mir-450a-5p within rat adipose tissue exosome-like ... · research article mir-450a-5p within rat...
TRANSCRIPT

RESEARCH ARTICLE
miR-450a-5p within rat adipose tissue exosome-like vesiclespromotes adipogenic differentiation by targeting WISP2Yan Zhang1,2,3,*, Mei Yu1,2,*,‡, Minjia Dai1,2,3, Chang Chen1,2,3, Qi Tang1,2,3, Wei Jing1,2,3, Hang Wang1,2,3 andWeidong Tian1,2,3,‡
ABSTRACTAdipose tissue is an active endocrine organ that can secrete a widenumber of factors to regulate adipogenesis via paracrine signals. Inaddition to soluble proteins in adipose tissue, microRNAs (miRNAs)enriched in extracellular vesicles (EVs), such as exosomes ormicrovesicles, could modulate intercellular communications. In thisstudy, we demonstrated that exosome-like vesicles derived fromadipose tissue (Exo-AT) were internalized by adipose tissue-derivedstem cells (ADSCs), and that these, in turn, induced adipogenesis.High-throughput sequencing showed that 45 miRNAs were enrichedin Exo-AT, and 31.11% of them were associated with adipogenesis,compared with ADSC-derived exosome-like vesicles (Exo-ADSC).miR-450a-5p, one of the most abundant miRNAs in Exo-AT, was aproadipogenic miRNA. Further study demonstrated that miR-450a-5ppromoted adipogenesis through repressing expression of WISP2 bytargeting its 3′ untranslated region. Additionally, Exo-AT could alsodownregulate the expression of WISP2, while miR-450a-5p inhibitorreversed this effect. Moreover, inhibition of miR-450a-5p impairedadipogenesis mediated by exosome-like vesicles. In conclusion,Exo-AT mediates adipogenic differentiation through a mechanisminvolving transfer of miR-450a-5p.
KEY WORDS: Exosome-like vesicle, Adipose tissue, Adipogenesis,miR-450a-5p, WISP2
INTRODUCTIONIn adipose tissue, adipocytes are constantly being created anddestroyed throughout life (Rosen and Spiegelman, 2014). Inhumans, ∼10% of adipocytes are turned over every year, whilenearly 0.6% of adipocytes are renewed every day in mice(Rigamonti et al., 2011, Spalding et al., 2008). Adipose tissue-derived stem cells (ADSCs) serve as a reservoir and allow thecontinued renewal of precursor cells that can differentiate intoadipocytes (Bowers et al., 2006). However, differentiation ofadipocytes is a complex and multi-step process with many factorsand signaling pathways involved (Gustafson et al., 2015b). Theterminal differentiation of adipocytes has been extensively
characterized with the use of hormonal cocktails that typicallycontains dexamethasone (DEX), insulin, indomethacin andisobutylmethylxanthin (IBMX) (Rosen and Spiegelman, 2014,Wu et al., 2010). The cocktails act as direct inducers of genes thatregulate adipogenesis such as peroxisome proliferator-activatedreceptor γ (PPARγ), and CCAAT-enhancer-binding protein β (C/EBPβ) and δ (C/EBPδ) (Moseti et al., 2016; Wu et al., 2010).However, the differentiation cocktails cannot represent the naturaladipogenic process in vivo (Sarkanen et al., 2012). We know littleabout the adipogenic process in vivo, as the means of studying thisare most indirectly (Rosen and Spiegelman, 2014). Understandingadipogenesis in adipose tissue may provide means to developstrategies for the treatment and prevention of obesity and theapplication of adipose tissue regeneration.
Previous studies have demonstrated that adipose tissue extractpromotes adipogenesis and angiogenesis, indicating that adiposetissue can produce bioactive factors to regulate adipogenesis viaparacrine signals (Sarkanen et al., 2012; Li et al., 2014). Forexample, BMP4, secreted by adipocytes, can induce ADSCs toundergo adipogenic commitment (Gustafson et al., 2015a). Inaddition to secreting soluble proteins, adipose tissue also releasesextracellular vesicles (EVs) [including exosomes, small membranevesicles (30–100 nm) that are released by multivesicular bodiesfusing with the cell membrane and microvesicles, 100–1000 nmvesicles originated by shedding of the plasma membrane] (Denget al., 2009; Connolly et al., 2015; Muller et al., 2011; Fleury et al.,2016). However, the size range of exosomes overlaps partially withthat of microvesicles, which hinders a complete size-discriminationbetween these two populations of EVs (Campoy et al., 2016).Therefore, enriched vesicles having similar morphology and sizeconsistent with exosomes are defined as exosome-like vesicles(Altadill et al., 2016). Exosomes can facilitate intercellularcommunication by transferring proteins and non-coding RNAs(such as microRNAs; miRNAs) (Kahlert and Kalluri, 2013; Braicuet al., 2014). Although some functions or mechanisms remainelusive, it has been demonstrated that exosomes can modulate theirneighboring cells by influencing major cellular process includingapoptosis, cell differentiation, proliferation and metabolism (Qinet al., 2016; Braicu et al., 2014). One obvious point about thecontent of exosomes is that they contain miRNAs, which can inhibittranslation of mRNAs by targeting 3′ untranslated regions (UTRs)(Batiz et al., 2015). Transported miRNAs are capable of targetingmRNAs in recipient cells and, hence, are associated with cell-to-cellsignaling and communication (He et al., 2014; Valadi et al., 2007).
Emerging evidence shows that inhibition of miRNAs biogenesisimpairs differentiation of stem cells into adipocytes, which supportsthe idea that miRNAs may directly modulate adipocytedifferentiation (Xie et al., 2009; Zhang et al., 2016; Arner andKulyté, 2015). For example, miR-30a is a positive regulator ofadipocyte lineage commitment that acts through modulation ofReceived 23 September 2016; Accepted 31 January 2017
1State Key Laboratory of Oral Disease, West China School of Stomatology, SichuanUniversity, Chengdu 610041, China. 2National Engineering Laboratory for OralRegenerative Medicine, West China Hospital of Stomatology, Sichuan University,No. 14, 3rd Sec., Ren Min Nan Road, Chengdu, Sichuan Province 610041, P.R.China. 3Department of Oral and Maxillofacial Surgery, West China Hospital ofStomatology, Sichuan University, Chengdu 610041, China.*These authors contributed equally to this work
‡Authors for correspondence ([email protected]; [email protected])
Y.Z., 0000-0003-3079-9960; M.Y., 0000-0003-0306-6817; M.D., 0000-0002-2613-7588; C.C., 0000-0002-6049-1193; Q.T., 0000-0003-3048-1813; H.W., 0000-0001-6916-0887; W.T., 0000-0003-0175-1132
1158
© 2017. Published by The Company of Biologists Ltd | Journal of Cell Science (2017) 130, 1158-1168 doi:10.1242/jcs.197764
Journal
ofCe
llScience
Runx2 while miR-143 is required for terminal differentiation bymodulating MAP2K5–ERK5 (ERK5 is also known as MAPK7)signaling (Chen et al., 2016; Chen et al., 2014; Hamam et al., 2014).In addition, exosomal miRNAs also contribute to the process of celldifferentiation (Forterre et al., 2014; Xu et al., 2014). Previousstudies have shown that exosomes can promote differentiationof mesenchymal stem cells (MSCs) (such as osteogenesis,neurogenesis and angiogenesis) through exosomal miRNAs(Narayanan et al., 2016; Takeda and Xu, 2015; Cui et al., 2016;Lopatina et al., 2014). In adipose tissue, adipocyte-derivedexosomes deliver anti-osteoblastic miRNAs (miR-30c, miR-125a,miR-125b and miR-31) to osteoblasts, which reduces the levels ofosteoblastic differentiation markers (osteocalcin and osteopontin)(Martin et al., 2015).Although adipose tissue extract induces adipogenic
differentiation of MSCs, the role that exosomes play in thisprocess remains unknown. In this study, we hypothesized thatexosome-like vesicles derived from adipose tissue (Exo-AT) couldbe enriched with miRNA that would induce adipogenicdifferentiation of MSCs. We demonstrated that Exo-AT containedadipogenesis-associated miRNAs that could promote adipogenicdifferentiation of ADSCs. In particular, miR-450a-5p, which wasenriched in Exo-AT, could promote adipogenesis through targetingWISP2, a negative regulator of adipogenesis (Hammarstedt et al.,2013). This study expands our understanding of the adipogenicprocess in the adipose tissue microenvironment.
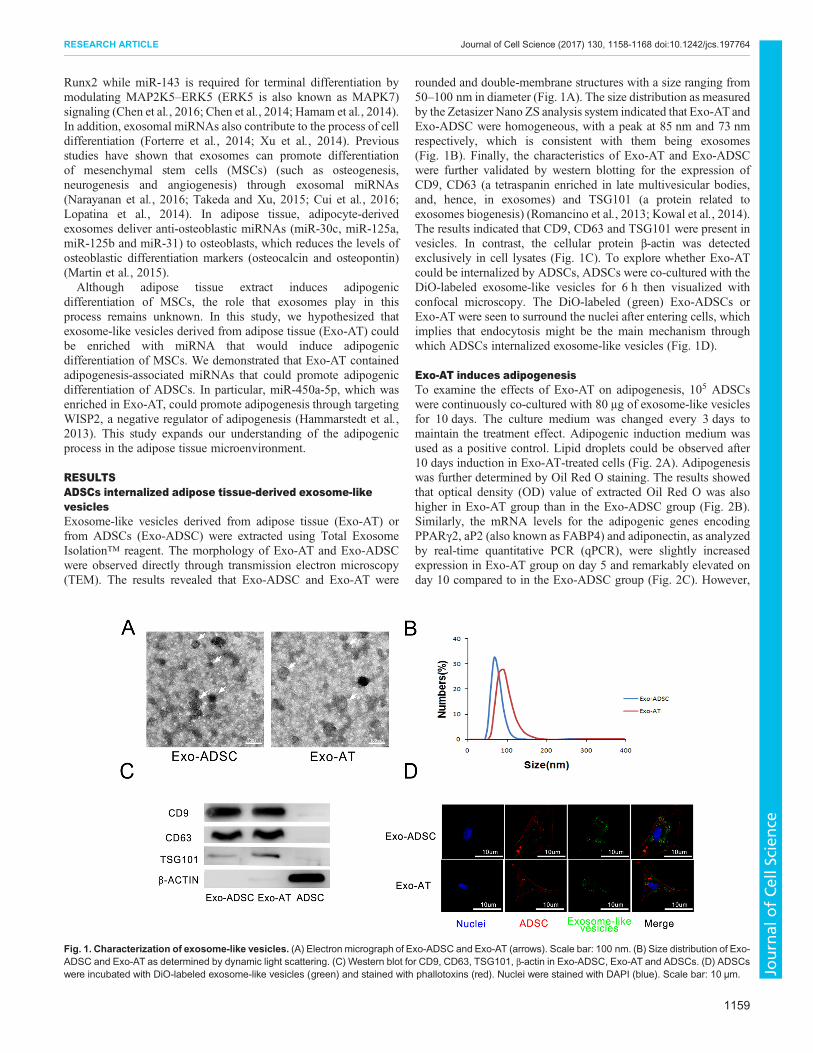
RESULTSADSCs internalized adipose tissue-derived exosome-likevesiclesExosome-like vesicles derived from adipose tissue (Exo-AT) orfrom ADSCs (Exo-ADSC) were extracted using Total ExosomeIsolation™ reagent. The morphology of Exo-AT and Exo-ADSCwere observed directly through transmission electron microscopy(TEM). The results revealed that Exo-ADSC and Exo-AT were
rounded and double-membrane structures with a size ranging from50–100 nm in diameter (Fig. 1A). The size distribution as measuredby the Zetasizer Nano ZS analysis system indicated that Exo-AT andExo-ADSC were homogeneous, with a peak at 85 nm and 73 nmrespectively, which is consistent with them being exosomes(Fig. 1B). Finally, the characteristics of Exo-AT and Exo-ADSCwere further validated by western blotting for the expression ofCD9, CD63 (a tetraspanin enriched in late multivesicular bodies,and, hence, in exosomes) and TSG101 (a protein related toexosomes biogenesis) (Romancino et al., 2013; Kowal et al., 2014).The results indicated that CD9, CD63 and TSG101 were present invesicles. In contrast, the cellular protein β-actin was detectedexclusively in cell lysates (Fig. 1C). To explore whether Exo-ATcould be internalized by ADSCs, ADSCs were co-cultured with theDiO-labeled exosome-like vesicles for 6 h then visualized withconfocal microscopy. The DiO-labeled (green) Exo-ADSCs orExo-AT were seen to surround the nuclei after entering cells, whichimplies that endocytosis might be the main mechanism throughwhich ADSCs internalized exosome-like vesicles (Fig. 1D).
Exo-AT induces adipogenesisTo examine the effects of Exo-AT on adipogenesis, 105 ADSCswere continuously co-cultured with 80 μg of exosome-like vesiclesfor 10 days. The culture medium was changed every 3 days tomaintain the treatment effect. Adipogenic induction medium wasused as a positive control. Lipid droplets could be observed after10 days induction in Exo-AT-treated cells (Fig. 2A). Adipogenesiswas further determined by Oil Red O staining. The results showedthat optical density (OD) value of extracted Oil Red O was alsohigher in Exo-AT group than in the Exo-ADSC group (Fig. 2B).Similarly, the mRNA levels for the adipogenic genes encodingPPARγ2, aP2 (also known as FABP4) and adiponectin, as analyzedby real-time quantitative PCR (qPCR), were slightly increasedexpression in Exo-AT group on day 5 and remarkably elevated onday 10 compared to in the Exo-ADSC group (Fig. 2C). However,
Fig. 1. Characterization of exosome-like vesicles. (A) Electron micrograph of Exo-ADSC and Exo-AT (arrows). Scale bar: 100 nm. (B) Size distribution of Exo-ADSC and Exo-AT as determined by dynamic light scattering. (C) Western blot for CD9, CD63, TSG101, β-actin in Exo-ADSC, Exo-AT and ADSCs. (D) ADSCswere incubated with DiO-labeled exosome-like vesicles (green) and stained with phallotoxins (red). Nuclei were stained with DAPI (blue). Scale bar: 10 µm.
1159
RESEARCH ARTICLE Journal of Cell Science (2017) 130, 1158-1168 doi:10.1242/jcs.197764
Journal
ofCe
llScience

the expression of mRNA encoding C/EBPδ, an early adipogenicgene, remained at a similar level (Fig. 2C). Western blottingrevealed that Exo-AT had an increased amount of PPARγ2,lipoprotein lipase (LPL) and adiponectin, while non-treatedADSCs and the Exo-ADSC group had a lower amount of theseproteins (Fig. 2D; Fig. S1A).
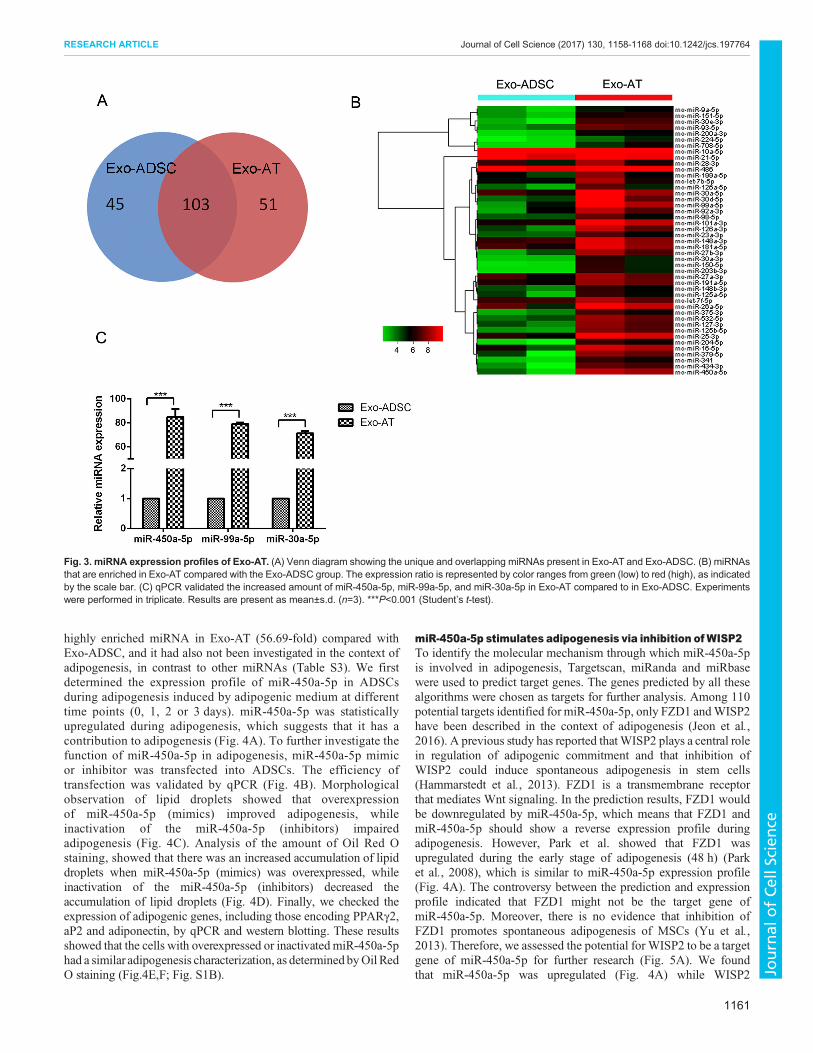
Adipogenic miRNAs were enriched in Exo-ATThe emerging evidence supports the theory that exosome cargomiRNAs function as important regulators in cell differentiation.Therefore, in order to determine the mechanism by which Exo-ATmediated adipogenesis, we profiled the miRNAs in Exo-AT in tworeplicates by using high-throughput sequencing (miRNA-seq);Exo-ADSC was also profiled as a comparative sample. Aftertrimming low-quality reads, contaminants, adaptors and readssmaller than 15 nt, the remaining reads were mapped to miRBasev21 (Table S2). A total of 148 and 154 types of known miRNAswere identified in Exo-ADSC and Exo-AT, respectively (Fig. 3A).Among these miRNAs, 103 miRNAs were detected in both Exo-ADSC and Exo-AT. Compared to Exo-ADSC, 45 conserved
miRNAs were enriched (expressed ≥2-fold, FDR<0.05) in Exo-AT (Fig. 3B). A total of 14 of the 45 miRNAs that were enriched inExo-AT (31.11%, such as miR-30a-5p and miR-148a-3p) arereported to participate in regulation of adipogenesis, whereas eightmiRNAs (17.78%, such as miR-93-5p and miR-150-3p) are knownto negatively control osteoblastic differentiation of MSCs, ninemiRNAs (20%, such as miR-150-3p, miR-126a-3p) are associatedwith angiogenesis, four miRNAs (8.89%, such as miR-224) arerelated to metabolism and 15 miRNAs have not been reported toregulate adipogenesis (Table S3). To validate these miRNA-seqresults, the expression of three most abundant miRNAs in Exo-AT(miR-450a-5p, miR-99a-5p and miR-30a-5p) were measured byqPCR. Consistent with the miRNA-seq results, qPCR showed thatExo-AT were enriched in miR-450a-5p, miR-99a-5p and miR-30a-5p compared with Exo-ADSC (Fig. 3C).
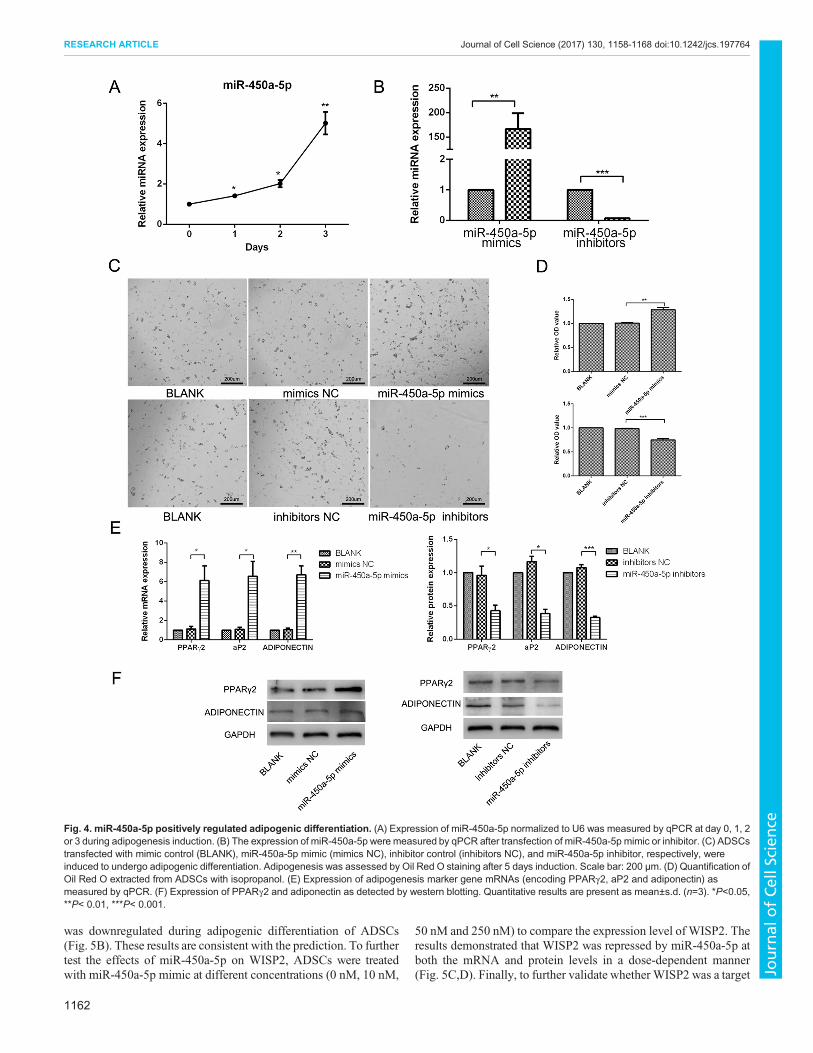
miR-450a-5p promotes adipogenesis of ADSCsIn order to further investigate the function of the miRNAs that wereenriched in Exo-AT, miR-450a-5p was selected as candidate since itis one of abundant miRNAs (reads count>1000) and also the most
Fig. 2. Exo-AT induces adipogenesis. (A) ADSCs cultured with Exo-ADSC and Exo-AT for 10 days were stained with Oil Red O to determine the level ofadipogenesis. ADSCs cultured with basal culture medium (NC) or adipogenic medium (ADIPO) were used as a negative and positive controls, respectively. Scalebar: 50 μm. (B) Quantification of the amount of Oil Red O extracted with isopropanol. (C) The relative expressions of mRNA encoding C/EBPδ, PPARγ2, aP2 andadiponectin was measured by qPCR on day 5 and 10 after induction. Results are present as mean±s.d. (n=3). *P<0.05; **P<0.01; ***P<0.001; ns, not significant.(D) Protein expression of PPARγ2, LPL and adiponectin was detected by western blotting on day 5 and 10 after induction.
1160
RESEARCH ARTICLE Journal of Cell Science (2017) 130, 1158-1168 doi:10.1242/jcs.197764
Journal
ofCe
llScience
highly enriched miRNA in Exo-AT (56.69-fold) compared withExo-ADSC, and it had also not been investigated in the context ofadipogenesis, in contrast to other miRNAs (Table S3). We firstdetermined the expression profile of miR-450a-5p in ADSCsduring adipogenesis induced by adipogenic medium at differenttime points (0, 1, 2 or 3 days). miR-450a-5p was statisticallyupregulated during adipogenesis, which suggests that it has acontribution to adipogenesis (Fig. 4A). To further investigate thefunction of miR-450a-5p in adipogenesis, miR-450a-5p mimicor inhibitor was transfected into ADSCs. The efficiency oftransfection was validated by qPCR (Fig. 4B). Morphologicalobservation of lipid droplets showed that overexpressionof miR-450a-5p (mimics) improved adipogenesis, whileinactivation of the miR-450a-5p (inhibitors) impairedadipogenesis (Fig. 4C). Analysis of the amount of Oil Red Ostaining, showed that there was an increased accumulation of lipiddroplets when miR-450a-5p (mimics) was overexpressed, whileinactivation of the miR-450a-5p (inhibitors) decreased theaccumulation of lipid droplets (Fig. 4D). Finally, we checked theexpression of adipogenic genes, including those encoding PPARγ2,aP2 and adiponectin, by qPCR and western blotting. These resultsshowed that the cells with overexpressed or inactivated miR-450a-5phad a similar adipogenesis characterization, as determined byOil RedO staining (Fig.4E,F; Fig. S1B).
miR-450a-5p stimulates adipogenesis via inhibition ofWISP2To identify the molecular mechanism through which miR-450a-5pis involved in adipogenesis, Targetscan, miRanda and miRbasewere used to predict target genes. The genes predicted by all thesealgorithms were chosen as targets for further analysis. Among 110potential targets identified for miR-450a-5p, only FZD1 andWISP2have been described in the context of adipogenesis (Jeon et al.,2016). A previous study has reported thatWISP2 plays a central rolein regulation of adipogenic commitment and that inhibition ofWISP2 could induce spontaneous adipogenesis in stem cells(Hammarstedt et al., 2013). FZD1 is a transmembrane receptorthat mediates Wnt signaling. In the prediction results, FZD1 wouldbe downregulated by miR-450a-5p, which means that FZD1 andmiR-450a-5p should show a reverse expression profile duringadipogenesis. However, Park et al. showed that FZD1 wasupregulated during the early stage of adipogenesis (48 h) (Parket al., 2008), which is similar to miR-450a-5p expression profile(Fig. 4A). The controversy between the prediction and expressionprofile indicated that FZD1 might not be the target gene ofmiR-450a-5p. Moreover, there is no evidence that inhibition ofFZD1 promotes spontaneous adipogenesis of MSCs (Yu et al.,2013). Therefore, we assessed the potential for WISP2 to be a targetgene of miR-450a-5p for further research (Fig. 5A). We foundthat miR-450a-5p was upregulated (Fig. 4A) while WISP2
Fig. 3. miRNA expression profiles of Exo-AT. (A) Venn diagram showing the unique and overlapping miRNAs present in Exo-AT and Exo-ADSC. (B) miRNAsthat are enriched in Exo-AT compared with the Exo-ADSC group. The expression ratio is represented by color ranges from green (low) to red (high), as indicatedby the scale bar. (C) qPCR validated the increased amount of miR-450a-5p, miR-99a-5p, and miR-30a-5p in Exo-AT compared to in Exo-ADSC. Experimentswere performed in triplicate. Results are present as mean±s.d. (n=3). ***P<0.001 (Student’s t-test).
1161
RESEARCH ARTICLE Journal of Cell Science (2017) 130, 1158-1168 doi:10.1242/jcs.197764
Journal
ofCe
llScience
was downregulated during adipogenic differentiation of ADSCs(Fig. 5B). These results are consistent with the prediction. To furthertest the effects of miR-450a-5p on WISP2, ADSCs were treatedwith miR-450a-5p mimic at different concentrations (0 nM, 10 nM,
50 nM and 250 nM) to compare the expression level of WISP2. Theresults demonstrated that WISP2 was repressed by miR-450a-5p atboth the mRNA and protein levels in a dose-dependent manner(Fig. 5C,D). Finally, to further validate whether WISP2 was a target
Fig. 4. miR-450a-5p positively regulated adipogenic differentiation. (A) Expression of miR-450a-5p normalized to U6 was measured by qPCR at day 0, 1, 2or 3 during adipogenesis induction. (B) The expression of miR-450a-5p were measured by qPCR after transfection of miR-450a-5p mimic or inhibitor. (C) ADSCstransfected with mimic control (BLANK), miR-450a-5p mimic (mimics NC), inhibitor control (inhibitors NC), and miR-450a-5p inhibitor, respectively, wereinduced to undergo adipogenic differentiation. Adipogenesis was assessed by Oil Red O staining after 5 days induction. Scale bar: 200 µm. (D) Quantification ofOil Red O extracted from ADSCs with isopropanol. (E) Expression of adipogenesis marker gene mRNAs (encoding PPARγ2, aP2 and adiponectin) asmeasured by qPCR. (F) Expression of PPARγ2 and adiponectin as detected by western blotting. Quantitative results are present as mean±s.d. (n=3). *P<0.05,**P< 0.01, ***P< 0.001.
1162
RESEARCH ARTICLE Journal of Cell Science (2017) 130, 1158-1168 doi:10.1242/jcs.197764
Journal
ofCe
llScience
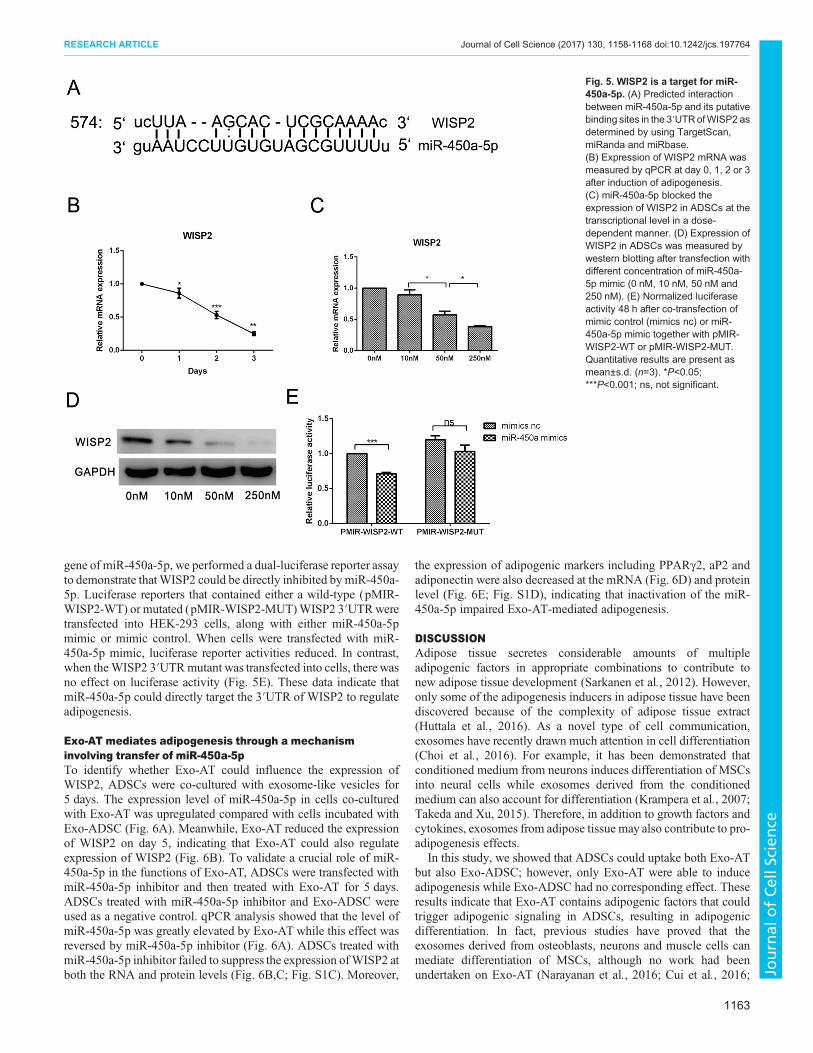
gene of miR-450a-5p, we performed a dual-luciferase reporter assayto demonstrate thatWISP2 could be directly inhibited bymiR-450a-5p. Luciferase reporters that contained either a wild-type (pMIR-WISP2-WT) or mutated (pMIR-WISP2-MUT)WISP2 3′UTRweretransfected into HEK-293 cells, along with either miR-450a-5pmimic or mimic control. When cells were transfected with miR-450a-5p mimic, luciferase reporter activities reduced. In contrast,when theWISP2 3′UTRmutant was transfected into cells, therewasno effect on luciferase activity (Fig. 5E). These data indicate thatmiR-450a-5p could directly target the 3′UTR of WISP2 to regulateadipogenesis.
Exo-AT mediates adipogenesis through a mechanisminvolving transfer of miR-450a-5pTo identify whether Exo-AT could influence the expression ofWISP2, ADSCs were co-cultured with exosome-like vesicles for5 days. The expression level of miR-450a-5p in cells co-culturedwith Exo-AT was upregulated compared with cells incubated withExo-ADSC (Fig. 6A). Meanwhile, Exo-AT reduced the expressionof WISP2 on day 5, indicating that Exo-AT could also regulateexpression of WISP2 (Fig. 6B). To validate a crucial role of miR-450a-5p in the functions of Exo-AT, ADSCs were transfected withmiR-450a-5p inhibitor and then treated with Exo-AT for 5 days.ADSCs treated with miR-450a-5p inhibitor and Exo-ADSC wereused as a negative control. qPCR analysis showed that the level ofmiR-450a-5p was greatly elevated by Exo-AT while this effect wasreversed by miR-450a-5p inhibitor (Fig. 6A). ADSCs treated withmiR-450a-5p inhibitor failed to suppress the expression ofWISP2 atboth the RNA and protein levels (Fig. 6B,C; Fig. S1C). Moreover,
the expression of adipogenic markers including PPARγ2, aP2 andadiponectin were also decreased at the mRNA (Fig. 6D) and proteinlevel (Fig. 6E; Fig. S1D), indicating that inactivation of the miR-450a-5p impaired Exo-AT-mediated adipogenesis.
DISCUSSIONAdipose tissue secretes considerable amounts of multipleadipogenic factors in appropriate combinations to contribute tonew adipose tissue development (Sarkanen et al., 2012). However,only some of the adipogenesis inducers in adipose tissue have beendiscovered because of the complexity of adipose tissue extract(Huttala et al., 2016). As a novel type of cell communication,exosomes have recently drawn much attention in cell differentiation(Choi et al., 2016). For example, it has been demonstrated thatconditioned medium from neurons induces differentiation of MSCsinto neural cells while exosomes derived from the conditionedmedium can also account for differentiation (Krampera et al., 2007;Takeda and Xu, 2015). Therefore, in addition to growth factors andcytokines, exosomes from adipose tissue may also contribute to pro-adipogenesis effects.
In this study, we showed that ADSCs could uptake both Exo-ATbut also Exo-ADSC; however, only Exo-AT were able to induceadipogenesis while Exo-ADSC had no corresponding effect. Theseresults indicate that Exo-AT contains adipogenic factors that couldtrigger adipogenic signaling in ADSCs, resulting in adipogenicdifferentiation. In fact, previous studies have proved that theexosomes derived from osteoblasts, neurons and muscle cells canmediate differentiation of MSCs, although no work had beenundertaken on Exo-AT (Narayanan et al., 2016; Cui et al., 2016;
Fig. 5. WISP2 is a target for miR-450a-5p. (A) Predicted interactionbetween miR-450a-5p and its putativebinding sites in the 3′UTRofWISP2 asdetermined by using TargetScan,miRanda and miRbase.(B) Expression of WISP2 mRNA wasmeasured by qPCR at day 0, 1, 2 or 3after induction of adipogenesis.(C) miR-450a-5p blocked theexpression of WISP2 in ADSCs at thetranscriptional level in a dose-dependent manner. (D) Expression ofWISP2 in ADSCs was measured bywestern blotting after transfection withdifferent concentration of miR-450a-5p mimic (0 nM, 10 nM, 50 nM and250 nM). (E) Normalized luciferaseactivity 48 h after co-transfection ofmimic control (mimics nc) or miR-450a-5p mimic together with pMIR-WISP2-WT or pMIR-WISP2-MUT.Quantitative results are present asmean±s.d. (n=3). *P<0.05;***P<0.001; ns, not significant.
1163
RESEARCH ARTICLE Journal of Cell Science (2017) 130, 1158-1168 doi:10.1242/jcs.197764
Journal
ofCe
llScience

Batiz et al., 2015; Choi et al., 2016). Therefore, we conclude thatexosomes may serve as a mediator of communication between stemcells and mature tissue cells. Once the stem cells differentiate intomature cells, they, in turn, induce stem cells to differentiate throughexosomes, thus a positive-feedback loop mechanism is formedduring differentiation (Fig. 7).Exosomes contain a wide contingent of functional proteins,
mRNAs and miRNAs (Sun et al., 2013). It has been indicated thatproteins and miRNAs are involved in exosome-mediateddifferentiation. For example, TGF-β1 in cancer cell exosomesinduce the differentiation of MSCs into cancer-associatedmyofibroblasts via the TGF-β1/SMAD signaling pathway(Chowdhury et al., 2015). Several studies have shown thatexosomal miRNAs participate in multiple differentiationprocesses, such as osteogenesis, neurogenesis, angiogenesis and
myogenesis (Lee et al., 2014; Qi et al., 2016), which indicates thatmiRNAs could be the main mechanism for exosome-mediateddifferentiation. However, whether miRNAs in Exo-AT acted as asignificant component in exosomes to promote adipogenesis hadnot been elucidated. Therefore, we focused on the role of exosomalmiRNAs in cell communication during adipogenesis. Interestingly,14 miRNAs enriched in Exo-AT had been previously reported topromote adipogenic differentiation of MSCs. Several of thesemiRNAs (fold change ≥20 compared to Exo-ADSC) had beendescribed as regulating adipogenesis (e.g. miR-125b-5p, miR-30e-3p and miR-204-5p), whereas several of them (e.g. miR-93-5p andmiR-203-3p) had been reported to negatively regulate osteogenesis(Table S3) (Kennell et al., 2008; Cho et al., 2016; Hamam et al.,2014; Yang et al., 2012; An et al., 2014). Eguchi et al. also analyzedthe miRNAs in adipocytes derived from EVs and detected several
Fig. 6. miR-450a-5p inhibitor impaired Exo-AT-mediated adipogenesis by targeting WISP2. ADSCs transfected with miR-450a-5p inhibitor (200 nM) werethen treatedwithExo-ATorExo-ADSC for 5 days. (A) Expression ofmiR-450a-5pwasmeasuredby qPCR. Expression ofWISP2mRNAwasmeasuredby qPCR (B)and western blotting (C). Expression of adipogenesis-induced genemRNA and protein (PPARγ2, aP2 and adiponectin) was measured by qPCR (D) and westernblotting (E). Quantitative results are present as mean±s.d. (n=3). *P<0.05, **P< 0.01, ***P<0.001.
1164
RESEARCH ARTICLE Journal of Cell Science (2017) 130, 1158-1168 doi:10.1242/jcs.197764
Journal
ofCe
llScience

adipogenic miRNAs (including miR-148a-3p, miR-181a-5p, miR-30a-5p, miR-125b-5p) (Eguchi et al., 2016), which is consistentwith our results (Table S3).Here, we identified that miR-450a-5p, one of the most abundant
miRNAs in Exo-AT, was a regulator of ADSC differentiationinto adipocytes. Normally, adipogenesis-related miRNAs aredownregulated in the obese subjects (Williams and Mitchell2012). Taken together with the fact that decreased expression ofmiR-450a-5p has been reported in obesity (Delic et al., 2016),miR-450a-5p might be involved in adipogenesis. Moreover, miR-450a-5p is downregulated when Wnt signaling, a negative signalregulating adipogenesis, is activated by Li+ during adipogenesis ofpreadipocytes (Qin et al., 2010). Li+ can activate Wnt signalingthrough the inhibition of GSK-3β and further blocks thephosphorylation of β-catenin (Ross et al., 2000). Therefore, thedownregulation of miR-450a-5p in Li+-treated pre-adipocytesindicates that miR-450a-5p might act as a positive regulator ofadipogenesis by suppressing Wnt signaling. In our results,inactivation of miR-450a-5p partially diminished the adipogenicactivity of Exo-AT. This suggests miR-450a-5p might be involvedin Exo-AT-mediated adipogenic differentiation.To determine the molecule mechanism for the action of exosomal
miR-450a-5p during adipogenesis, we performed a bioinformaticsanalysis for potential targets of miR-450a-5p and found WISP2.WISP2, also known as CCN5, is a novel adipokine secreted by adiposetissue (Lehr et al., 2012) that is highly expressed in MSCs andpreadipocytes (Hammarstedt et al., 2013). A previous study hassuggested that WISP2 promoted proliferation and inhibitedadipogenesis, while inhibition of WISP2 induced spontaneousadipogenesis (Hammarstedt et al., 2013). WISP2 is also known toregulate BMP4-dependent adipogenesis by forming a complex withZNF423, a transcriptional activator of PPARγ2, or by directlyactivating Wnt signaling (Gustafson et al., 2013). Hence, we
hypothesize that miR-450a-5p transferred from exosome-like vesiclesmight promote adipogenesis through downregulation of WISP2. Asexpected, ADSCs treatedwith Exo-ATor overexpressingmiR-450a-5phad a substantial decrease in the expression of WISP2. These findingsindicate a new role of miR-450a-5p transferred from exosome-likevesicles in regulating adipogenesis by modulating its target WISP2.
Although our data suggest that miR-450a-5p had an important rolein the Exo-AT-mediated adipogenesis, we cannot rule out thecontribution of other exosomal cargoes. Inhibition of miR-450a-5pdoes not arrest Exo-AT-mediated adipogenesis completely. Eguchiet al. performed a comprehensive protein content characterization ofEVs derived from stressed adipocytes using tandem massspectroscopy (LC-MS/MS) and found that the functions of proteinsin EVs were mainly involved in translation, carbohydrate and lipidmetabolism, cytoskeleton, extracellular matrix, ribonucleoproteinsand nucleosomes (Eguchi et al., 2016). Connolly et al. have reportedthat EVs from adipocytes are enriched in markers of adipogenesis(such as PREF-1, also known as DLK1, and PPARγ). The amount ofPREF-1, a negative regulator of adipogenesis, decreased in EVsderived from cells during the later stages of differentiation (Connollyet al., 2015). Other adipogenesis-related proteins were also detectedin adipocyte-derived exosomes including adenylate kinase 2 (AK2),and matrix metalloproteinase-2 and -9 (MMP-2 and MMP-9) (Leeet al., 2015; Aoki et al., 2007). Furthermore, mRNAs encoding theadipogenesis-related proteins PPARγ, leptin, C/EBP-α and C/EBP-δhave also been detected in adipocyte-derived EVs and as beingdelivered to osteoblasts (Martin et al., 2015). Therefore, componentsother than miRNAs in exosome-like vesicles may also participate inregulating adipogenesis.
Our results provide novel insights into the intercommunicationbetween ADSCs and adipose tissue cells in the adipose tissuemicroenvironment. Considering that exosome-like vesicles arecompletely cell-free and not highly immunogenic, exosome-likevesicles could be an important tool for use during adipose tissueengineering (Konala et al., 2016). Therefore, further research isneeded to study the possibility that exosome-like vesicles act as aproadipogenic factor to enable use during adipose tissue engineering.
MATERIALS AND METHODSIsolation of adipose tissue-derived stem cellsThe study was approved by the Ethical Committees of the State KeyLaboratory of Oral Diseases, West China School of Stomatology, SichuanUniversity, China. ADSCs were prepared and cultured as described in ourprevious studies (Jing et al., 2011; Zuk et al., 2002). Briefly, inguinal fat padswere collected from 4-week-old Sprague-Dawley (SD) rats and washedextensively with sterile phosphate-buffered saline (PBS) to remove the debrisand red blood cells. Then the samples were cut into small pieces (1–2 mm3)and treated with 0.075% collagenase (type I) for 30 min at 37°C. The cellswere maintained at 37°C in a CO2 incubator. When the cells reached 80%confluence, the cells were trypsinized and passaged at a 1:3 ratio.
To collect the culture medium, cells were cultured to 90% confluence,washed twice with PBS, switched to serum-free medium (α-MEM,100 U/ml penicillin and 100 μg/ml streptomycin) and cultured for 3 days.The serum-free culture medium was centrifuged at 150 g for 10 min toremove cell debris and kept at 4°C for further experiments.
Isolation of exosome-like vesiclesExo-AT or Exo-ADSC were obtained from adipose tissue extract (ATE) orADSC culture medium, respectively.
To obtain adipose tissue extract, 5 g adipose tissue explants, achieved bymincing adipose tissue from 4-week-old SD rats, was added into the CelstirSpinner Flask (Wheaton, IL) supplemented with 75 ml α-minimal essentialmedium (α-MEM, Hyclone), 100 U/ml penicillin and 100 μg/mlstreptomycin, and cultured with shaking at 100 rpm for 3 days. ATE was
Fig. 7. Schematic of hypothesis for the action of miR-450a-5p in ADSCs.Adipose tissue secretes exosome-like vesicles, which are taken up by ADSCs.miR-450a-5p contained in the exosome-like vesicles promotes adipogenesisby downregulating WISP2 in ADSCs.
1165
RESEARCH ARTICLE Journal of Cell Science (2017) 130, 1158-1168 doi:10.1242/jcs.197764
Journal
ofCe
llScience

centrifuged at 150 g for 10 min to remove the cell debris and stored at−80°C, after determining its protein concentration by the BCA method.
ATEorADSC culturemediumwas first introduced intoAmicon®Ultra-15Centrifugal Filter Units with an Ultracel-3 membrane (3 kDa molecular masscut off membrane, Millipore) and centrifuged at 5000 g for 30 min. Theconcentrate was then mixed with 0.5 volume of Total Exosome Isolation™reagent (Life Technologies), incubated overnight at 4°C and spundown for 1 hat 10,000 g at 4°C. The pellet was re-suspended in 100 μl PBS and stored at−80°C, after determining its protein concentration by the BCA method.
Transmission electron microscopyExosome-like vesicles were fixed with 1% glutaraldehyde at 4°C overnight.After washing, vesicles were loaded onto formvarcarbon-coated grids,negatively stained with aqueous phosphotungstic acid for 60 s and imagedwith a transmission electron microscope (Hitachi H7500 TEM, Japan).
Size distributionThe size of vesicles was determined by a dynamic light scattering techniqueusing a ZetasizerNano ZS analysis system (Zetasizer version 6.12; MalvernInstruments, UK). A size distribution plot, with the x-axis showing thedistribution of estimated particle radius (nm) and the y-axis showing therelative percentage, was made.
Exosome-like vesicle labeling and cellular uptakeExosome-like vesicles were labeled with a membrane-labeling dye DiO(Invitrogen) as previously described (Choi et al., 2016), and were thenwashedand resuspended in serum free α-MEM. Next, ADSCs were co-cultured withDiO-labeled vesicles for 6 h, washed with PBS three times, fixed in 4%paraformaldehyde, stained with phallotoxins (Invitrogen), washed with PBSand imaged by confocal microscopy (Olympus FV1000, Japan).
Induction of adipogenic differentiationADSCs at passage 3 were plated in six-well plates at a density of 105 cells/well, cultured for 24 h, then rinsed with PBS and incubated with 2 ml of oneof four different culture media for up to 10 days. The media used were: (1)basal medium [α-MEM supplemented with 10% fetal bovine serum (FBS)],as a negative control; (2) basal medium supplemented with Exo-ADSC(the concentration of Exo-ADSC was 40 µg/ml); (3) basal mediumsupplemented with Exo-AT (the concentration of Exo-AT was 40 µg/ml);and (4) adipogenic medium (α-MEM supplemented with 10% FBS, 1 mMDEX, 10 mM insulin, 200 mM indomethacin and 0.5 mM IBMX), as apositive control (PC) (Zuk et al., 2002). The medium was changed every3 days. The cells were collected at day 5 or 10 for qPCR analysis or westernblotting. After 10 days in culture, adipogenic differentiation was determinedby Oil Red O (Sigma-Aldrich) staining. Then, Oil Red O in cells wasextracted with 100% isopropanol for 15 min. The absorbance was measuredat 510 nm with a spectrophotometer (MultiskanGO, Thermo Scientific).
Total RNA isolation, small RNA library construction andsequencingTotal RNA from Exo-AT and Exo-AT was isolated using TRIzol reagent(Invitrogen) according to the manufacturer’s instructions. Total RNAwas fractionated on a 15% Tris-borate-EDTA (TBE) polyacrylamide gel(Invitrogen) and a band corresponding to small RNAs (18–30 nt) was excised.Isolated small RNAs were terminal repaired by adding 5′- and 3′-adapter,reverse transcribed into cDNA and amplified. Raw small RNA sequence datawere obtained by using a Illumina HiSeq™2000 machine. After sequencing,the Solexa CHASTITY quality filtered reads were harvested as Clean Reads.The trimmed reads were aligned to miRBase v21 using Novoalign software(v2.07.11) allowing, at most, one mismatch. miRNAs in Exo-AT and Exo-ADSCs were profiled in two biological replicates. Our miRNA profiling datahave been deposited at the Gene Expression Omnibus (GEO) repository, withaccessionnumbers fromGSM2425598–GSM2425601 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE92313). We tested for a differencebetween the Exo-AT and Exo-ADSC with a paired two-sided t-test andcorrected for multiple testing by using the Bonferonni method. Fold changeand P-values were calculated for each miRNA. These P-values were used in
testing the false discovery rate (FDR) values for each miRNA, which wasfurther used as a filter to get significantmiRNAhits with a fold change≥2 andFDR<0.05.We analyzed expression datawithMulti ExperimentView (MEV)cluster software. To identify putative targets of different expressed miRNAs,TargetScan (http://www.targetscan.org/), miRanda (http://www.microrna.org/microrna/) and miRbase (http://www.mirbase.org/) were applied. The genespredicted by all these algorithms were chosen as target genes.
miRNA transfection assayADSCs (5×104 per well) were seeded in 12-well plates and culturedovernight. 500 μl transfection mix (Lipofectamine 3000; Invitrogen) withmiRNA mimic or inhibitor (Ruibo, China) was added into cells andincubated at 37°C for 6 h. The transfection efficiency of mimic (50 nM) andinhibitor (200 nM) was confirmed by qPCR. To explore functions of miR-450a-5p during adipogenesis, ADSCs were transfected with mimic orinhibitor then cultured with 1 ml adipogenic medium (α-MEMsupplemented with 10% FBS, 1 mM DEX, 10 mM insulin, 200 mMindomethacin and 0.5 mM IBMX) for 5 days (fresh adipogenic mediumwere changed every 3 days). RNA and protein were extracted at day 5 afterinduction. To explore the role of miR-450a-5p in Exo-AT-mediatedadipogenesis, ADSCs (5×104 per well, 12-well plates) were transfectedwith miR-450a-5p inhibitor (200 nM), then cultured in 1 ml mediumsupplemented with exosome-like vesicles (40 µg/ml) for 5 days. Themedium was changed every 3 days.
Dual-luciferase reporter assay756 bp of the WISP2 3′UTR region was amplified and cloned into a fireflyluciferase reporter vector (Obio Technology, China), named as pMIR-WISP2-WT. A mutant version of the WISP2 3′UTR reporter plasmid(pMIR-WISP2-MUT) was also generated by mutating the seed region formiR-450a-5p. To evaluate the direct interaction between miR-450a-5p andthe 3′UTR from WISP2, HEK-293 cells were co-transfected with 50 nMmiRNA mimic, 1 μg of firefly luciferase reporter vector (pMIR-WISP2-WTor pMIR-WISP2-MUT) and with 100 ng of Renilla luciferase pRL-CMV vector (Obio Technology, China). Luciferase activity was measuredusing the Dual-Glo luciferase assay system (Promega) at 48 h posttransfection. Normalized firefly luciferase activity was compared betweendifferent groups.
Western blot analysisTo identify exosome-like vesicles, 30 μg vesicles, as determined by the BCAmethod, was dissolved in RIPA Lysis Buffer (KeyGEN, China), resolved on a10% polyacrylamide gel and blotted on to a nitrocellulose membrane.The membranes were blocked and then incubated with primary antibodiesagainst CD9 (1:1000, Zen Bioscience, China, 220642), TSG101 (1:1000,Zen Bioscience, China, 341000), CD63 (1:1000, Zen Bioscience, China,615509) and β-actin (1:1000, Abcam, ab3280) at 4°C overnight, followed byhorseradish peroxidase (HRP)-conjugated secondary antibodies for 1 h atroom temperature. Immobilon Western Chemiluminescent HRP Substrate(Millipore) was used for the detection following the manufacturer’sinstructions. Signals were visualized with a ImageQuant LAS 4000 minimachine (GEHealthcare). To detect adipogenic markers during adipogenesis,total proteinswere extracted from treated cells at various times by using a TotalProtein Extraction Kit (KeyGEN, China). 30 µg cellular protein from eachsample was resolved by SDS-PGAE followed by immunoblotting as above.Primary antibodies against PPARγ2 (Abcam, ab45036), adiponectin (Abcam,ab62551), WISP2 (Zen Bioscience, China, 507456) (all used at a dilution of1:1000) and GAPDH (1:5000, Zen Bioscience, China, 200306-7E4) wereused in this study. Band intensities were determined using ImageJ softwareand normalized to internal control GAPDH.
qPCRTotal cellular RNAs were extracted using RNAiso Plus (TaKaRaBiotechnology) according to the manufacturer’s instruction. The RNAswere transcribed into cDNAs with a First Strand cDNA Synthesis Kit(Thermo Scientific). Specific cDNAs were amplified with SYBR PremixExTaq (TaKaRa Biotechnology, Japan) utilizing an Eco Real-time PCR
1166
RESEARCH ARTICLE Journal of Cell Science (2017) 130, 1158-1168 doi:10.1242/jcs.197764
Journal
ofCe
llScience

System (Illumina). Reaction conditions were: 95°C for 2 min; followed by40 cycles of 95°C for 5 s, 60°C for 30 s. The results were analyzed by usingthe 2−ΔΔCT relative quantitative method, with GAPDH as an internal control.miRNAs from 1 mg vesicles were extracted with an miRNA Isolation kit(OMEGA) according to the manufacturer’s instructions, then transcribedinto cDNA by using an miRcute Plus miRNA First-Strand cDNA SynthesisKit (TIANGEN, China). cDNAs were amplified with an miRcute miRNAqPCRDetection Kit (SYBRGreen) (TIANGEN, China) by utilizing an Ecoreal-time PCR system (Illumina). U6 was used as an internal control. Primersequences are listed in Table S1.
Statistical analysisEach experiment was repeated at least three times. Data are expressed asmean±s.d. Statistical analysis was performed with a paired Student’s t-test.P<0.05 was considered statistically significant.
Competing interestsThe authors declare no competing or financial interests.
Author contributionsConceptualization: M.Y.; Methodology: Y.Z., M.D.; Data curation: M.Y.;Investigation: Y.Z.; Validation: M.D.; Visualization: C.C., Q.T., W.J., H.W.; Writing -original draft: Y.Z.; Writing - review and editing: M.Y.; Supervision: W.T.
FundingThis work was supported by National Natural Science Foundation of China(81300848).
Data availabilitymiRNA profiling data has been deposited at the Gene Expression Omnibus (GEO)repository, with accession numbers from GSM2425598–GSM2425601 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE92313).
Supplementary informationSupplementary information available online athttp://jcs.biologists.org/lookup/doi/10.1242/jcs.197764.supplemental
ReferencesAltadill, T., Campoy, I., Lanau, L., Gill, K., Rigau, M., Gil-Moreno, A., Reventos,J., Byers, S., Colas, E. and Cheema, A. K. (2016). Enabling metabolomicsbased biomarker discovery studies using molecular phenotyping of exosome-likevesicles. PLoS ONE 11, e0151339.
An, J. H., Ohn, J. H., Song, J. A., Yang, J.-Y., Park, H., Choi, H. J., Kim, S. W.,Kim, S. Y., Park, W.-Y. and Shin, C. S. (2014). Changes of microRNA profile andmicroRNA-mRNA regulatory network in bones of ovariectomized mice. J. BoneMiner. Res. 29, 644-656.
Aoki, N., Jin-no, S., Nakagawa, Y., Asai, N., Arakawa, E., Tamura, N., Tamura, T.and Matsuda, T. (2007). Identification and characterization of microvesiclessecreted by 3T3-L1 adipocytes: redox- and hormone-dependent induction of milkfat globule-epidermal growth factor 8-associated microvesicles. Endocrinology148, 3850-3862.
Arner, P. and Kulyte, A. (2015). MicroRNA regulatory networks in human adiposetissue and obesity. Nat. Rev. Endocrinol. 11, 276-288.
Batiz, L. F., Castro, M. A., Burgos, P. V., Velasquez, Z. D., Munoz, R. I.,Lafourcade, C. A., Troncoso-Escudero, P. andWyneken, U. (2015). Exosomesas novel regulators of adult neurogenic niches. Front Cell Neurosci. 9, 501.
Bowers, R. R., Kim, J. W., Otto, T. C. and Lane, M. D. (2006). Stable stem cellcommitment to the adipocyte lineage by inhibition of DNA methylation: role of theBMP-4 gene. Proc. Natl. Acad. Sci. USA 103, 13022-13027.
Braicu, C., Tomuleasa, C., Monroig, P., Cucuianu, A., Berindan-Neagoe, I. andCalin, G. A. (2014). Exosomes as divine messengers: are they the Hermes ofmodern molecular oncology? Cell Death Differ. 22, 34-45.
Campoy, I., Lanau, L., Altadill, T., Sequeiros, T., Cabrera, S., Cubo-Abert, M.,Perez-Benavente, A., Garcia, A., Borros, S., Santamaria, A. et al. (2016).Exosome-like vesicles in uterine aspirates: a comparison of ultracentrifugation-based isolation protocols. J. Transl. Med. 14, 180.
Chen, L., Hou, J., Ye, L., Chen, Y., Cui, J., Tian, W., Li, C. and Liu, L. (2014).MicroRNA-143 regulates adipogenesis by modulating the MAP2K5-ERK5signaling. Sci. Rep. 4, 3819.
Chen, Q., Shou, P., Zheng, C., Jiang, M., Cao, G., Yang, Q., Cao, J., Xie, N.,Velletri, T., Zhang, X. et al. (2016). Fate decision of mesenchymal stem cells:adipocytes or osteoblasts? Cell Death Differ. 23, 1128-1139.
Cho, Y. M., Kim, T. M. and Hun Kim, D. (2016). miR-148a is a downstream effectorof X-box-binding protein 1 that silences Wnt10b during adipogenesis of 3T3-L1cells. Exp. Mol. Med. 48, e226.
Choi, J. S., Yoon, H. I., Lee, K. S., Choi, Y. C., Yang, S. H., Kim, I.-S. and Cho,Y. W. (2016). Exosomes from differentiating human skeletal muscle cells triggermyogenesis of stem cells and provide biochemical cues for skeletal muscleregeneration. J. Control Release 222, 107-115.
Chowdhury, R.,Webber, J. P., Gurney,M., Mason,M. D., Tabi, Z. andClayton, A.(2015). Cancer exosomes trigger mesenchymal stem cell differentiation into pro-angiogenic and pro-invasive myofibroblasts. Oncotarget 6, 715-731.
Connolly, K. D., Guschina, I. A., Yeung, V., Clayton, A., Draman, M. S., VonRuhland, C., Ludgate, M., James, P. E. andRees, D. A. (2015). Characterisationof adipocyte-derived extracellular vesicles released pre- and post-adipogenesis.J. Extracell. Vesicles 4, 29159.
Cui, Y., Luan, J., Li, H., Zhou, X. and Han, J. (2016). Exosomes derived frommineralizing osteoblasts promote ST2 cell osteogenic differentiation by alterationof microRNA expression. FEBS Lett. 590, 185-192.
Delic, D., Eisele, C., Schmid, R., Luippold, G., Mayoux, E. and Grempler, R.(2016). Characterization of Micro-RNA changes during the progression of type 2diabetes in zucker diabetic fatty rats. Int. J. Mol. Sci. 17, E665.
Deng,Z.B., Poliakov,A.,Hardy,R.W.,Clements,R., Liu,C., Liu, Y.,Wang, J., Xiang,X., Zhang,S., Zhuang,X. et al. (2009).Adipose tissueexosome-like vesiclesmediateactivation of macrophage-induced insulin resistance. Diabetes 58, 2498-2505.
Eguchi, A., Lazic, M., Armando, A. M., Phillips, S. A., Katebian, R., Maraka, S.,Quehenberger, O., Sears, D. D. and Feldstein, A. E. (2016). Circulatingadipocyte-derived extracellular vesicles are novel markers of metabolic stress.J. Mol. Med. 94, 1241-1253.
Fleury, A., Hoch, L., Martinez, M. C., Faure, H., Taddei, M., Petricci, E., Manetti,F., Girard, N., Mann, A., Jacques, C. et al. (2016). Hedgehog associated tomicroparticles inhibits adipocyte differentiation via a non-canonical pathway. Sci.Rep. 6, 23479.
Forterre, A., Jalabert, A., Chikh, K., Pesenti, S., Euthine, V., Granjon, A.,Errazuriz, E., Lefai, E., Vidal, H. and Rome, S. (2014). Myotube-derivedexosomal miRNAs downregulate Sirtuin1 in myoblasts during muscle celldifferentiation. Cell Cycle 13, 78-89.
Gustafson, B., Hammarstedt, A., Hedjazifar, S. and Smith, U. (2013). Restrictedadipogenesis in hypertrophic obesity: the role of WISP2, WNT, and BMP4.Diabetes 62, 2997-3004.
Gustafson,B., Hammarstedt,A., Hedjazifar,S.,Hoffmann, J.M., Svensson,P.-A.,Grimsby, J., Rondinone, C. and Smith, U. (2015a). BMP4 and BMP antagonistsregulate human white and beige adipogenesis. Diabetes 64, 1670-1681.
Gustafson, B., Hedjazifar, S., Gogg, S., Hammarstedt, A. and Smith, U. (2015b).Insulin resistance and impaired adipogenesis. Trends Endocrinol. Metab. 26,193-200.
Hamam, D., Ali, D., Vishnubalaji, R., Hamam, R., Al-Nbaheen, M., Chen, L.,Kassem, M., Aldahmash, A. and Alajez, N. M. (2014). microRNA-320/RUNX2axis regulates adipocytic differentiation of human mesenchymal (skeletal) stemcells. Cell Death Dis. 5, e1499.
Hammarstedt, A., Hedjazifar, S., Jenndahl, L., Gogg, S., Grunberg, J.,Gustafson, B., Klimcakova, E., Stich, V., Langin, D., Laakso, M. et al.(2013). WISP2 regulates preadipocyte commitment and PPARγ activation byBMP4. Proc. Natl. Acad. Sci. USA 110, 2563-2568.
He, W. A., Calore, F., Londhe, P., Canella, A., Guttridge, D. C. and Croce, C. M.(2014). Microvesicles containing miRNAs promote muscle cell death in cancercachexia via TLR7. Proc. Natl. Acad. Sci. USA 111, 4525-4529.
Huttala, O., Mysore, R., Sarkanen, J. R., Heinonen, T., Olkkonen, V. M. andYlikomi, T. (2016). Differentiation of human adipose stromal cells in vitro intoinsulin-sensitive adipocytes. Cell Tissue Res. 366:63-74.
Jeon, M., Rahman, N. and Kim, Y.-S. (2016). Wnt/β-catenin signaling plays adistinct role in methyl gallate-mediated inhibition of adipogenesis. Biochem.Biophys. Res. Commun. 479, 22-27.
Jing, W., Xiao, J., Xiong, Z., Yang, X., Huang, Y., Zhou, M., Chen, S., Lin, Y. andTian, W. (2011). Explant culture: an efficient method to isolate adipose-derivedstromal cells for tissue engineering. Artif. Organs 35, 105-112.
Kahlert, C. and Kalluri, R. (2013). Exosomes in tumor microenvironment influencecancer progression and metastasis. J. Mol. Med. 91, 431-437.
Kennell, J. A., Gerin, I., MacDougald, O. A. and Cadigan, K. M. (2008). ThemicroRNA miR-8 is a conserved negative regulator of Wnt signaling. Proc. Natl.Acad. Sci. USA 105, 15417-15422.
Konala, V. B. R., Mamidi, M. K., Bhonde, R., Das, A. K., Pochampally, R. and Pal,R. (2016). The current landscape of the mesenchymal stromal cell secretome: anew paradigm for cell-free regeneration. Cytotherapy 18, 13-24.
Kowal, J., Tkach, M. and Thery, C. (2014). Biogenesis and secretion of exosomes.Curr. Opin. Cell Biol. 29, 116-125.
Krampera, M., Marconi, S., Pasini, A., Galie, M., Rigotti, G., Mosna, F., Tinelli,M., Lovato, L., Anghileri, E., Andreini, A. et al. (2007). Induction of neural-likedifferentiation in human mesenchymal stem cells derived from bone marrow, fat,spleen and thymus. Bone 40, 382-390.
1167
RESEARCH ARTICLE Journal of Cell Science (2017) 130, 1158-1168 doi:10.1242/jcs.197764
Journal
ofCe
llScience

Lee, H. K., Finniss, S., Cazacu, S., Xiang, C. and Brodie, C. (2014). Mesenchymalstem cells deliver exogenousmiRNAs to neural cells and induce their differentiationand glutamate transporter expression. Stem Cells Dev. 23, 2851-2861.
Lee, J.-E., Moon, P.-G., Lee, I.-K. and Baek, M.-C. (2015). Proteomic analysis ofextracellular vesicles released by adipocytes of otsuka long-evans tokushima fatty(OLETF) rats. Protein J. 34, 220-235.
Lehr, S., Hartwig, S., Lamers, D., Famulla, S., Muller, S., Hanisch, F.-G.,Cuvelier, C., Ruige, J., Eckardt, K., Ouwens, D. M. et al. (2012). Identificationand validation of novel adipokines released from primary human adipocytes.Mol.Cell Proteomics 11, M111.010504.
Li, J., Qiao, X., Yu, M., Li, F., Wang, H., Guo, W. and Tian, W. (2014). Secretoryfactors from rat adipose tissue explants promote adipogenesis and angiogenesis.Artif. Organs 38, E33-E45.
Lopatina, T., Bruno, S., Tetta, C., Kalinina, N., Porta, M. and Camussi, G. (2014).Platelet-derived growth factor regulates the secretion of extracellular vesicles byadipose mesenchymal stem cells and enhances their angiogenic potential. CellCommun. Signal. 12, 26.
Martin, P. J., Haren, N., Ghali, O., Clabaut, A., Chauveau, C., Hardouin, P.,Broux, O. (2015). Adipogenic RNAs are transferred in osteoblasts via bonemarrow adipocytes-derived extracellular vesicles (EVs). BMC Cell Biol. 16, 10.
Moseti, D., Regassa, A. and Kim, W. K. (2016). Molecular regulation ofadipogenesis and potential anti-adipogenic bioactive molecules. Int. J. Mol. Sci.17, E124.
Muller, G., Schneider, M., Biemer-Daub, G. and Wied, S. (2011). Microvesiclesreleased from rat adipocytes and harboring glycosylphosphatidylinositol-anchored proteins transfer RNA stimulating lipid synthesis. Cell Signal. 23,1207-1223.
Narayanan, R., Huang, C.-C. and Ravindran, S. (2016). Hijacking the cellular mail:exosome mediated differentiation of mesenchymal stem cells. Stem Cells Int.2016, 3808674.
Park, J.-R., Jung, J.-W., Lee, Y.-S. and Kang, K.-S. (2008). The roles of Wntantagonists Dkk1 and sFRP4 during adipogenesis of human adipose tissue-derived mesenchymal stem cells. Cell Prolif. 41, 859-874.
Qi, X., Zhang, J., Yuan, H., Xu, Z., Li, Q., Niu, X., Hu, B., Wang, Y. and Li, X.(2016). Exosomes secreted by human-induced pluripotent stem cell-derivedmesenchymal stem cells repair critical-sized bone defects through enhancedangiogenesis and osteogenesis in osteoporotic rats. Int. J. Biol. Sci. 12, 836-849.
Qin, L., Chen, Y., Niu, Y., Chen, W., Wang, Q., Xiao, S., Li, A., Xie, Y., Li, J., Zhao,X. et al. (2010). A deep investigation into the adipogenesis mechanism: profile ofmicroRNAs regulating adipogenesis by modulating the canonical Wnt/beta-catenin signaling pathway. BMC Genomics 11, 320.
Qin, Y., Wang, L., Gao, Z., Chen, G. and Zhang, C. (2016). Bone marrow stromal/stem cell-derived extracellular vesicles regulate osteoblast activity anddifferentiation in vitro and promote bone regeneration in vivo. Sci. Rep. 6, 21961.
Rigamonti, A., Brennand, K., Lau, F. and Cowan, C. A. (2011). Rapid cellularturnover in adipose tissue. PLoS ONE 6, e17637.
Romancino, D. P., Paterniti, G., Campos, Y., De Luca, A., Di Felice, V., d’Azzo, A.andBongiovanni, A. (2013). Identification and characterization of the nano-sizedvesicles released by muscle cells. FEBS Lett. 587, 1379-1384.
Rosen, E. D. andSpiegelman, B.M. (2014).What we talk about whenwe talk aboutfat. Cell 156, 20-44.
Ross, S. E., Hemati, N., Longo, K. A., Bennett, C. N., Lucas, P. C., Erickson, R. L.and MacDougald, O. A. (2000). Inhibition of adipogenesis by Wnt signaling.Science 289, 950-953.
Sarkanen, J.-R., Kaila, V., Mannerstrom, B., Raty, S., Kuokkanen, H., Miettinen,S. and Ylikomi, T. (2012). Human adipose tissue extract induces angiogenesisand adipogenesis in vitro. Tissue Eng. Part A 18, 17-25.
Spalding, K. L., Arner, E., Westermark, P. O., Bernard, S., Buchholz, B. A.,Bergmann, O., Blomqvist, L., Hoffstedt, J., Naslund, E., Britton, T. et al.(2008). Dynamics of fat cell turnover in humans. Nature 453, 783-787.
Sun, D., Zhuang, X., Zhang, S., Deng, Z.-B., Grizzle, W., Miller, D. and Zhang, H.-G. (2013). Exosomes are endogenous nanoparticles that can deliver biologicalinformation between cells. Adv. Drug Deliv. Rev. 65, 342-347.
Takeda, Y. S. and Xu, Q. (2015). Neuronal differentiation of human mesenchymalstem cells using exosomes derived from differentiating neuronal cells. PLoS ONE10, e0135111.
Valadi, H., Ekstrom, K., Bossios, A., Sjostrand, M., Lee, J. J. and Lotvall, J. O.(2007). Exosome-mediated transfer of mRNAs and microRNAs is a novelmechanism of genetic exchange between cells. Nat. Cell Biol. 9, 654-659.
Williams, M. D. and Mitchell, G. M. (2012). MicroRNAs in insulin resistance andobesity. Exp. Diabetes Res. 2012, 484696.
Wu, H.-T., Chen, W., Cheng, K. C., Yeh, C.-H., Shen, K.-H. and Cheng, J.-T.(2010). Indomethacin activates peroxisome proliferator-activated receptor gammato improve insulin resistance in cotton pellet granuloma model. Horm Metab. Res.42, 775-780.
Xie, H., Lim, B. and Lodish, H. F. (2009). MicroRNAs induced during adipogenesisthat accelerate fat cell development are downregulated in obesity. Diabetes 58,1050-1057.
Xu, J.-F., Yang, G. H., Pan, X.-H., Zhang, S.-J., Zhao, C., Qiu, B.-S., Gu, H.-F.,Hong, J.-F., Cao, L., Chen, Y. et al. (2014). Altered microRNA expression profilein exosomes during osteogenic differentiation of human bone marrow-derivedmesenchymal stem cells. PLoS ONE 9, e114627.
Yang, L., Cheng, P., Chen, C., He, H.-B., Xie, G.-Q., Zhou, H.-D., Xie, H., Wu, X. P.and Luo, X. H. (2012). miR-93/Sp7 function loop mediates osteoblastmineralization. J. Bone Miner. Res. 27, 1598-1606.
Yu, S., Yerges-Armstrong, L. M., Chu, Y., Zmuda, J. M. and Zhang, Y. (2013).E2F1 effects on osteoblast differentiation andmineralization aremediated throughup-regulation of frizzled-1. Bone 56, 234-241.
Zhang, Y., Yu, M. and Tian,W. D. (2016). The physiological and pathological impactof exosomes from adipose tissue. Cell Prolif. 49, 3-13.
Zuk, P. A., Zhu, M., Ashjian, P., De Ugarte, D. A., Huang, J. I., Mizuno, H.,Alfonso, Z. C., Fraser, J. K., Benhaim, P. and Hedrick, M. H. (2002). Humanadipose tissue is a source of multipotent stem cells.Mol. Biol. Cell 13, 4279-4295.
1168
RESEARCH ARTICLE Journal of Cell Science (2017) 130, 1158-1168 doi:10.1242/jcs.197764
Journal
ofCe
llScience