molecular bacteria-fungi interactions: effects on
TRANSCRIPT

MI67CH18-Hertweck ARI 12 August 2013 11:12
Molecular Bacteria-FungiInteractions: Effects onEnvironment, Food,and MedicineKirstin Scherlach, Katharina Graupner,and Christian HertweckLeibniz Institute for Natural Product Research and Infection Biology, Hans Knoll Institute,07745 Jena, Germany; email: [email protected]
Annu. Rev. Microbiol. 2013. 67:375–97
First published online as a Review in Advance onJune 26, 2013
The Annual Review of Microbiology is online atmicro.annualreviews.org
This article’s doi:10.1146/annurev-micro-092412-155702
Copyright c© 2013 by Annual Reviews.All rights reserved
Keywords
endofungal bacteria, endophytes, mushroom diseases, quorum sensing,secondary metabolites, symbiosis
Abstract
This review focuses on bacteria-fungi interactions mediated by secondarymetabolites that occur in the environment and have implications formedicine and biotechnology. Bipartite interactions that affect agricultureas well as relationships involving additional partners (plants and animals) arediscussed. The advantages of microbial interplay for food production andthe risks regarding food safety are presented. Furthermore, recent devel-opments in decoding the impact of bacteria-fungi interactions on infectionprocesses and their implications for human health are highlighted. In addi-tion, this reviews aims to demonstrate how the understanding of complexmicrobial interactions found in nature can be exploited for the discovery ofnew therapeutics.
375
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
Secondarymetabolites: organiccompounds that arenot essential for thegrowth of an organismbut are often endowedwith potent biologicalactivities to warrantsurvival of the species
Contents
INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 376BACTERIA-FUNGI INTERACTIONS IN THE ENVIRONMENT . . . . . . . . . . . . . . 376
Bipartite Bacteria-Fungi Interactions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 376Tripartite Bacteria-Fungi-Plant Interactions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 379Tripartite Bacteria-Fungi-Animal Interactions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 382
BACTERIA-FUNGI INTERACTIONS: EFFECTS ON FOODPRODUCTION AND SAFETY . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 384
BACTERIA-FUNGI INTERACTIONS: EFFECTS ON MEDICINE . . . . . . . . . . . . . 386Bacteria and Fungi as Coacting Human Pathogens . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 386Exploiting Bacteria-Fungi Interactions for Drug Discovery . . . . . . . . . . . . . . . . . . . . . . . 388
CONCLUSIONS AND PERSPECTIVES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 389
INTRODUCTION
The discovery of penicillin is one of the first documented observations of an interaction betweenbacteria and fungi mediated by small molecules. Historically molecular intergeneric interactionswere regarded mainly as growth-inhibiting interactions; however, modern research revealed thatmicrobial cross talk forms an integral part of our environment and covers various aspects beyondsimple antibiosis. There are instances in which natural products alter phenotypes and developmen-tal processes, such as sporulation or biofilm formation, and serve as virulence factors in symbioticand pathogenic associations involving additional partners. Specialized mutualistic relationshipshave evolved in which a host organism harbors a symbiont to make use of its chemical synthe-sis capabilities to combat competitors or to maintain a certain lifestyle. The growing number ofstudies published in the past few years that report discoveries in this field points to an excitingemerging area of research. Our increasing understanding of the complex networks in microbialecology will not only help us understand fundamental biological processes but also lead to the dis-covery of new virulence factors and drug candidates. This review highlights recent contributionsto the understanding of bacteria-fungi interactions mediated by secondary metabolites that occurin the environment and affect medicine and biotechnology.
BACTERIA-FUNGI INTERACTIONS IN THE ENVIRONMENT
Bipartite Bacteria-Fungi Interactions
Microbes ubiquitously occur in the environment and colonize almost every ecological niche. Be-cause of this high abundance, different species coinhabit certain habitats and as a consequenceinteract with each other. Such encounters probably represent the driving force to produce sec-ondary metabolites that regulate the coexistence and survival of different species.
Interactions via antibiosis. Production of antimicrobial compounds provides a growth advan-tage for an organism that enables its survival in a competitive environment (112). A huge numberof antibiotic and antifungal natural products have been identified since the beginning of thegolden age of antibiotics, and many of these compounds have been applied in medicine (11).Besides the medicinal importance, these works provided valuable insights into the ecological
376 Scherlach · Graupner · Hertweck
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
Imaging massspectrometry:analytical techniquethat combines theuniversal detectioncapability of massspectrometry withmicroscopic imagingtechniques and thusallows the visualizationof biomolecules inbiological matrices
function of the compounds and demonstrated how intricate and finely tuned bacteria-fungi in-teractions in nature are. This is reflected by the different strategies that bacteria and fungi haveevolved to combat secreted antimicrobial compounds. For example, Schoonbeek et al. (145) re-ported that broad-spectrum antibiotics produced by Pseudomonas spp. (2,4-diacetylphloroglucinol,phenazine-1-carboxylic acid, and phenazine-1-carboxamide) induce the expression of several ABCtransporter genes in the plant pathogenic fungus Botrytis cinerea, thus ensuring the efflux of toxicmetabolites. Fusarium spp. produce the mycotoxin fusaric acid, which is able to repress the pro-duction of 2,4-diacetylphloroglucinol in Pseudomonas fluorescens CHA0 by reducing the expressionof the responsible biosynthetic genes (110). In return, the biocontrol strain P. fluorescens WCS365profits from the mycotoxin, as it appears to be a chemoattractant for this species (42). Theseworks also denote that antibiotics not only are chemical warfare agents, but presumably havenumerous additional functions in the microbial interplay. For instance, at subinhibitory con-centrations, antibiotics may stimulate or depress bacterial gene expression at the transcriptionlevel and thus affect virulence and metabolic and adaptive functions (58). Thus, secreted sec-ondary metabolites might have manifold functions that we are only beginning to understand,and further studies are needed to uncover the precise roles of natural products in microbialecology.
Bacteria as inducers of mushroom diseases. Bacteria-fungi interactions adapt to changingenvironmental conditions. Within one genus of bacteria, organisms can be found that exert ei-ther beneficial or detrimental effects on a fungal host. Well-studied examples are the interactionsbetween various Pseudomonas species and the cultivated mushrooms Agaricus bisporus and Pleu-rotus ostreatus (3). Whereas Pseudomonas putida stimulates the fructification of A. bisporus (109,129), other pseudomonads cause mushroom diseases (94, 136). Brown blotch disease, initiated byP. tolaasii and other Pseudomonas spp., accounts for significant crop losses in mushroom farming.Typical symptoms include a dark brown discoloration of the mushroom caps accompanied bycharacteristic lesions on the basidiocarp (152). The causal agents are tensioactive lipopeptidesnamed tolaasins that are produced by the bacteria (30). Tolaasins are cyclic lipodepsipeptidesconsisting of 18 amino acids with β-hydroxy-octanoic acid at the N terminus (9). Tolaasin I,which is considered the main virulence factor, exhibits antimicrobial activity and is highly resis-tant to enzymatic degradation (128). The characteristic symptoms of the disease are due to theability of tolaasin to disrupt the fungal cell membrane by forming transmembrane pores (20, 33).Other known bacterial mushroom pathogens include Pseudomonas gingeri (ginger blotch disease),P. costantinii, P. aeruginosa, and P. fluorescens (56, 106). A related bacterium, Pseudomonas ‘reactans,’has been proposed as a biocontrol strain to reduce blotch diseases, even though it is classified asa mushroom pathogen (151). P. ‘reactans’ produces in culture an extracellular peptide called thewhite line inducing principle (WLIP), which is able to precipitate tolaasins (98). Pretreatment ofA. bisporus with the isolated peptide protects the mushroom from brown blotch symptoms causedby infection with P. tolaasii (3). WLIP consists of nine amino acids and an N-terminal fatty acid,representing a potent biosurfactant with antimicrobial properties (98). The molecular basis forthe biosynthesis of this peptide in P. putida was recently established (133).
Graupner et al. (60) described the first insights into the pathogenicity factors of Janthinobac-terium agaricidamnosum, which causes soft rot disease of A. bisporus (97). By monitoring theinfection process with imaging mass spectrometry, the authors identified the lipopeptide jagaricinas a potent virulence factor. Genome mining and molecular genetic studies unequivocally estab-lished that jagaricin is biosynthesized by a nonribosomal peptide synthetase. Furthermore, potentantifungal properties of jagaricin were described, which indicates that the discovery of virulence
www.annualreviews.org • Molecular Bacteria-Fungi Interactions 377
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
Defensins: cationicpeptides produced byeukaryotes thatfunction as hostdefense molecules
γ-Butyrolactones:signaling moleculesproduced mainly bystreptomycetes thatregulate antibioticproduction andmorphologicaldifferentiation
Nonribosomalpeptide synthetases(NRPS):multifunctionalenzymes that catalyzethe biosynthesis ofpeptides
Polyketide synthase(PKS):multifunctionalenzyme that catalyzesthe oligomerization ofactivated smallcarboxylic acids;analogous to fatty acidsynthesis
factors involved in mushroom diseases may also contribute to the development of antifungaltherapeutics.
Similarly, the discovery of the first defensin from a fungus, plectasin, underlined the impor-tance of studying natural bacteria-fungi interactions. Plectasin was isolated from the saprophyticascomycete Pseudoplectania nigrella and acts by directly binding the bacterial cell wall precursorlipid II (65, 107, 143). Owing to this rare mode of action, the fungal defensin represents a promisinglead for the development of new antibacterial agents.
Bacteria-fungi interactions in lichens. Lichens are mutualistic associations between fungi andphotoautotrophic organisms, i.e., green algae and cyanobacteria (43). The symbiotic partners formcomplex and exposed structures that enable growth in diverse and even extreme environments thatusually are not favorable for the individual species (61). Secondary metabolites are hypothesizedto contribute to the ability of lichens to survive in such rather hostile environments. For ex-ample, lichens can endure high-UV exposition by synthesizing pigments that act as UV filters(19). Moreover, it was shown several decades ago that lichen extracts possess antibiotic properties(25), suggesting potential defense strategies of lichens. In many cases the mycobiont produces thesecondary metabolites (1, 38). However, more recent studies indicate that the photoautotrophicbacterial partner may also contribute biosynthetic capabilities. Yang et al. (165) reported theformation of chlorinated γ-butyrolactones called nostoclides by cultures of the cyanobacteriumNostoc sp. that were isolated from the lichen Peltigera canina. These compounds display moder-ate cytotoxic properties and may function as signalling molecules. Another well-known class ofcyanobacterial metabolites are the microcystins, which are produced by a Nostoc species recoveredfrom the lichen Pannaria pezizoides (116). These cyclic heptapeptides were isolated from multiplegenera of cyanobacteria and are strong hepatotoxins (45, 157). The biosynthesis of microcystinsby a large multifunctional enzyme complex containing both nonribosomal peptide synthetase(NRPS) and polyketide synthase (PKS) modules is well studied (44). Kaasalainen et al. (82) haveshown that microcystins are also produced in situ in the lichen and suggest that these peptidesact as deterrents for animal grazers. Cox et al. (35) analyzed Nostoc strains, among others, thatwere isolated from diverse lichens and found that the formation of a neurotoxic amino acid, β-N-methylamino-L-alanine, seems to be a common principle in cyanobacteria. Cryptophycins arehighly potent tubulin-depolymerizing polyketides isolated from cultures of the lichen cyanobac-terial symbiont Nostoc sp. ATCC 53789 (101). Because of their strong cytostatic activities, severalsemisynthetic derivatives were evaluated as potential anticancer agents (150). The finding thatlichen-derived cyanobacteria produce a number of highly toxic metabolites suggests an importantfunction of the bacterial partner for the ecological fitness of the mutualistic association.
Furthermore, the close intergeneric association between fungi and photosynthetic organismsmay be influenced by additional microbial partners (61). In many cases, lichens harbor a stableconsortium of various nonphotoautotrophic microorganisms including parasitic fungi and evenendosymbiotic organisms (61). These coinhabiting species contribute to the mutualism, for ex-ample, via secretion of secondary metabolites. Gonzalez et al. (59) isolated actinomycetes fromlichens collected in tropical and cold areas and evaluated their biosynthetic capabilities. By PCRtargeting secondary-metabolite-encoding biosynthetic genes, the authors proved the high biosyn-thetic potential of the lichen-associated bacteria. Furthermore, the prevalence of these genes wascorrelated to antimicrobial activities of the culture extracts (59). He et al. (68) isolated the bis-naphthopyrones lichenicolins A and B from a lichen-associated fungus and reported antibacterialactivity of lichenicolin A against gram-positive bacteria, suggesting an ecological function of theassociated microorganisms.
378 Scherlach · Graupner · Hertweck
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
Siderophores: ferricion-chelating agentsproduced by bacteriaand fungi to sequesteriron from theenvironment
Tripartite Bacteria-Fungi-Plant Interactions
In many cases, the microbial interplay is even more complex and involves additional partners.Both bacteria and fungi live in close association with higher organisms and may exert beneficialas well as detrimental effects on the coinhabiting species.
Bacteria-fungi interactions in the mycorrhizosphere. Fungi and bacteria are essential com-ponents of terrestrial ecosystems (17). The root system of several plant species interacts with fungito form a symbiotic alliance called mycorrhiza (17, 153). Through this mutualistic interaction,plants benefit from enhanced access to minerals, and fungi profit from the carbohydrates of rootexudates (17). Whereas such trophic interactions have been extensively studied and were the focusof a number of comprehensive reviews (5, 17, 48), only a little is known about the mechanismscontrolling these associations.
Among soil microbes, the arbuscular mycorrhizal fungi (AM fungi) represent one of the mostimportant classes, as they not only significantly influence the growth of plants but also impact thedevelopment of other microbes coinhabiting the terrestrial ecosystem (5, 17). AM fungi belongto the phylum Glomeromycota, and the establishment of a symbiotic relationship with plants isusually not host specific (103). To engage in this mutualism, both partners have to communicate,for example, via the exchange of signal molecules (67, 123). Plants secrete phytohormones suchas strigolactones to stimulate fungal metabolism and hyphal branching (2, 12). Fungi respond viaso-called Myc factors that control plant gene expression (17). In many cases this bipartite mutu-alism is extended to a third bacterial partner. Several studies indicated the presence of bacteria inthe mycorrhizosphere (40, 108). Bacteria colonize mycorrhized roots, fungal hyphae, spores, andfruiting bodies and even occur in the fungal cytoplasm. Their effect on the plant-fungus micro-habitat is dynamic and multifaceted, ranging from the production of plant hormones to alleviatingstress and controlling pathogens (5, 17). The cooperation may vary in accordance with the currentecophysiological state. Specific bacterial populations may facilitate the mycorrhizal establishment(mycorrhization helper bacteria), for example, by excretion of organic acids that serve as a carbonsource for fungi (49). To ensure a stable physical interaction with the fungi, bacteria produceextracellular polymers that aid attachment to the fungal surface (14). Soil prokaryotes may alsostimulate the growth of the fungi by triggering the germination of fungal spores (24, 70, 71).Tylka et al. (158) proposed that volatile compounds secreted by streptomycetes positively impactAM fungal germination. Later, bacteria-produced auxofurans were identified as positive effectorson fungal development (130) (Figure 1). Other important functions of such soil bacteria are thedetoxification of the fungal habitat and the protection against plant pathogens. Removal of fun-gal waste products or changing the level of siderophores may facilitate mycorrhizal growth (50).Budi et al. (24) reported that a Paenibacillus strain isolated from Glomus mosseae spores inhibits anumber of different plant fungal pathogens, thus showing a broad spectrum of activity. Bharadwajet al. (13) also studied the effect of AM fungal spore–associated bacteria on plant pathogens andevaluated the formation of siderophores. They found that a high number of bacteria inhibit thegrowth of the plant pathogen Rhizoctonia solani, although the active compounds have not beenidentified in this study. In addition, it was shown that 16 of 57 antagonistic isolates (fluorescentpseudomonads) produce siderophores (13). Because such bacterial chelators have a higher affin-ity to ferric iron than fungal siderophores, it was suggested that the production of these smallmolecules might contribute to the antagonistic activity against fungal pathogens (13). Citernesiet al. (31) studied the influence of the biocontrol compound iturin A2, secreted by Bacillus subtilisstrain M51, on AM fungi (31). The saprophytic growth of the fungus G. mosseae was inhibited byiturin A2 and no retardation in growth or establishment of symbiosis was noticed in the presence
www.annualreviews.org • Molecular Bacteria-Fungi Interactions 379
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.
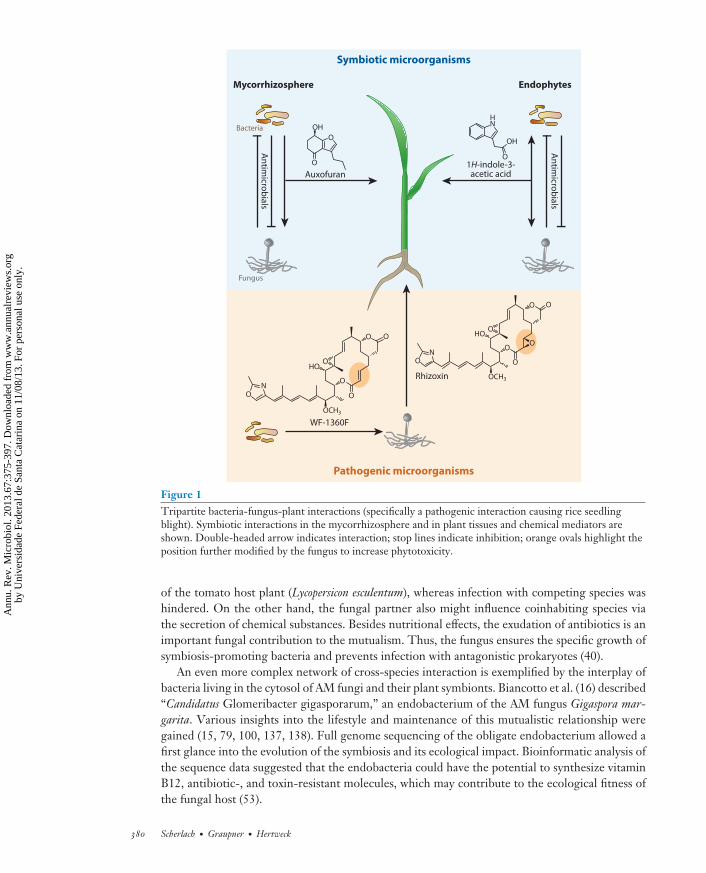
MI67CH18-Hertweck ARI 12 August 2013 11:12
O
OCH3
ON
O
HOO
O O
O
HN
O
OH
O
OH
O
O
OCH3
ON
O
HOO
O O
Bacteria
Fungus
An
timicro
bia
ls
An
timicro
bia
ls
Symbiotic microorganisms
Pathogenic microorganisms
Mycorrhizosphere Endophytes
Rhizoxin
WF-1360F
Auxofuran1H-indole-3-
acetic acid
Figure 1Tripartite bacteria-fungus-plant interactions (specifically a pathogenic interaction causing rice seedlingblight). Symbiotic interactions in the mycorrhizosphere and in plant tissues and chemical mediators areshown. Double-headed arrow indicates interaction; stop lines indicate inhibition; orange ovals highlight theposition further modified by the fungus to increase phytotoxicity.
of the tomato host plant (Lycopersicon esculentum), whereas infection with competing species washindered. On the other hand, the fungal partner also might influence coinhabiting species viathe secretion of chemical substances. Besides nutritional effects, the exudation of antibiotics is animportant fungal contribution to the mutualism. Thus, the fungus ensures the specific growth ofsymbiosis-promoting bacteria and prevents infection with antagonistic prokaryotes (40).
An even more complex network of cross-species interaction is exemplified by the interplay ofbacteria living in the cytosol of AM fungi and their plant symbionts. Biancotto et al. (16) described“Candidatus Glomeribacter gigasporarum,” an endobacterium of the AM fungus Gigaspora mar-garita. Various insights into the lifestyle and maintenance of this mutualistic relationship weregained (15, 79, 100, 137, 138). Full genome sequencing of the obligate endobacterium allowed afirst glance into the evolution of the symbiosis and its ecological impact. Bioinformatic analysis ofthe sequence data suggested that the endobacteria could have the potential to synthesize vitaminB12, antibiotic-, and toxin-resistant molecules, which may contribute to the ecological fitness ofthe fungal host (53).
380 Scherlach · Graupner · Hertweck
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
Endosymbioticbacteria: bacteria thatlive inside anotherorganism
Type III secretionsystem: a proteinstructure found inbacteria that is used totranslocate proteinsinto the cytosol ofeukaryotes (also calledinjectisome)
Bacteria as mediators of fungal virulence. A similarly intriguing tripartite relationship has beendescribed in the context of rice seedling blight. This plant disease is caused by the zygomyceteRhizopus microsporus and accounts for severe losses in agriculture in Asia (76). The fungus utilizesantimitotic toxins synthesized by its bacterial endosymbionts to destroy the rice seedlings (76, 77,120) (Figure 1). The polyketide macrolide rhizoxin binds to the β-tubulin of the rice plant cellsand causes mitotic arrest (155, 156). This leads to the characteristic symptom of the plant disease,a swelling of the seedling tips, and finally results in the death of the plant offspring. Isolationand axenic cultivation of the endosymbiotic bacteria Burkholderia rhizoxinica (119) provided clearevidence that the toxin complex is actually produced by the mutualistic bacteria (141). Toxinogenicendofungal bacteria were shown to occur in a number of Rhizopus spp. isolated from geographicallydistant regions, which points to the ecological importance of the bacteria-fungi relationship (90).
Cloning, sequencing, and functional analysis of the rhizoxin biosynthetic gene cluster provideddetailed insights into the formation of the toxin by a huge hybrid PKS/NRPS assembly line (87,88, 121). In addition to metabolic dependence, other remarkable facets of the symbiotic alliancehave been disclosed. Sequencing of the genome of the intracellular symbiont provided valuableinsights into the bacterial genetic equipment (92) and allowed deductions regarding the evolutionand metabolic interconnection of the host-symbiont relationship (93). Schmitt et al. (142) revealedhow the fungal host acquired resistance to the antimitotic toxin. By correlating rhizoxin sensitivitywith the type of β-tubulin sequences, the authors showed that rhizoxin resistance is predictableaccording to the character state of β-tubulin residue 100 (142). These findings also suggest thatendosymbiosis became possible through a shift from parasitism to mutualism in insensitive fungi.To guarantee the persistence of the mutualism during host reproduction, a finely tuned systemof fungal vegetative reproduction and transmission of endosymbionts has evolved. It was foundthat the bacteria strictly control the sporulation of Rhizopus and ensure the exclusive dispersal ofspores harboring endosymbionts (122). Moreover, the unusual mutualism has been refined throughadditional symbiosis factors such as a Type III secretion system (91) and a novel lipopolysaccharideO-antigen that sets the symbionts into stealth mode by decorating the outer membrane of theendosymbionts (96).
In a recent study, the close intergeneric interaction of the allied organisms was further disclosed.Driven by the observation that the bacteria solely produce rhizoxin derivatives harboring onlyone epoxy moiety, whereas in coculture bis-epoxides are formed, Scherlach et al. (139) studied thecontribution of the fungus to the biosynthesis of the endobacterial toxin. Through a combinationof genetic and chemical analyses it was demonstrated that the fungal host specifically tailors thebacterial metabolite to increase the phytotoxic potency (139) (Figure 1). This first example of asymbiotic synergism in the biosynthesis of a secondary metabolite that has biological significancehighlights the importance of metabolic bacteria-fungi interactions in the ecosystem.
Interactions among endophytes. Endophytes are microbes that colonize living tissues of plantswithout causing any immediate or visible negative effects (126). The impact of such symbioticassociation on the plant ecosystem is diverse and is not fully understood (4). Many endophytes havethe potential to produce bioactive secondary metabolites and are therefore regarded as a valuablesource for drug discovery (86, 126, 154). Especially, compounds with antimicrobial propertieswere frequently isolated from endophytes, suggesting these metabolites protect against harmfulmicrobes such as phytopathogens. Some endophytes were even described as producing typical plantmetabolites when cultured outside their host plant (166). In most cases a plant is colonized not bya single endophyte, but rather by a high number of different species either coinhabiting differenttissue compartments or even occurring as endobacteria in the hyphae of a fungal endophyte (72).Therefore, it is conceivable that complex patterns of microbial interactions occur within the plant
www.annualreviews.org • Molecular Bacteria-Fungi Interactions 381
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
Indoleacetic acid:a phytohormone thataffects growth anddevelopment of plants
to ensure the beneficial outcome of the mutualism. Diffusible chemical signals that coordinatethe individuals in these populations might mediate the interaction. Bandara et al. (7) undertookfirst attempts to study the interaction between endophytic fungi and bacteria in vitro showing thecolonization of fungal mycelia by bacteria and the influence of biofilm formation on antimicrobialeffects and production of indoleacetic acid–like substances (7) (Figure 1). Up to now the potentialof endophytic interspecies cross talk had hardly been exploited for the discovery of novel naturalproducts (86). Most studies focusing on the isolation of secondary metabolites from endophyteshave been carried out utilizing only axenic cultures. However, it can be assumed that a cocultureof different species usually coinhabiting a plant might lead to the formation of novel compoundsas has recently been described for cocultures of actinomycetes and ascomycetes (see below).
Tripartite Bacteria-Fungi-Animal Interactions
The lifestyle and the survival of animals in tripartite animal-bacteria-fungi interactions dependon the bacteria-fungi interplay. A remarkable example of the impact of bacteria-fungi interactionson insects is the symbiosis of the European beewolf digger wasp (Philanthus triangulum) withStreptomyces spp. (83). The female wasps carry the bacteria in their antennal glands and distributethem to the brood cell within the ground. Larvae harbor symbiotic bacteria in their cocoons thatprotect them against pathogenic microorganisms. It was shown that the streptomycete symbiontis capable of producing a mixture of antibiotics containing streptochlorin (Figure 2) and eightderivatives of piericidin under environmental conditions (85). Inhibition tests pointed toward a
Bacteria
Fungus
AnimalsMycangimycin
Streptochlorin
Violacein
Dentigerumycin
HN
NH
NH
OHO
O
O
NCl
NH
HO
O O O
HN NN
OO
HNOH
O
NHO
O
N
NH
O
HN O
OHO
OH
OO
NH
Figure 2Schematic showing different tripartite interactions among bacteria, fungi, and animals. Selected naturalproducts mediating these interplays are displayed. Double-headed arrows indicate symbiotic interaction;bolts indicate damaging effect (dashed bolts indicate decreased damage due to bacteria-fungi interaction);stop lines indicate inhibition.
382 Scherlach · Graupner · Hertweck
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
Chytridiomycosis:infectious disease ofamphibians causingtissue erosion, weightloss, and death
Ovipositional gallery:breeding galleryconstructed by beetlesunder the bark of a tree
synergistic mode of action for the produced broad-spectrum antibiotic mixture, which is activeagainst not only entomopathogenic fungi but also bacteria (85). Current research in this field re-vealed that natural-product-producing Actinobacteria, which live in association with insects, mightbe a common theme in nature (e.g., 127). Poulsen et al. (127) cultivated Streptomyces isolates fromsolitary mud dauber wasps and demonstrated that several isolates produce antifungal and antibac-terial secondary metabolites. The screening even led to the discovery of the novel macrocycliclactam sceliphrolactam. However, evidence for a protective role of these newly discovered naturalproducts remains to be proven (127).
With respect to the amphibians a more specific bacteria-fungi interaction can be found. Thecutaneous bacterium Janthinobacterium lividum protects its amphibian hosts against the fungusBatrachochytrium dendrobatidis, which causes the lethal disease chytridiomycosis (22, 66). The bac-terial secondary metabolites violacein (Figure 2) and indole-3-carboxaldehyde have been identi-fied as the active compounds on the skin of the red-backed salamander (Plethodon cinereus) (22) andmountain yellow-legged frog (Rana muscosa) (66). Knowledge about this beneficial bacteria-fungiantagonism could prove to be helpful to cope with chytridiomycosis. This disease is an impor-tant issue as it threatens about one-third of the amphibian population worldwide with extinction.Becker et al. (10) and Harris et al. (66) demonstrated independently that amphibians treated withJ. lividum are protected against a B. dendrobatidis infection. Hence, a carefully considered use ofJ. lividum as a biological chytridiomycosis protectant could help control the disease and preventthe decline of amphibian populations. Although other amphibian skin-inhabiting bacteria havebeen discovered, J. lividum has been studied most intensely (10, 21, 22, 66).
In the marine environment few bacteria-fungi interactions that impact animals have beenreported. Gil-Turnes et al. (55) discovered a symbiosis between Alteromonas sp. and embryos of theshrimp Palaemon macrodactylus, in which bacteria inhabiting the shrimps’ surface confer resistanceagainst infections of the fungus Lagenidium callinectes by producing 2,3-indolinedione (55).
A more complex interaction between bacteria and fungi occurs in association with the southernpine beetle (Dendroctonus frontalis). The beetle harbors the symbiotic fungus Entomocorticum sp. Aand inoculates the ovipositional galleries with the symbiont. Within these galleries, Entomocorticumsp. A supplies the developing beetle larvae with nutrition. However, this mutualistic relationshipis endangered by the pathogenic fungus Ophiostoma minus, as this fungus is capable of overgrow-ing Entomocorticum sp. A (84). Scott et al. (147) reported that the beneficial mutualism betweenD. frontalis and Entomocorticum sp. A is protected by a third participant, a previously unknownactinomycete, that produces the antifungal compound mycangimycin (Figure 2). This naturalproduct inhibits the antagonistic fungus O. minus but has only minor effects on the symbiontEntomocorticum sp. A (147).
The probably best-studied and most sophisticated insect-bacteria-fungus interaction is the tri-partite mutualism between the leaf-cutting ants (Acromyrmex spp.), the garden fungus Leucoagaricusgongylophorus, and actinomycete bacteria. The ants grow the fungus L. gongylophorus in so-calledfungal gardens and supply it with food. In turn, L. gongylophorus serves itself as a nutritional sourcefor the ants. In order to protect their fungal gardens from necrotrophic fungi such as Escovopsis spp.,ants have formed a symbiosis with antibiotic-producing Actinobacteria of the genera Streptomyces,Pseudonocardia, and Amycolatopsis (8). These bacteria were acquired by either vertical transmission(Pseudonocardia) or sampling from the soil (Streptomyces and Amycolatopsis) (8). Several antifungalsecondary metabolites, such as candicidin (63), dentigerumycin (115), valinomycin, antimycin,actinomycin (144), and a nystatin-like compound (8), have been isolated (Figure 2). Yet, Seipkeet al. (148) showed that there are still more natural products remaining to be discovered fromActinobacteria engaged in symbioses with insects. Sen et al. (149) gave this story an interestingtwist; they showed that the secondary metabolites produced by the investigated bacterial species
www.annualreviews.org • Molecular Bacteria-Fungi Interactions 383
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
Lactic acid bacteria(LAB): gram-positivebacteria that producelactic acid as the mainend product duringfermentation
Pseudonocardia and Amycolatopsis display nonspecific broad antifungal activity. Subsequently, theproduced natural products can inhibit or kill the beneficial fungus L. gongylophorus as well. Hence,ants could damage their fungal garden by subjecting them to actinobacterial secretions.
BACTERIA-FUNGI INTERACTIONS: EFFECTS ON FOODPRODUCTION AND SAFETY
Many food fermentations are carried out with mixed consortia of microorganisms, which can beyeasts, filamentous fungi, lactic acid bacteria (LAB), and/or acetic acid bacteria. These coculturescontribute to the aroma, flavor, and texture of food, and usually they also enhance the shelf life offermented products (39) (Figure 2). Mixed bacteria-fungus fermenting cultures are employed inthe production of alcoholic beverages such as wine (46) and beer (134), dairy products (161) suchas cheese (34) and kefir (99), and sourdough (52, 159), among others. However, investigationsof the molecular interactions between bacteria and fungi during fermentation are limited. Thegrowth of bacterial starter cultures can be enhanced by yeasts, but the molecular mechanism hasnot yet been elucidated (34, 161).
The molecular interplay between antifungal metabolite-producing LAB and spoilage fungiin baked goods, beer, and cheese has been investigated in more detail (39, 52, 134, 135, 159).Organic acids (lactic, acetic, propionic, and phenyllactic), hydroxy fatty acids, hydrogen peroxide,cyclic dipeptides, and reuterin, as well as uncharacterized phenolic and proteinaceous substances,are antifungal compounds produced by LAB (39) (Figure 3). In addition, yeasts can inhibit thegrowth of the spoilage bacteria Listeria monocytogenes in smear cheese (27, 57). Increasing effort isbeing devoted to research on fermenting microorganisms that inhibit spoilage bacteria and fungi,as the demand for additive-free food rises. Moreover, the usage of inhibitory starter cultures couldreduce the risk of food spoilage by mycotoxin-producing molds or pathogenic bacteria. Such foodpoisoning represents a substantial public health risk and causes severe economic losses. Anotheremerging research field to counter food poisoning is the biotransformation and biodegradationof mycotoxins by microorganisms (78) (Figure 2). Mycotoxins such as aflatoxin, zearalenone,fumonisin, and trichothecene contaminate food and feedstuff, leading to toxic reactions uponuptake. Several bacteria are able to transform or even degrade such fungal toxins (75). However,the microbial degradation processes have to be carefully evaluated as not all biotransformationreactions warrant a detoxification of the mycotoxins. Furthermore, it has to be assessed whether thetransformed product is still stable after uptake by humans and animals. Although many investigatedbacteria show promising transformation and degradation abilities, the usage of biotransformingenzymes isolated from these organisms seems to be more advantageous to treatments with livingmicroorganisms (78).
In contrast to the beneficial effects of microbial cocultures for food production, bacteria-fungiinteractions may also cause food spoilage and thus have a major impact on human health (89)(Figure 3). Tempe bongkrek, a popular, coconut-derived dish in Southeast Asia, is produced byfermentation of coconut press cake using the fungus Rhizopus oligosporus (23, 51). Unfortunately,the fungal culture is often contaminated with a bacterium, Burkholderia gladioli pv. cocovene-nans (32). These bacteria are known to produce two toxins, bongkrekic acid (bon) and toxoflavin,which account for a number of lethal intoxications after consumption of tempe bongkrek (51).Bongkrekic acid is a polyunsaturated fatty acid and efficiently inhibits oxidative phosphorylationin mitochondria through blockage of adenine nucleotide translocation (41). Oral intake causesstrong hyperglycemia followed by hypoglycemia and death. The foodborne toxin is derived froma polyketide pathway, as was found by stable isotope labeling (131) and molecular studies of bonbiosynthesis (104). Through sequencing of the genome of B. gladioli and functional analyses of
384 Scherlach · Graupner · Hertweck
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
Fungus
Bacteria
Foods
Aroma
Flavor
Texture
Biotransformation ofmycotoxins
Unknowncompound
Rhizoxin S2
Rhizonin
Toxoflavin
Reuterin
Propionic acid
cyclo(Phe-Pro)
ON
OCH3
O
O
OHO
OHCOOH
NHHN
H3CN
H3CN
NH
HNNCH3
CH3
NCH3
OO
O
OO
O
OO O
N
N
NNO
O
O
O
O
OO
OCH3
H
H
HN
N
O
OH
OHO
O
OH
Aflatoxin
Figure 3Schematic illustration of bacteria-fungi interactions that affect food production and spoilage. Double-headed arrows indicate mutualinteraction; bolts indicate spoilage (dashed bolts indicate decreased spoilage due to bacteria-fungi interaction; bold bolts indicateincreased spoilage due to bacteria-fungi interaction); stop lines indicate inhibition.
trans-AT PKS:a subclass of modulartype I polyketidesynthases in which themodules lackindividual AT domains
the bon gene locus new insights into the complex polyketide assembly by a trans-AT PKS weregained. The other toxin produced by the contamination is the azapteridine toxoflavin, a yellowpigment that was discovered as early as 1934. Latuasan & Berends (95) postulated that toxoflavinis poisonous due to the generation of hydrogen peroxide. Toxoflavin is also considered the mainvirulence factor of Burkholderia glumae, a bacterium causing bacterial panicle blight of rice (64).
Two other metabolites of fungus-associated Burkholderia spp., rhizonin and rhizoxin, are ofsimilar toxicological importance. Actually, the cyclopeptide rhizonin was believed to be the firstmycotoxin identified from the Zygomycota (81, 163). However, rhizonin is in fact produced byendosymbiotic bacteria living in the fungal cytosol of Rhizopus microsporus, as had been shown
www.annualreviews.org • Molecular Bacteria-Fungi Interactions 385
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
Sufu: a fermentedsoybean productproduced by solid statefermentation of tofufollowed by an agingstep in brinecontaining salt andalcohol
Mucormycosis:fungal infection causedby Mucorales
previously for the phytotoxin rhizoxin (118). Because some subspecies of R. microsporus are usedfor the fermentation of soybeans (e.g., for sufu and tempe production), a potential risk for humanhealth arises from the toxinogenic symbiosis. Rhizonin is produced by Burkholderia endofungorum,the endosymbiont of a R. microsporus strain originally isolated from groundnuts in Mozambique(119). The cyclic peptide is highly toxic to mammals as it causes serious hepatic lesions (163).The cytotoxic polyketide rhizoxin from Burkholderia rhizoxinica is produced during fermentationof soybeans with Rhizopus sp. for sufu production (132). Because some of the produced derivativesbelong to the strongest antimitotic agents known to date (141), a severe health risk may arisefor the consumer. These results underline the urgent necessity to consider potential detrimentaleffects resulting from bacteria-fungi interactions during food production to warrant food safety(89).
BACTERIA-FUNGI INTERACTIONS: EFFECTS ON MEDICINE
Bacteria and Fungi as Coacting Human Pathogens
Even though humans are colonized by numerous microorganisms, little is known about the molec-ular interaction among these organisms. However, this research area deserves particular attentionbecause often mixed-species infections generate a complex scenario. The significance of under-standing microbial interactions within human hosts is highlighted even further by the fact thatthe disease outcome of mixed bacteria-fungi infections can differ from single-species infections(124), so that alternative treatment methods have to be applied. Usually, opportunistic pathogensare involved in such multispecies infections. As a consequence, immunocompromised individualsare more prone to becoming infected. The bacteria-fungi interactions during an infection canbe characterized as either neutral (the interaction has no impact on the disease outcome), syner-gistic (bacterial and fungal pathogens act together against the human host), or antagonistic (themicroorganisms inhibit each other).
Rhizopus oryzae is responsible for about 60–80% of all human mucormycosis cases (75). Takinginto consideration that Rhizopus fungi have been found in association with endosymbiotic, toxin-producing bacteria (89), it appears plausible that R. oryzae too may harbor endosymbionts. Exam-inations of clinical isolates identified bacteria-associated fungi as well as bacteria-free fungi (75,117). Utilizing mouse and fly models, Ibrahim et al. (75) investigated the impact of the endosym-biotic bacteria on the infection outcome. No difference in virulence was observed between thefungi harboring bacterial endosymbionts and those without bacteria, although the endobacteria-produced rhizoxin displays cytotoxic activity itself. Apparently, the Rhizopus sp.–Burkholderia sp.interaction is an example of a neutral interaction during infection (Figure 4). Still, it remainsunclear whether the same behavior can be expected during human infections.
Several synergistic interactions between human pathogenic fungi and bacteria have been dis-covered and described. One intensely studied interaction is the coinfection of the yeast Cryptococcusneoformans and the bacterium Klebsiella aerogenes (Figure 4). Thereby, melanin is an important vir-ulence factor for C. neoformans (28). Surprisingly, the yeast is not capable of synthesizing melaninon its own, so that C. neoformans depends on exogenous substrate. K. aerogenes supplies dopamine,which can be utilized for melanization by C. neoformans (47). The pigmentation protects the mi-croorganisms not only against environmental stress but also against the human immune defense(28). Another example of enhanced fungal virulence due to interspecies interplay is mixed biofilmsof the yeast Candida albicans and the bacterium Streptococcus gordonii, which occur together in theoral cavity (Figure 4). It was shown that S. gordonii promotes hyphal growth and biofilm formationof C. albicans (6). Both filamentous growth and biofilm formation contribute to the virulence of
386 Scherlach · Graupner · Hertweck
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
Fungus
Human
Bacteria
No BFI
Neutral BFI
Antagonistic BFI
Synergistic BFI
Rhizoxi
n S2
Farnesol
Pyocyanin
3-oxo-C12-HSL
Melanin
AI-2
Dopamine
ON
OCH 3
O OOH
OO
H COO
H
NCH
3
N
OH
NH
O
O
OO
OH
HN
O
ON
HHO
HO
NH2
O
OBO
HO
HO OH
HO
Figure 4Different medically important bacteria-fungi interactions (BFI). Double-headed arrows indicate interaction;bolts indicate disease (dashed bolts indicate less virulence due to bacteria-fungi interaction; bold boltsindicate enhanced virulence due to bacteria-fungi interaction); stop lines indicate inhibition. Abbreviations:3-oxo-C12-HSL, 3-oxo-C12-homoserine lactone; AI-2, autoinducer-2.
Autoinducer-2:small, diffusiblequorum-sensingmolecule forinterspeciescommunication
the yeast (124). Bamford et al. (6) demonstrated that this interaction occurs via both physical andchemical signals. The physical interaction takes place via adherence, and the interspecies signalmolecule autoinducer-2 serves as chemical signal (6).
The yeast C. albicans is involved in antagonistic bacteria-fungi interactions, too. An intenselystudied opponent of C. albicans is Pseudomonas aeruginosa (Figure 4). Both opportunistic pathogenshave been frequently isolated from burn victims (62) and cystic fibrosis patients (74). P. aeruginosaproduces a range of secondary metabolites that inhibit or even kill C. albicans. One of thesenatural products is the antifungal phenazine pyocyanin (124). Notably, the pyocyanin precursor5-methyl-phenazine-1-carboxylic acid (5MPCA) is even more potent than its end product (54).Morales et al. (105) revealed the mechanism by which the antifungal activity is conferred. Theredox-active 5MPCA generates reactive oxygen species (ROS) such as H2O2 and O2
−. In addition,5MPCA reacts with amine moieties within proteins, thus impairing important cellular structures
www.annualreviews.org • Molecular Bacteria-Fungi Interactions 387
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
Quorum-sensing(QS) molecules:small diffusiblemolecules employedby bacteria to regulatetheir cell-density-dependent behavior
Silent biosyntheticpathways: conditionin which secondarymetabolite geneclusters are notexpressed (e.g., undercommon laboratoryculture conditions)
and enzymes. The covalently bound 5MPCA derivatives keep their redox activity and hence theirability to produce ROS. Both effects promote fungal cell death (105).
In addition, C. albicans and P. aeruginosa interact via the quorum-sensing (QS) molecules far-nesol and 3-oxo-C12-homoserine lactone (3-oxo-C12-HSL), which are produced by C. albicansand P. aeruginosa, respectively (73). The fungal QS molecule inhibits the swarming ability ofP. aeruginosa (102) and reduces the production of the antifungal pyocyanin (37). The bacterial QSmolecule 3-oxo-C12-HSL is structurally related to farnesol and might act as a structural analog(73). 3-oxo-C12-HSL reduces the yeast’s virulence by repressing filamentation (73). Likewise,other bacteria, such as Xanthomonas campestris, Burkholderia cenocepacia, and Streptococcus mutans,produce farnesol mimics that repress hyphal growth of C. albicans. These interspecies signals havebeen identified as cis-2-methyl-11-dodecenoic acid (162), cis-2-dodecenoic acid (18), and trans-2-decenoic acid (160), respectively. Furthermore, S. mutans produces another QS molecule, thecompetence-stimulating peptide, which represses germ tube formation of C. albicans (80).
Investigations of these diverse bacteria-fungi interactions occurring in humans have shownthat the outcome of an interspecies infection ranges from virulence enhancement to antagonisticinteraction. Therefore, treatment for mixed-species infection should be carefully measured, as anincreasing growth of C. albicans, for example, was observed after antibacterial therapy of a mixedinfection of P. aeruginosa and the yeast (26). However, in infections where bacteria contribute tothe virulence of a fungus, an additional antibacterial treatment might be required. Also, furtherresearch on molecular antagonistic interactions might serve new treatment options or hithertounknown points of attack.
Exploiting Bacteria-Fungi Interactions for Drug Discovery
The interplay between different microorganisms at the molecular level has generated a vast chem-ical diversity. The complex interactions of fungi and bacteria in nature are mediated by secondarymetabolites that have a specific function for the producing organism, for example, to ensure sur-vival in a competitive environment. Therefore, many microorganisms have a huge biosyntheticpotential as has been revealed by various whole-genome sequencing projects (69, 164). The expres-sion of the biosynthetic gene clusters is usually tightly regulated to meet the varying requirementsof a changing environment, and it appears that most natural product biosynthesis genes are silentin pure cultures. Thus, the majority of secondary metabolites remains untapped.
Surprisingly, only a limited number of studies have exploited the potential of bacteria-fungiinteractions for drug discovery. Although coculturing approaches have been widely used forindustrial purposes (e.g., production of food and beverages), little is known about secondarymetabolism in cocultures (140). Several years ago it was shown that mixed fermentations couldhave an influence on the production of secondary metabolites. Whereas initially the impact of thecocultured organism was assessed mainly by an increase in the biological (i.e., antibiotic) activity ofa culture extract or a yield improvement in the production of a certain compound (125), followingresearch focused on the discovery of new natural products resulting from either combinedbiosynthetic pathways or the activation of previously silent pathways. Cueto et al. (36) cultured amarine Pestalotia species together with an unidentified, antibiotic-resistant marine bacterium andthus elicited the biosynthesis of the benzophenone pestalone. The compound, which is producedonly in coculture, displays potent antibacterial activity against methicillin-resistant Staphylo-coccus aureus and vancomycin-resistant Enterococcus faecium. In another study, a marine-derivedEmericella species was challenged by coculturing it with the actinomycete Salinispora arenicola(114). Thus, the production of two new cyclic depsipeptides, emericellamides A and B, wasenhanced 100-fold, which enabled their structural elucidation. Emericellamides possess moderate
388 Scherlach · Graupner · Hertweck
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
Histoneacetyltransferases:acetylate-conservedlysine residues ofhistone
antibacterial activities against methicillin-resistant S. aureus. The emericellamide biosyntheticgenes were discovered via a gene deletion approach in the model organism Aspergillus nidulans,offering the possibility to engineer novel analogues (29). Four new diterpenoids, libertellenonesA–D, were discovered by coculturing the marine fungus Libertella sp. with an unidentified marinebacterium (113). This bacterium seemed to specifically induce the formation of these compoundsin this fungal species as no new metabolites could be detected in 49 other cocultures with thesame bacterium. Zuck et al. (167) described the production of formyl-xanthocillin analogues byusing a mixed culture of Aspergillus fumigatus and Streptomyces peucetius.
In a more systematic approach, Schroeckh et al. (146) employed microarray-based monitoringto study the expression of cryptic biosynthetic gene clusters in the model fungus A. nidulansafter induction through the interaction with actinomycetes sharing the same habitat. Of 58 testedspecies, one strain (Streptomyces rapamycinicus) was detected that triggers the formation of thearchetypal polyketide orsellinic acid along with the typical lichen metabolite lecanoric acid andtwo complex polyphenols with anti-osteoporosis activity. Molecular genetic studies allowed forthe first time the identification of the long-sought-after genetic locus for the biosynthesis oforsellinic acid, one of the simplest fungal polyketides. Further insights into the mechanism ofthe intimate interaction of both organisms were gained by studying the regulation of fungalgene expression. By a systematic deletion of 36 out of 40 acetyltransferase-encoding genes andby chromatin immunoprecipitation experiments, it was revealed that the bacterium induces ahistone modification in the fungus via the main histone acetyltransferase complex Saga/Ada (111).These results demonstrate the complexity of bacteria-fungi interactions and imply the yet-to-be-discovered potential of studying microbial cross talk.
CONCLUSIONS AND PERSPECTIVES
Various examples of bacteria-fungi interactions highlighted in this review illustrate that mixedconsortia of microorganisms are widespread and can be found in a variety of places including soil,plants, animals, and humans and in food production. The more we understand these interactionsamong bacteria and fungi, the more we realize that such interplays affect not only environmentalprocesses (e.g., growth of plants or diseases of plants, animals, fungi), but also our everyday life (e.g.,food production, food spoilage, human disease control). However, the molecular basis of someimportant interkingdom interactions remains elusive. Moreover, one can presume that a plethoraof bacteria-fungi interactions awaits discovery. From a translational point of view, knowledge ofbacteria-fungi interactions can be exploited for beneficial use in many ways. Microorganisms thatcontrol growth of spoilage or pathogenic organisms can replace synthetic chemicals used in thefood industry or during animal husbandry, fulfilling consumer demand for additive-free food andorganic farming. Obviously, the application of living organisms must be carefully considered. Insome cases it might be more suitable to use specific enzymes or natural products isolated froman organism instead of the whole microbe. Natural product discovery has always been essentialfor drug development. Utilizing cocultures in natural product-screening processes can aid thediscovery of novel secondary metabolites that are not produced in the absence of particular stimuli.Another important medical benefit that can be gained from understanding the molecular basicsof bacteria-fungi interactions involved in human diseases is the identification of new therapeutictargets to combat pathogens.
However, humans will be able to take advantage of bacteria-fungi interactions only ifthey fully appreciate the molecular interplay. We expect future research to contribute greatlyto our understanding of bacteria-fungi interactions and to provide new avenues for novelapplications.
www.annualreviews.org • Molecular Bacteria-Fungi Interactions 389
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships, funding, or financial holdings thatmight be perceived as affecting the objectivity of this review.
ACKNOWLEDGMENTS
K.S. and K.G. contributed equally to this review. We thank the International Leibniz ResearchSchool for Microbial and Biomolecular Interactions (ILRS) and the Jena School for MicrobialCommunication ( JSMC) of the German Excellence Initiative for supporting the authors’ originalresearch in this area.
LITERATURE CITED
1. Ahmadjian V, Reynolds JT. 1961. Production of biologically active compounds by isolated lichenizedfungi. Science 133:700–1
2. Akiyama K, Matsuzaki K, Hayashi H. 2005. Plant sesquiterpenes induce hyphal branching in arbuscularmycorrhizal fungi. Nature 435:824–27
3. Andolfi A, Cimmino A, Cantore PL, Iacobellis NS, Evidente A. 2008. Bioactive and structural metabolitesof Pseudomonas and Burkholderia species causal agents of cultivated mushrooms diseases. Perspect. Med.Chem. 2:81–112
4. Arnold AE, Mejia LC, Kyllo D, Rojas EI, Maynard Z, et al. 2003. Fungal endophytes limit pathogendamage in a tropical tree. Proc. Natl. Acad. Sci. USA 100:15649–54
5. Artursson V, Finlay RD, Jansson JK. 2006. Interactions between arbuscular mycorrhizal fungi and bac-teria and their potential for stimulating plant growth. Environ. Microbiol. 8:1–10
6. Bamford CV, d’Mello A, Nobbs AH, Dutton LC, Vickerman MM, Jenkinson HF. 2009. Streptococcus gor-donii modulates Candida albicans biofilm formation through intergeneric communication. Infect. Immun.77:3696–704
7. Bandara WM, Seneviratne G, Kulasooriya SA. 2006. Interactions among endophytic bacteria and fungi:effects and potentials. J. Biosci. 31:645–50
8. Barke J, Seipke RF, Gruschow S, Heavens D, Drou N, et al. 2010. A mixed community of actinomycetesproduce multiple antibiotics for the fungus farming ant Acromyrmex octospinosus. BMC Biol. 8:109
9. Bassarello C, Lazzaroni S, Bifulco G, Lo Cantore P, Iacobellis NS, et al. 2004. Tolaasins A-E, five newlipodepsipeptides produced by Pseudomonas tolaasii. J. Nat. Prod. 67:811–16
10. Becker MH, Brucker RM, Schwantes CR, Harris RN, Minbiole KPC. 2009. The bacterially producedmetabolite violacein is associated with survival of amphibians infected with a lethal fungus. Appl. Environ.Microbiol. 75:6635–38
11. Berdy J. 2005. Bioactive microbial metabolites. J. Antibiot. 58:1–2612. Besserer A, Puech-Pages V, Kiefer P, Gomez-Roldan V, Jauneau A, et al. 2006. Strigolactones stimulate
arbuscular mycorrhizal fungi by activating mitochondria. PLoS Biol. 4:e22613. Bharadwaj DP, Lundquist PO, Persson P, Alstrom S. 2008. Evidence for specificity of cultivable bacteria
associated with arbuscular mycorrhizal fungal spores. FEMS Microbiol. Ecol. 65:310–2214. Bianciotto V, Andreotti S, Balestrini R, Bonfante P, Perotto S. 2001. Extracellular polysaccharides
are involved in the attachment of Azospirillum brasilense and Rhizobium leguminosarum to arbuscularmycorrhizal structures. Eur. J. Histochem. 45:39–49
15. Bianciotto V, Genre A, Jargeat P, Lumini E, Becard G, Bonfante P. 2004. Vertical transmission ofendobacteria in the arbuscular mycorrhizal fungus Gigaspora margarita through generation of vegetativespores. Appl. Environ. Microbiol. 70:3600–8
16. Bianciotto V, Lumini E, Bonfante P, Vandamme P. 2003. ‘Candidatus Glomeribacter gigasporarum’ gen.nov., sp. nov., an endosymbiont of arbuscular mycorrhizal fungi. Int. J. Syst. Evol. Microbiol. 53:121–24
17. Bonfante P, Anca IA. 2009. Plants, mycorrhizal fungi, and bacteria: a network of interactions. Annu. Rev.Microbiol. 63:363–83
390 Scherlach · Graupner · Hertweck
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
18. Boon C, Deng Y, Wang LH, He Y, Xu JL, et al. 2008. A novel DSF-like signal from Burkholderiacenocepacia interferes with Candida albicans morphological transition. ISME J. 2:27–36
19. Boustie J, Tomasi S, Grube M. 2011. Bioactive lichen metabolites: alpine habitats as an untapped source.Phytochem. Rev. 10:287–307
20. Brodey CL, Rainey PB, Tester M, Johnstone K. 1991. Bacterial blotch disease of the cultivated mushroomis caused by an ion channel forming lipodepsipeptide toxin. Mol. Plant Microbe Interact. 4:407–11
21. Brucker RM, Baylor CM, Walters RL, Lauer A, Harris RN, Minbiole KP. 2008. The identification of2,4-diacetylphloroglucinol as an antifungal metabolite produced by cutaneous bacteria of the salamanderPlethodon cinereus. J. Chem. Ecol. 34:39–43
22. Brucker RM, Harris RN, Schwantes CR, Gallaher TN, Flaherty DC, et al. 2008. Amphibian chemi-cal defense: antifungal metabolites of the microsymbiont Janthinobacterium lividum on the salamanderPlethodon cinereus. J. Chem. Ecol. 34:1422–29
23. Buckle KA, Kartadarma EK. 1990. Inhibition of bongkrek acid and toxoflavin production in tempebongkrek containing Pseudomonas cocovenenans. J. Appl. Bacteriol. 68:571–76
24. Budi SW, van Tuinen D, Martinotti G, Gianinazzi S. 1999. Isolation from the Sorghum bicolor mycorrhi-zosphere of a bacterium compatible with arbuscular mycorrhiza development and antagonistic towardssoilborne fungal pathogens. Appl. Environ. Microbiol. 65:5148–50
25. Burkholder PR, Evans AW, McVeigh I, Thornton HK. 1944. Antibiotic activity of lichens. Proc. Natl.Acad. Sci. USA 30:250–55
26. Burns JL, Van Dalfsen JM, Shawar RM, Otto KL, Garber RL, et al. 1999. Effect of chronic intermit-tent administration of inhaled tobramycin on respiratory microbial flora in patients with cystic fibrosis.J. Infect. Dis. 179:1190–96
27. Callon C, Saubusse M, Didienne R, Buchin S, Montel MC. 2011. Simplification of a complex microbialantilisterial consortium to evaluate the contribution of its flora in uncooked pressed cheese. Int. J. FoodMicrobiol. 145:379–89
28. Casadevall A, Rosas AL, Nosanchuk JD. 2000. Melanin and virulence in Cryptococcus neoformans. Curr.Opin. Microbiol. 3:354–58
29. Chiang YM, Szewczyk E, Nayak T, Davidson AD, Sanchez JF, et al. 2008. Molecular genetic miningof the Aspergillus secondary metabolome: discovery of the emericellamide biosynthetic pathway. Chem.Biol. 15:527–32
30. Cho KH, Kim ST, Kim YK. 2007. Purification of a pore-forming peptide toxin, tolaasin, produced byPseudomonas tolaasii 6264. J. Biochem. Mol. Biol. 40:113–18
31. Citernesi AS, Filippi C, Bagnoli G, Giovannetti M. 1994. Effects of the antimycotic molecule Iturin A2,secreted by Bacillus subtilis strain M51, on arbuscular mycorrhizal fungi. Microbiol. Res. 149:241–46
32. Coenye T, Holmes B, Kersters K, Govan JR, Vandamme P. 1999. Burkholderia cocovenenans (van Dammeet al. 1960) Gillis et al. 1995 and Burkholderia vandii Urakami et al. 1994 are junior synonyms of Burkholde-ria gladioli (Severini 1913) Yabuuchi et al. 1993 and Burkholderia plantarii (Azegami et al. 1987) Urakamiet al. 1994, respectively. Int. J. Syst. Bacteriol. 49(Pt. 1):37–42
33. Coraiola M, Lo Cantore P, Lazzaroni S, Evidente A, Iacobellis NS, Dalla Serra M. 2006. WLIP andtolaasin I, lipodepsipeptides from Pseudomonas reactans and Pseudomonas tolaasii, permeabilise model mem-branes. Biochim. Biophys. Acta 1758:1713–22
34. Corsetti A, Rossi J, Gobbetti M. 2001. Interactions between yeasts and bacteria in the smear surface-ripened cheeses. Int. J. Food Microbiol. 69:1–10
35. Cox PA, Banack SA, Murch SJ, Rasmussen U, Tien G, et al. 2005. Diverse taxa of cyanobacteria producebeta-N-methylamino-L-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 102:5074–78
36. Cueto M, Jensen PR, Kauffman C, Fenical W, Lobkovsky E, Clardy J. 2001. Pestalone, a new antibioticproduced by a marine fungus in response to bacterial challenge. J. Nat. Prod. 64:1444–46
37. Cugini C, Calfee MW, Farrow JM 3rd, Morales DK, Pesci EC, Hogan DA. 2007. Farnesol, a commonsesquiterpene, inhibits PQS production in Pseudomonas aeruginosa. Mol. Microbiol. 65:896–906
38. Culberson CF, Armaleo D. 1992. Induction of a complete secondary-product pathway in cultures of alichen fungus. Exp. Mycol. 16:52–63
39. Excellent surveyreviewing the role ofLAB in food safety.39. Dalie DKD, Deschamps AM, Richard-Forget F. 2010. Lactic acid bacteria—potential for control
of mould growth and mycotoxins: a review. Food Control 21:370–80
www.annualreviews.org • Molecular Bacteria-Fungi Interactions 391
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
40. de Boer W, Folman LB, Summerbell RC, Boddy L. 2005. Living in a fungal world: impact of fungi onsoil bacterial niche development. FEMS Microbiol. Ecol. 29:795–811
41. de Bruijn J, Frost DJ, Nugteren DH, Gaudemer A, Lijmbach GWM, Cox HC. 1973. The structure ofbongkrekic acid. Tetrahedron 29:1541–47
42. de Weert S, Kuiper I, Lagendijk EL, Lamers GE, Lugtenberg BJ. 2004. Role of chemotaxis toward fusaricacid in colonization of hyphae of Fusarium oxysporum f. sp. radicis-lycopersici by Pseudomonas fluorescensWCS365. Mol. Plant Microbe Interact. 17:1185–91
43. DePriest PT. 2004. Early molecular investigations of lichen-forming symbionts: 1986–2001. Annu. Rev.Microbiol. 58:273–301
44. Dittmann E, Fewer DP, Neilan BA. 2013. Cyanobacterial toxins: biosynthetic routes and evolutionaryroots. FEMS Microbiol. Rev. 37:23–43
45. Dittmann E, Wiegand C. 2006. Cyanobacterial toxins—occurrence, biosynthesis and impact on humanaffairs. Mol. Nutr. Food Res. 50:7–17
46. Fleet GH. 2003. Yeast interactions and wine flavour. Int. J. Food Microbiol. 86:11–2247. Frases S, Chaskes S, Dadachova E, Casadevall A. 2006. Induction by Klebsiella aerogenes of a melanin-like
pigment in Cryptococcus neoformans. Appl. Environ. Microbiol. 72:1542–5048. Reviews variousaspects of bacteria-fungiinterplay and its effecton other organisms andthe environment.
48. Frey-Klett P, Burlinson P, Deveau A, Barret M, Tarkka M, Sarniguet A. 2011. Bacterial-fungalinteractions: hyphens between agricultural, clinical, environmental, and food microbiologists.Microbiol. Mol. Biol. Rev. 75:583–609
49. Frey-Klett P, Garbaye J, Tarkka M. 2007. The mycorrhiza helper bacteria revisited. New Phytol. 176:22–36
50. Garbaye J. 1994. Helper bacteria: a new dimension to the mycorrhizal symbiosis. New Phytol. 128:197–210
51. Garcia RA, Hotchkiss JH, Steinkraus KH. 1999. The effect of lipids on bongkrekic (Bongkrek) acid toxinproduction by Burkholderia cocovenenans in coconut media. Food Addit. Contam. 16:63–69
52. Gerez CL, Torino MI, Rollan G, de Valdez GF. 2009. Prevention of bread mould spoilage by usinglactic acid bacteria with antifungal properties. Food Control 20:144–48
53. Ghignone S, Salvioli A, Anca I, Lumini E, Ortu G, et al. 2012. The genome of the obligate endobacteriumof an AM fungus reveals an interphylum network of nutritional interactions. ISME J. 6:136–45
54. Gibson J, Sood A, Hogan DA. 2009. Pseudomonas aeruginosa–Candida albicans interactions: localizationand fungal toxicity of a phenazine derivative. Appl. Environ. Microbiol. 75:504–13
55. Gil-Turnes MS, Hay ME, Fenical W. 1989. Symbiotic marine bacteria chemically defend crustaceanembryos from a pathogenic fungus. Science 246:116–18
56. Godfrey SA, Harrow SA, Marshall JW, Klena JD. 2001. Characterization by 16S rRNA sequence analysisof pseudomonads causing blotch disease of cultivated Agaricus bisporus. Appl. Environ. Microbiol. 67:4316–23
57. Goerges S, Aigner U, Silakowski B, Scherer S. 2006. Inhibition of Listeria monocytogenes by food-borneyeasts. Appl. Environ. Microbiol. 72:313–18
58. Goh EB, Yim G, Tsui W, McClure J, Surette MG, Davies J. 2002. Transcriptional modulation of bacterialgene expression by subinhibitory concentrations of antibiotics. Proc. Natl. Acad. Sci. USA 99:17025–30
59. Gonzalez I, Ayuso-Sacido A, Anderson A, Genilloud O. 2005. Actinomycetes isolated from lichens:evaluation of their diversity and detection of biosynthetic gene sequences. FEMS Microbiol. Ecol. 54:401–15
60. Graupner K, Scherlach K, Bretschneider T, Lackner G, Roth M, et al. 2012. Imaging mass spectrometryand genome mining reveal highly antifungal virulence factor of mushroom soft rot pathogen. Angew.Chem. Int. Ed. 51:13173–77
61. Grube M, Berg G. 2009. Microbial consortia of bacteria and fungi with focus on the lichen symbiosis.Fungal Biol. Rev. 23:72–85
62. Gupta N, Haque A, Mukhopadhyay G, Narayan RP, Prasad R. 2005. Interactions between bacteria andCandida in the burn wound. Burns 31:375–78
63. Haeder S, Wirth R, Herz H, Spiteller D. 2009. Candicidin-producing Streptomyces support leaf-cuttingants to protect their fungus garden against the pathogenic fungus Escovopsis. Proc. Natl. Acad. Sci. USA106:4742–46
392 Scherlach · Graupner · Hertweck
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
64. Ham JH, Melanson RA, Rush MC. 2011. Burkholderia glumae: next major pathogen of rice? Mol. PlantPathol. 12:329–39
65. Hara S, Mukae H, Sakamoto N, Ishimoto H, Amenomori M, et al. 2008. Plectasin has antibacterialactivity and no affect on cell viability or IL-8 production. Biochem. Biophys. Res. Commun. 374:709–13
66. Harris RN, Brucker RM, Walke JB, Becker MH, Schwantes CR, et al. 2009. Skin microbes on frogsprevent morbidity and mortality caused by a lethal skin fungus. ISME J. 3:818–24
67. Harrison MJ. 2005. Signaling in the arbuscular mycorrhizal symbiosis. Annu. Rev. Microbiol. 59:19–4268. He H, Bigelis R, Yang HY, Chang LP, Singh MP. 2005. Lichenicolins A and B, new bisnaphthopyrones
from an unidentified lichenicolous fungus, strain LL-RB0668. J. Antibiot. 58:731–3669. Hertweck C. 2009. Hidden biosynthetic treasures brought to light. Nat. Chem. Biol. 5:450–5270. Hildebrandt U, Janetta K, Bothe H. 2002. Towards growth of arbuscular mycorrhizal fungi independent
of a plant host. Appl. Environ. Microbiol. 68:1919–2471. Hildebrandt U, Ouziad F, Marner FJ, Bothe H. 2006. The bacterium Paenibacillus validus stimulates
growth of the arbuscular mycorrhizal fungus Glomus intraradices up to the formation of fertile spores.FEMS Microbiol. Lett. 254:258–67
72. Hoffman MT, Arnold AE. 2010. Diverse bacteria inhabit living hyphae of phylogenetically diverse fungalendophytes. Appl. Environ. Microbiol. 76:4063–75
73. Hogan DA, Vik A, Kolter R. 2004. A Pseudomonas aeruginosa quorum-sensing molecule influences Candidaalbicans morphology. Mol. Microbiol. 54:1212–23
74. Hughes WT, Kim HK. 1973. Mycoflora in cystic fibrosis: some ecologic aspects of Pseudomonas aeruginosaand Candida albicans. Mycopathol. Mycol. Appl. 50:261–69
75. Ibrahim AS, Gebremariam T, Liu M, Chamilos G, Kontoyiannis D, et al. 2008. Bacterial endosymbiosisis widely present among zygomycetes but does not contribute to the pathogenesis of mucormycosis.J. Infect. Dis. 198:1083–90
76. Iwasaki S, Kobayashi H, Furukawa J, Namikoshi M, Okuda S, et al. 1984. Studies on macrocyclic lactoneantibiotics. VII. Structure of a phytotoxin “rhizoxin” produced by Rhizopus chinensis. J. Antibiot. 37:354–62
77. Iwasaki S, Namikoshi M, Kobayashi H, Furukawa J, Okuda S. 1986. Studies on macrocyclic lactoneantibiotics. VIII. Absolute structures of rhizoxin and a related compound. J. Antibiot. 39:424–29
78. Jard G, Liboz T, Mathieu F, Guyonvarc’h A, Lebrihi A. 2011. Review of mycotoxin reduction in foodand feed: from prevention in the field to detoxification by adsorption or transformation. Food Addit.Contam. A 28:1590–609
79. Jargeat P, Cosseau C, Ola’h B, Jauneau A, Bonfante P, et al. 2004. Isolation, free-living capacities,and genome structure of “Candidatus Glomeribacter gigasporarum,” the endocellular bacterium of themycorrhizal fungus Gigaspora margarita. J. Bacteriol. 186:6876–84
80. Jarosz LM, Deng DM, van der Mei HC, Crielaard W, Krom BP. 2009. Streptococcus mutans competence-stimulating peptide inhibits Candida albicans hypha formation. Eukaryot. Cell 8:1658–64
81. Jennessen J, Nielsen KF, Houbraken J, Lyhne EK, Schnurer J, et al. 2005. Secondary metabolite andmycotoxin production by the Rhizopus microsporus group. J. Agric. Food Chem. 53:1833–40
82. Kaasalainen U, Fewer DP, Jokela J, Wahlsten M, Sivonen K, Rikkinen J. 2012. Cyanobacteria producea high variety of hepatotoxic peptides in lichen symbiosis. Proc. Natl. Acad. Sci. USA 109:5886–91
83. Kaltenpoth M, Gottler W, Herzner G, Strohm E. 2005. Symbiotic bacteria protect wasp larvae fromfungal infestation. Curr. Biol. 15:475–79
84. Kleipzig KD, Wilkens RT. 1997. Competitive interaction among symbiotic fungi of the southern pinebeetle. Appl. Environ. Microbiol. 63:621–27
85. Kroiss J, Kaltenpoth M, Schneider B, Schwinger MG, Hertweck C, et al. 2010. Symbiotic streptomycetesprovide antibiotic combination prophylaxis for wasp offspring. Nat. Chem. Biol. 6:261–63
86. Kusari S, Hertweck C, Spiteller M. 2012. Chemical ecology of endophytic fungi: origins of secondarymetabolites. Chem. Biol. 19:792–98
87. Kusebauch B, Busch B, Scherlach K, Roth M, Hertweck C. 2009. Polyketide-chain branching by anenzymatic Michael addition. Angew. Chem. Int. Ed. 48:5001–4
88. Kusebauch B, Busch B, Scherlach K, Roth M, Hertweck C. 2010. Functionally distinct modules operatetwo consecutive α,β→β,γ double-bond shifts in the rhizoxin polyketide assembly line. Angew. Chem.Int. Ed. 49:1460–64
www.annualreviews.org • Molecular Bacteria-Fungi Interactions 393
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
89. Lackner G, Hertweck C. 2011. Impact of endofungal bacteria on infection biology, food safety, and drugdevelopment. PLoS Pathog. 7:e1002096
90. Lackner G, Mobius N, Scherlach K, Partida-Martinez LP, Winkler R, et al. 2009. Global distributionand evolution of a toxinogenic Burkholderia-Rhizopus symbiosis. Appl. Environ. Microbiol. 75:2982–86
91. Lackner G, Moebius N, Hertweck C. 2011. Endofungal bacterium controls its host by an hrp type IIIsecretion system. ISME J. 5:252–61
92. Lackner G, Moebius N, Partida-Martinez L, Hertweck C. 2011. Complete genome sequence ofBurkholderia rhizoxinica, an endosymbiont of Rhizopus microsporus. J. Bacteriol. 193:783–84
93. Lackner G, Moebius N, Partida-Martinez LP, Boland S, Hertweck C. 2011. Evolution of an endofungallifestyle: deductions from the Burkholderia rhizoxinica genome. BMC Genomics 12:210
94. Largeteau ML, Savoie JM. 2010. Microbially induced diseases of Agaricus bisporus: biochemical mecha-nisms and impact on commercial mushroom production. Appl. Microbiol. Biotechnol. 86:63–73
95. Latuasan HE, Berends W. 1961. On the origin of the toxicity of toxoflavin. Biochim. Biophys. Acta 52:502–896. Leone MR, Lackner G, Silipo A, Lanzetta R, Molinaro A, Hertweck C. 2010. An unusual galactofuranose
lipopolysaccharide that ensures the intracellular survival of toxin-producing bacteria in their fungal host.Angew. Chem. Int. Ed. 49:7476–80
97. Lincoln SP, Fermor TR, Tindall BJ. 1999. Janthinobacterium agaricidamnosum sp. nov., a soft rot pathogenof Agaricus bisporus. Int. J. Syst. Bacteriol. 49:1577–89
98. Lo Cantore P, Lazzaroni S, Coraiola M, Dalla Serra M, Cafarchia C, et al. 2006. Biological character-ization of white line-inducing principle (WLIP) produced by Pseudomonas reactans NCPPB1311. Mol.Plant Microbe Interact. 19:1113–20
99. Lopitz-Otsoa F, Rementeria A, Elguezabal N, Garaizar J. 2006. Kefir: a symbiotic yeasts-bacteria com-munity with alleged healthy capabilities. Rev. Iberoam. Micol. 23:67–74
100. Lumini E, Bianciotto V, Jargeat P, Novero M, Salvioli A, et al. 2007. Presymbiotic growth and sporalmorphology are affected in the arbuscular mycorrhizal fungus Gigaspora margarita cured of its endobac-teria. Cell Microbiol. 9:1716–29
101. Magarvey NA, Beck ZQ, Golakoti T, Ding Y, Huber U, et al. 2006. Biosynthetic characterization andchemoenzymatic assembly of the cryptophycins. Potent anticancer agents from cyanobionts. ACS Chem.Biol. 1:766–79
102. McAlester G, O’Gara F, Morrissey JP. 2008. Signal-mediated interactions between Pseudomonas aerugi-nosa and Candida albicans. J. Med. Microbiol. 57:563–69
103. Miransari M. 2011. Interactions between arbuscular mycorrhizal fungi and soil bacteria. Appl. Microbiol.Biotechnol. 89:917–30
104. Moebius N, Ross C, Scherlach K, Rohm B, Roth M, Hertweck C. 2012. Biosynthesis of the respiratorytoxin bongkrekic acid in the pathogenic bacterium Burkholderia gladioli. Chem. Biol. 19:1164–74
105. Elucidates themode of action ofphenazine and itsprecursor 5MPCA.
105. Morales DK, Jacobs NJ, Rajamani S, Krishnamurthy M, Cubillos-Ruiz JR, Hogan DA. 2010.Antifungal mechanisms by which a novel Pseudomonas aeruginosa phenazine toxin kills Candidaalbicans in biofilms. Mol. Microbiol. 78:1379–92
106. Munsch P, Alatossava T, Marttinen N, Meyer JM, Christen R, Gardan L. 2002. Pseudomonas costantiniisp. nov., another causal agent of brown blotch disease, isolated from cultivated mushroom sporophoresin Finland. Int. J. Syst. Evol. Microbiol. 52:1973–83
107. Mygind PH, Fischer RL, Schnorr KM, Hansen MT, Sonksen CP, et al. 2005. Plectasin is a peptideantibiotic with therapeutic potential from a saprophytic fungus. Nature 437:975–80
108. Nazir R, Warmink JA, Boersma H, van Elsas JD. 2010. Mechanisms that promote bacterial fitness infungal-affected soil microhabitats. FEMS Microbiol. Ecol. 71:169–85
109. Noble R, Dobrovin-Pennington A, Hobbs PJ, Pederby J, Rodger A. 2009. Volatile C8 compounds andpseudomonads influence primordium formation of Agaricus bisporus. Mycologia 101:583–91
110. Notz R, Maurhofer M, Dubach H, Haas D, Defago G. 2002. Fusaric acid-producing strains of Fusariumoxysporum alter 2,4-diacetylphloroglucinol biosynthetic gene expression in Pseudomonas fluorescens CHA0in vitro and in the rhizosphere of wheat. Appl. Environ. Microbiol. 68:2229–35
111. Provides the firstexperimental evidencethat bacteria can inducehistone modifications infungi.
111. Nutzmann HW, Reyes-Dominguez Y, Scherlach K, Schroeckh V, Horn F, et al. 2011. Bacteria-induced natural product formation in the fungus Aspergillus nidulans requires Saga/Ada-mediated histone acetylation. Proc. Natl. Acad. Sci. USA 108:14282–87
394 Scherlach · Graupner · Hertweck
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
112. O’Brien J, Wright GD. 2011. An ecological perspective of microbial secondary metabolism. Curr. Opin.Biotechnol. 22:552–58
113. Oh DC, Jensen PR, Kauffman CA, Fenical W. 2005. Libertellenones A-D: induction of cytotoxic diter-penoid biosynthesis by marine microbial competition. Bioorg. Med. Chem. 13:5267–73
114. Oh DC, Kauffman CA, Jensen PR, Fenical W. 2007. Induced production of emericellamides A and Bfrom the marine-derived fungus Emericella sp. in competing co-culture. J. Nat. Prod. 70:515–20
115. Oh DC, Poulsen M, Currie CR, Clardy J. 2009. Dentigerumycin: a bacterial mediator of an ant-fungussymbiosis. Nat. Chem. Biol. 5:391–93
116. Oksanen I, Jokela J, Fewer DP, Wahlsten M, Rikkinen J, Sivonen K. 2004. Discovery of rare andhighly toxic microcystins from lichen-associated cyanobacterium Nostoc sp. strain IO-102-I. Appl. Environ.Microbiol. 70:5756–63
117. Partida-Martinez LP, Bandemer S, Ruchel R, Dannaoui E, Hertweck C. 2008. Lack of evidence ofendosymbiotic toxin-producing bacteria in clinical Rhizopus isolates. Mycoses 51:266–69
118. Partida-Martinez LP, de Looss CF, Ishida K, Ishida M, Roth M, et al. 2007. Rhizonin, the first mycotoxinisolated from Zygomycota, is not a fungal metabolite, but produced by bacterial endosymbionts. Appl.Environ. Microbiol. 73:793–97
119. Partida-Martinez LP, Groth I, Schmitt I, Richter W, Roth M, Hertweck C. 2007. Burkholderia rhizoxinicasp. nov. and Burkholderia endofungorum sp. nov., bacterial endosymbionts of the plant-pathogenic fungusRhizopus microsporus. Int. J. Syst. Evol. Microbiol. 57:2583–90
120. Partida-Martinez LP, Hertweck C. 2005. Pathogenic fungus harbours endosymbiotic bacteria for toxinproduction. Nature 437:884–88
121. Partida-Martinez LP, Hertweck C. 2007. A gene cluster encoding rhizoxin biosynthesis in “Burkholderiarhizoxina”, the bacterial endosymbiont of the fungus Rhizopus microsporus. ChemBioChem 8:41–45
122. Partida-Martinez LP, Monajembashi S, Greulich KO, Hertweck C. 2007. Endosymbiont-dependenthost reproduction maintains bacterial-fungal mutualism. Curr. Biol. 17:773–77
123. Paszkowski U. 2006. A journey through signaling in arbuscular mycorrhizal symbioses 2006. New Phytol.172:35–46
124. Offers an overviewof bacteria-fungiinteractionscontributing to humaninfections.
124. Peleg AY, Hogan DA, Mylonakis E. 2010. Medically important bacterial-fungal interactions.Nat. Rev. Microbiol. 8:340–49
125. Pettit RK. 2009. Mixed fermentation for natural product drug discovery. Appl. Microbiol. Biotechnol.83:19–25
126. Porras-Alfaro A, Bayman P. 2011. Hidden fungi, emergent properties: endophytes and microbiomes.Annu. Rev. Phytopathol. 49:291–315
127. Poulsen M, Oh DC, Clardy J, Currie CR. 2011. Chemical analyses of wasp-associated streptomycesbacteria reveal a prolific potential for natural products discovery. PLoS One 6:e16763
128. Rainey PB, Brodey CL, Johnstone K. 1991. Biological properties and spectrum of activity of tolaasin,a lipodepsipeptide toxin produced by the mushroom pathogen Pseudomonas tolaasii. Physiol. Mol. PlantPathol. 39:57–70
129. Rainey PB, Cole ALJ, Fermor TR, Wood DA. 1990. A model system for examining involvment ofbacteria in basidiome initiation of Agaricus bisporus. Mycol. Res. 94:191–95
130. Riedlinger J, Schrey SD, Tarkka MT, Hampp R, Kapur M, Fiedler HP. 2006. Auxofuran, a novelmetabolite that stimulates the growth of fly agaric, is produced by the mycorrhiza helper bacteriumStreptomyces strain AcH 505. Appl. Environ. Microbiol. 72:3550–57
131. Rohm B, Scherlach K, Hertweck C. 2010. Biosynthesis of the mitochondrial adenine nucleotide translo-case (ATPase) inhibitor bongkrekic acid in Burkholderia gladioli. Org. Biomol. Chem. 8:1520–22
132. Rohm B, Scherlach K, Mobius N, Partida-Martinez LP, Hertweck C. 2010. Toxin production by bacterialendosymbionts of a Rhizopus microsporus strain used for tempe/sufu processing. Int. J. Food Microbiol.136:368–71
133. Rokni-Zadeh H, Li W, Sanchez-Rodriguez A, Sinnaeve D, Rozenski J, et al. 2012. Genetic and func-tional characterization of cyclic lipopeptide white-line-inducing principle (WLIP) production by ricerhizosphere isolate Pseudomonas putida RW10S2. Appl. Environ. Microbiol. 78:4826–34
134. Rouse S, Harnett D, Vaughan A, van Sinderen D. 2008. Lactic acid bacteria with potential to eliminatefungal spoilage in foods. J. Appl. Microbiol. 104:915–23
www.annualreviews.org • Molecular Bacteria-Fungi Interactions 395
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
135. Ryan LA, Zannini E, Dal Bello F, Pawlowska A, Koehler P, Arendt EK. 2011. Lactobacillus amylovorusDSM 19280 as a novel food-grade antifungal agent for bakery products. Int. J. Food. Microbiol. 146:276–83
136. Sajben E, Manczinger L, Nagy A, Kredics L, Vagvolgyi C. 2010. Characterization of pseudomonadsisolated from decaying sporocarps of oyster mushroom. Microbiol. Res. 166:255–67
137. Salvioli A, Chiapello M, Fontaine J, Hadj-Sahraoui AL, Grandmougin-Ferjani A, et al. 2010. Endobac-teria affect the metabolic profile of their host Gigaspora margarita, an arbuscular mycorrhizal fungus.Environ. Microbiol. 12:2083–95
138. Salvioli A, Lumini E, Anca IA, Bianciotto V, Bonfante P. 2008. Simultaneous detection and quantificationof the unculturable microbe Candidatus Glomeribacter gigasporarum inside its fungal host Gigasporamargarita. New Phytol. 180:248–57
139. Scherlach K, Busch B, Lackner G, Paszkowski U, Hertweck C. 2012. Symbiotic cooperation in thebiosynthesis of a phytotoxin. Angew. Chem. Int. Ed. 51:9615–18
140. Scherlach K, Hertweck C. 2009. Triggering cryptic natural product biosynthesis in microorganisms.Org. Biomol. Chem. 7:1753–60
141. Scherlach K, Partida-Martinez LP, Dahse HM, Hertweck C. 2006. Antimitotic rhizoxin derivatives froma cultured bacterial endosymbiont of the rice pathogenic fungus Rhizopus microsporus. J. Am. Chem. Soc.128:11529–36
142. Schmitt I, Partida-Martinez LP, Winkler R, Voigt K, Einax E, et al. 2008. Evolution of host resistancein a toxin-producing bacterial-fungal alliance. ISME J. 2:632–41
143. Reveals bacterialcell wall biosynthesis asthe pathway targeted bythe fungal defensinplectasin.
143. Schneider T, Kruse T, Wimmer R, Wiedemann I, Sass V, et al. 2010. Plectasin, a fungal defensin,targets the bacterial cell wall precursor Lipid II. Science 328:1168–72
144. Schoenian I, Spiteller M, Ghaste M, Wirth R, Herz H, Spiteller D. 2011. Chemical basis of the synergismand antagonism in microbial communities in the nests of leaf-cutting ants. Proc. Natl. Acad. Sci. USA108:1955–60
145. Schoonbeek HJ, Raaijmakers JM, De Waard MA. 2002. Fungal ABC transporters and microbial inter-actions in natural environments. Mol. Plant Microbe Interact. 15:1165–72
146. Schroeckh V, Scherlach K, Nutzmann HW, Shelest E, Schmidt-Heck W, et al. 2009. Intimate bacterial-fungal interaction triggers biosynthesis of archetypal polyketides in Aspergillus nidulans. Proc. Natl. Acad.Sci. USA 106:14558–63
147. Scott JJ, Oh DC, Yuceer MC, Klepzig KD, Clardy J, Currie CR. 2008. Bacterial protection of beetle-fungus mutualism. Science 322:63
148. Seipke RF, Barke J, Brearley C, Hill L, Yu DW, et al. 2011. A single Streptomyces symbiont makes multipleantifungals to support the fungus farming ant Acromyrmex octospinosus. PLoS One 6:e22028
149. Demonstrates thatbacterial symbionts ofthe fungus-growing antdisplay a broad andnonspecific antibioticactivity even against thegarden fungus.
149. Sen R, Ishak HD, Estrada D, Dowd SE, Hong E, Mueller UG. 2009. Generalized antifungal ac-tivity and 454-screening of Pseudonocardia and Amycolatopsis bacteria in nests of fungus-growingants. Proc. Natl. Acad. Sci. USA 106:17805–10
150. Shih C, Teicher BA. 2001. Cryptophycins: a novel class of potent antimitotic antitumor depsipeptides.Curr. Pharm. Des. 7:1259–76
151. Soler-Rivas C, Arpin N, Olivier JM, Wichers HJ. 1999. WLIP, a lipodepsipeptide of Pseudomonas ‘re-actans’, as inhibitor of the symptoms of the brown blotch disease of Agaricus bisporus. J. Appl. Microbiol.86:635–41
152. Soler-Rivas C, Jolivet S, Arpin N, Olivier JM, Wichers HJ. 1999. Biochemical and physiological aspectsof brown blotch disease of Agaricus bisporus. FEMS Microbiol. Rev. 23:591–614
153. Strack D, Fester T, Hause B, Schliemann W, Walter MH. 2003. Arbuscular mycorrhiza: biological,chemical, and molecular aspects. J. Chem. Ecol. 29:1955–79
154. Strobel G, Daisy B, Castillo U, Harper J. 2004. Natural products from endophytic microorganisms.J. Nat. Prod. 67:257–68
155. Takahashi M, Iwasaki S, Kobayashi H, Okuda S. 1987. Studies on macrocyclic lactone antibiotics.XI. Anti-mitotic and anti-tubulin activity of new antitumor antibiotics, rhizoxin and its homologues.J. Antibiot. 40:66–72
156. Takahashi M, Matsumoto S, Iwasaki S, Yahara I. 1990. Molecular basis for determining the sensitivityof eucaryotes to the antimitotic drug rhizoxin. Mol. Gen. Genet. 222:169–75
396 Scherlach · Graupner · Hertweck
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67CH18-Hertweck ARI 12 August 2013 11:12
157. Theiss WC, Carmichael WW, Wyman J, Bruner R. 1988. Blood pressure and hepatocellular effects ofthe cyclic heptapeptide toxin produced by the freshwater cyanobacterium (blue-green alga) Microcystisaeruginosa strain PCC-7820. Toxicon 26:603–13
158. Tylka GL, Hussey RS, Roncadori RW. 1991. Axenic germination of vesicular-arbuscular mycorrhizalfungi: effects of selected Streptomyces species. Phytopathology 81:754–59
159. Valerio F, Favilla M, De Bellis P, Sisto A, de Candia S, Lavermicocca P. 2009. Antifungal activity ofstrains of lactic acid bacteria isolated from a semolina ecosystem against Penicillium roqueforti, Aspergillusniger and Endomyces fibuliger contaminating bakery products. Syst. Appl. Microbiol. 32:438–48
160. Vilchez R, Lemme A, Ballhausen B, Thiel V, Schulz S, et al. 2010. Streptococcus mutans inhibits Candida al-bicans hyphal formation by the fatty acid signaling molecule trans-2-decenoic acid (SDSF). ChemBioChem11:1552–62
161. Viljoen BC. 2001. The interaction between yeasts and bacteria in dairy environments. Int. J. Food Mi-crobiol. 69:37–44
162. Wang LH, He Y, Gao Y, Wu JE, Dong YH, et al. 2004. A bacterial cell-cell communication signal withcross-kingdom structural analogues. Mol. Microbiol. 51:903–12
163. Wilson T, Rabie CJ, Fincham JE, Steyn PS, Schipper MA. 1984. Toxicity of rhizonin A, isolated fromRhizopus microsporus, in laboratory animals. Food Chem. Toxicol. 22:275–81
164. Winter JM, Behnken S, Hertweck C. 2011. Genomics-inspired discovery of natural products. Curr.Opin. Chem. Biol. 15:22–31
165. Yang X, Shimizu Y, Steiner JR, Clardy J. 1993. Nostoclide I and II, extracellular metabolites from asymbiotic cyanobacterium, Nostoc sp., from the lichen Peltigera canina. Tetrahedron Lett. 34:761–64
166. Zhao J, Shan T, Mou Y, Zhou L. 2011. Plant-derived bioactive compounds produced by endophyticfungi. Mini Rev. Med. Chem. 11:159–68
167. Zuck KM, Shipley S, Newman DJ. 2011. Induced production of N-formyl alkaloids from Aspergillusfumigatus by co-culture with Streptomyces peucetius. J. Nat. Prod. 74:1653–57
www.annualreviews.org • Molecular Bacteria-Fungi Interactions 397
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67-FrontMatter ARI 12 August 2013 12:26
Annual Review ofMicrobiology
Volume 67, 2013 Contents
Fifty Years Fused to LacJonathan Beckwith � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 1
3′ Cap-Independent Translation Enhancers of Plant VirusesAnne E. Simon and W. Allen Miller � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �21
Acyl-Homoserine Lactone Quorum Sensing: From Evolutionto ApplicationMartin Schuster, D. Joseph Sexton, Stephen P. Diggle, and E. Peter Greenberg � � � � � � � � �43
Mechanisms of Acid Resistance in Escherichia coliUsheer Kanjee and Walid A. Houry � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �65
The Biology of the PmrA/PmrB Two-Component System: The MajorRegulator of Lipopolysaccharide ModificationsH. Deborah Chen and Eduardo A. Groisman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �83
Transcription Regulation at the Core: Similarities Among Bacterial,Archaeal, and Eukaryotic RNA PolymerasesKimberly B. Decker and Deborah M. Hinton � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 113
Bacterial Responses to Reactive Chlorine SpeciesMichael J. Gray, Wei-Yun Wholey, and Ursula Jakob � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 141
It Takes a Village: Ecological and Fitness Impactsof Multipartite MutualismElizabeth A. Hussa and Heidi Goodrich-Blair � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 161
Electrophysiology of BacteriaAnne H. Delcour � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 179
Microbial Contributions to Phosphorus Cycling in Eutrophic Lakesand WastewaterKatherine D. McMahon and Emily K. Read � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 199
Structure and Operation of Bacterial Tripartite PumpsPhilip Hinchliffe, Martyn F. Symmons, Colin Hughes, and Vassilis Koronakis � � � � � � � � � 221
Plasmodium Nesting: Remaking the Erythrocyte from the Inside OutJustin A. Boddey and Alan F. Cowman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 243
vi
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67-FrontMatter ARI 12 August 2013 12:26
The Algal Past and Parasite Present of the ApicoplastGiel G. van Dooren and Boris Striepen � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 271
Hypoxia and Gene Expression in Eukaryotic MicrobesGeraldine Butler � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 291
Wall Teichoic Acids of Gram-Positive BacteriaStephanie Brown, John P. Santa Maria Jr., and Suzanne Walker � � � � � � � � � � � � � � � � � � � � � � 313
Archaeal Biofilms: The Great UnexploredAlvaro Orell, Sabrina Frols, and Sonja-Verena Albers � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 337
An Inquiry into the Molecular Basis of HSV Latency and ReactivationBernard Roizman and Richard J. Whitley � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 355
Molecular Bacteria-Fungi Interactions: Effects on Environment, Food,and MedicineKirstin Scherlach, Katharina Graupner, and Christian Hertweck � � � � � � � � � � � � � � � � � � � � � � � 375
Fusarium PathogenomicsLi-Jun Ma, David M. Geiser, Robert H. Proctor, Alejandro P. Rooney,
Kerry O’Donnell, Frances Trail, Donald M. Gardiner, John M. Manners,and Kemal Kazan � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 399
Biological Consequences and Advantages of AsymmetricBacterial GrowthDavid T. Kysela, Pamela J.B. Brown, Kerwyn Casey Huang, and Yves V. Brun � � � � � � � 417
Archaea in Biogeochemical CyclesPierre Offre, Anja Spang, and Christa Schleper � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 437
Experimental Approaches for Defining Functional Roles of Microbesin the Human GutGautam Dantas, Morten O.A. Sommer, Patrick H. Degnan,
and Andrew L. Goodman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 459
Plant Cell Wall Deconstruction by Ascomycete FungiN. Louise Glass, Monika Schmoll, Jamie H.D. Cate, and Samuel Coradetti � � � � � � � � � � � � 477
Cnidarian-Microbe Interactions and the Origin of InnateImmunity in MetazoansThomas C.G. Bosch � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 499
On the Biological Success of VirusesBrian R. Wasik and Paul E. Turner � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 519
Prions and the Potential Transmissibility of ProteinMisfolding DiseasesAllison Kraus, Bradley R. Groveman, and Byron Caughey � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 543
Contents vii
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.

MI67-FrontMatter ARI 12 August 2013 12:26
The Wonderful World of Archaeal VirusesDavid Prangishvili � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 565
Tip Growth in Filamentous Fungi: A Road Trip to the ApexMeritxell Riquelme � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 587
A Paradigm for Endosymbiotic Life: Cell Differentiation of RhizobiumBacteria Provoked by Host Plant FactorsEva Kondorosi, Peter Mergaert, and Attila Kereszt � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 611
Neutrophils Versus Staphylococcus aureus: A Biological Tug of WarAndras N. Spaan, Bas G.J. Surewaard, Reindert Nijland,
and Jos A.G. van Strijp � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 629
Index
Cumulative Index of Contributing Authors, Volumes 63–67 � � � � � � � � � � � � � � � � � � � � � � � � � � � 651
Errata
An online log of corrections to Annual Review of Microbiology articles may be found athttp://micro.annualreviews.org/
viii Contents
Ann
u. R
ev. M
icro
biol
. 201
3.67
:375
-397
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
idad
e Fe
dera
l de
Sant
a C
atar
ina
on 1
1/08
/13.
For
per
sona
l use
onl
y.