mixtures of hemoglobin-based oxygen carriers and perfluorocarbons exhibit a synergistic effect in...
TRANSCRIPT

ARTICLE
Mixtures of Hemoglobin-Based Oxygen Carriersand Perfluorocarbons Exhibit a Synergistic Effect inOxygenating Hepatic Hollow Fiber Bioreactors
Guo Chen, Andre F. Palmer
231A Koffolt Laboratories, William G. Lowrie Department of Chemical and Biomolecular
Engineering, The Ohio State University, 140 West 19th Avenue, Columbus, Ohio 43210;
telephone: 614-292-6033; fax: 614-292-3769; e-mail: [email protected]
Received 3 August 2009; revision received 25 September 2009; accepted 28 September 2009
Published online 8 October 2009 in Wiley InterScience (www.interscience.wiley.com)
. DOI 10.1002/bit.22571ABSTRACT: Hepatic hollow fiber (HF) bioreactors arebeing developed for use as bioartificial liver assist devices(BLADs). In general, BLADs suffer from O2 limited trans-port, which reduces their performance. This modeling studyseeks to investigate if O2 carrying solutions consisting ofmixtures of hemoglobin-based oxygen carriers (HBOCs)and perfluorocarbons (PFCs) can enhance O2 transportto hepatocytes cultured in the extra capillary space (ECS)of HF bioreactors. We simulated supplementing the circu-lating cell culture media stream of the HF bioreactor with amixture containing these two types of oxygen carriers(HBOCs and PFCs). A mathematical model was developedbased on the dimensions and physical characteristics of acommercial HF bioreactor. The resulting set of partialdifferential equations, which describes fluid transport; aswell as, mass transport of dissolved O2 in the pseudo-homogeneous PFC/water phase and oxygenated HBOC,was solved to yield the O2 concentration field in the threeHF domains (lumen, membrane and ECS). Our resultsshow that mixtures of HBOC and PFC display a synergisticeffect in oxygenating the ECS. Therefore, the presence ofboth HBOC and PFC in the circulating cell culture mediadramatically improves transport of O2 to cultured hepato-cytes. Moreover, the in vivo O2 spectrum in a liver sinusoidcan be recapitulated by supplementing the HF bioreactorwith a mixture of HBOCs and PFCs at an inlet pO2 of80mmHg. Therefore, we expect that PFC-based oxygencarriers will be more efficient at transporting O2 athigher O2 levels (e.g., at an inlet pO2 of 760mmHg, whichcorresponds to pure O2 in equilibrium with aqueous cellculture media at 1 atm).
Biotechnol. Bioeng. 2010;105: 534–542.
� 2009 Wiley Periodicals, Inc.
KEYWORDS: hemoglobin-based oxygen carrier; perfluoro-carbon; hollow fiber bioreactor; bioartificial liver assistdevice; oxygen transport; mathematical model
Correspondence to: A.F. Palmer
534 Biotechnology and Bioengineering, Vol. 105, No. 3, February 15, 2010
Introduction
O2 is transport limited during the large-scale culture ofmammalian cells in bioreactors, primarily due to itspoor solubility in aqueous cell culture media (Centis andVermette, 2009). Hepatic hollow fiber (HF) bioreactorsusually suffer from this transport limitation (Hay et al.,2000). These devices are composed of bundles of individualHFs that are seeded with hepatocytes in the extra capillaryspace (ECS) in order to recapitulate the structure of theliver sinusoid (Chan et al., 2004; van de Kerkhove et al.,2004). It is hypothesized that HF bioreactors can be used asa bioartificial liver assist device (BLAD) to bridgepatients suffering from acute liver failure (ALF) towardsorthotopic liver transplantation (Millis and Losanoff, 2005).Hepatocytes cultured in HF bioreactors are situated in a 3-Denvironment, which is physically separated from the cellculture media/patient plasma by the HF membrane whichacts as another barrier that limits O2 transport.
In vivo, it is argued that the local O2 concentration alongthe length of the sinusoid controls the liver’s variedmetabolic and synthetic functions (Kietzmann et al.,2006). Metabolic zonation is found in the liver sinusoidwith respect to gradients of key liver enzymes, translocators,and receptors that vary along the length of the liver sinusoidfrom the inlet (portal tract) to the outlet (central vein;Jungermann and Kietzmann, 1996). Variation in theconcentration of key enzymes along the length of theliver sinusoid, which includes: dehydrogenase (energymetabolism), glucose-6-phosphatase (gluconeogenesis),albumin (protein synthesis), gluokinase (glucose uptake),and cytochrome P450 (xenobiotic metabolism), representthe zonation of metabolic and synthetic functions. Forexample, glucose-producing enzymes have higher levelsupstream in the sinusoid, where hepatocytes possess a highercapacity for glucose production via gluconeogenesis. Theregulation of liver zonation is controlled at the transcrip-tional level, and it seems to be tightly coupled to the O2
� 2009 Wiley Periodicals, Inc.

gradient via O2-sensitive transcription factors, where the O2
partial pressure varies from �65mmHg at the inlet of thesinusoid to �25mmHg at the outlet of the sinusoid(Kietzmann et al., 2006).
Since the O2 gradient is a major regulator of functionalzonation in the liver sinusoid, the aim of this simulationstudy is to model O2 transport in a hepatic HF bioreactorsupplemented with a mixture of two distinct types of O2
carriers in order to recapitulate the in vivo O2 gradient (pO2
ranging from 20–70mmHg) along the length of a single HF.One class of O2 carrier consists of hemoglobin-based O2
carriers (HBOCs), and the other consists of perfluorocar-bons (PFCs). O2 carriers are currently being developed totransport O2 to tissues and organs in vivo as red blood cellsubstitutes (Cohn and Cushing, 2009; Sakai et al., 2009). Inthis work we propose to use O2 carriers to oxygenate HF-based BLADs.
The fundamental difference between HBOCs and PFCsdepends on whether the O2 molecule is covalently bound tothe carrier molecule or physically dissolved within it (Riess,2001). For HBOCs, O2 is covalently bound to the hemegroup of the hemoglobin (Hb) molecule (Riess, 2001). Incontrast, O2 is physically dissolved in PFCs, whichpossess O2 solubilities that are about 20 times greater thanthat of water (Riess, 2006). PFCs are synthetic, inert, andextremely hydrophobic molecules so that the only way toutilize PFCs as an O2 carrier in aqueous solution is toemulsify them with a surfactant in order to form anemulsion.
Previous modeling studies have evaluated how muchbenefit can be derived from the use of O2 carriers intransfusion applications. For example, mathematical modelswere developed by Patel andMehar (1998a,b) to simulate O2
transport in a tubular blood vessel and its uptake bysurrounding tissue when transfusing a PFC emulsioninto the blood stream. The results of these studies showedthat a large increase in O2 delivery to tissues can be obtainedby transfusing a PFC emulsion into the blood streamcombined with breathing in a 100% O2 atmosphere.Another mathematical model (Gundersen et al., 2009)simulated O2 and NO transport in an arteriole transfusedwith HBOCs. The results of this work showed thatadministration of HBOCs can increase O2 delivery andmaintain tissue oxygenation during severe blood loss atphysiological O2 levels, despite the fact that HBOCsscavenge large amounts of NO which could result invasoconstriction.
Computational fluid dynamics has been used to redesignthe HF bioreactor in order to improve O2 transport. Mareelset al. (2006) suggested that increasing the number of gascapillaries and increasing the inlet pO2 maximally improvesO2 availability based on simulation of the AMC-bioartificialliver.
In our group, mathematical models were developed todescribe O2 transport in hepatic HF bioreactor supple-mented with either pure HBOCs or PFCs (Chen and Palmer,2009a,b; Sullivan and Palmer, 2006; Sullivan et al., 2006,
2007, 2008). It has been shown that both HBOCs and PFCs,by themselves, can enhance O2 delivery to culturedhepatocytes versus the case of no O2 carrier present.
The aim of this study is to explore the benefit ofsupplementing the circulating cell culture media withmixtures of HBOCs and PFCs. A steady state, 2-Dmodel wasdeveloped to calculate the O2 distribution in the HFbioreactor and evaluate the potential of mixtures of HBOCsand PFCs to recapitulate the in vivo pO2 spectrum of theliver sinusoid (pO2 ranging from 20 to 70mmHg). It wasfound that mixtures of HBOCs and PFCs increased O2
delivery to cultured hepatocytes and further improvedthe O2 distribution in the ECS space.
Computational Methods
Several assumptions have been made to simplify the O2
transport model to yield a manageable set of partialdifferential equations (PDEs) which describe fluid transportin an individual HF; as well as, mass transport of O2
dissolved in the aqueous cell culture media, PFC particlesand O2 bound to the HBOC (oxyHBOC): (1) parallel HFs inthe bioreactor bundle are considered to be evenly distributedand interactions between HFs are neglected so that a singleHF is modeled; (2) the system is at steady state, andaxisymmetric in the cylindrical coordinate system; (3) thefluid in the HF lumen contains a mixture of cell culturemedia, HBOC, and PFC and is treated as a pseudo-homogeneous fluid, since the PFC particles are very smallcompared to the HF diameter (�1/1,000 aspect ratio); (4)the permeability and diffusivity of dissolved O2 in themembrane/ECS is assumed to be uniform; (5) the totalHBOC concentration and PFC particle density are assumedto be constant throughout the HF lumen.
Figure 1A shows a schematic of the hepatic HF bioreactorsystem, whereas Figure 1B shows the HF cartridge (the HFcartridge modeled in this study contains 2,025 individualHFs), and Figure 1C shows the modeled domain beingcomposed of a single HF with three cylindrical compart-ments, that is the lumen (where the cell culture media flowsthrough), membrane (which separates the cultured cell massin the ECS from the circulating cell culture media/HBOC/PFC in the lumen), and ECS (where cultured cells reside).The geometry of the HF system to be modeled is based on acommercial HF bioreactor product (cat. no. 400-011,Spectrum Laboratories, Rancho Dominguez, CA).
The velocity profile in each of the three subdomains of themodel is calculated from a set of momentum transportPDEs. As the fluid in the HF lumen is within the laminarflow regime and assumed to be pseudo-homogeneous, theNavier–Stokes equation is applied to the lumen subdomain(0< r<RL) with effective viscosity (meff) (Eq. 1A). Both theHF membrane and ECS are treated as porous media withuniform intrinsic permeability, so that fluid motion inthese subdomains is described by Brinkman’s equation(RL< r<RE) (Bear, 1972), shown in Equation (1B), where v
Chen and Palmer: Effect of HBOCs and PFCs in HF Bioreactors 535
Biotechnology and Bioengineering
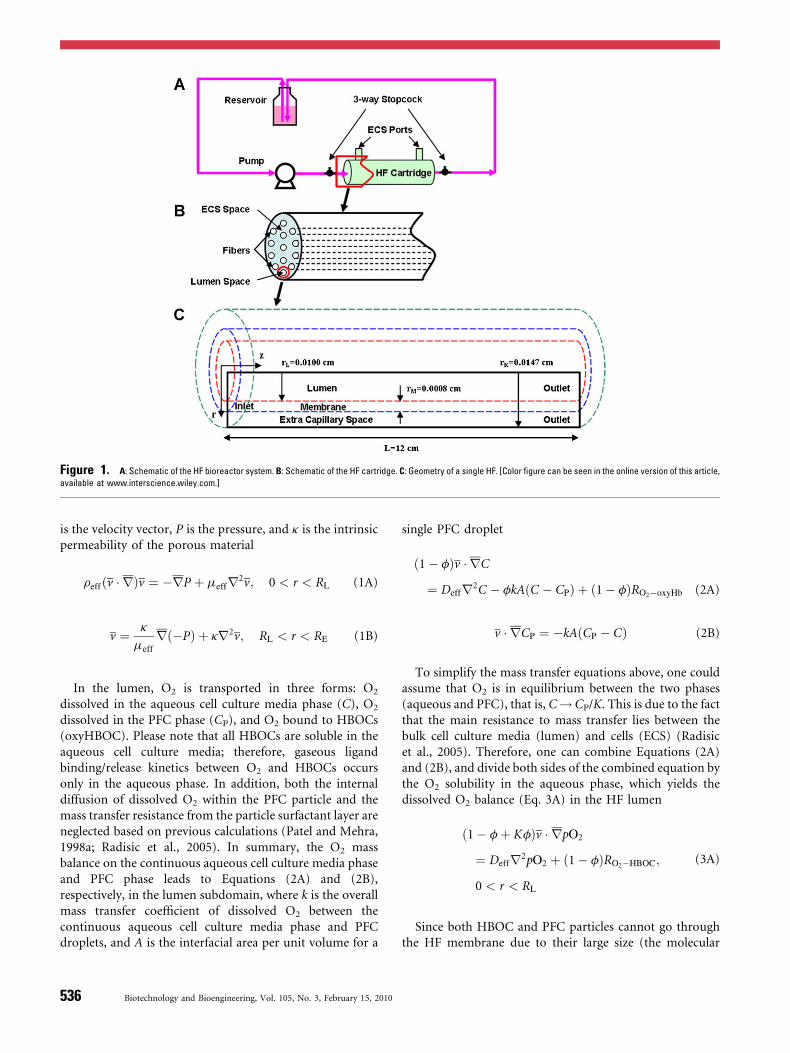
Figure 1. A: Schematic of the HF bioreactor system. B: Schematic of the HF cartridge. C: Geometry of a single HF. [Color figure can be seen in the online version of this article,
available at www.interscience.wiley.com.]
is the velocity vector, P is the pressure, and k is the intrinsicpermeability of the porous material
reffðv � rÞv ¼ �rP þ meffr2v; 0 < r < RL (1A)
v ¼ k
meff
rð�PÞ þ kr2v; RL < r < RE (1B)
In the lumen, O2 is transported in three forms: O2
dissolved in the aqueous cell culture media phase (C), O2
dissolved in the PFC phase (CP), and O2 bound to HBOCs(oxyHBOC). Please note that all HBOCs are soluble in theaqueous cell culture media; therefore, gaseous ligandbinding/release kinetics between O2 and HBOCs occursonly in the aqueous phase. In addition, both the internaldiffusion of dissolved O2 within the PFC particle and themass transfer resistance from the particle surfactant layer areneglected based on previous calculations (Patel and Mehra,1998a; Radisic et al., 2005). In summary, the O2 massbalance on the continuous aqueous cell culture media phaseand PFC phase leads to Equations (2A) and (2B),respectively, in the lumen subdomain, where k is the overallmass transfer coefficient of dissolved O2 between thecontinuous aqueous cell culture media phase and PFCdroplets, and A is the interfacial area per unit volume for a
536 Biotechnology and Bioengineering, Vol. 105, No. 3, February 15, 2010
single PFC droplet
ð1� fÞv � rC
¼ Deffr2C � fkAðC � CPÞ þ ð1� fÞRO2�oxyHb (2A)
v � rCP ¼ �kAðCP � CÞ (2B)
To simplify the mass transfer equations above, one couldassume that O2 is in equilibrium between the two phases(aqueous and PFC), that is, C!CP/K. This is due to the factthat the main resistance to mass transfer lies between thebulk cell culture media (lumen) and cells (ECS) (Radisicet al., 2005). Therefore, one can combine Equations (2A)and (2B), and divide both sides of the combined equation bythe O2 solubility in the aqueous phase, which yields thedissolved O2 balance (Eq. 3A) in the HF lumen
ð1� fþ KfÞv � rpO2
¼ Deffr2pO2 þ ð1� fÞRO2�HBOC;
0 < r < RL
(3A)
Since both HBOC and PFC particles cannot go throughthe HF membrane due to their large size (the molecular

weight cut-off of the HF membrane is 30 kDa), thedissolved O2 balance in the HF membrane and ECSsubdomain simplifies to Equation (3B), only the reactionrate of O2 ðRO2Þ in the HF membrane is zero while in theECS it obeys Michaelis–Menten kinetics (Eq. 4)
v � rpO2 ¼ Dr2pO2 þ RO2 ; RL < r < RE (3B)
aRO2 ¼0; RL < r < RM
� VMpO2
kMþpO2; RM < r < RE
�(4)
In Equation (3A), the reaction term can be described byEquation (5A) (Chen and Palmer, 2009a), given that the O2-HBOC equilibrium curve is known, which itself is a functionof pO2 (Eq. 5B, where a1–a4 are the Adair constants)
� RoxyHb�O2 ¼ aRO2�oxyHb
¼ �k�½HBOC� Seq1� Seq
ð1� SÞ � S
� �(5A)
Seq ¼a1pO2 þ 2a2pO
22 þ 3a3pO
32 þ 4a4pO
42
4ða1pO2 þ a2pO22 þ a3pO
32 þ a4pO
42Þ
(5B)
Here, another unknown parameter S (molar ratio ofoxygenated HBOC (oxyHBOC) to total HBOC) is intro-duced, and it then becomes necessary to include the massbalance of oxyHBOC in the transport model, shown inEquation (6). Please note that oxyHBOC is confined withinthe lumen subdomain
v � rCoxyHBOC ¼ DHBOC � r2CoxyHBOC þ RoxyHBOC�O2 (6)
Table I. Constants and parameters used in the O2 transport model.
Symbol Simulation parameter Valu
Physical constants
a O2 solubility in plain medium 1.71�m0 Viscosity of plain medium 6.92�r0 Density of plain medium 1.0
m1 Viscosity of pure PFC 1.92�r1 Pure PFC density 1.9
K Partition coefficient of O2 in water/PFC 20
kM Intrinsic permeability of HF membrane 1.04� 1
eM Membrane porosity 0.8
kE Intrinsic permeability of HF ECS 1.4� 1
eE ECS porosity 0.4
Da O2 diffusivity in plain medium 3.0� 1
Dp O2 diffusivity in pure PFC 3.7� 1
DM O2 diffusivity in membrane 2.5� 1
DE O2 diffusivity in ECS 2.0� 1
Vmax Maximum O2 consumption rate 0.03
kM Michaelis constant 3
Varied parameters
Q Total volumetric flow rate through entire HF cartridge 4.72, 16.
f PFC volume fraction 0–20%
pO2,in Inlet O2 partial pressure 5–15
[HBOC] HBOC concentration 0–10
The centerline of the lumen is axisymmetric, and the ECSouter boundary is assumed to connect to another HF ECSunit so as to be symmetric. The entrance of the lumen is setto parabolic flow with uniform pO2/PFC volume fraction/HBOC concentration, and both the HBOCs and PFCs areassumed to be in equilibrium with the local O2 concentra-tion at the entrance of the HF. The outlet of the lumen is setto an arbitrary pressure with convective mass flux.Interfacial boundaries between subdomains are continuousfor O2 and momentum, and the rest of the boundaries aretaken as no-slip at the walls. For oxyHBOC, the interfacialcylinder between the lumen and membrane is set to no-slipat the wall.
Constants/parameters used in the simulations are listed inTable I. Calculation of fluid properties can be found in theAppendix. The physical properties of the PFC are taken frompure F-octyl bromide (PFOB) which is the majorcomponent of the commercial PFC product OxygentTM
(Alliance Pharmaceutical Corp., San Diego, CA). Fourdifferent types of HBOCs were simulated: MP4 (apoly(ethylene glycol)-conjugated human Hb) (Bjorkholmet al., 2005; Vandegriff et al., 2003), human Hb (hHb),bovine Hb (BvHb), and polymerized bovine Hb (PolyBvHb)(Buehler et al., 2005). The size, O2-HBOC equilibriumconstants, and O2 dissociation rate constant for each HBOCare listed in Table II.
Results and Discussion
The coupled set of PDEs was solved by the finite elementmethod in Comsol Multiphysics (Comsol, Inc., Burlington,MA). First, the velocity and pressure profiles were calculatedfrom fluid transport equations (Eq. 1) and the continuity
e Units Source
10�3 mol/(m3mmHg) Sander (1999)
10�4 kg/(m s) Bird et al. (2002)
g/cm3
10�3 kg/(m s) Riess (2001)
2 g/cm3
0�20 m2 Bear (1972)
0�13 m2 Nield and Bejan (2006)
3
0�5 cm2/s Foy et al. (1994) and Piret and Cooney (1991)
0�5 cm2/s Middleman (1998)
0�5 cm2/s Hay et al. (2000)
0�5 cm2/s Piret and Cooney (1991)
5 mol/(m3 s) Smith et al. (1996)
mmHg Hay et al. (2000)
8, 28.6 mL/min
(v/v)
5 mmHg
0% �8.8mol/m3
Chen and Palmer: Effect of HBOCs and PFCs in HF Bioreactors 537
Biotechnology and Bioengineering
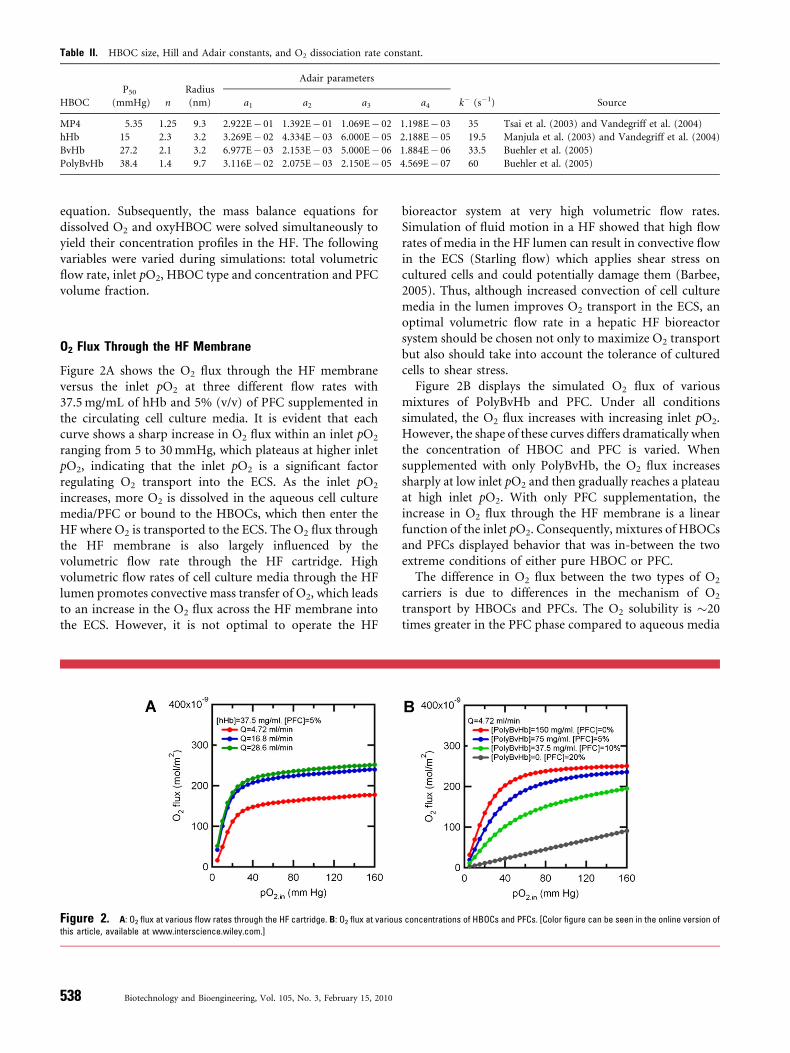
Table II. HBOC size, Hill and Adair constants, and O2 dissociation rate constant.
HBOC
P50(mmHg) n
Radius
(nm)
Adair parameters
k� (s�1) Sourcea1 a2 a3 a4
MP4 5.35 1.25 9.3 2.922E� 01 1.392E� 01 1.069E� 02 1.198E� 03 35 Tsai et al. (2003) and Vandegriff et al. (2004)
hHb 15 2.3 3.2 3.269E� 02 4.334E� 03 6.000E� 05 2.188E� 05 19.5 Manjula et al. (2003) and Vandegriff et al. (2004)
BvHb 27.2 2.1 3.2 6.977E� 03 2.153E� 03 5.000E� 06 1.884E� 06 33.5 Buehler et al. (2005)
PolyBvHb 38.4 1.4 9.7 3.116E� 02 2.075E� 03 2.150E� 05 4.569E� 07 60 Buehler et al. (2005)
equation. Subsequently, the mass balance equations fordissolved O2 and oxyHBOC were solved simultaneously toyield their concentration profiles in the HF. The followingvariables were varied during simulations: total volumetricflow rate, inlet pO2, HBOC type and concentration and PFCvolume fraction.
O2 Flux Through the HF Membrane
Figure 2A shows the O2 flux through the HF membraneversus the inlet pO2 at three different flow rates with37.5mg/mL of hHb and 5% (v/v) of PFC supplemented inthe circulating cell culture media. It is evident that eachcurve shows a sharp increase in O2 flux within an inlet pO2
ranging from 5 to 30mmHg, which plateaus at higher inletpO2, indicating that the inlet pO2 is a significant factorregulating O2 transport into the ECS. As the inlet pO2
increases, more O2 is dissolved in the aqueous cell culturemedia/PFC or bound to the HBOCs, which then enter theHF where O2 is transported to the ECS. The O2 flux throughthe HF membrane is also largely influenced by thevolumetric flow rate through the HF cartridge. Highvolumetric flow rates of cell culture media through the HFlumen promotes convective mass transfer of O2, which leadsto an increase in the O2 flux across the HF membrane intothe ECS. However, it is not optimal to operate the HF
Figure 2. A: O2 flux at various flow rates through the HF cartridge. B: O2 flux at variou
this article, available at www.interscience.wiley.com.]
538 Biotechnology and Bioengineering, Vol. 105, No. 3, February 15, 2010
bioreactor system at very high volumetric flow rates.Simulation of fluid motion in a HF showed that high flowrates of media in the HF lumen can result in convective flowin the ECS (Starling flow) which applies shear stress oncultured cells and could potentially damage them (Barbee,2005). Thus, although increased convection of cell culturemedia in the lumen improves O2 transport in the ECS, anoptimal volumetric flow rate in a hepatic HF bioreactorsystem should be chosen not only to maximize O2 transportbut also should take into account the tolerance of culturedcells to shear stress.
Figure 2B displays the simulated O2 flux of variousmixtures of PolyBvHb and PFC. Under all conditionssimulated, the O2 flux increases with increasing inlet pO2.However, the shape of these curves differs dramatically whenthe concentration of HBOC and PFC is varied. Whensupplemented with only PolyBvHb, the O2 flux increasessharply at low inlet pO2 and then gradually reaches a plateauat high inlet pO2. With only PFC supplementation, theincrease in O2 flux through the HF membrane is a linearfunction of the inlet pO2. Consequently, mixtures of HBOCsand PFCs displayed behavior that was in-between the twoextreme conditions of either pure HBOC or PFC.
The difference in O2 flux between the two types of O2
carriers is due to differences in the mechanism of O2
transport by HBOCs and PFCs. The O2 solubility is �20times greater in the PFC phase compared to aqueous media
s concentrations of HBOCs and PFCs. [Color figure can be seen in the online version of

because of its extremely strong intramolecular bonding (C–F) and weak intermolecular interactions (Riess, 2001). PFCsupplementation in the flowing media stream simplyincreases the solubility of O2 in solution. In contrast, O2
binds to HBOCs via covalent bonds, which allows loading/unloading of large amounts of O2 within the physiologicalpO2 range. Generally within the simulated range ofvariables, the O2 flux increases as the concentration ofHBOC increases in the HBOC/PFC mixture. O2 transportin the bioreactor is much more improved with onlyPolyBvHb ([PolyBvHb]¼ 150mg/mL) in solution com-pared with only PFC in solution ([PFC]¼ 20%, v/v). Inother words, HBOCs are more efficient at delivering O2 tothe ECS space under the simulated range of O2 tensionscompared to PFCs.
Normalized O2 Flux Through the HF Membrane
Our group has demonstrated that different HBOCs havevarious oxygenation capabilities due to their O2 affinity (P50,pO2 at which the HBOC is half-saturated with O2) and O2
dissociation kinetics (k�) (Chen and Palmer, 2009a). InFigure 3A, the O2 flux was calculated from a series ofsimulations with the same PFC concentration but withdifferent types of HBOCs in the HBOC/PFC mixture, andthen normalized by the O2 flux from plain cell culturemedia. The enhancement of O2 transport is dramatic withsupplementation of HBOC and PFC in the circulating cellculture media, especially under low inlet pO2. The O2 fluxthrough the HF membrane increased up to 100-fold with amixture of MP4 and PFC as shown in the figure. Thenormalized O2 flux first increases, reaches a maximumvalue, and then decreases. Each HBOC exhibits its ownmaximum normalized O2 flux at different inlet pO2 levels,suggesting an optimal pO2 window at which oxygenation ismaximized for that particular HBOC. Generally, at low inletpO2’s (5–10mmHg), the MP4 and PFC mixture had thehighest O2 flux through the HFmembrane. Mixtures of hHband PFC best improves O2 transport at an inlet pO2 ranging
Figure 3. A: Normalized O2 flux with various HBOCs. B: Normalized O2 flux with HBOC
www.interscience.wiley.com.]
from 10 to 30mmHg. Mixtures of BvHb and PFC result inthe most oxygenated HF at an inlet pO2 ranging from 40 to60mmHg. When the inlet pO2 exceeds 70mmHg, mixturesof PolyBvHb and PFC show the most improved O2 deliveryin the HF system. Moreover, at an inlet pO2 levelgreater than 70mmHg, better oxygenation in the HFECS is apparent as the P50 of the HBOC increases(PolyBvHb>BvHb> hHb>MP4). In this case, one shouldcarefully choose the type of HBOC based on the inlet pO2 inorder to maximize enhancement of O2 transport.
Figure 3B displays the normalized O2 flux fromsimulations with 75mg/mL of BvHb, 10% (v/v) PFC andthe BvHb/PFCmixture. The normalized O2 flux with respectto pure PFC remains constant over the inlet pO2’s studied,since PFCs only increase the O2 solubility. The enhancementof O2 delivery by PFCs is not sensitive to the inlet pO2. Thenormalized O2 flux from pure HBOCs and mixtures ofHBOCs and PFCs almost overlap as the inlet pO2 rangesfrom 5 to 160mmHg, which suggests that the oxygenationcapability of pure HBOCs andmixtures of HBOCs and PFCsare somewhat similar under the simulated conditions. Fromthis point of view, supplementing the circulating cell culturemedia with a mixture of HBOCs and PFCs has limited effecton O2 transport in hepatic HF bioreactors, compared withsupplementing HBOCs alone. This is understandable giventhat the simulated pO2 window is relatively low (0–21% O2)and the maximum PFC fraction is only 20% (v/v) and not alarger volume fraction. It is expected that supplementing thecirculating cell culture media with a mixture of PFC andHBOC would be more efficient in improving O2 transfer athigher dissolved O2 levels (Patel and Mehra, 1998a). Forinstance, improvement in O2 delivery by the PFC wouldincrease up to fivefold if the cell culture media at the inlet ofthe HF was in equilibrium with pure O2 at 1 atm to yield aninlet pO2 of 760mmHg. However, in mammalian cellculture, high O2 levels induces lipid peroxidation and resultsin the formation of cell-damaging free radicals (D’Angio andFinkelstein, 2000). Hence, operating bioreactors at high O2
levels is not advisable.
s and PFCs. [Color figure can be seen in the online version of this article, available at
Chen and Palmer: Effect of HBOCs and PFCs in HF Bioreactors 539
Biotechnology and Bioengineering

Figure 4. pO2 distribution in a single HF with various concentrations of BvHb and
PFC. [Color figure can be seen in the online version of this article, available at
www.interscience.wiley.com.]
O2 Concentration Profile in the HF
The pO2 level throughout the HF domain is displayed inFigure 4 at various BvHb concentrations and PFC volumefractions. In vivo, the pO2 level for primary hepatocyteslining the liver sinusoid usually spans 20–70mmHg (Allenand Bhatia, 2003). As mentioned earlier, hyperoxia results inlipid peroxidation, which leads to cell damage or even death.Therefore, we only showed simulation results at an inletpO2¼ 80mmHg. When there is no O2 carrier present (thecontrol is shown in the upper left hand corner of the figure),most of the HF space is hypoxic. Therefore, cell growth inthis hypoxic region will be limited by an insufficient O2
supply. As the HBOC and PFC concentration in thecirculating cell culture media increases, the ECS becomesbetter oxygenated. Even though the magnitude of thenormalized O2 flux is nearly the same for BvHb by itself andfor the mixture of BvHb and PFC in Figure 3B, improve-ment in the O2 distribution in the ECS is noticeable as thePFC concentration increases in the mixture. However, aconcentration of 75mg/mL of BvHb by itself in thecirculating cell culture media is sufficient to properly
Figure 5. pO2 distribution in a single HF domain and ECS zonation breakdown. [Co
cience.wiley.com.]
540 Biotechnology and Bioengineering, Vol. 105, No. 3, February 15, 2010
oxygenate the ECS, while the maximum volume fraction of20% PFC by itself in the circulating cell culture media stillleaves a considerably sized hypoxic region in the ECS. Inother words, HBOCs are more efficient oxygenating the ECSof the HF bioreactor compared to PFCs. However, mixturesof HBOCs and PFCs synergistically improve O2 transportinto the ECS. The hypoxic region is completely eliminated atHBOC concentrations >37.5mg/mL and PFC volumefractions >10%.
O2 Concentration Profile and ECS Zonation Breakdown
Figure 5 shows the O2 distribution within the HF withdifferent types of HBOCs (different P50’s) mixed withvarious concentrations of PFCs. As the P50 and PFCconcentration increases (top to bottom/left to right), higherconcentrations of O2 in the ECS region are realized at thesimulated inlet pO2 (80mmHg).
One of the goals of this study is to optimize the operatingparameters of the HF bioreactor system in order tomaximize the ECS volume where liver cells are exposedto the in vivo pO2 spectrum observed in the liver sinusoid(pO2 ranging from 20 to 70mmHg; where 20–35mmHgrepresents the perivenous region; 35–60mmHg representsthe pericentral region; and 60–70mmHg represents theperiportal region). Thus, identifying the percentage of cellsin the ECS that are exposed to each of the three oxygenationzones observed in vivo should help us to choose bioreactoroperating conditions that attempt to recapitulate these threeoxygenation regimes and the overall in vivo pO2 spectrum.
The right graph (ECS zonation breakdown at a PFCvolume fraction of 20%) shows that supplementation of thecirculating cell culture media with a mixture of PFC andeither MP4 or hHb results in a considerable fraction of theECS space that falls below the perivenous pO2 level, wherethe O2 supply is considered insufficient for hepatocytes tomaintain their various functions. On the other hand, themixture of PFC and either BvHb or PolyBvHb (high P50HBOCs) oxygenates the ECS space better than the low P50HBOCs, where the majority of the ECS space is within the
lor figure can be seen in the online version of this article, available at www.inters-

three oxygenation regimes observed in vivo (the pO2 in theECS ranges from 20 to 70mmHg). Therefore, the in vivo O2
gradient is recapitulated in the HF ECS under the simulatedconditions, although the fraction of the ECS experiencingboth perivenous and periportal oxygenation is very smallcompared to the pericentral region.
Conclusion
HBOCs and PFCs are under commercial development asoxygen-carrying solutions. The benefits of using each type ofoxygen carrier in oxygenating the ECS of HF bioreactorshave been previously explored in prior work. In this work, itwas observed that mixtures of HBOCs and PFCs yield asynergistic effect in improving O2 transport in a hepaticHF bioreactor. The trans-membrane O2 flux increasesdramatically with supplementation of HBOCs and PFCsin the circulating cell culture media. In addition, the O2
distribution in the ECS space was markedly improved withsupplementation of mixtures of HBOCs and PFCs. Morespecifically, an increase in the HBOC’s P50, the concentra-tion of HBOC and PFC improves O2 transport to the ECSand also recapitulates the in vivo O2 gradient of the liversinusoid.
Appendix: Calculation of Some Fluid Propertiesin the Transport Model
Effective density of the flowing cell culture media with amixture of HBOC and PFC
reff ¼ r0ð1� fÞ þ r1f
Effective viscosity of the flowing cell culture media with amixture of HBOC and PFC:
meff
m0
¼ 1þ m0 þ ð5=2Þm1
m0 þ m1
� �f
Effective O2 diffusivity in the HF lumen:
Deff ¼ Da 1þ 3ðg � 1Þðg þ 2Þf
� �; g ¼ KDp=Da
References
Allen JW, Bhatia SN. 2003. Formation of steady-state oxygen gradients in
vitro: Application to liver zonation. Biotechnol Bioeng 82(3):253–262.
Barbee KA. 2005. Mechanical cell injury. Ann N Y Acad Sci 1066:67–84.
Bear J. 1972. Dynamics of fluids in porous media. New York: American
Elsevier Pub. Co.
Bird RB, Stewart WE, Lightfoot EN. 2002. Transport phenomena.
New York: John Wiley & Sons, Inc.
BjorkholmM, Fagrell B, Przybelski R, Winslow N, Young M, Winslow RM.
2005. A phase I single blind clinical trial of a new oxygen transport
agent (MP4), human hemoglobin modified with maleimide-activated
poly(ethylene glycol). Haematologica 90(4):505–515.
Buehler PW, Boykins RA, Jia Y, Norris S, Freedberg DI, Alayash AI. 2005.
Structural and functional characterization of glutaraldehyde-polymer-
ized bovine hemoglobin and its isolated fractions. Anal Chem 77(11):
3466–3478.
Centis V, Vermette P. 2009. Enhancing oxygen solubility using hemoglobin-
and perfluorocarbon-based carriers. Front Biosci 14:665–688.
Chan C, Berthiaume F, Nath BD, Tilles AW, Toner M, Yarmush ML. 2004.
Hepatic tissue engineering for adjunct and temporary liver support:
Critical technologies. Liver Transpl 10(11):1331–1342.
Chen G, Palmer AF. 2009a. Hemoglobin-based oxygen carrier and con-
vection enhanced oxygen transport in a hollow fiber bioreactor.
Biotechnol Bioeng 102(6):1603–1612.
Chen G, Palmer AF. 2009b. Perfluorocarbon facilitated O(2) transport
in a hepatic hollow fiber bioreactor. Biotechnol Prog 25(5):1317–
1321.
Cohn CS, Cushing MM. 2009. Oxygen therapeutics: Perfluorocarbons and
blood substitute safety. Crit Care Clin 25(2):399–414.
D’Angio CT, Finkelstein JN. 2000. Oxygen regulation of gene expression: A
study in opposites. Mol Genet Metab 71:371–380.
Foy BD, Rotem A, Toner M, Tompkins RG, YarmushML. 1994. A device to
measure the oxygen uptake rate of attached cells: Importance in
bioartificial organ design. Cell Transpl 3(6):515–527.
Gundersen SI, Chen G, Palmer AF. 2009. Mathematical model of NO
and O2 transport in an arteriole facilitated by hemoglobin based O2
carriers. Biophys Chem 143(1–2):1–17.
Hay PD, Veitch AR, Smith MD, Cousins RB, Gaylor JD. 2000. Oxygen
transfer in a diffusion-limited hollow fiber bioartificial liver. Artif
Organs 24(4):278–288.
Jungermann K, Kietzmann T. 1996. Zonation of parenchymal and non-
parenchymal metabolism in liver. Annu Rev Nutr 16:179–203.
Kietzmann T, Dimova EY, Flugel D, Scharf JG. 2006. Oxygen: Modulator of
physiological and pathophysiological processes in the liver.
Z Gastroenterol 44(1):67–76.
Mareels G, Poyck PP, Eloot S, Chamuleau RA, Verdonck PR. 2006. Three-
dimensional numerical modeling and computational fluid dynamics
simulations to analyze and improve oxygen availability in the AMC
bioartificial liver. Ann Biomed Eng 34(11):1729–1744.
Middleman S. 1998. An introduction tomass and heat transfer: Principles of
analysis and design. John Wiley & Sons, Inc.
Millis JM, Losanoff JE. 2005. Technology insight: Liver support systems. Nat
Clin Pract Gastroenterol Hepatol 2(9):398–405.
Nield DA, Bejan A. 2006. Convection in porous media. New York: Springer.
15 p.
Patel S, Mehra A. 1998a. Modeling of oxygen transport in blood-perfluor-
ocarbon emulsion mixtures. Part 1: Oxygen uptake in tubular vessels.
ASAIO J 44:144–156.
Patel S, Mehra A. 1998b. Modeling of oxygen transport in blood-perfluor-
ocarbon emulsion mixtures. Part 2: Tissue oxygenation. ASAIO J
44:157–165.
Piret JM, Cooney CL. 1991. Model of oxygen-transport limitations in
hollow fiber bioreactors. Biotechnol Bioeng 37(1):80–92.
Radisic M, Deen W, Langer R, Vunjak-Novakovic G. 2005. Mathematical
model of oxygen distribution in engineered cardiac tissue with parallel
channel array perfused with culture medium containing oxygen car-
riers. Am J Physiol Heart Circ Physiol 288:H1278–H1289.
Riess JG. 2001. Oxygen carriers (‘blood substitutes’)—Raison d’etre,
chemistry, and some physiology. Chem Rev 101:2797–2919.
Riess JG. 2006. Perfluorocarbon-based oxygen delivery. Artif Cells Blood
Substit Immobil Biotechnol 34:567–580.
Sakai H, Sou K, Horinouchi H, Kobayashi K, Tsuchida E. 2009. Review of
hemoglobin-vesicles as artificial oxygen carriers. Artif Organs 33(2):
139–145.
Sander R. 1999. Compilation of Henry’s law constants for inorganic and
organic species of potential importance in environmental chemistry.
http://www.mpch-mainz.mpg.de/�sander/res/henry.html
Smith MD, Smirthwaite AD, Cairns DE, Cousins RB, Gaylor JD. 1996.
Techniques for measurement of oxygen consumption rates of hepa-
tocytes during attachment and post-attachment. Int J Artif Organs
19(1):36–44.
Sullivan JP, Palmer AF. 2006. Targeted oxygen delivery within hepatic
hollow fiber bioreactors via supplementation of hemoglobin-based
oxygen carriers. Biotechnol Prog 22:1374–1387.
Chen and Palmer: Effect of HBOCs and PFCs in HF Bioreactors 541
Biotechnology and Bioengineering

Sullivan JP, Gordon JE, Palmer AF. 2006. Simulation of oxygen carrier
mediated oxygen transport to C3A hepatoma cells housed within a
hollow fiber bioreactor. Biotechnol Bioeng 93(2):306–317.
Sullivan JP, Gordon JE, Bou-Akl T, Matthew HWT, Palmer AF. 2007.
Enhanced oxygen delivery to primary hepatocytes within a hollow fiber
bioreactor facilitated via hemoglobin-based oxygen carriers. Artif Cells
Blood Substit Immobil Biotechnol 35:585–606.
Sullivan JP, Harris DR, Palmer AF. 2008. Convection and hemoglobin-
based oxygen carrier enhanced oxygen transport in a hepatic hollow
fiber bioreactor. Artif Cells Blood Substit Immobil Biotechnol 36(4):
386–402.
542 Biotechnology and Bioengineering, Vol. 105, No. 3, February 15, 2010
Tsai AG, Vandegriff KD, Intaglietta M, Winslow RM. 2003. Targeted O2
delivery by low-P50 hemoglobin: A new basis for O2 therapeutics. Am J
Physiol Heart Circ Physiol 285(4):H1411–1419.
van de Kerkhove MP, Hoekstra R, Chamuleau RA, van Gulik TM. 2004.
Clinical application of bioartificial liver support systems. Ann Surg
240(2):216–230.
Vandegriff KD, Bellelli A, Samaja M, Malavalli A, Brunori M, Winslow RM.
2004. Kinetics of NO and O2 binding to a maleimide poly(ethylene
glycol)-conjugated human haemoglobin. Biochem J 382:183–189.
Vandegriff KD, Malavalli A, Wooldridge J, Lohman J, Winslow RM. 2003.
MP4, a new nonvasoactive PEG-Hb conjugate. Transfusion 43:509–516.