modulation of growth factor responsiveness of murine mammary carcinoma cells by cell matrix...
TRANSCRIPT

JOURNAL OF CELLULAR PHYSIOLOGY 152:292-301 t 1992)
Modulation of Growth Factor Responsiveness of Murine Mammary Carcinoma Cells by Cell
Matrix Interactions: Correlation of Cell Proliferation and Spreading
BRUCE ELLIOTT, ARNE OSTMAN, BENCT WESTERMARK, AND KRISTOFER RUBIN* Dcpartmcnt of Medrcal and Phy3rologrcal Chenirctry, Unrversrty ot Uppsala, S-75 7 23 I lppsala ( H E , K R 1, Ludwrg Instrtute for Cancer Rerearch, S-75 7 24 Uppsala (A 0 1,
Department of Pathology, University Hosprtal, 5-75 7 H i 1Jpptala (6 W ), 5wetlen
We have examined the role of growth factors and extracellular matrix in the proliferation and cell adhesion of a murine mammary carcinoma, SP1, and a stable highly metastatic variant, SPI -3M. O n fibronectin, both cell typcs prolifer- ated strongly in response to basic fibroblast growth factor (bFGF) and platelet- derived growth factor BB (PDGF-BB) after culture for 24 h and 72 h. In contrast, on collagen type I, SP1 cells proliferated only weakly to PDGF-BB at either lime, and SP1-3M cells showed a response to PDGF-BB only at 72 h. The proliferative response to bFGF was also consistently lower when the cells were cultured on collagen than on fibronectin. No significant proliferative responses were detected to epithelial growth factor (EGF), transforming growth factor-p (TGF-p), or estro- gen on any substratum. The lack of responsiveness to PDGF-BB of cells cultured on collagen type I was not due to differences in numbers or affinity of PDGF receptors. We therefore examined the adhesion and spreading properties of SP1 and SPI-3M cells. Without exogenous growth factors, both cell lines adhered to fibronectin and laminin. SP1-3M cells did not bind to collagen type I, whereas SP1 cells did. Attachment to all three substrata was inhibited by anti-p, integrin IgG, suggesting that the primary adhesion to these substrata is mediated by p, integrins. SP1 and SP1-3M cells showed similar integrin pattcrns following immunoprecipi- tation by anti-p, integrin IgG. bFGF stimulated increased adhesion and spreading of both SP1 and SPI-3M cells to collagen type I within 24 h, whereas PDGF-BB was less capable of this effect. Our results suggest that the proliferative response of SP1 and SP1-3M cells to PDCF-BB and bFGF is dependent on the extracellular matrix environment, and imply that modification of extracellular matrix and/or surface integrin receptors may regulate responsiveness to these growth factors in the SP1 tumor model. GI 1992 Wilev-Lis. Inc.
Tissue mesenchyme can dramatically contribute to the selection of tumor subpopulations with increased growth and invasive phenotypes (Price e t al., 1990; Miller and McInerney, 1988). Growth factors and extra- cellular matrix are two important types of stromal products that are likely to be involved in this process. For instance, members of the fibroblast growth factor (FGF) family, which are synthesized and deposited in subendothelial extracellular matrix (Folkman et al., 1988), appear to be important mediators of angiogene- sis and of regulation of growth in certain tumor cells (Wellstein et al., 1990). Production of another growth substance, platelet-derived growth factor (PDGF), has been reported in some malignancies including breast tumors (Rozengurt e t al., 1985; Peres et al., 19871, al- though the function of this growth factor pathway in tumor progression is poorly understood.
Extracellular matrix (ECM) proteins, such as colla- gen types I and IV (Bissell et al., 1982, Salomon et al., 1981) and fibronectin (Brooks et al., 1990; Ruoslahti, 0 1992 WILEY-LISS, INC
19881, can modulate the growth and differentiation of normal and malignant cells. Many receptors for ECM components belong to the integrin family of adhesion receptors (reviewed in Hynes, 1987; Ruoslahti and Pierschbacher, 1987; Albelda and Buck, 1990; Ak- iyama et al., 1990). Integrins are composed of two non- covalently associated subunits, denoted a and p. The integrins have been classified into a t least six subfami- lies, designated PI-& according to the type of P-sub- unit present (Albelda and Buck, 1990; McLean et al., 1990; Sheppard et al., 1990). Receptors for ECM pro-
Received October 17,1991; accepted February 18,1992 *To whom reprint requestsicorrespondence should be addressed. Bruce Elliott’s present address is Cancer Research Laboratories, Department of Pathology, Queen’s University, Kingston, Canada, K7L 3N6.

CELL MATRIX AND GROWTH FACTOR RESPONSE 293
teins including fibronectin, laminin, and collagen are found predominantly within the PI subfamily of inte- grins (Hynes, 1987; Ruoslahti and Pierschbacher, 1987; Albelda and Buck, 1990; Akiyama et al., 1990; Shep- pard et al., 1990). There is much evidence that inte- grins play important roles in mediating mesenchyme- tumor interactions (Giancotti and Ruoslahti, 1990; Dedhar and Saulnier, 1990; Plantefaber and Hynes, 1989). The mechanisms by which interactions among growth factors, ECM proteins, and their integrin recep- tors lead to increased growth and invasiveness of tu- mors are poorly understood.
Previously, Elliott et al. (1988) developed a murine mammary carcinoma (SP1 j cell line which grows faster in the mammary gland than in the subcutis, and metas- tasizes more efficiently from the mammary gland than from the subcutis. A metastatic variant cell line (SP1- 3M) was isolated from a single metastatic nodule which had undergone three cycles of selection from primary SP1 tumors in the mammary gland. SP1-3M cells showed additional chromosome and cell evolution and displayed a stable, more aggressive, metastatic pheno- type in both subcutaneous and intramammary sites (Elliott et al., 1991). These paired metastatic and non- metastatic tumor cell lines present a physiologically relevant model for the study of stromal cell interactions in the development of metastases.
In the present study we examined the role of ECM and growth factors in the proliferation and cell-to-sub- stratum adhesion of SP1 and SPl-3M cells. We ob- served that fibronectin was more capable of supporting an early proliferative response of SP1 and SP1-3M at 24 h to basic FGF (bFGF) and PDGF-BB than collagen type I, although both cell lines were capable of respond- ing to bFGF on collagen type I at 3 days. To assess the role of cell attachment and spreading in the initial stimulation of SP1 cells with growth factors, we exam- ined the effect of bFGF and PDGF-BB on adhesion and spreading of SP1 and SP1-3M cells on collagen type I. Our findings show that bFGF is more capable of stimu- lating a spreading response of SP1 and SP1-3M cells on collagen type I than PDGF-BB, and that the spreading effect correlates with the resulting proliferative re- sponse to these growth factors.
MATERIALS AND METHODS Reagents
The following reagents were purchased from the sup- pliers indicated: collagen type I (Vitrogen 100, Colla- gen Corporation, Palo Alto, CA, USA); fetal calf serum (FCS) (Flow Lab, Irvine, UK); protein-A Sepharose (Pharmacia AB, Uppsala, Sweden); lactoperoxidase, glucose oxidase 11, and pepstatin A (Sigma, St. Louis, MO, USA); leupeptin and bovine serum albumin (BSA) (Boerhringer-Mannheim, Mannheim, FRG); aprotinin (Bayer, Leverkusen, FRG); trypsin-EDTA (GIBCO, Paisley, UK).
Recombinant PDGF-AA and PDGF-BB and trans- forming growth factor-pl (TGF-PI ) were kindly pro- vided by Dr. C.-H. Heldin. Acidic FGF (aFGF), bFGF, and EGF were purchased from Promega (NJ, USA). Human fibronectin was purified from human plasma by affinity chromatography on gelatin-Sepharose, as described in Miekka et al. (1982). Laminin was purified
from EHS tumor extracts as described in Kleinman et al. (1982). A rabbit antiserum to rat a5P1 was kindly donated by Dr. Staffan Johansson (University of Upp- sala, Sweden).
Tissue culture Tumor cells were cultured in RPMI-1640 tissue cul-
ture medium supplemented with transferrin 15 pgiml (Sigma), hydrocortisone 3 x M (Sigma), sodium selenite M (Sigma), glutamine 2 mM (GIBCO), penicillin 0.5 pgiml (GIBCO), and streptomycin 0.6 pgiml (GIBCO). Growth factors or FCS were added as indicates in tables and figures.
Cell lines SP1 was derived from a spontaneous non-metastatic
intraductal mammary adenocarcinoma which arose in a retired female CBAiJ breeder in the Queen’s Univer- sity animal colony. The histological and ultrastructural morphology of SP1 has been described (Carlow et al., 1985).
Metastatic lines were generated from SP1 cells by sequential passage in the mammary gland; this process has been shown to result in metastasis formation (El- liott et al., 1988). Lung metastases were excised; a cell line was established from one nodule, and those cells were reinjected into the mammary glands of a second set of mice. This process was repeated for three serial passages to yield SP1-3M on the third cycle. The cytoge- netic and in vivo growth properties of the cell lines used in this study have been described (Elliott et al., 1992a). The growth and metastatic properties of the cells are stable for a t least three months in culture, and could be recovered from cells frozen for at least two years.
Stock cell lines were maintained in culture medium with 10% FCS without antibiotics and subcultured ev- ery 4-5 days. Cells were detached by treatment (3-5 min) with trypsin (0.05%)/EDTA (0.02%) in Puck’s sa- line (GIBCO) and washed once in RPMI with 10% FCS. For serum-free studies, cells were washed twice in ice- cold medium without serum before being added (2 x lo4 cells per well) directly to 12- or 24-well culture plates (Costar, Cambridge, MA, USA). Proliferation was as- sessed by determining the incorporation of [”]-thymi- dine (T3H]-TdR) into cells following a 12 h pulse with 0.2 pCiiml of L3H]-TdR (Amersham, UK).
Cell attachment The quantitation of cell attachment and the effects of
various antibodies and peptides on this attachment have been described previously (Gullberg et al., 1989). Briefly, a plate-based assay was used in which proteins were coated on 24-well tissue culture plates (Costar) overnight at 4°C in medium A (0.137 M NaCl, 4.7 mM KC1,0.65 mM MgCl,, 1.2 mM CaC12, 10 mM HEPES a t pH 7.4) at the following concentrations: collagen type I, 100 pgiml; fibronectin, 50 pgiml; and laminin, 50 kg/ ml. Remaining protein binding sites on the plate were blocked with 0.1% BSA. Plates were washed three times in PBS and cells (105/well) suspended in medium A were added. In some experiments rabbit anti-p, inte- grin IgG, or pre-immune IgG, or synthetic hexapeptides of the composition GRGDTP or GRGESP (control) were added at the concentrations indicated in figures. Cells

294 ELLIOTT ET AL
TABLE 1. Stimulation of a murine mammary carcinoma cell line SP1 and a metastatic variant SP1-3M with erowth factors (day 3 assay)
3H-thymidine incorporation (cpm X lo-')' SP 1 SP1-3M Conc.
Growth factor' (ndml) Exp. 1 Exp. 2 Exp. 1 Exp. 2
None - 0.5 !C 0.2 0.2 f 0.1 1.4 * 0.6 7.6 -i. 1.9 PDGF-AA3 50 0.5 zk 0.1 0.1 f 0.1 2.5 + 0.4 8.2 5 0.2
bFGF 1 47.9 f 11.22 150.1 f 25' 32.8 f 8.6' 50 5 15.2' EGF3 10 0.6 f 0.2 0.3 + 0.03 1.2 f 0.3 11.9 + 3.2 TGF-p13 10 0.8 zk 0.02 0.1 i 0.01 0.5 + 0.2 4.1 f 0.2 Estrogen3 50 2.3 * 0.1 ND 1.3 k 0.2 ND FCS 1% 157 31.1' 109.1 f 9.22 166 zk 61.7' 86.5 f 17.3'
'SP1 and SPl-3M cells were subcultured at 2 X lo4 cells per culture in 12-well Costar culture plates with various growth factors or estrogen at the concentrations indicated. DNA synthesis was measured at 3 days by adding 0 2 p C 13H]-TdR (Amersham) and harvesting after 24 h. Values show mean cpm X of [3H]-TdR incorporated: mean * s.e.m. of 3 wells. 'Values greater than threefold above background (no growth factor added) were considered positive. 'Values were less than threefold above background at concentrations of this growth factor or hormone hetween 1 and 50 ng per ml.
PDGF-BR 50 1.2 * 1 0.04 f 0.01 13.5 f 5.4' 33.3 * 4.92
were incubated at 37°C for various times as indicated. The plates were washed and the number of cells was quantitated by the hexoseaminidase enzyme assay (Gullberg et al., 1989).
Cell surface labelling and immunoprecipitation For labelling cell surface proteins, cells were cul-
tured in 60 or 100 mm tissue culture plates (Falcon Plastics) until close to confluence. Cells were washed with warm PBS with 5 mM p-glucose and labelled on the plates by the lactoperoxidase method: 0.2 mCi (Amersham), 50 pg lactoperoxidase, and 2 pg glucose oxidase were added to the cells in 1 ml of PBS-glucose and incubated for 20 min a t room temperature. Cells were washed three times in cold PBS, then solubilized for 20 min in 1 ml of lysing buffer (1 mM MgCl,, 10 mM Tris-HC1, and 0.1% Triton X-100, pH 7.4, containing 1 mM phenylmethylsulfonyl fluoride, 1% aprotinin, 1 pg/ml pepstatin, and 23 pgiml leupeptin). Insoluble material was removed by centrifugation a t 14,OOOg in an Ependorff centrifuge. Immunoprecipitation was car- ried out as described previously (Gullberg et al., 1990a).
PDGF receptor binding assay Purified recombinant PDGF-BB and -AA (short
splice variant) (Ostman et al., 1989) were labelled with 1251 as described by Bolton and Hunter (1973) (PDGF- BB), and by the chloramine T method (Hunter and Greenwood, 1962) (PDGF-AA), to specific activities of 16-80 x lo3 cpming.
Binding experiments were performed in triplicate on SP1 and SP1-3M cells grown in 24-well tissue culture plates (50-130 x lo3 cellsiwell). To study specifically the numbers and binding affinities of PDGF P-recep- tors, we preincubated cells a t 37°C for 60 min in 0.2 ml DMEM supplemented with 0.1 mgiml BSA and 100 ng/ml of PDGF-AA to down-regulate surface expression of a-receptors. Cells were then rinsed once in binding buffer (PBS containing 1 mg/ml of BSA, 0.9 mM CaCl,, 0.49 mM MgC1,) and incubated for 2 h at 0°C in 0.2 ml of binding buffer containing 1-2 ng of radiolabelled ligand and various concentrations of unlabelled ligand. Cultures were then washed five times in binding buffer. Cell-associated radioactivity was then extracted by 30 min incubation a t room temperature in 1% Triton
X-100, 10% glycerol, and 20 mM Tris-HC1, pH 7.5, and quantified with a y-counter. Binding data were sub- jected to Scatchard analysis (Scatchard, 1949).
RESULTS Growth factor response pattern of a murine mammary carcinoma SPl and a metastatic
variant SP1-3M As a first step in studying the growth behavior of SP1
and SP1-3M cells in vitro, we compared the responsive- ness of these two cell lines to various growth factors. To exclude the involvement of serum components, we es- tablished serum-free medium conditions which allow optimum survival of tumor cells in 24-72 h cultures and which show low backgrounds of ['HI-TdR incorpo- ration. The optimum medium included selenium, hy- drocortisone, and transferrin. This medium was capa- ble of maintaining two- to fivefold greater proliferation of SP1 cells in response to a 0.1% FCS stimulus com- pared to RPMI without these additives (data not shown).
Table 1 shows the response pattern of SP1 and SP1-3M cells to stimulation for three days with various growth factors on plastic. A proliferative response to bFGF by both SP1 and SP1-3M cells occurred. A mar- ginal proliferative response to aFGF occurred; this re- sponse was strongly facilitated by the addition of hep- arin (1 Fgiml) (data not shown). SP1-3M cells, but not SP1 cells, responded to PDGF-BB. No proliferative re- sponses to PDGF-AA, EGF, TGF-P1, or estrogen were observed. The proliferative response to 1% FCS is shown for comparison. These results show that SPl-3M cells were more capable than SP1 cells of responding to PDGF-BB in a three-day proliferation assay on plastic.
Specificity of adhesion of SP1 and SPl-3M cells to collagen type I, fibronectin, and laminin, and
analysis of receptors To investigate the role of ECM in SP1 cell growth, we
first examined the capacity of SP1 and SP1-3M cells to adhere to substrata consisting of various ECM proteins.
The results in Figure 1 show that only 3 0 4 0 % of SP1 cells attach to native or heat-denatured (56°C for 30 min) collagen type I substrata. No attachment of S P l S M cells to collagen type I was observed. Equiva-

CELL MATRIX AND GROWTH FACTOR RESPONSE 295
2
2 120 a 4 100
v) W
4
0
80
60
40
20
0
COLl E
TIME (MIN)
FN F
LN C
0 100
LN G
ANTI-p1 IgG (ug I ml x
Fig. 1. Adhesion of SP1 and SPI-3M cells to collagen type I, fibronec- tin, and laminin. Cells were harvested by brief trypsinization and added (10” cells per well) to 24-well Costar plates precoated with proteins as indicated. In panels A-D, cells were incubated for the indicated time period in medium A; in panels E-H, anti-p, integrin IgG (at the concentrations indicated) was mixed with the cells and preincubated for 30 min in tubes on ice, transferred to wells coated with matrix proteins, and incubated for a further 40 min a t 37°C. A,E: Collagen type I. B , F Human plasma fibronectin. C,G EHS laminin.
lent proportions (90-100%) of SP1 and SP1-3M cells adhered to fibronectin and to laminin. Attachment to fibronectin was partially inhibited by RGD-containing peptides (Fig. 2) ; this finding suggests that the attach- ment to fibronectin is in part mediated by RGD-di- rected receptors.
The attachment of SP1 cells to collagen type I, fi- bronectin, and laminin, as well as that of SP1-3M cells to fibronectin and laminin, was inhibited by monospe- cific anti-p, integrin IgG (Fig. 1). Thus, PI integrins are
D Anti-P1
100
H Anti-P1
D,H: Anti+, integrin IgG. Open symbols (0 A 0) represent SP1-3M cells; closed symbols (0 A m) represent SPl cells. Squares in A show attachment to heat-denatured collagen (preincubated at 56°C for 30 min). Triangles in B show attachment to BSA. Cell number was mea- sured by the hexoseaminidase enzyme assay and results are expressed as the mean of duplicate points ? range. In panels A-D, values are expressed as a o/o of the maximum attachment to anti+, integrin IgG (100%)). In panels C H , values are expressed as a % of attachment in the absence of anti-& integrin IgG.
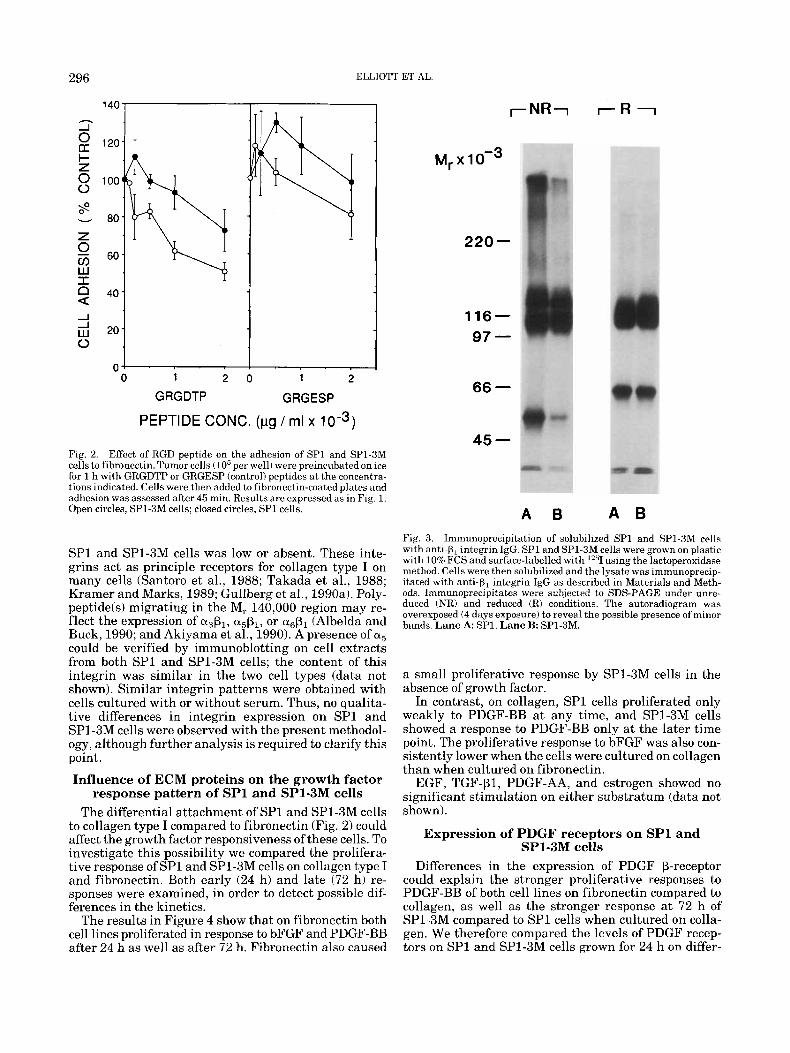
important for the interactions of the tumor cells with these ECM-components. We therefore determined the expression of various PI integrins on SPl and SP1-3M cells by immunoprecipitation with anti+, IgG of ex- tracts from surface-labelled cells (Fig. 3). In extracts from both cell types, a M, 115,000 band at the position expected for the P1-chain was detected (Fig. 3). An ab- sence of prominent bands in the M, range of 150,000- 200,000 in gels run under reduced conditions indicates that the surface expression of alp1 and azP1 by both
296
140
2 2 120 I- z 0 100 0
8 - 80 z 0 cn 6o w I 2 40
J 20
0
n
ELLIOTT ET AL
- 0 1 2 0 1 2
GRGDTP GRGESP
PEPTIDE CONC. (pg / mt x 10-3)
Fig. 2. Effect of RGD peptide on the adhesion of SP1 and SP1-3M cells to fibronectin. Tumor cells (lo5 per well) were preincubated on ice for 1 h with GRGDTP or GRGESP (control) peptides a t the concentra- tions indicated. Cells were then added to fibronectin-coated plates arid adhesion was assessed after 45 min. Results are expressed as in Fig. 1. Open circles, SP1-3M cells; closed circles, SP l cells.
SP1 and SP1-3M cells was low or absent. These inte- grins act as principle receptors for collagen type I on many cells (Santoro et al., 1988; Takada et al., 1988; Kramer and Marks, 1989; Gullberg et al., 1990a). Poly- peptideb) migrating in the M, 140,000 region may re- flect the expression of a&, a&, or a& (Albelda and Buck, 1990; and Akiyama et al., 1990). A presence of a5 could be verified by immunoblotting on cell extracts from both SP1 and SP1-3M cells; the content of this integrin was similar in the two cell types (data not shown). Similar integrin patterns were obtained with cells cultured with or without serum. Thus, no qualita- tive differences in integrin expression on SP1 and SP1-3M cells were observed with the present methodol- ogy, although further analysis is required to clarify this point.
Influence of ECM proteins on the growth factor response pattern of SP1 and S P l S M cells
The differential attachment of SP1 and SP1-3M cells to collagen type I compared to fibronectin (Fig. 2 ) could affect the growth factor responsiveness of these cells. To investigate this possibility we compared the prolifera- tive response of SP1 and SP1-3M cells on collagen type I and fibronectin. Both early (24 h) and late (72 h) re- sponses were examined, in order to detect possible dif- ferences in the kinetics.
The results in Figure 4 show that on fibronectin both cell lines proliferated in response to bFGF and PDGF-BB after 24 h as well as after 72 h. Fibronectin also caused
220-
16- 97 -
66 -
45 -
A 6 A B Fig. 3. Immunoprecipitation of solubilized S P l and SP1-3M cells with anti-p, integrin IgG. SP1 and SP1-3M cells were grown on plastic with 10% FCS and surface-labelled with '''1 using the lactoperoxidase method. Cells were then solubilized and the lysate was immunoprecip- itated with anti-p, integrin IgG as described in Materials and Meth- ods. Imrnunoprecipitates were subjected to SDS-PAGE under unre- duced (NR) and reduced (R) conditions. The autoradiogram was overexposed (4 days exposure) to reveal the possible presence of minor bands. Lane A SP1. Lane B: SP1-3M.
a small proliferative response by SP1-3M cells in the absence of growth factor.
In contrast, on collagen, SPl cells proliferated only weakly to PIIGF-BB a t any time, and SP1-3M cells showed a response to PDGF-BB only a t the later time point. The proliferative response to bFGF was also con- sistently lower when the cells were cultured on collagen than when cultured on fibronectin.
EGF, TGF-P1, PDGF-AA, and estrogen showed no significant stimulation on either substratum (data not shown).
Expression of PDGF receptors on SP1 and SP1-3M cells
Differences in the expression of PDGF p-receptor could explain the stronger proliferative responses to PDGF-BB of both cell lines on fibronectin compared to collagen, as well as the stronger response at 72 h of SPl-3M compared to SP1 cells when cultured on colla- gen. We therefore compared the levels of PDGF recep- tors on SP1 and SP1-3M cells grown for 24 h on differ-

CELL MATRIX AND GROWTH FACTOR RESPONSE 297
SPI /24 H 4 ,
3
h
P 2
2 1 a Y z 0
0
X 7
s? - BB FGF - BB FGF k M SP1-3M/24 H g 4 M 0 O 3 z n k 2
CT
I m 1
0 - BB FGF - BB FGF
Fig. 4. Response of SP1 and SPI-3M cells to PDGF-BB and bFGF on collagen type I or fibronectin substratum. Cells (2 x lo4 per well) were cultured in serum-free medium alone, or with bFGF (1 ng/ml), or with PDGF-BB (10 ngiml) in 24-well Costar plates precoated with collagen type I(100 p,g/ml) or fibronectin (50 p,g/ml). At 12 hand 60 h,
TABLE 2. Expression of PDGF preceptors on SP1 and SP1-3M cells during growth on collagen type I, fibronectin, and laminin'
Matrix SP1 SP1-3M component K D No. receptors K D No. receptors
Collagen type I 0.4 34 0.5 78 Fib r o n ec tin 1.1 62 0.8 75 Laminin 0.8 56 0.4 43
'Cells (lo5 per well) were cultured in 24-well Costar tissue culture plates precoated withcollagen typeI(lOOpg/mI) orfibronectin(50pg/mI) or larninin(50gg/ml). For the PDGFR assay, plates were washed and the amo~ntof'~~1-PDGF-BB bound(cpm) per number of cells plated was determined in each group. Scatchard analysis was performed as previously described (Ostman et al., 1989).
ent substrata; this time corresponded to the early time point in Figure 4. When I2'1-PDGF-BB was used as a ligand, we found that the number and affinity of the receptors were not significantly different on cells of SP1 and SPlSM during growth on collagen type I, fibronec- tin, or laminin (Table 2). Only trace levels of lZ5I- PDGF-AA bound to any of the cells, suggesting that expression of PDGF a-receptors was low or absent. To- gether these results suggest that the PDGF receptors expressed on both cell lines are predominantly of the p type and that neither receptor density nor affinity con- tributes to the altered growth properties of SP1 and SP1-3M cells on fibronectin and collagen type I.
SPI /72 H 8
6
4
2
0
- BB FGF - BB FGF
- BB FGF - BB FGF
SPI-3M/72 H T
cultures were pulsed with [jHI-TdR (final concentration 0.2 pCiwel1) for an additional 12 h, and the number of cpm per well was deter- mined. Results are expressed as the mean of triplicates & SD (at 24 h), or the mean of duplicates t range (at 72 h).
Spreading by SP1 and SP1-3M cells in response to bFGF and PDGF-BB on collagen type I
Since SP1 and SP1-3M cells adhere strongly to fi- bronectin but weakly, or not at all, to collagen type I, we determined whether the proliferative response of SP1 and SP1-3M cells to bFGF on collagen type I corre- lated with increased adhesion and spreading. In the absence of growth factor, the morphology of SP1 cells growing on collagen type I was markedly different from that of SP1-3M cells (Fig. 5). A significant proportion (3040%) of SP1 cells adhered to collagen type I within 6 h of culture. No adhesion or spreading of SP1-3M cells on collagen type I was observed. In contrast, all SP1 and SP1-3M cells adhered and spread on fibronectin within 6 h (Fig. 5). The same morphology of SP1 and SP1-3M cells cultured on collagen type I was also ob- served at 12 h (Fig. 6A,G). By 24 h of culture on colla- gen type I, SP1-3M cells began to grow in tight three- dimensional spheres (Fig. 6J). SP1 cells also formed some aggregates of cells (Fig. 6D): however, the cells in the aggregates did not appear to be bound so tightly as in SP1-3M (Fig. 6J).
In the presence of bFGF the proportion of both SP1 and SP1-3M cells spreading on collagen type I in- creased markedly by 24 h (Fig. 6E,K); no significant effect was seen at 12 h (Fig. 6B,H). This spreading effect was dependent on the concentration of bFGF (data not shown). Spreading of SP1-3M cells on collagen

298 ELLIOTT ET AL.
COL I FN
SPI
SPI-3M
Fig. 5. Morphology at 6 h of SP1 and SP1-3M cells growing on collagen type I and fibronectin without growth factor added. SP1 and SP1-YM cells were added to Costar plates prepared as described in Fig. 4. Photographs were taken after 6 h of culture. A SP1 on collagen type I. B: SP1 on fibronectin. C: SPJ -3M on collagen type I. D: SP1-3M on fibronectin. Magnification is x 150.
type I in response to bFGF appeared to originate from cell spheres (Fig. 6K). PDGF-BB also produced an in- crease in the proportion of cells spreading at 24 h (Fig. 6F,L), although the effect was not as pronounced as with bFGF.
DISCUSSION In the present report we have compared the cell-to-
substratum adhesion and proliferative responses to PDGF-BB and bFGF of a murine mammary carcinoma (SP1) cell line and a stable metastatic variant (SP1- 3M). SP1 cells adhered weakly, and SP1-3M cells not a t all, to collagen type I; both cells adhered strongly to fibronectin and laminin. Fibronectin, but not collagen type I, promoted an early (24 h) proliferative response of SP1 and SP1-3M cells to PDGF-BB; this effect was not due to differences in PDGF @-receptor density or affinity. Similarly, fibronectin was more efficient than collagen type I in promoting an early (24 h) prolifera- tive response to bFGF. Both growth factors were capa- ble of inducing increased spreading of SP1 and SPl-3M cells after 24 h of culture on collagen type I. The spread- ing response of SPl and SP1-3M cells to bFGF on colla- gen was followed by a strong proliferative response de- tectable after 72 h.
The pioneering work of Folkman and Moscona (1978) demonstrated the importance of cell spreading in cell proliferation. By varying the adhesion of cells to tissue
culture plastic, these investigators were able t o show that the degree of cell spreading correlated with DNA synthesis induced by serum. Ingber (1990) showed that the mitogenic effect of bFGF depends on cell spreading, which in turn depends on the amount of fibronectin on the substratum. The present results are in agreement with these findings; we found that cell adhesion and spreading were closely correlated with the early (24 h) proliferative response of both SP1 and SP1-3M cells to bFGF and PDGF-BB. Thus, fibronectin promoted strong adhesion and early proliferative responses to bFGF and PDGF-BB, whereas collagen type I did not.
bFGF induced a spreading of both SP1 and SP1-3M cells visible at 24 h on collagen type I. This was followed by a strong proliferative response of both cell types a t 72 h. PDGF-BB also induced some spreading of SP1 and SP1-3M at 24 h on collagen. However, SP1 cells showed only a small proliferative response to PDGF-BB on this substratum at 72 h; a stronger proliferative response of SP1-3M cells to PDGF-BB on collagen was observed at 72 h. This observation suggests that SPI-3M cells may be less restricted by ECM than SP1 cells in their capac- ity to proliferate in response to PIIGF-RB. SP1-3M cells may be capable of remodelling their own matrix envi- ronment (e.g., synthesis of fibronectin or of proteinases that break down collagen type I), or may express func- tionally different integrin receptors compared to SP1 cells.

SPI
CELL MATRIX AND GROWTII FACTOR RESPONSE
NO GF bFGF PDGF-BB
12h
24h
SPI-3M
12h
24h
Fig. 6. Influence of bFGF and PDGF-BB on the morphology of SP1 and SP1-3M cells growing on collagen type I for 12 and 24 h. SP1 (A-F) and SP1-3M (G-L) cells from thc experiment shown in Fig. 4 were photographed at 12 h and 24 h of culture, corresponding to the beginning and the end of the first pulse with ['HI-TdR. First column (A,D,G,J), no growth factor present (NO GF); second column (B,E,H,K), bFGF i l ngiml) added; third column (C,F,I,L), PDGF-BB (10 ng/ml) added.
299

300 ELLIOTT ET AL.
Taken together, our results clearly show that adhe- sion of SP1 and SP1-3M carcinoma cells is not only affected by the nature of the solid support but is also modulated in response to defined growth factors. This finding suggests the presence of a two-stage mecha- nism of growth factor-regulated mitogenesis: the growth factor stimulates cell spreading and thereby augments the mitogenic response.
The different abilities of SP1 and SP1-3M cells to adhere to collagen type I could not be attributed to differences in expression of p1 integrins. Both cell types lacked detectable expression of alp1 and a2p1, two inte- grins that have been ascribed roles as collagen type I receptors on human and rat cells (Santoro et al., 1988; Takada et al., 1988; Kramer and Marks, 1989; Gullberg et al., 1990a). a3P1 and avPl have also been reported to act as collagen receptors (Takada et al., 1988; Albelda and Buck, 1990; Akiyama et al., 1990; Dedhar and Gray, 19901, and it is possible that one or both of these integrins mediates the attachment of SP1 cells to colla- gen. SP1-3M cells should then lack these integrins, or express variant forms not binding to collagen. The in- crease in spreading of cells on collagen type I induced by bFGF or PDGF-BB could be mediated by an in- creased production of fibronectin; our results clearly show that fibronectin efficiently promotes spreading and proliferation of SP1 cells. In a previous study we demonstrated that anti-p, integrin IgG could inhibit PDGF-stimulated collagen gel contraction (Gullberg et al., 1990b). The effect of the IgG could be overcome by increasing the dose of PDGF, a finding which suggests that PDGF may increase the activity of integrins. By analogy it is possible to hypothesize that the effect of PDGF-BB on cell spreading reported here may also be due to altered activity or expression of collagen-binding integrins. Other changes, including altered cytoskele- ton organization, may also be important.
The mouse mammary carcinoma cells used in the present study showed an unexpected response pattern to EGF and estrogen. Neither growth substance had an effect in this system, in contrast to many human mam- mary carcinoma cell lines which show EGF-dependent growth (Dickson and Lippman, 1987). It has recently been shown that estrogen can promote growth of the SP1 and SP1-3M cells in ovariectomized animals (El- liott et al., 1992); however, this effect may be mediated indirectly via stromal cells.
The mitogenic response of SP1 and SP1-3M cells to bFGF is consistent with other reports that this growth factor can stimulate proliferation in many human breast carcinoma cell lines (Dickson and Lippman, 1987), and may be an important regulator of neovascu- lar formation, a crucial step in solid-tumor develop- ment (Kandel et al., 1991). There are fewer studies on the role of PDGF-BB in breast cancer, since previous studies have shown that human mammary carcinoma cells secrete PDGF but have no PDGF receptors (Rozen- gurt et al., 1985; Peres et a]., 1987). The present results clearly demonstrate the presence of functional PDGF p-receptors on both SP1 and SPl-3M cells which are of mouse mammary origin (Elliott et al., 1988). Similar observations have recently been reported for both hu- man (Ginsburg and Vonderhaar, 1991) and mouse (Taverna et al., 1991) mammary carcinoma cells.
PDGF may therefore have a broader role in the growth of mammary carcinomas than was previously thought. Both stromal cells and carcinoma cells themselves may respond to locally produced bFGF and PDGF. Experi- ments are in progress to examine the role of these growth factors in the in vivo growth and metastasis of SP1 cells.
In summary, we have shown that extracellular ma- trix proteins can modulate the proliferative and spread- ing responsiveness of the SP1 mammary carcinoma cell line to PDGF-BB and bFGF. Fibronectin promotes a proliferative response more efficiently than collagen type I. Spreading of cells on the matrix substratum appears to be very closely correlated with the prolifera- tive response. This model provides a useful system to analyze the intracellular pathways that are activated by the PDGF and FGF receptor tyrosine kinases, and the role of signals from the extracellular matrix in pro- moting the growth, competence, and metastasis of tu- mor cells.
ACKNOWLEDGMENTS The skillful technical assistance of Ms. Karina dhlen
and Ms. Ann-Marie Gustafsson is gratefully acknowl- edged. This work was supported by grants from the Swedish Cancer Society, Konung Gustaf V:s 80-Brs- fond, The Lundberg Foundation, and Nordisk Insulin- fond. B.E. is a Terry Fox Scientist of the National Can- cer Institute of Canada and was a recipient of an Eleanor Roosevelt Exchange Fellowship from the Inter- national Union Against Cancer.
LITERATURE CITED Akiyama, S.M., Nagata, K., and Yamada, K.M. (1990) Cell surface
receptors for extracellular matrix components. Biochem. Biophys. Acta, 1031:91-110.
Albelda, S.M., and Buck, C.A. (1990) Integrins and other cell adhesion molecules. FASEB J., 4:286%2880.
Bissell, M.J., Hall, H.G., and Parry, G. (1982) How does the extracellu- lar matrix direct gene expression? J. Theor. Biol., 99:3168.
Bolton, A.E., and Hunter, W.M. (1973) Thc labeling of proteins to high specific radioactivities by conjugation to a L251-containing alcylating reagent. Biochem. J. , 133529-539.
Brooks, R.F., Howard, M., Leake, D.S., and Riddle, P.N. (1990) Failure of platelet-derived growth factor plus insulin to stimulate sustained proliferation of Swiss 3T3 cells. Requirement for hydrocortisone, prostaglandin El, lipoproteins, fibronectin and an unidentified com- ponent derived from serum. J . Cell. Sci., 97t71-78.
Carlow, D.A., Kerbel, R.S., Feltis, J.T., and Elliott, B.E. (1985) En- hanced expression of class I MHC (Dk) gene products on immuno- genic variants of a spontaneous murine carcinoma. J. Natl. Cancer Inst., 75:291-303.
Dedhar, S., and Gray, V. (1990) Isolation of a novel integrin receptor mediating Arg-Gly-Asp-directed cell adhesion to fibronectin and type I collagen from neuroblastoma cells. Association of a novel p,-related subunit with 01". J. Cell Biol., 110t2185-2193.
Dedhar, S., and Saulnier, R. (1990) Alterations in integrin receptor expression on chemically transformed human cells: Specific en- hancement of laminin and collagen receptor complexes. J. Cell Biol., 1 lot48 1 4 8 9 .
Dickson, R.B., and Lippman, M.E. (1987) Estrogenic regulation of growth and polypeptide growth factor secretion in human breast cancer. Endocr. Res., 8:2943.
Elliott, B.E., Maxwell, L., Arnold, M., Wei, W.Z., and Miller, F.R. (1988) Expression of epithelial-like markers and class I major histo- compatibility antigens by a murine carcinoma growing in the mam- mary gland and in metastases: Orthotropic site eff'ects. Cancer Res., 4Nr7237-7245.
Elliott, B.E., Xu, W., Mudrik, K., Marshall, J , , Vekmans, J., and Holden, J.J.A. (1992a) Karyotypic evolution of a murine mammary adenocarcinoma in vitro and during progression from primary t o metastatic growth in vivo. Genes Chromosom Cancer, 4:l-9.

301 CELL MATRIX AND GROWTH FACTOR RESPONSE
Elliott, B.E., Tam, S.-P., Dexter, D., and Chen, Z.Q. (1992b3 Capacity of adipose tissue to promote growth and metastasis of a murine mammary carcinoma: Effect of estrogen and progesterone. Int. 3. Cancer, 52t1-9.
Folkman, J., and Moscona, A. (1978) Role of cell shape in growth control. Nature, 273.345-349.
Folkman, J., Klagsbrun, M., Sasse, J., Wadzinski, M., Ingber, D., and Vlodavsky, I. (19881 A heparin-binding angiogenic protein-basic fibroblast growth factor-is stored within basement membrane. Am. J. Pathol., 130t393-400.
Giancotti, F., and Ruoslahti, E. (1990) Elevated levels of the aspl fibronectin receptor suppress the transformed phenotype of Chinese hamster ovary cells. Cell, 60t849--859.
Ginsburg, E., and Vonderhaar, B.K. (1991) Stimulation of growth of human breast cancer cells (T47Dl by platelet-derived growth factor. Cancer Lett., 58t137-144.
Gullberg, D., Terracio, L., Borg, T.K., and Rubin, K . (1989) Integrin- like collagen receptors on primary cells: Identification of integrins on hepatocytes and fibroblasts with affinity for interstitial colla- gens. J. Biol. Chem., 264t12686-12694.
Gullberg, D., Turner, D.C., Borg, T.K., Terracio, L., and Rubin, K. (1990a) Different P,-integrin collagen receptors on rat hepatocytes and cardiac fibroblasts. Exp. Cell Res., 190:254-264.
Gullberg, D., Tingstrom, A., Thuresson, A.-C., Olsson, L., Terracio, L., Borg, T.K., and Rubin, K. (1990b) (3, -1ntegrin-mediated collagen gel contraction is stimulated by PDGF. Exp. Cell Res., 186264-272.
Hunter, W.M., and Greenwood, F.C. (1962) Preparation of iodine-131 labelled human growth hormone of high specific activity. Nature, 194:496-496.
Hynes, R.O. (1987) Integrins: A family of cell surface receptors. Cell, 48t549-554.
Ingber, D.E. (1990) Control of fibroblast growth factor action by extra- cellular matrix. In: Growth Factors in Health and Disease. B. Wes- termark, C. Betsholtz, and B. Hokfelt, eds. Elsevier Sci. Pub. (Am- sterdam), pp. 289-302.
Kandel, J., Bossy-Wetzel, E., Radvanyl, F., Klagsbrun, M., Folkman, J., and Hanahan, D. (1991) Neovascularization is associated with a switch to the export of bFGF in the multistep development of fibro- sarcoma. Cell, 66t1095-1104.
Kleinman, H.K., McGarvey, M.L., Liotta, L.A., Gehron Robey. P., Tryggvason, K., and Martin, G.R. (1982) Isolation and characteriza- tion of type IV procollagen, laminin, and heparan sulfate proteogly- can from the EHS sarcoma. Biochemistry, 21r6188-6193.
Kramer, R.H., and Marks, N. (1989) Identification of integrin collagen receptors on human melanoma cells. J. Biol. Chem., 264:46844688.
McLean, J.W., Vestal, D.J., Cheresh, D.A., and Bodary, S.C. (1990) cDNA sequence of the human integrin ps subunit. J . Biol. Chem., 265: 17 I 26-17 13 1.
McLean, J.W., Vestal, D.J., Cheresh, D.A., and Bodary, S.C. (1990) cDNA sequence of the human integrin pS subunit. J. Biol. Chem., 265:1772&17131.
Miekka, S.I., Ingham, K.C., and Menache, D. (1982) Rapid methods for isolation of human plasma fibronectin. Thromb. Res., 27:l-14.
Miller, F.R., and McInerney, D. (19881 Epithelial component of host- tumor interactions in the orthotopic site of preference of a mouse mammary tumor. Cancer Kes., 48r3698-3701.
Ostman, A., Backstrom, G., Fong, N., Betsholtz, C., Wernstedt, C., Hellman, U., Westermark, B., Valenzuela, P. and Heldin, C.-H. (1989) Expression of three recombinant homodimcric isoforms of PIIGF in Saccharomyces cereuisiae: Evidence for differences in re- ceptor binding and functional activities. Growth Factors, 1 t271- 281.
Peres, R., Betsholtz, C., Westermark, B., and Heldin, C.-H. (1987) Frequent expression of growth factors for mesenchymal cells in hu- man mammary carcinoma cell lines. Cancer Res., 47,3425-3429.
Plantefaber, L.C., and Hynes, R.O. 11989) Changes in integrin recep- tors on oncogenically transformed cells. Cell, 56281-290.
Price, J.E., Polyzos, A., Zbang, R.D., and Daniels, L.M. (1990) Tumor- igenicity and metastasis of human breast carcinoma cell lines in nude mice. Cancer Res.. 50t717-721.
Rozengurt, E., Sinnett-Smith, J., and Taylor-Papadimitriou, J. (1985) Production of PDGF-like growth factor by breast cancer cells lines. Int. J. Cancer, 36:247-252.
Ruoslahti, E. (1988) Fibronectin and its receptors. Annu. Rev. Bio- chem., 57t375-413.
Ruoslahti, E., and Pierschbacher, M. (1987) New perspectives in cell adhesion: RGD and integrins. Science, 238t491-497.
Salomon, D.S., Liotta, L.A., and Kidwell, W.R. (1981) Differential response to growth factor by rat mammary epithelium plated on different collagen substrata in serum-free medium. Proc. Natl. Acad. Sci. U.S.A., 78.382-386.
Santoro, S.A., Rajpara: S.M., Staatz, W.D., and Woods, V.L., J r . (1988) Isolation and characterization of a platelet surface collagen binding complex related to VLA-2. Biochem. Biophys. Res. Commun., 153.2 17-2 23.
Scatcbard, G. (1949) The attractions of proteins for small molecules and ions. Ann. N.Y. Acad. Sci., 51r660-672.
Sheppard, D., Rozzo, C., Starr, L., Quaranta, V., Erle, D.J., and Pytela, R. (1990) Complete amino acid sequence of a novel integrin P-sub- unit ipJ identified in epithelial cells using the polymerase chain reaction. J. Bid. Chem., 265t11502-11507.
Takada, Y., Wayner, E.A., Carter, W.G., and Hemler, M.E. (1988) Extracellular matrix receptors? ECMRII and ECMRI, for collagen and fibronectin correspond to VLA-2 and VLA-3 in the VLA family of heterodimers. J . Cell. Biochem., 37t385-393.
Taverna, D., Groner, B., and Hynes, N. (1991) Epidermal growth fac- tor receptor, platelet-derived growth factor receptor, and c-erbB-2 receptor activation all promote growth but have distinctive effects upon mouse mammary epithelial cell differentiation. Cell Growth Differ., 2r145-154.
Wellstein, A., Lupu, R., Zugmaier, G., Flamm, S.L., Cheville, A.L., Delli Bovi, P., Basilico, C., Lippman, M.E., and Kern, F.G. (1990) Autocrine growth stimulation by secreted Kaposi fibroblast growth factor but not by endogenous basic fibroblast growth factor. Cell Growth Differ.. 1:63-71.