAC CTG TTT GCT Met Asp Gly His Leu Phe Ala
TAC A3G CTA GTC AGT GGA CAC Tyr Arg Leu Val Sot GIy His
TGG GCC CAG GAC CCG TCC CAG Trp Ala Gln Asp Pro Sor Gln
ACA GAG ATG GCT CTG GCC CAG Thr Glu Met Ala Leu Ala Gln
CGA ATG CCT ATC TAC CCA AAA Arg Met Pro Ile Tyr Pro Lys
ACC CCA CTG GGC TAC CGG GCC Thr Pzo Leu Gly Tyr Arg Ala
GGG GAC CCC AAC ATG GGC AAC GIy ASp Pro Ash Met Gly ASh
ATC AAT AAG AAA ATA ACC AGC Ile ASh Lys Lys Ile Thr Ser
GAG ATG CTG CCC ACT GTG GTT Glu Met; Leu Pro Thr Val Val
GAC TCC CAG GGT GGT CCT GTC Asp Set Gin GIy GIy Pro Val
TTT C~ GGC CTC A~C TGC TGC Phe Leu GIy Leu Tar Cys cys
GAG GGC GTC AAC AAG AAA CTC G_!u Oly Val Asn Lys Lys Leu
AAG ACC CT0 GAG AAT CCT CAG Lys Thr Leu Glu Asn Pro Gin
CAA GCC ACC ATC ACC CAG GAY Gln Ala Thr Ile Thr Gin Asp
AAG CAA GTG TCT CAT GAC CTG Lys Gin Val Ser His Asp Leu
AA'2 TTT CTC AAG AAT TAC CTG ASh Phe Leu Lys ASh Tyr Leu
CGT GTC GGA CCC TTG GGT TTC Arg Val GIy Pro Leu Gly Phe
GCT ATT GCC TGG GTG AAG AGG Ala Ile Ala Trp Val Lys Arg
GGA GCT GCC AGT GTC TCT CTG GIy Ala Ala Set Val Sot Leu
GTG GCA CTG AGC CCC TGG GCC VOI Ala Leu Set Pro Trp Ala
ACA G~G GAT ACC GCC AAG ATG Thr Glu Asp Thr Ala Lys Het
TTG AAA AGC CAG GAG TAC CCC Leu Lys SeE Gln Glu Tyr Pro
CCC ATC AAC CTG TAC GAC AAC Pro Ilc Ash Leu Tyr Asp Asn
ACA GTT GAC GTG CCC GCC ATC Thz Val Asp Val Pro Ala Ile
ACT GTC GCC AAG GGG CTT AAA Thr Val Ala Lys Gly Lou Lys
GAG AAC ATG AAG AAG ACA GTG Glu Ash Met Lys Lys Thr Val
CAC AGA GCC CAT GCC AAG AGT His Arg Ala His Ala Lys Ser
TGG ATG GGG GCA GAC CAC GCT Trp Met Gly Ala Asp His Ala
CAA GAC AGG ACT GTC TCC AAG Gln Asp Arg Thr Val Set Lys
TCA CC ° GTG CCC ACA CAC TGG Set Pro Val Pro Thr His Trp
ACC mCC ATG AAG GAG CAC CTA ThE Se: Met LyS GLI His Leu
GGT GAC CAC ACT CCC CCT GAG Gly Asp His ThE Pro Pro GIu
CCA CCT ACA GAT GAC TCT CAG Pro Pro Thr Asp Asp Set Gin
231
TTG GCA GCT GCT TGT GCT GCA 80 Leu Ala Ala Ala Cys Ala Ala 1
AGT CTC TTG GGT GGT GAC TCT 158 SeE Leu Leu GIy GIy Asp Her 27
CGT CAC CCT GGC TGG CAAGGG 236 Arg His Pro Oly Trp Gln Gky 53
GAg ACC TAT GGG CAA OAA GAC 314 Asp Thr TyE GIy Gln Olu Asp 79
CCTGHG ATG GTC TGG ATC TAT 392 Pro Val Net Val Trp Ile Tyr 105
TAT GAT GGG GAA GAG ATC GCC 470 Tyr Asp Gly Glu Olu Ile Ala 131
CTT AGC ACC GGA GAT GCT AAC 548 Leu Ser Thr Gly Asp Ala Ash 157
AAC ATT GCA GCC TTT GGA G GA 626 Ash Ile AI~ Ala Phe GIy GI7 183
CAG ACC CTC TCC CCA TAC AAC 704 Gin Thr Leu Set Pro Tyr Ash 209
ATC CAG GAG AAT CCA CTT TTC 782 zle Gin Glu Ash Pro Leu Phe 235
GCT GGG TGT CTG AAG ATC ACA 860 Ala Gly Cys I,eu Lys Ile Thr 261
ATT GTG CAC TAC CTG GCC TTC 938 lle Val H£s Tyr Leu Ala Phe 287
GCT GCT GAC ATT GAC TAC TTA 1016 A1a Ala Asp Ile AspTyr Leu 313
GAC AAG GCC AAG CAG GAT GTC 1094 Asp Lys A1a Lys Gln Asp Val 339
GGC ACC CAA GCC ACC TTT GAC 1172 GI7 Thr Gln Ala Thr Phe Asp 365
GTG GCC TTT GAG ACT GAC ATA 1250 Val Ala Phe Glu Thr Asp Zle 391
GCC AAG ACC TAC TCT TAC CTG 1328 A1a Lys Thr Tyr Set Tyr Leu 417
GAT GAC CTC CAG TAC GTC TTT 1406 Asp Asp Leu Gln ~yr Val Phe 443
GCC ATG ATT GC~ TAC TGG ACC 1484 Ala Met Ile Ala Tyr Trp Thr 469
TAC CCT TAT ACC ACG GA~ AAT 1562 Tyr Pro TMr Thr Thr Glu ASh 495
AGG GAA AAG TTC CTC AAG TTC 1640 Arg Glu Lys Phe Leu Lys Phe 521
GAT GAC TCA GAG GCT GCC CCC 1718 Asp Asp Set Olu Ala Ala Pro 547
ACA ACA CCG GTG CCC CCA ACA 1796 Thr Thr Pro Val Pro Pro Tbr 573
1797 GAC AAC TCT CAG GCT GGT GAC TCT GTG GAG GCT CAG ATG CCT GGT CCC ATT GGC TTC TAA AGTCCTATAAACCGGGGCTAGAG 1879 574 Asp ASh Set Gin Ala GIy Asp Set Val Glu Ala Gin Met Pro GIy Pro Zle GIy Phe * 592
1880 ATG~cTCA~GAGCTAAGAGCTCTTCTTCCACTGTTCTTCTGAAGGTCCTGTGTTcAATTC~CAG~ACcCACATTGCTGcTTACAGcTGT~TGATTCCGT~ACT 1983
1984 GGTGTGC AGATGTAC ACGuAGAC AAAGCGCCTATATATAGAAAATTCATAC ATAAATAAAGTCTTATAA.
Fig. 2. The cDNA nucleotide sequence and deduced amino acid sequence of the open reading frame for rat pancreatic cholesterol esterase. The numbering system begins with nucleotide 1 of the full-length cDNA. The N-terminus amino acid in the mature protein ;~ labeled as residue | in the amino acid sequence. Peptide sequences identical to those obtained from protein sequencing techniques are underlined. The polyadenylation signa| is highlighted by a fine above the nucleotide sequence. The * below amino acid residue 189 denotes the last amino acid in the possible
N-glucosylation signal of the protein.
cDNA, isolated from A clone 5, was subcloned into the plasmid vector pSVL at the multiple cloning site. Orien- tation of the cDNA insert in the plasmid was de- termined by restriction mapping techniques. The vector
containing the cholesterol esterase cDNA in the proper orientation was then introduced into COS cells using the calcium phosphate precipitation method. Expression of the cDNA was driven by the SV40 promoter located
232
upstream from the 5' end of the cDNA. Cell extracts were isolated 48 h after transfection and cholesterol esterase activity was determined. Results, as shown in Fig. 3, indicated that cells transfected with the cholesterol estera~e cDNA expressed high levels of bile salt-stimulated cholesterol esterase activity. In contrast, control COS cells without plasmid transfection did not show any ~;etectable bile salt-stimulated cholesterol esterase activity in our assay system. Furthermore, cells transfected wi& the pSVL plasmid were also negative for bile salt-stimulated cholesterol esterase (Fig. 3).
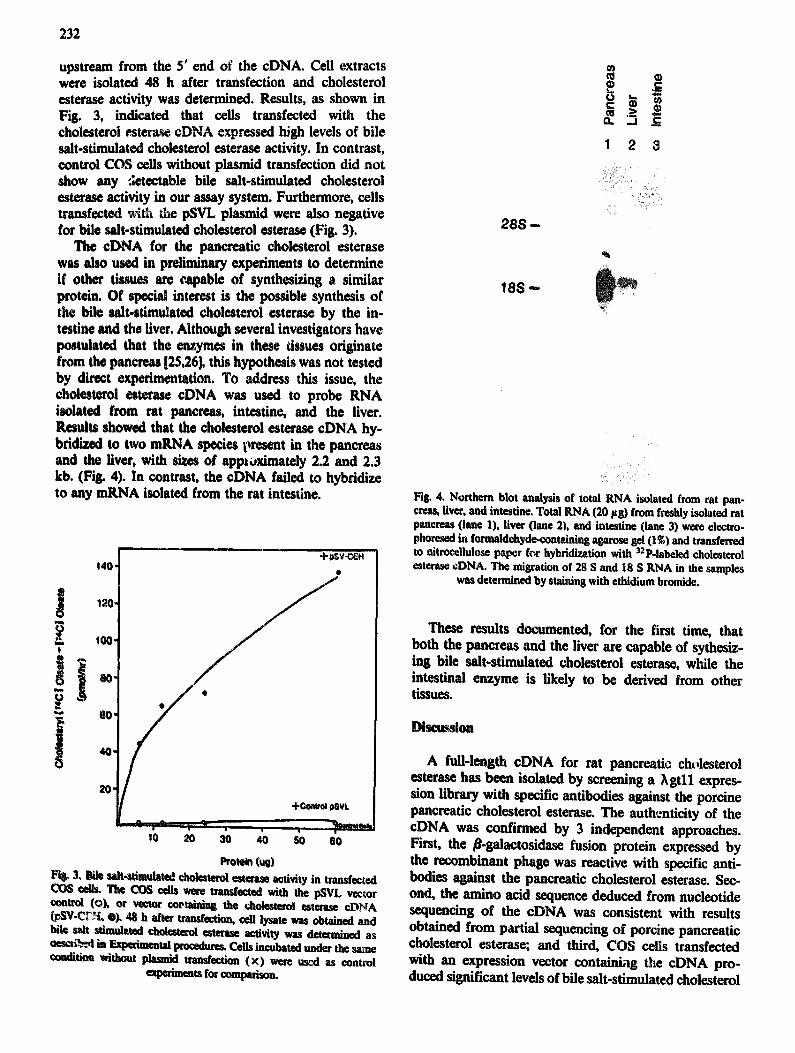
The cDNA for the panereatic cholesterol esterase was also used in preliminary experiments to determine if other tissues are capable of synthesizing a similar protein. Of special interest is the possible synthesis of the bile =fit-stimulated cholesterol esterase by the in- testine and the liver. Although several investigators have postulated that the enzymes in these tissues originate from the pancreas [25,26], this hypothesis was not tested by direct experimentation. To address this issue, the cholesterol esterase eDNA was used to probe RNA isolated from rat pancreas, intestine, and the liver. Results showed that the cholesterol esterase eDNA hy- bridized to two mRNA species present in the pancreas and the liver, with sizes of approximately 2.2 and 2.3 kb. (Fig. 4). In contrast, the eDNA failed to hybridize to any mRNA isolated from the rat intestine.
140" *
~ 100- t
"~ CO,
, o . s o e o
Protein ( t~ Fit~ 3, Bile ult-,~timalated cholesterol esterase activity in transfeeted
~ The COS cells were tran~ected with the pSVL vector ¢oatmi (ok of vector ¢oetaiDJng the dmlesteml esterase eDNA ¢L~'V-C~'£ o). 48 It after tmnsfection, cell iysate was obtained and bile salt =tiudated cholesterol esterase activity was determined as ¢tes=i-%,~,4 in Expeimental ~ooedutes. Cells incubated under the same ¢=adition without plasmid tran~eetion (×) were used as control
eqperimem for mmpvho~
288 - -
18S -
o " $ -
1 2 3
i!i ¸i ilil i i
~i ~̧ ~i~! i
Fig. 4. Northern blot analysis of total RNA isolated from rat pan- ¢reas, fiver, and intestine. Total RNA (20 Fg) from freshly isolated rat pancreas (lane 1), liver (lane 2), and intestine (lane 3) were electro. phoresed in formaldehyde-containing agamse gel (I~) and transferred to nitrocellulose paper for hybrld!vation with SaP-labeled cholesterol esterase eDNA. The misration of 28 S and 18 S RNA in the samples
was determined by staining with ethidium bromide.
These results documented, for the first time, that both the pancreas and the liver are capable of sythesiz- ing bile salt-stimulated cholesterol esterase, while the intestinal enzyme is likely to be derived from other tissues.
Discussion
A full-length cDNA for rat pancreatic cholesterol esterase has been isolated by screening a ?~gtll expres- sion library with specific antibodies against the porcine pancreatic cholesterol esterase. The authenticity of the cDNA was confirmed by 3 independent approaches. First, the fl-galactosidase fusion protein expressed by the recombinant phage was reactive with specific anti- bodies against the pancreatic cholesterol esterase. Sec- ond, the amino acid sequence deduced from nucleotide sequencing of the eDNA was consistent with results obtained from partial sequencing of porcine pancreatic cholesterol esterase; and third, COS cells transfected with an expression vector containi,~g the eDNA pro- duced significant levels of bile salt-stimulated cholesterol

TABLE 1I
Sequence differences belween cholesterol esterose and lysophosphofipase
Cholesterol esterase
Nucleotides
66°GCT GGA GCT GCC AGT GT'C TCT 6e°
Amino acids
19SAla Gly ALa Ala Sot Vat Set a°°
233
Lysophospholipase
Cholezterol esterase
Lysophospholipase
Cholesterol esterase
Lysophospholipase
GCT 6GA 66T 6CC ATT 6TC TCT
lS4aTAT A¢C ACG GAG AAT 156a .
TAT ACC ATG GAG AAT
19SaTTGCTGCTTAC 1¢62
TTGCTTCTTAC
Ata GLy G~y ALa gLe VaL Set
t~9'DTyP Thr Thr 6Lu ASh 495
Tyr Thr Net 6 tu Asn
Untranslated
Untranslated
esterase activity, similar to that of the pancreatic en- zyme. Based on these observations, we concluded that the isolated eDNA represents the complementary DNA of rat pancreatic cholesterol esterase.
A computer search of the GenBank and the EMBL databases revealed that the sequence of the isolated cholesterol estetase eDNA was highly homologous to the sequence of a putative lysophospholipase eDNA from rat pancreas [11]. Differences were observed in only four residues resulting in a conserved change of three amino acid residues (Table II). Ala-197 in the cholesterol esterase sequence was reported to be giycine in the lysophospholipase sequence. Likewise, Ser-199 and Thr-493 in cholesterol esterase was substituted by Ile and Met, ;espectively, in lysophospholipase. The other nueleotide difference between the two eDNA sequences was observed at nucleotide 1957, in which a G was reported here for the cholesterol esterase eDNA instead of the T in the lysophospholipase sequence [11]. This nueleotide substitution occurred at the Y-untrans- lated region of the mRNA, which would not affect the translated sequence of the proteins. Since these dif- ferences were most likely due to polymorphisms ob- served between the different cDNA libraries, or to possible sequencing errors, it is probable that the eDNA isolated in these studies were derived from identical mRNA. Our results reported in this communication used a combination of antibody reactivity, protein se- quencing, and expression of enzyme activities to con- firm the identity of the cholesterol esterase eDNA. Likewise, in the study by Hun et al. [11], transfection of the eDNA into Chinese Hamster Ovary cells also re- sulted in the expression of !ysophospholipase activity. Therefore, it is probable that the same protein can hydrolyze both cholesteryl esters and lysophospholipids. Previous results using enzymological approaches have shown similarities between the pancreatic cholesterol esterase and lysophospholipase [1,7]. Our results re- ported herein documented unequivocally that a single protein is responsibe for both enzyme activities.
The search of databases for sequence similarities also revealed that the cholesterol esterase structure is similar to that of other serine esterases. In particularly, a 62-
amino-acid sequence, residues 159-221, in the primary structure of cholesterol esterase showed a high degree of similarity to the active site regions of acetylcholinester- ase [27] and cholinesterase [28]. Both these enzymes are serine esterases, with the active site located at residue 200 and 198, respectively. The active site serine i~ centered within the sequence T-I/L-F-G-E-S-A-G- G/A-A. Interestingly, this consensus sequence was highly conserved in the structure of cholesterol esterase (Fig. 5). Since cholesterol esterase is also a serine esterase enzyme, this similarity suggests that the active site of cholesterol esterase may reside in this domain. The serine residue at position 194 may be the important serine in the catalytic hydrolysis of cholesteryl esters by this protein. It is noteworthy that this 62-amino-acid sequence within the structure of cholesterol esterase is also homologous to the liver microsomal carboxyl- esterase residues 168-230 [29]. The microsomal enzyme is another serine esterase with several non-specific lipo- lytic enzyme activities including the hydrolysis of cholesteryl esters. The similarities in the sequences of these two proteins further suggest that the active site of cholesterol esterase is located at this domain. Further- more, the similarities of active site sequences between the serine esterases suggest that these proteins may be derived from a common ancestral gene. Whether the proteins are derived from differential splicing of the same gene, or more likely products of related genes due to oxen shuffling [30] of common ancestral genes, re- mains to be determined.
Comparison of the structure of cholesterol esterase with the recently sequenced hormone-sensitive lipase of rat adipocytes [31] revealed no similarities between the two proteins. It is interesting to note that both cholesterol esterase (bile salt-stimulated lipase) and hormone-sensitive lipas¢ are active in the hydrolysis of cholesteryl esters. The bile salt-stimulated lipase cata- lyzes cholesteryl ester hydrolysis in the pancreas, the intestine, and the liver for cholesterol absorption and recirculation, while the hormone sensitive lipase cata- lyzes cholesteryl ester hydrolysis in steroidogenic tis- sues. However, comparison of their sequences using the rigorous ALIGN program and other similarity searches

234
failed to identify any common domains between the two proteins. The only similarity between the proteins is located at or near the active site serine, where cholesterol esterase has the sequence G-E-S-A-G and the sequence of hormone-sensitive lipase is G-D-S-A-G. It seems probable that this five-amino-acid sequence is sufficient to confer cholesteryl ester hydrolysis. However, this sequence also fits the consensus sequence for the lipid interracial binding sites of other ripases such as the pancreatic lipase [32], cholesterol acyltransferase [33], lingual lipase [34], hepatic triglyceride lipase [35,36] and lipoprotein lipase [37]. These ripases lack cholesterol esterase activity, therefore suggesting that this sequence may be necessary, but not sufficient, to account for the enzyme activity in cholesterol esterase. It is possible that this sequence is responsible for the absorption of the protein into lipid interfaces as suggested by other investigators [36,37]. On the other hand, using sequence comparison approaches, we have identified the same serine residue as possible active site attd as the lipid- binding domain. Although previous data have shown that the active site of cholesterol esterase is distinct from its lipid interracial binding site [1], these observa- tions are not necessarily in contradiction because the active site of the enzyme may simply He in close proxim- ity to the ripid-binding site and may be located elsewhere within this 63.amino-acid serine esterase homologous domain. The availability of the eDNA for cholesterol esterase will allow a direct determination of active site and lipid-binding domain of the protein in further experiments.
In addition to the identification of the possible active site and lipid-binding domain of the cholesterol esterase, analysis of the cholesterol esterase sequence also re- vealed several regions with a high concentration of Charged amino acids. Of particular interest is the do-
main at amino acid residues 61-66. The sequence in this domain, K-K-R-C-L-Q, fits the cilarge distribution of the consensus heparin-binding domain as identified by Cardin and his associates [36,38]. This finding suggests that cholesterol esterase may also be a protein capable of interacting with heparin-like molecules. A recent report by Bosner et al. [39] has suggested that the pancreatic cholesterol esterase may bind to heparin-like structures on the surface of intestinal cells where it catalyzes the hydrolysis and absorption of alimentary cholesteryl esters. Therefore, it is possible to postulate that cholesterol esterase binding to the intestinal cells are mediated by residues 61-66 in the protein.
The sequence of the cholesterol esterase also showed four repetitive sequences toward the C-terminal end of the protein which are enriched with Pro, Asp, Glu, Ser, and Thr residues (Table II1). According to the PEST hypothesis championed by Roger's, Wells, and Rech- steiner, regions enriched in these residues may serve as signals for selective proteolysis of the protein [40]. In fact, a significant positive score was obtained for cholesterol esterase when the sequence was analyzed using the PEST search program supplied by Dr. Rech- steiner (University of Utah). The PEST score of the cholesterol esterase compares favorably with those re- corded for other short-rived proteins (Table III). There- fore, the presence of these structural domains in cholesterol esterase suggests that the enzyme may have a short half-life If this hypothesis proves to be correct by direct experimentation, then small changes in the synthetic rate of cholesterol esterase will be important in regulation of cholesterol absorption. Experimer~ts designed to test these hypotheses are currently in pro- gress.
Results reported in this manuscript also showed two distinct mRNAs for the bile salt-stimulated cholesterol
TABLI~ lit Analysis of PEgT.r~eh reglo~ in ¢~olesterol esterase and other short./load protebts
Sequenc~ and PEST scores of representative short.lived proteins were extracted from data in Ref. 4(/. A complete list of the proteins could be found in the same reference. PEST score for cholesterol esterase was derived by computer analysi~ using the program supplied by Dr. M. R(~lnteiaer.
Protein Sequen~ PEST scor~ Half-life (h)
Cholesterol esterase 538-~14 P P E O O S E 547-$$$ P V P P T 0 D S (I 15.5 $~)8-M)6 P V P P T 0 G S q 56~677 P V P P T O f l $ Q
&denovim~ early protein EtA 126-141 E A G F P P S 0 D E D E E S E E 11.6 186-201 S P V S E P E P E P E P E P E P 11.8
Murine omithine decarboxylase 426--444 P P E V E E Q 0 0 G T L P H $ C A Q E 5.2
HMG-CoA teducta~e 3gl-395 K L S S V E E E P G V S q I) R 4.2 429-442 R T q E L E I E L P S E P R 5.5
0.5
0.5
L2-3.0

g c e e l t l c h o l i n e s t e r a s e ( 1 6 5 - 2 2 7 ) : ~ V ~ R [~ A L 0 W V ~
e h o l i n e s t e r a s e { 1 6 3 - 2 2 5 ) : L~G NIMLG ~ D QI{~ L[A L QW V OIK 1,I I A A F G G]N]P K S V !
,icrosomal esterase(168-230):~,[y~.ILt~A~W V ~ I ~ I I. A ~ F G G ~ P D S ~
235
C h o l e s t e r o l e s t e r a s e I F e lg S A G A A S V -~ r.]O Tit. S PlY NI.~G L~I~R R A I S O S ~
Fig. 5. Similarities between the amino acid sequences of torpedo acetyleholinesterase, human cholinesterase, rat liver microsomal esterase and rat Fancreatic cholesterol esterase. The numbers in parentheses signify the position of the sequences within each protein.
esterase in pancreas and in liver. Although it is very likely that the two mRNAs differ in size due to differen- tial transcription initiation or to alternative termination sites, it is possible that the two mRNAs encode for two different forms of the protein. Heterogeneity of cholesterol esterase have been noted for the porcine pancreas previously [15]. These proteins have identical amino-terminal sequences but differ in molecular weight [15]. If two forms of the cholesterol esterase also exist in the rat, the two mRNA species may represent different products due to differential gone splicing. These possi- bilities remain to be determined in future studies.
The observation that cholesterol esterase mRNA was present in rat pancreas, but absent from, or in low abundance in the intestine, is consistent with physio- logical studies showing that the intestinal cholesterol esterase was derived from the pancreas [25]. The identi- fication of the bile salt-stimulated cholesterol esterase mRNA in the liver was a surprising result. The liver possesses two disfin¢:t cholesterol esterases in addition to the acid lipase in the lysosomes [41]. One cholesterol esterase is present in the hep,~tic microsomes. This protein appears to be identical to other non-specific microsomal esterases. The eDNA has recently been isolated and the sequence determined [30]. The ho- mology between the sequence of the microsomal carboxylesterase and the pancreatic cholesterol esterase is shown in Fig. 5. The cholesterol esterase in the cytosol of the fiver is apparently the same enzyme as the bile salt-stimulated cholesterol esterase of the pancreas. The hepatic protein has recently been purifi,ed to ho- mogeneity [12]. The N-terminal sequence of this protein is identical to the pancreatic cholesterol esterase. Al- though the enzyme was detected in the liver [26], the cholesterol esterase had been presumed to be derived from the pancreas via the enterohepatic circulation sys- tem [26]. This report documents that the bi|e salt- stimulted cholesterol esterase in fiver cytosol is derived directly from de novo synthesis by the liver.
Acknowledgements
This research was supported in part by a grant from the Lapp Endowment Fund, a Grant-in-Aid (SW-87-21) from tiae Ohio Affiliate of the American Heart Associa- tion, the Biomedical Research Support Grant Program (S07-RR05408-27), and grant HL17371 (to H.L.B.) from the National Institutes of Health. D.Y.H. is an Estab- lished Investigator of the American Heart Association.
References
1 ~udd, E.A. and Brockman, H.L. (1984) in Lipases (Borgstrom, B. and Brockman, H.L., eds.), pp. 185-204, Elsevier. Amsterdam.
2 Gallo, L.L., Newbill, T. Hyun, J.. Vahouny, G.V. and Treadwell. C.R. (1977) Proc. Sot:. Exp. Biol. Med. 156, 277 281.
3 Borja, C.R., Vahouny, G.V. and Treadweil, C.R. (1964) Am. J. Physiol. 206, 223-228.
4 Chaikoff, I.L. and Kaplan, A. (1935) J. Biol. Chem. 112, 155-165. 5 Gallo. L.L., Cheriathundam, E., Vahouny, G.V. and Treadwell,
C.R. (1978) Arch. Biochem. Biophys. 191, 42-48. 6 Van den Bosch, H., Aarsman, A.J, and Van Deenen, LL.M.
(1974) Biochim. Biophys. Acta 348, 197-209. 7 Lombardo, D., Fauvel, J. and Guy, O. (1980) Biochim. Biophys.
Acta 611,136-146. 8 Albro, P.W., Corbett, B.J. and Latimer, A.D. (1976) Biochim.
Biophys. Acta 424, 351-365. 9 Mathur, S.N., Joshi, P.S., Murthy, S.K. and Ganguly, J~ (1974)
Indian J. Biochem. Biophys. 11, 105-109. 10 Aarsman, A.J., Hi,e, J.D.R. and Van den Bosch, H. (1977)
Bioclum. Biophys. Acta 489, 242-246. 11 Ham J.H., Stratowa, C. and Rutter, W.J. (1987) Biochemistry 26.
1617-1625. 12 CamuUi, E.D., Linke, M.J., Brockman, H.L. and Hui, D.Y. (1989)
Biochim. Biophys. Acta 1005, 177-182. 13 Maniatis, T., Fritsch, E.F. and Sambrook, J. (1982) Molecular
Cloning. A Laboratory Manual, Cold Spring Harbor Laboratory. Cold Spring Harbor, NY.
14 Sanger. F., Nicklen, S. and Coulson. A.R. (1977) Proc, Natl. Acad. Sci. USA 74, 5463-5467.
15 Rudd, E A., Mizuno, N.K. and Brockman. H.L. (1987) Biochim. Biophys. Acta 918. 106-114.

236
16 Deisiqlei, U., Weber, W., Havinga, J.R., lhrke, G., Hui, D.Y., Wernette-Hammond, M.E., Turck, C.W., Innerarity, T.L. and Mahley, R.W. (1988) Arteriosclerosis 8, 288-297.
17 Lathe, R. (1985) J. Mol. Biol. 183, 1-12. 18 Graham, F.L. and Van der Eb, A.L (1973) Virology 52, 456-467. 19 Blanef, W.S., Prystowsky, J.H., Smith, J.E. and Goodman, D.S.
(1984) Biochim. Biophys. Acta 794, 419-427. 20 Lowry, O.H., Rosebroush, N.J., Farr, A.L. and Randall, ILL
(1931) J. Biol. Chem. 193, 265-275. 21 Young, IL.A. and Davis, ILW. (1983) Proc. Natl. Acad. Sci. USA
80, 1194..1198. 22 Gallo, L.L. (1981) Methods Enzymol. 71, 664-674. 23 Abouakil, N., l~wflska, E., Bonicel, J. and Lombardo, D. (1988)
Biochim. Biophy~. Acta 961, 299-308. 24 Kozak, M. (1986) Nucleic Acids Res. 12, 857-872. 25 Lechene de la Porte, I% Abouakil, N., Lafont, H, and Lombardo,
D, (1987) Biochim, Biophys, Acta 920, 237-246, 26 Ha~aen, E,H, (1988) Biochim. Biophys. Acta 963, 28-34. 27 Schurnachet, M., Camp, $,, 2~/aulet, Y., Newton, M., MacPhee-
QuttlJey, K,, Taylor, $.S., Friednumn, T. and Taylor, P. (1986) Nature 319, 407-409.
28 Lockfldge, O,, P~rtels, C,F,, Vaughan, T.A~, Won8, C.K., Norton, S,E, and Johnson, L.L, (1987) J, Biol, Chem. 262, 549-557.
29 long, R.M., Satoh, H,, Martin, B.M., Kimura, S., Gonzalez, F.L and Pohl, L.R. (1988) Bivchem. Biophys. Res. Commun. 156, 866-873.
30 Sudhof, T,C., Goldstein, J.L., Brown, M.S. and Russell, D.W. (1985) Science 228, 818-822.
31 Holm, C., Kirchgessner, T.G., Svenson, K.L., Fredrikson, G., Nilsson, S., Miller, C.G., Shively, J.E., Heinzmann, C., Sparkes, R.S., Mohandas, T., Lusis, A.J., Beifrage, P. and Scl~otz, M.C. (1988) Science 241, 1503-1506.
32 De Caro, L, Boudouard, M., Bonicel, J., Guidoni, A., Desnuelle, P. and Rovery, M. (1981) Biochim. Biophys. Acta 671, 129-138.
33 McLean, J., Fielding, CJ., Dr,~yna, D., Dieplinger, H., Baer, B., Kohr, W., Henzel, W. and Lawn, R. (1986) Proc. Natl. Acad. Sci. USA 83, 2335-2339.
34 Docheny, A.LP., Bodmer, M.W., Angal, S., Verger, R., Riviere, C., Lowe, P.A., Lyons, A., Emtage, J.S. and Harris, T.J.R. (1985) Nucleic Acit~s Res. 13, 1891-1903.
35 Komaromy, M.C. and Schotz, M.C. (1987) Proc. Natl. Acad. Sci. USA 84, 1526-1530.
36 Martin, G.A., Busch, S.J., Meredith, G.D., Cardin, A.D., Blanken- ship, D.T., Mao, SJ.T., Rechtin, A.E., Woods, C.W., Racke, M,M., Schafer, M.P., Fit~erald, M.C., Burke, D.M., Flanagan, M.A. and Jackson, R.L. (1988) J. Biol. Chem. 263, 10907-10914.
37 Kirchgessner, T.G., Lusis, A.L, Svenson, K.L. and Schotz, M.C. (1987) J. Biol. Chem. 262, 8463-8466.
38 Cardin, A.D. and W¢instraeb, HJ.R. (1989) Arteriosclerosis 9, 21-32.
39 Oosner, M.S., Gulick, T., Riley, DJ.S., Spilburg, C.A. and Lange, L.G., IlL (1988) Prec, Natl. Aced. SCI. USA 85, 7438-7442.
40 Rogers, S., Wells, R, and Rechsteiner, M. (1986) Science 234, 364-368.
41 Deykin, D. and Goodman, D.S. (1962) J. Biol. Chem. 237, 3649-3656.