microevolution in ï¬mh gene of mucosa-associated
TRANSCRIPT

Microevolution in fimH Gene of Mucosa-Associated Escherichia coliStrains Isolated from Pediatric Patients with Inflammatory BowelDisease
Valerio Iebba,a Maria Pia Conte,a Maria Stefania Lepanto,a Giovanni Di Nardo,b Floriana Santangelo,a Marina Aloi,b Valentina Totino,a
Monica Proietti Checchi,a Catia Longhi,a Salvatore Cucchiara,b and Serena Schippaa
Microbiology Unit, Department of Public Health Sciences, Sapienza University of Rome, Rome, Italy,a and Pediatric Gastroenterology and Liver Unit, Department ofPediatrics, Sapienza University of Rome, Rome, Italyb
Several studies reported increased numbers of mucosa-associated Escherichia coli strains in patients with inflammatory boweldisease (IBD), encompassing Crohn’s disease (CD) and ulcerative colitis (UC). The majority of E. coli strains possess type 1 fim-briae, whose tip fibrillum protein, FimH, naturally undergoes amino acid replacements, an important process in the adaptationof commensal E. coli strains to environmental changes, like those observed in IBD and urinary tract infections. In this study, weanalyzed mutational patterns in the fimH gene of 52 mucosa-associated E. coli strains isolated from IBD and non-IBD pediatricpatients, in order to investigate microevolution of this genetic trait. FimH-positive strains were also phylogenetically typed andtested for their adhesive ability on Caco-2 cells. Specific FimH alleles for each grouping feature were found. Mutations G66S andV27A were related to CD, while mutations A242V, V163A, and T74I were attributed to UC. Otherwise, the G66S, N70S, and S78Nmutations were specifically attributed to B2/D phylogroups. The N70S and A119V mutations were related to adhesive E. colistrains. Phylogroup B2, adhesive, and IBD E. coli strains showed a higher site substitution rate (SSR) in the fimH gene, togetherwith a higher number of mutations. The degree of naïve mucosal inflammation was related to specific FimH alleles. Moreover,we could suggest that the V27A mutation is pathoadaptive for the CD intestinal habitat, while we could also suggest that both theN70S and S78N mutations are related to the more virulent E. coli B2 phylogroup. In conclusion, we found some FimH variantsthat seem to be more involved than others in the evolution of IBD pathogenesis.
The inflammatory bowel disease (IBD) spectrum, of whichCrohn’s disease (CD) and ulcerative colitis (UC) are the two
main phenotypes (21), is the result of a complex interaction be-tween elements such as host susceptibility, mucosal immunity,and the intestinal milieu. Several lines of evidence support thehypothesis that intestinal bacteria play a role in the pathogenesisof adult IBD and pediatric IBD (6, 12, 18, 32, 36, 37, 42). Increasednumbers of mucosa-associated Escherichia coli isolates were ob-served in both CD and UC (17, 25). The E. coli dominant genotypeassociated with strains isolated from IBD patients was also de-scribed. In addition, a new E. coli pathotype referred to as adher-ent-invasive E. coli (AIEC) has been recognized (2, 4, 5, 13, 14, 29,30). Strong adherence of bacteria to surfaces is an essential pre-liminary stage in colonization (24, 27) and an integral and essen-tial step in infection. The adherence is mediated by structurescalled adhesins (28). E. coli expresses a number of different adhe-sive organelles, including P, type 1, S, F1C, and long polar fimbriae(1, 8, 22). A mean of 95% of all isolates of E. coli express type 1fimbriae, also called mannose-sensitive fimbriae or, commonly,fimbriae. It was reported how pilus type 1 is the major factorresponsible for the enhanced adhesive and invasive properties ofE. coli (3, 7, 26, 39). The type 1 pilus consists of five parts. They are,starting from the cellular body, FimD (inserted in the outer mem-brane), FimA (pilus rod), and FimF, FimG, and FimH (tip fibril-lum). Recently, it has become clear that pilus type 1 exhibits sev-eral different phenotypes, due to allelic variation of the genes fimAand fimH, and that these phenotypes are differentially distributedamong E. coli isolates (15, 33). The 273-amino-acid-long FimHprotein is structured in two main domains: the mannose-bindinglectin domain (Ld; residues 1 to 156) and the fimbria-incorporat-
ing pilin domain (Pd; residues 160 to 273), which are connectedvia a 3-amino-acid interdomain linker peptide chain (residues 157to 159) (31). Naturally occurring phenotypic variants of the FimHprotein have recently been recognized among intestinal and uro-pathogenic E. coli strains (10). More precisely, these natural vari-ations confer stronger monomannose and three-mannose bind-ing (33), as seen in urinary tract infections, depending on the shearstress which E. coli cells undergo (1, 43). Thus, FimH mutationsmay be adaptive in secondary habitats of E. coli. The aim of thiswork was to study the fimH gene (903 nucleotides) mutationalpatterns of 52 mucosa-associated E. coli strains isolated from pe-diatric patients diagnosed with IBD and those without IBD, inorder to evaluate potential associations between fimH mutationalprofiles and particular features, such as disease status, E. coli phy-logroup, and in vitro static adhesion levels of the isolated strains.
Received 21 November 2011 Returned for modification 1 January 2012Accepted 13 January 2012
Published ahead of print 30 January 2012
Editor: S. M. Payne
Address correspondence to Valerio Iebba, [email protected].
V.I. and M.P.C. contributed equally to this article. S.C. and S.S. contributed equallyto this article.
Supplemental material for this article may be found at http://iai.asm.org/.
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/IAI.06181-11
1408 iai.asm.org 0019-9567/12/$12.00 Infection and Immunity p. 1408–1417
on April 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

MATERIALS AND METHODSPatients. Thirty-eight pediatric patients (age range, 8 to 17 years) referredto the Pediatric Gastroenterology and Liver Unit of the Sapienza Univer-sity of Rome, Italy, for suspected IBD were studied: active CD was diag-nosed in 12, and active UC was diagnosed in 7. The remaining 19 subjectswith functional gastrointestinal disorders (undetermined colitis, lympho-nodular hyperplasia) and normal colonoscopy and histology findingsserved as controls. As reported in Table S1 in the supplemental material,the baseline demographic characteristics were similar in the three groups.The patient groups did not differ significantly by age and disease duration.All children with CD had ileocolonic involvement, and all had diseaseactivity in the moderate to severe range. All UC patients had endoscopicevidence of pancolitis, showing a backwash ileitis, and the disease wasclassified severe. The diagnostic workup of UC and CD was done accord-ing to international protocols. Children with CD were assessed using thePediatric Crohn’s Disease Activity Index (PCDAI), which is a multi-itemscore based on recall of the preceding week’s symptoms, laboratory pa-rameters (erythrocyte sedimentation rate, hematocrit and albumin lev-els), and physical examination. A score of 10 implies inactive disease, oneof 11 to 30 implies mild disease, and one of �30 implies moderate tosevere disease. It is noteworthy that our CD pediatric patients did not havea previous endoscopic assessment (see Table S1 in the supplemental ma-terial), nor any kind of treatment (sulfasalazine was given to 2/12 CDpatients after biopsy sampling for this study), so their naïve mucosal in-flammation reflected the actual selective pressure on the E. coli strainsisolated. Patients with UC were evaluated using the Pediatric UlcerativeColitis Activity Index (PUCAI), which is based only on clinical symptoms.A score of 10 indicates inactive disease, one of 11 to 34 implies milddisease, one of 35 to 64 implies moderate disease, and one of �65 impliessevere disease. Only one UC patient out of seven received a previoustreatment with steroids as enema; thus, also for the UC cohort, the naïvemucosal inflammation reflected a naïve habitat from which E. coli strainswere isolated. All patients underwent ileocolonoscopy after parental in-formed written consent was provided. During ileocolonoscopy, two bi-opsy samples were taken from each region (ileum and descending colon)for routine histological assessment and bacteriological study. Specimenswere collected in 2-ml screw-cap vials filled with 0.85 ml of brain heartinfusion broth (Oxoid, Cambridge, United Kingdom) and 0.15 ml ofglycerol (Sigma-Aldrich, St. Louis, MO) and immediately stored at�80°C.
Treatment of biopsy specimens and bacterial culture. For E. coli spe-cies isolation, biopsy specimens (15 mg each) from CD, UC, and controlpatients and from different regions (ileum and colon) were first washed in500 �l of physiologic saline with 0.016% dithioerythritol to remove themucus and then shaken 3 times in 500 �l of physiological saline for 30 s.After a fourth wash, the biopsy specimens were hypotonically lysed byvortexing for 30 min in 500 �l distilled water. One hundred microliters ofthe cell debris left after hypotonic lysis was plated in 10-fold dilution stepsonto MacConkey agar. After 24 h of incubation at 37°C, all bacterial col-onies were isolated and subcultured onto nutrient agar and successivelyidentified by use of the API 32 ID system (bioMérieux, Milan, Italy), inorder to recognize and isolate E. coli species. Sixty E. coli strains wererandomly chosen (2 to 3 from each patient), taking into account bothbiopsy specimens (from the ileum and descending colon). E. coli strainsfrom controls were retrieved only from descending colon biopsy speci-mens.
Amplification of E. coli fimH gene. Total DNA from E. coli strains wasobtained by putting a single colony picked up from a petri dish in a 0.2-mltube containing 50 �l of 0.22-�m-pore-size-filtered and autoclaved dou-ble-distilled H2O. A subsequent cell-disrupting step was achieved by dis-ruption for 10 min at 95°C in an Eppendorf thermocycler, followed by a4°C step and a centrifugation step for 10 min at 10,000 rpm. Supernatantcontaining genomic and plasmid DNA was collected in new 0.2-ml tubesand stored at �20°C until PCR amplification. The fimH gene (903 bp) wasamplified in a 25-�l-final-volume PCR mixture containing 1� Phusion
High-Fidelity PCR master mix with HF buffer (New England BioLabs,Ipswich, MA), primers AdH_FWnoGC (5=-ATGAAACGAGTTATTACCCTGTTTGCTG-3=) and Pil_RW (5=-ATTGATAAACAAAAGTCACGCCAATAATCG-3=) at 0.5 �M, and 3 �l of supernatant, used as DNA tem-plate. Run conditions were as follows: 95°C for 5 min; 25 cycles at 94°C for20 s, 59°C for 20 s, and 72°C for 40 s; and then a final step at 72°C for 1 min.The whole PCR mixture was run on a 1% agarose gel, and the resultingband was excised and purified with a MinElute gel extraction kit (Qiagen,Hilden, Germany) following the manufacturer’s instructions. The elutedDNA was sequenced in both directions by the MWG-Biotech (Eurofins)service to have coverage of the entire fimH gene. Electropherograms for allsingleton mutations were also inspected through Geneious (version 4.8.3)software (Biomatters Ltd.) for consistency between strands, and any am-biguous nucleotides were resolved by resequencing.
Analysis of fimH gene sequences. Upon receipt, sequences underwenta BLAST search (www.ncbi.nih.gov/blast) to confirm that they belongedto the fimH gene of E. coli and were then aligned with Geneious (version4.8.3) software (Biomatters Ltd.) for DNA mutational pattern analysis.The fimH gene sequence of E. coli strain MG1655 was taken as the refer-ence one (GeneID 948847). The silent and nonsilent mutations weretransformed in a binary matrix of presence/absence of such point muta-tions, so as to have a comprehensive view of mutational patterns. Wesubaligned all 52 fimH sequences, plus those of strains MG1655 and LF82,according to status (CD, UC, control), phylogenetic group (A, B1, B2, D),and adhesion class (class 1 and class 2), performing various match-by-match analyses on subalignments.
(i) Mutational pattern analysis. A nonsupervised method, agglomer-ative hierarchical clustering (AHC), was made to test the clustering be-havior of the nucleotide and amino acid sequences of the 52 strains plusthose of MG1655 and LF82. Dendrograms were generated on mean-cen-tered and variance-scaled data by means of the partial least-squares dis-criminant analysis (PLS-DA) algorithm implemented in Simca-P� soft-ware (Umetrics), taking into account Euclidean distances and theagglomeration method of Ward. PSL-DA is a regression analysis that cor-relates a number of variables x (point mutations) to a set of variables y(fimH sequences), obtaining the covariance between x and y. The PSL-DAapproach creates a predictive model, validated through Fisher’s test, use-ful to classify E. coli strains according to the mutational patterns in thefimH gene. The two-tailed �2 test was used to discover whether the distri-bution of E. coli strains differed between the clusters identified by thedendrogram. A P value of less than or equal to 0.05 was considered statis-tically significant.
(ii) Discrimination of nucleotides and amino acids. PLS-DA was alsoused to find nucleotide and amino acid mutations that made a majorcontribution in dividing subalignments. Data were automatically meancentered and unit variance (UV) scaled by the statistical software. Eachnucleotide mutation was hierarchically classified on the basis of a soft-ware-assigned variable importance (VIP) value. The variables with VIPvalues of �1 were chosen as being discriminatory.
(iii) Transitions, transversions, and dN/dS ratio. Calculation oftransitions (AG/GA, CT/TC) and transversions was performed on eachsubalignment by means of the on-line software Los Alamos National Labo-ratory Highlighter tool (http://hcv.lanl.gov/content/sequence/HIGHLIGHT/highlighter.html), to visualize transition and transversion mutations in a setof nucleotide sequences that are aligned and in frame. Computation of silent(synonymous [dS]) and nonsilent (nonsynonymous [dN]) mutations, alongwith the dN/dS ratio, was performed on each subalignment by means of theonline software Los Alamos National Laboratory SNAP tool (http://hcv.lanl.gov/content/sequence/SNAP/SNAP.html), useful to give a quantificationof positive selection for pathoadaptive changes in the fimH gene. A dN/dSratio of �1 indicates purifying or neutral selection favoring amino acidconservation, while a dN/dS ratio of �1 indicates positive selection favor-ing amino acid substitutions (20, 38). To assess differences between dN/dSratios, a two-tailed t test was performed on means and standard deviations
Microevolution of fimH Gene in IBD Pediatric Patients
April 2012 Volume 80 Number 4 iai.asm.org 1409
on April 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

(SDs), and a P value of less than or equal to 0.05 was considered statisti-cally significant.
(iv) Dice index. The Dice similarity index (mean percent � SD) wascalculated within each subalignment to assess intersequence similarity bythe formula (2nAB)/(nA � nB), where nA is the total number of nucleo-tide mutations in pattern A, nB is the total number of nucleotide muta-tions in pattern B, and nAB is the number of nucleotide mutations com-mon to patterns A and B. Next, a mean intragroup Dice index wascalculated in order to compare all patient groups. A two-tailed t test wasperformed to find differences, and a P value of �0.05 was consideredstatistically significant.
(v) Phylogenetic tree. Phylogenetic analysis of the fimH gene sequence(799 bp) was achieved by means of Geneious (version 4.8.3) software(Biomatters Ltd.), and an unconstrained-branch-length Bayesian phylo-genetic tree was a posteriori constructed after generation of 5,501 raw treeswith nucleotide substitution model JC96 and a gamma-rate variation. Thesite substitution rate (SSR) was also computed for each fimH sequence,taking the sum of distances from the root. The two-tailed �2 test was usedto assess differences between SSR groups belonging to a defined subalign-ment, and a P value of �0.05 was considered statistically significant.
(vi) Clustered image mapping (CIM). Cross-correlations of fimHmutational patterns between subalignments were obtained by the onlinesoftware CIMminer (http://discover.nci.nih.gov/cimminer), based on ascaled and centered set of data on the presence/absence of nucleotidemutations. Weight coefficients were computed by the PLS-DA algorithmwith Simca-P� software (Umetrics) for each mutated nucleotide (70 xvariables) on each classification group (9 y variables). x and y variableswere two-dimensionally clustered on the basis of the Euclidean distancedissimilarity matrix and the agglomeration method of Ward.
Static adhesion assay. Monolayers of differentiated Caco-2 (humancolonic adenocarcinoma) cells were cultured in 24-well plates in mini-mum essential medium (MEM; Euroclone, Milan, Italy) supplementedwith NaHCO3 at 1.2 g/liter, 2 mM glutamine, penicillin at 100 IU/ml,streptomycin at 0.1 mg/ml, and 10% heat-inactivated fetal calf serum(Euroclone) in a 5% CO2 incubator. For static adhesion assays, the cellswere seeded at a density of 2 � 104 cells/well and cultured to completedifferentiation for 15 days before infection. For maximal fimbrial expres-sion, bacterial colonies were grown overnight in nutrient agar, resus-pended in sterile saline solution, and left for 48 h at room temperature.Each monolayer was infected with 1 � 108 bacteria per well. After 2 h ofincubation at 37°C, monolayers were washed 3 times with phosphate-buffered saline (PBS), cells were lysed with 0.1% Triton X-100, and thenumbers of CFU were determined by plating. For each experiment, themean number of Caco-2 cells after 15 days of culture was also determined.Bacterial adhesion was defined as the percentage of attached bacteria com-pared with the initial inoculum, which was taken to be 100%, and a cutoffvalue equal to or greater than 0.8% was used to define a strain as adhesive(class 2), while a cutoff value of less than 0.8% defined a strain as nonad-hesive (class 1). Adhesion assays were performed in triplicate.
Phylogenetic PCR grouping. The phylogenetic groups of E. colistrains were identified using a multiplex PCR, as already described (11).The PCR products were 279 bp for the chuA gene, 211 bp for the yjaA gene,and 152 bp for the TspE4C2 fragment. Phylogenetic grouping was madeon the basis of the presence of specific PCR-amplified fragments: group Awas chuA negative, yjaA positive or negative, and TspE4C2 negative;group B1 was chuA negative, yjaA positive or negative, and TspE4C2 pos-itive; group B2 was chuA positive, yjaA positive, and TspE4C2 positive ornegative; and group D was chuA positive, yjaA negative, and TspE4C2positive or negative.
RESULTSMucosa-associated E. coli strain characterization. A total of 60mucosal E. coli strains were isolated by cultural techniques: 28from patients with CD, 19 from patients with UC, and 13 fromnon-IBD controls. The prevalence of E. coli in each cohort was
83% in CD patients (10/12), 100% in UC patients (7/7), and 53%in controls (10/19) (�2 � 6.35, P � 0.04). The E. coli bacterial loadvaried from patient to patient and ranged from 1 � 104 to 1 � 106
CFU/ml in IBD patient specimens and from 0 to 1 � 103 CFU/mlin control specimens. Higher bacterial concentrations were foundin biopsy specimens from patients with IBD. Among all isolates ofthe E. coli strains, 52/60 (86.7%) were positive for the fimH geneand underwent further characterization. The adhesive propertiesof the fimH-positive E. coli isolates were investigated in the Caco-2cell model. The mean adhesion rate of the referent nonadhesivestrain, E. coli DH5�, was 0.4% � 0.3% of the original inoculum,whereas the referent adhesive strain enteropathogenic E. coli32O55 had a mean adhesion index of 2.7% � 0.7%. E. coli strainstested in this study were considered adherent when the mean ad-hesion index was 0.8% or more of the original inoculum. Table S2in the supplemental material reports the results of the E. coli ad-hesion assays. E. coli strains were separated into two adhesionclasses. Class 1, with adhesion levels ranging from 0 to 0.8% (non-adhesive strains), included 16 strains: 8/23 (34.8%) from patientswith CD, 2/16 (12.5%) from patients with UC, and 6/13 (46.2%)from controls. Class 2, with adhesion levels of �0.8% (adhesivestrains), comprehended 36 strains: 15/23 (65.2%) from patientswith CD, 14/16 (87.5%) from patients with UC, and 7/13 (53.8%)from controls. The strains isolated from CD patients and controlswere randomly distributed between the two classes, while thestrains from patients with UC were significantly present in adhe-sion class 2 (�2 � 15.1, P � 0.0001). Table S2 in the supplementalmaterial also reports the phylogroup distributions among the pa-tients. We decided, supported by literature data (23), to join to-gether phylogenetic groups B2 and D, typically shown by virulentE. coli types. Therefore, phylogenetic groups A and B1 were joined,too. The B2/D group included 29 strains: 15/23 (65.2%) frompatients with CD, 9/16 (56.2%) from patients with UC, and 5/13(38.5%) from controls. The A/B1 group included 23 strains: 8/23(34.8%) from patients with CD, 7/16 (43.7%) from patients withUC, and 8/13 (61.5%) from controls. The four phylogroups wererandomly distributed among the cohorts (�2 � 2.41, P � 0.21),and, in particular, the B2 group was exclusively present, but notsignificantly, in overall IBD strains.
FimH protein harbors characteristic mutations shaped fromselective habitats. All 52 FimH sequences were compared to the E.coli MG1655 FimH sequence, and we observed 15 distinct FimHprotein allelic variants, but only alleles retrieved two or moretimes were reported (Table 1). FimH sequences did not differ bysource (ileum or descending colon), as assessed by the Wilcoxonsigned-rank test on the mean intragroup Dice index (P � 0.145),meaning a high intrapatient similarity of FimH protein sequence.By PLS-DA, performed on FimH sequences, we generated predic-tive models based on different grouping features, as stated in Ma-terials and Methods. The patients’ status was well predicted(78.5%, Fisher’s P � 5.9 � 10�8), with G66S and V27A mutationsspecifically ascribed to the CD cohort, while A242V, V163A, andT74I mutations were attributed to the UC cohort (Fig. 1A). Mu-tation Y195F was considered predictive for controls (datum notshown). E. coli phylogroups (A/B1 and B2/D) were correctly pre-dicted by FimH sequences (85%, Fisher’s P � 5.1 � 10�4), withthe G66S, N70S, and S78N mutations specifically being attributedto B2/D, while A119V was ascribed to A/B1 (Fig. 1B). Finally, E.coli adhesion class 2, as stated in Materials and Methods, wasproperly predicted (81.7%, Fisher’s P � 9.2 � 10�3) by amino
Iebba et al.
1410 iai.asm.org Infection and Immunity
on April 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

acid sequences, with N70S and A119V mutations (Fig. 1C) beingdistinctive for the adhesive class. The nonadhesive class (class 1)gave no significant results (data not shown). By PLS-DA, differentFimH mutational patterns were found for each E. coli character-istic (status, phylogroup, adhesion class); thus, it could be hypoth-esized that, under specific selective pressure, the FimH proteinundergoes selective amino acid mutations.
FimH protein mutations arise from a fine-tuned microevo-lution on different parts of fimH gene. In order to evaluate mi-croevolution in the fimH gene, which is responsible for selectiveamino acid mutations, we analyzed all 52 gene sequences col-lected. Among the 52 isolates, 26 distinct fimH allelic variants wereobserved, with 72 unique single nucleotide polymorphisms(SNPs) found at 70 polymorphic sites. Fourteen of the 72 SNPs(19.4%) were singletons (observed in only one fimH type) with 9amino acid replacements; thus, 64.3% of singletons had pheno-typic effects. All 72 mutations were point substitutions; 17(23.6%) were transversions, and 55 (76.4%) were transitions.More precisely, we found 33 C/T transitions (45.8%), 22 A/G
transitions (30.6%), and 17 transversions (23.6%). The codon dis-tribution of the 72 SNPs was 11/72 (15.3%) in the 1st base, 14/72(19.4%) in the 2nd base, and 47/72 (65.3%) in the 3rd base. Out of72 mutations, 21 resulted in amino acid replacements and 51 weresilent substitutions. Among the amino acid replacements, onlyfour were caused by transversion.
fimH sequences did not differ by source (ileum or descendingcolon), as assessed by the Wilcoxon signed-rank test on the meanintragroup Dice index (P � 0.362), meaning a high intrapatientsimilarity of the fimH gene sequence. By means of Dice index (seesupplemental material and Fig. S2 in the supplemental material)we found that CD and UC strains had roughly 50% similarity infimH mutational profiles, while controls had only 36% (Mann-Whitney U test, P � 0.0001). In addition, B2 phylogroup fimHsequences were similar at 63%, against roughly 47% of the otherphylogroups (Mann-Whitney U test, P � 0.0001).
To find out similarity patterns in the fimH gene, we generatedhierarchical dendrograms by PLS-DA analysis, based on differentgrouping features, as stated in Materials and Methods. As in the
TABLE 1 FimH alleles of the pediatric E. coli isolates
Allelea
No. (%) of strains Mutation at amino acid residue:
CD(n � 23)
UC(n � 16)
Controls(n � 13)
B2/D(n � 29)
A/B1(n � 23)
Class 2(n � 35)
Class 1(n � 16) V27 G66 N70 T74 S78 A119 V163 Y195 A202 A242
1 (n � 16) 10 (43.5) 2 (12.5) 4 (30.8) 11 (37.9) 5 (21.7) 6 (17.1) 10 (62.5) A2 (n � 12) 4 (17.4) 6 (37.5) 2 (15.4) 4 (13.8) 8 (34.8) 11 (31.4) 1 (6.2) A V3 (n � 3) 3 (13.0) 0 0 3 (10.3) 0 3 (8.6) 0 A S S N4 (n � 2) 2 (8.7) 0 0 2 (6.9) 0 2 (5.7) 0 A S V5 (n � 3) 1 (4.3) 1 (6.2) 1 (7.7) 2 (6.9) 1 (4.3) 3 (8.6) 0 A S N6 (n � 2) 0 2 (12.5) 0 2 (6.9) 0 1 (2.9)b 0 A S I N A7 (n � 3) 0 3 (18.7) 0 2 (6.9) 1 (4.3) 2 (5.7) 1 (6.2) A N V8 (n � 2) 0 0 2 (15.4) 1 (3.4) 1 (4.3) 1 (2.9) 1 (6.2) A Fa Alleles are defined as combinations of amino acid mutations. Only alleles retrieved two or more times were reported.b Strain 41, harboring FimH allele 6, was cytotoxic for Caco-2 cells, so it was not referred to any adhesion class.
FIG 1 Discriminating amino acid mutations in FimH protein. A three-dimensional graphical representation of the low-affinity state of FimH protein (ProteinData Bank accession number 3JWN) showed amino acid residues with the power to discriminate a patient’s status (A) (red, CD; blue, UC), the E. coli phylogroup(B) (red, B2/D; blue, A/B1), and the E. coli for adhesion capability (C) (red, class 2). The donor strand of the FimG protein, which incorporates into the pilin�-barrel domain, is depicted in green.
Microevolution of fimH Gene in IBD Pediatric Patients
April 2012 Volume 80 Number 4 iai.asm.org 1411
on April 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

case of the PLS-DA based on protein sequences, fimH mutationalpatterns were able to predict the characteristics of fimH-harboringE. coli strains (see supplemental material and Fig. S1 in the sup-plemental material). Mutated nucleotides with discriminatorypower are reported in Table 2. Nucleotide mutation T80C, corre-sponding to the amino acid variation V27A, was distinctive for UCand for the overall IBD group (Fisher’s P � 1.1 � 10�11), whileA209G (corresponding to FimH mutation N70S) and G233A(corresponding to FimH mutated residue S78N) were both re-lated to the B2 phylogroup (Fisher’s P � 3.6 � 10�8). These re-sults could suggest a pathoadaptive effect of the V27A mutationon E. coli fitness to the inflamed intestinal habitat, while the com-bination of N70S and S78N was linked to the phylogroup whichtypically presents virulent E. coli types (11, 23).
The latter analysis provided only fimH nucleotides with ahigher power to divide E. coli features, such as status, phylogroup,and adhesion class, but it gave no information about the fine-tuned modulation of the fimH gene. Trying to understand theunderlying molecular strategies used by E. coli to microevolve thefimH gene under different conditions, we computed the meannumbers of transitions and transversions and the dN/dS ratio, asreported in Table 3. The mean dN/dS ratio was always less than 1,indicating a purifying selection (34) on the overall fimH gene,although we found a higher mutation number in specific hot-spotsites (T80C, T144C, T255G, C276G, T471C, A540G, T651A/C,A654G, G744A, T768C), in which we retrieved a mutation morethan 50% of the time. Ninety percent (9/10) of these hot spotswere silent, due to their presence in the 3rd base; thus, a positiveselection (a dN/dS ratio of more than 1) for the overall fimH genewould be masked, even if a selective pressure acted on specificnucleotides. It was noteworthy that fimH from patients with UCshowed higher mean values for all parameters, especially for thedN/dS ratio, in comparison to fimH from patients with CD (t �2.327, P � 0.0218) and controls (t � 2.425, P � 0.0253). More-over, adhesive strains (class 2) had a significantly higher dN/dSratio than nonadhesive ones (t � �6.252, P � 0.0001), togetherwith an average number of mutations. These results could suggesta tendency for E. coli strains from patients with IBD, mainly frompatients with UC, to enhance adhesion by increasing the numberof mutations and the number of nonsynonymous substitutionsper nonsynonymous site (dN). In fact, UC strains were signifi-cantly present in adhesion class 2 (�2 � 15.1, P � 0.0001), asindicated above. Interestingly, the fimH gene underwent a do-
main-specific mutational pattern (see Fig. S3 in the supplementalmaterial). The lectin domain (nucleotides 1 to 468) showed ahigher number of mutations (�2 � 32.33, P � 0.0001) and hotspots of C/T transitions and transversions compared with the pilindomain (nucleotides 478 to 902). The latter had a tendency toharbor A/G transitions rather than transversions (�2 � 10.33, P �0.008).
In order to have a snapshot of such fimH mutational profiles,we calculated how well each one of the 70 polymorphic sites wasimportant in predicting E. coli habitat source (CD, UC, or con-trol). We performed a cross-correlation between the patients’ sta-tus and nucleotide mutations by means of CIMminer online soft-ware, based on coefficient loadings collected from Simca-P�software (Umetrics) (Fig. 2). The existence of particular red zones(coefficient values higher than 5.0) within each status group evi-denced specific mutational patterns as a result of fine-tuned mi-croevolution in different regions of the fimH gene. It was notewor-thy that, within each red zone, CD E. coli strains had a fimHmutability biased toward both C/T transitions (47.4%) and trans-versions (31.6%), while UC strains had a preponderance of C/Ttransitions (61.5%) (�2 � 8.78, P � 0.003). These results couldsuggest that the intestinal habitat of CD, UC, and control patientsexerted a status-specific stress-response on the fimH gene of E.coli, thus leading to a specific mutagenic profile.
fimH gene from B2 and CD E. coli strains has enhanced mu-tational rate. Trying to understand the fimH gene mutability, wethen calculated the site substitution rate (SSR) for all fimH se-quences, and a Bayesian phylogenetic tree was generated (Fig. 3).Three main nucleotide (NT) clusters were obtained: NT1 (encom-passing NT1a and NT1b; mean SSR � 0.712 � 0.028), NT2 (meanSSR � 0.567 � 0.017), and NT3 (mean SSR � 0.236 � 0.018).Differences in mean SSR values between the three clusters were allsignificant (P � 0.0001). A fourth cluster (strains 1, 14, and 21)was considered an outgroup. FimH mutation V27A was a com-mon background for all clusters, and it was the first to appear inthe phylogenetic tree (Fig. 3). Then, two separate clades evolvedthe mutations A119V (cluster NT2) and A202V (cluster NT1b),while the ultimate clade raised FimH mutations N70S and S78N(cluster NT1a). We also analyzed the percent distribution of E. colistrains in the phylogenetic tree in relation to the aforementionedclassification criteria (status, phylogroup, adhesion class). CDstrains were significantly present in NT1 (mainly NT1b; �2 � 5.12,P � 0.024), while UC strains were equally distributed. Consider-ing the UC and CD strains as an IBD group, we found a significantpresence (43.5%) of IBD strains in NT1 and NT2, which hadhigher SSRs than NT3 (�2 � 4.82, P � 0.0282). In addition, 57%of E. coli phylogroup B2/D strains were present in cluster NT1
TABLE 2 Mutated nucleotides with discriminatory power highlightedby PLS-DA
Grouping feature Discriminating NT mutations (PLS-DA)a
DiseaseCD C369T, C426TUC T80C (V27A), T471CControls C525T
Phylogenetic groupA/B1 T255G, C276GB2 G345T, A249T, A209G (N70S), G233A (S78N)D G639C, C408T
Adhesion class 2 T144C, T471Ca Underlined mutations are nonsilent, mutations between parentheses are thecorresponding FimH mutated amino acids, and mutations in bold are transversions.
TABLE 3 Mutation parameters for each subalignmenta
Feature
Total no.ofmutations
No. ofAG/GAtransitions
No. oftransversions
No. ofCT/TCtransitions
dN/dS(mean � SD)
CD 13.78 3.48 2.87 7.43 0.0580 � 0.0204UC 15.25 4.00 3.25 8.00 0.0761 � 0.0296Control 11.23 3.31 2.69 5.23 0.0526 � 0.0232A/B1 12.22 3.22 2.87 6.13 0.0672 � 0.0254B2/D 14.69 3.90 3.00 7.79 0.0574 � 0.0204Class 1 11.99 3.25 3.12 5.62 0.0301 � 0.0131Class 2 14.12 3.69 2.83 7.60 0.0689 � 0.0239a Values for total numbers of mutations, transitions, and transversions are reported asthe average for each fimH sequence within a specific subalignment.
Iebba et al.
1412 iai.asm.org Infection and Immunity
on April 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

(�2 � 7.18, P � 0.0074), while strains belonging to the phyloge-netic groups A and B1 were significantly present in NT2 (�2 �7.54, P � 0.006). The strain distribution in relation to E. coli ad-hesive classes was found to be random (data not shown). Theseoverall data could suggest a tendency for CD and B2 strains todevelop a higher SSR in response to a definite intestinal habitat,with specific FimH mutations emerging.
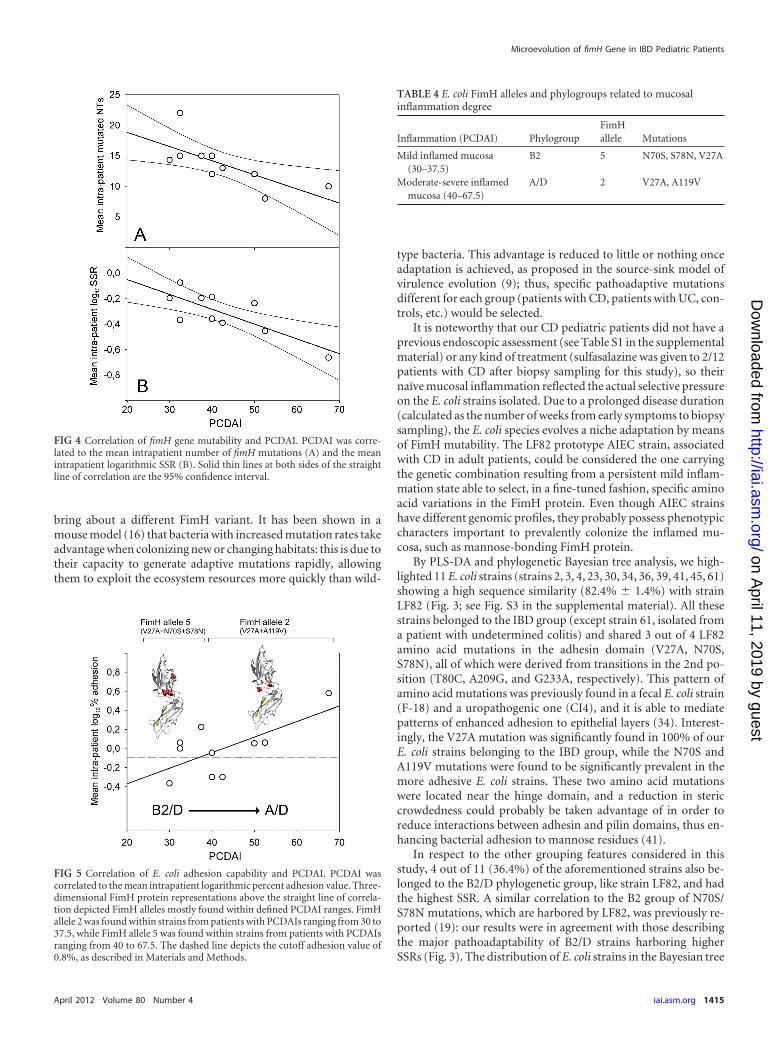
Specific FimH alleles are related to mucosal inflammation.Trying to understand the underlying molecular strategy of an E.coli strain to better fit and adhere to a CD patient naïve inflamedepithelium (that is, one that never underwent pharmacologicaltreatment), we set a graphical relation between PCDAI and themean intrapatient number of mutations, SSRs, and adhesion lev-els (Fig. 4). The equation y � mx � q, where m is the slope and qis the intercept on the y axis, was implemented. Taking into ac-count that two or three E. coli strains were isolated from each CDpatient (see Materials and Methods), we computed the mean in-trapatient values of numbers of fimH mutations, SSRs, and E. coliadhesion percentages. The results showed an inverse correlationfor both mean intrapatient number of mutations (r2 � 0.9677;m � �0.23, P � 0.0234; q � 23.44, P � 0.0002) and mean intrapa-tient log10 SSR (r2 � 0.9192; m � �0.0116, P � 0.0073; q �0.1821, P � 0.2385) in respect to PCDAI (Fig. 4). Within thePCDAI interval from 30 to 37.5, akin to a mild inflammation, themean SSR was 0.656 � 0.063, while within the PCDAI range from40 to 67.5, akin to a moderate to severe inflammation, the meanSSR was 0.374 � 0.065, and this difference was statistically signif-icant (Mann-Whitney U test, P � 0.0016). A direct correlationbetween mean intrapatient E. coli adhesion level and PCDAI wasfound (r2 � 0.4398; m � 0.0164, P � 0.0366; q � �0.6985, P �0.0406) (Fig. 5). Considering the overall FimH mutational pat-terns, we found that the A119V mutation was absent when bothN70S and S78N were present, and vice versa, presenting a mutualexclusion. FimH allele 2 (V27A and A119V) was preponderant instrains from patients with PCDAI values ranging from 40 to 67.5,reciprocally excluding FimH allele 5 (V27A, N70S, and S78N),which was present in strains from patients with a range of PCDAIsof from 30 to 37.5 (�2 � 16.9, P � 0.0001) (Table 4). PhylogroupB2 was prevalent among strains from patients with PCDAI valuesranging from 30 to 37.5, while phylogroup A was entirely com-prised of strains from patients within the PCDAI interval from 40to 67.5 (�2 � 8.20, P � 0.017). Phylogroup D was equally distrib-uted among strains from patients along the PCDAI range. Fromthese results, it could be argued that V27A is the primary patho-adaptive FimH mutation arising in pediatric patients with CDcovering the entire spectrum of mucosal inflammation. In addi-tion, B2 E. coli strains, harboring N70S and S78N mutations to-gether with V27A, could take advantage of only a mildly inflamedmucosa, because we did not find any B2 phylogroup in strainsfrom patients with PCDAIs ranging from 40 to 67.5. Finally, itcould be suggested that in a naïve pediatric patient with CD (withno previous pharmacological treatment) with a moderately to se-verely inflamed mucosa, strong adhesion can be achieved by an E.coli strain of the A/D phylogroup harboring FimH allele 2 (V27Aand A119V).
DISCUSSION
Adhesion capability is a critical step for bacterial fitness in naturalenvironments. E. coli species were found to be enormously abun-dant in the intestinal mucosa of pediatric patients with IBD, thus
FIG 2 Clustered image mapping of fimH gene mutation weights. Simca-P�software was used to compute weight coefficients for each mutation (70 xvariables) on a patient’s status (3 y variables) with a scaled and centered dataset. These coefficients were useful to interpret the influence of the x variableson the y ones. Coefficients for different responses (y variables) were also com-parable, as the y variables were normalized (scaled). As depicted in the color-coded legend, the higher that the coefficient value is, the higher the weight(red), while the lower that the value is, the lower the weight (turquoise).
Microevolution of fimH Gene in IBD Pediatric Patients
April 2012 Volume 80 Number 4 iai.asm.org 1413
on April 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

leading to the hypothesis of an enhanced adhesion capability ofsuch strains. It is well-known from the literature that naturallyoccurring amino acid replacements in the FimH protein, the mainE. coli adhesive hook, can modify its tropism toward epithelialtissue (43). In this study, we found that fimH gene and FimHprotein variants were able to predict a patient’s disease status, theE. coli phylogroup, and the percent adhesion to Caco-2 cells. Thesignificantly higher similarity observed between fimH alignmentsfrom patients with CD could be explained by a reduced intestinalallowance, with a lower number of ecological niches being avail-able. A higher homogeneity between fimH alignments in the B2group and adhesive E. coli strains was also found. Moreover, align-ments of fimH from CD and B2 strains showed a significantlyhigher mean SSR. In this context, only a few E. coli FimH variantswill survive and colonize. This could also explain why E. coli B2strains are the ones almost always recovered in a pathological con-text and related to the virulent type (11, 23). Those strains couldbe part of E. coli subpopulations with higher SSRs that, in a chang-ing habitat, will be better able to survive.
We observed an inverse correlation both for the mean intrapa-tient number of mutations and the mean intrapatient log10 SSR in
respect to PCDAI (Fig. 4), while a direct correlation between themean intrapatient E. coli adhesion level and PCDAI was found(Fig. 5). It was proposed not only how rapidly pathoadaptiveFimH mutations could arise (positive selection) but also how rap-idly they could be lost (purifying selection), once selective pres-sure is weakened (35). Table 4 reports the phylogroups and FimHvariants predominant in mucosa with different degrees of inflam-mation. From these results, it could be argued that V27A is theprimary pathoadaptive FimH mutation arising in pediatric pa-tients with CD with mucosal inflammation covering the entirespectrum. In addition, B2 E. coli strains, harboring N70S andS78N mutations together with V27A, could take advantage of onlya mildly inflamed mucosa, because we did not find any B2 phylo-group in strains from patients with PCDAIs ranging from 40 to67.5. Finally, it could be suggested that, in a naïve pediatric patientwith CD (with no previous treatment) with moderately to severelyinflamed mucosa, a strong adhesion can be achieved by an E. colistrain of the A/D phylogroup harboring FimH allele 2 (V27A andA119V). These results could imply that a shift is occurring in theintestinal habitat when the inflammation level is indicated by aPCDAI greater than 40, and a new naïve selective pressure would
FIG 3 Bayesian phylogenetic tree. A Bayesian tree generated by Geneious (version 4.8.3) software as described in Materials and Methods is depicted. This rootedphylogenetic tree of the fimH sequences of 52 strains (plus the fimH sequences of strains MG1655 and LF82) showed four distinct groups: NT1a, NT1b (NT1aand NT1b were joined into NT1 in the text for SSR analysis), NT2, and NT3. These groups were progressively denoted from higher SSRs to lower ones. Inside eachgroup, depicted with an ellipse, were reported the amino acid mutations found in �90% of strains belonging to the group itself. The black circle indicates the root.
Iebba et al.
1414 iai.asm.org Infection and Immunity
on April 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
bring about a different FimH variant. It has been shown in amouse model (16) that bacteria with increased mutation rates takeadvantage when colonizing new or changing habitats: this is due totheir capacity to generate adaptive mutations rapidly, allowingthem to exploit the ecosystem resources more quickly than wild-
type bacteria. This advantage is reduced to little or nothing onceadaptation is achieved, as proposed in the source-sink model ofvirulence evolution (9); thus, specific pathoadaptive mutationsdifferent for each group (patients with CD, patients with UC, con-trols, etc.) would be selected.
It is noteworthy that our CD pediatric patients did not have aprevious endoscopic assessment (see Table S1 in the supplementalmaterial) or any kind of treatment (sulfasalazine was given to 2/12patients with CD after biopsy sampling for this study), so theirnaïve mucosal inflammation reflected the actual selective pressureon the E. coli strains isolated. Due to a prolonged disease duration(calculated as the number of weeks from early symptoms to biopsysampling), the E. coli species evolves a niche adaptation by meansof FimH mutability. The LF82 prototype AIEC strain, associatedwith CD in adult patients, could be considered the one carryingthe genetic combination resulting from a persistent mild inflam-mation state able to select, in a fine-tuned fashion, specific aminoacid variations in the FimH protein. Even though AIEC strainshave different genomic profiles, they probably possess phenotypiccharacters important to prevalently colonize the inflamed mu-cosa, such as mannose-bonding FimH protein.
By PLS-DA and phylogenetic Bayesian tree analysis, we high-lighted 11 E. coli strains (strains 2, 3, 4, 23, 30, 34, 36, 39, 41, 45, 61)showing a high sequence similarity (82.4% � 1.4%) with strainLF82 (Fig. 3; see Fig. S3 in the supplemental material). All thesestrains belonged to the IBD group (except strain 61, isolated froma patient with undetermined colitis) and shared 3 out of 4 LF82amino acid mutations in the adhesin domain (V27A, N70S,S78N), all of which were derived from transitions in the 2nd po-sition (T80C, A209G, and G233A, respectively). This pattern ofamino acid mutations was previously found in a fecal E. coli strain(F-18) and a uropathogenic one (CI4), and it is able to mediatepatterns of enhanced adhesion to epithelial layers (34). Interest-ingly, the V27A mutation was significantly found in 100% of ourE. coli strains belonging to the IBD group, while the N70S andA119V mutations were found to be significantly prevalent in themore adhesive E. coli strains. These two amino acid mutationswere located near the hinge domain, and a reduction in stericcrowdedness could probably be taken advantage of in order toreduce interactions between adhesin and pilin domains, thus en-hancing bacterial adhesion to mannose residues (41).
In respect to the other grouping features considered in thisstudy, 4 out of 11 (36.4%) of the aforementioned strains also be-longed to the B2/D phylogenetic group, like strain LF82, and hadthe highest SSR. A similar correlation to the B2 group of N70S/S78N mutations, which are harbored by LF82, was previously re-ported (19): our results were in agreement with those describingthe major pathoadaptability of B2/D strains harboring higherSSRs (Fig. 3). The distribution of E. coli strains in the Bayesian tree
FIG 4 Correlation of fimH gene mutability and PCDAI. PCDAI was corre-lated to the mean intrapatient number of fimH mutations (A) and the meanintrapatient logarithmic SSR (B). Solid thin lines at both sides of the straightline of correlation are the 95% confidence interval.
FIG 5 Correlation of E. coli adhesion capability and PCDAI. PCDAI wascorrelated to the mean intrapatient logarithmic percent adhesion value. Three-dimensional FimH protein representations above the straight line of correla-tion depicted FimH alleles mostly found within defined PCDAI ranges. FimHallele 2 was found within strains from patients with PCDAIs ranging from 30 to37.5, while FimH allele 5 was found within strains from patients with PCDAIsranging from 40 to 67.5. The dashed line depicts the cutoff adhesion value of0.8%, as described in Materials and Methods.
TABLE 4 E. coli FimH alleles and phylogroups related to mucosalinflammation degree
Inflammation (PCDAI) PhylogroupFimHallele Mutations
Mild inflamed mucosa(30–37.5)
B2 5 N70S, S78N, V27A
Moderate-severe inflamedmucosa (40–67.5)
A/D 2 V27A, A119V
Microevolution of fimH Gene in IBD Pediatric Patients
April 2012 Volume 80 Number 4 iai.asm.org 1415
on April 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

in respect to disease status showed the presence of E. coli strainsisolated from patients with IBD mainly in the groups with higherSSRs. Similar results were found for phylogroups B2 and D, whilein respect to adhesive classes, the distribution of E. coli strainsoccurred in a random fashion. The SSR could be considered animportant issue in the ability to adapt to environmental changes.Only bacterial strains with high SSRs could develop the skills to bepositively selected in the newly forming habitat (16).
The E. coli mutagenesis response to a specific environmentalstress led to specific transitions and transversions, leading to dis-tinct effects on fimH mutational patterns (Table 1; see Fig. S3 inthe supplemental material). We found only a few fimH gene vari-ants in the intestine in the context of IBD, and one may think thata specific stress (inflammation context) led to a specific mutagenicprofile. Our data could be correlated with the already acceptedidea that the frequency of transition and transversion mutation isa function of both environmental factors and the stress-responsesof organisms (depending on the single-locus SSR value of thestrains). Interestingly, in agreement with the findings ofSokurenko et al., the dN/dS ratio, calculated to determine fimHvariation across our E. coli isolates, was much less than 1, probablydue to an overwhelming purifying selection above the nonsynony-mous mutations (35). Strains from patients with UC showed in-creased numbers of mutations and dN/dS ratios, and strains frompatients with CD had an enhanced mutational rate as a commonstrategy to survive in a pathological habitat, but different FimHmutations arose. This phenomenon could imply a different be-havior adopted by E. coli to survive under different shear stressesand different inflammation conditions. Further studies should beperformed to find out if the specific mutagenic profile found couldlead to distinct and predictable changes in protein geometry andfunctionality within the same pediatric patient (40). Genetic vari-ation in an originally commensal trait could change the adhesive-ness of E. coli, shifting it toward a phenotype of potential intestinalvirulence. Specific allelic variants could be a hint of specific E. coliadaptation to the inflammatory state of the IBD intestine, thusrepresenting promising new targets for therapeutic interventionin the early stages of the disease.
ACKNOWLEDGMENTS
This work was supported by Broad Medical Research Program (BMRP)grant IBD-0225R3-2 and university MIUR grants to S. Schippa and M. P.Conte.
AIEC strain LF82 was kindly provided by Arlette Darfeuille-Michaud.
REFERENCES1. Aprikian P, et al. 2007. Interdomain interaction in the FimH adhesin of
Escherichia coli regulates the affinity to mannose. J. Biol. Chem. 282:23437–23446.
2. Barnich N, et al. 2007. CEACAM6 acts as a receptor for adherent-invasiveE. coli, supporting ileal mucosa colonization in Crohn disease. J. Clin.Invest. 117:1566 –1574.
3. Boudeau J, Barnich N, Darfeuille-Michaud A. 2001. Type 1 pili-mediated adherence of Escherichia coli strain LF82 isolated from Crohn’sdisease is involved in bacterial invasion of intestinal epithelial cells. Mol.Microbiol. 39:1272–1284.
4. Boudeau J, Glasser AL, Masseret E, Joly B, Darfeuille-Michaud A. 1999.Invasive ability of an Escherichia coli strain isolated from the ileal mucosaof a patient with Crohn’s disease. Infect. Immun. 67:4499 – 4509.
5. Bringer MA, Glasser AL, Tung CH, Méresse S, Darfeuille-Michaud A.2006. The Crohn’s disease-associated adherent-invasive Escherichia colistrain LF82 replicates in mature phagolysosomes within J774 macro-phages. Cell. Microbiol. 8:471– 484.
6. Cartun RW, Van Kruiningen HJ, Pedersen CA, Berman MM. 1993. Animmunocytochemical search for infectious agents in Crohn’s disease.Mod. Pathol. 6:212–219.
7. Carvalho FA, et al. 2009. Crohn’s disease adherent-invasive Escherichiacoli colonize and induce strong gut inflammation in transgenic mice ex-pressing human CEACAM. J. Exp. Med. 206:2179 –2189.
8. Chassaing B, et al. 2011. Crohn disease-associated adherent-invasive E.coli bacteria target mouse and human Peyer’s patches via long polar fim-briae. J. Clin. Invest. 121:966 –975.
9. Chattopadhyay S, et al. 2009. High frequency of hotspot mutations incore genes of Escherichia coli due to short-term positive selection. Proc.Natl. Acad. Sci. U. S. A. 106:12412–12417.
10. Chen SL, et al. 2009. Positive selection identifies an in vivo role for FimHduring urinary tract infection in addition to mannose binding. Proc. Natl.Acad. Sci. U. S. A. 106:22439 –22444.
11. Clermont O, Bonacorsi S, Bingen E. 2000. Rapid and simple determina-tion of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol.66:4555– 4558.
12. Conte MP, et al. 2006. Gut-associated bacterial microbiota in paediatricpatients with inflammatory bowel disease. Gut 55:1760 –1767.
13. Darfeuille-Michaud A, et al. 2004. High prevalence of adherent-invasiveEscherichia coli associated with ileal mucosa in Crohn’s disease. Gastroen-terology 127:412– 421.
14. Darfeuille-Michaud A, et al. 1998. Presence of adherent Escherichia colistrains in ileal mucosa of patients with Crohn’s disease. Gastroenterology115:1405–1413.
15. Dias RC, Moreira BM, Riley LW. 2010. Use of fimH single-nucleotidepolymorphisms for strain typing of clinical isolates of Escherichia coli forepidemiologic investigation. J. Clin. Microbiol. 48:483– 488.
16. Giraud A, et al. 2001. Costs and benefits of high mutation rates: adaptiveevolution of bacteria in the mouse gut. Science 291:2606 –2608.
17. Glasser AL, et al. 2001. Adherent invasive Escherichia coli strains frompatients with Crohn’s disease survive and replicate within macrophageswithout inducing host cell death. Infect. Immun. 69:5529 –5537.
18. Hartley MG, et al. 1992. The rectal mucosa-associated microflora inpatients with ulcerative colitis. J. Med. Microbiol. 36:96 –103.
19. Hommais F, et al. 2003. The FimH A27V mutation is pathoadaptive forurovirulence in Escherichia coli B2 phylogenetic group isolates. Infect. Im-mun. 71:3619 –3622.
20. Hughes AL, Nei M. 1989. Nucleotide substitution at major histocompat-ibility complex class II loci: evidence for overdominant selection. Proc.Natl. Acad. Sci. U. S. A. 86:958 –962.
21. Kaser A, Zeissig S, Blumberg RS. 2010. Inflammatory bowel disease.Annu. Rev. Immunol. 28:573– 621.
22. Klemm P. 1994. Fimbriae, adhesion, genetics, biogenesis and vaccines.CRC Press, Boca Raton, FL.
23. Kotlowski R, Bernstein CN, Sepehri S, Krause DO. 2007. High preva-lence of Escherichia coli belonging to the B2�D phylogenetic group ininflammatory bowel disease. Gut 56:610 – 612.
24. Martin HM, et al. 2004. Enhanced Escherichia coli adherence and invasionin Crohn’s disease and colon cancer. Gastroenterology 127:80 –93.
25. Masseret E, et al. 2001. Genetically related Escherichia coli strains associ-ated with Crohn’s disease. Gut 48:320 –325.
26. Mossman KL, et al. 2008. Cutting edge: FimH adhesin of type 1 fimbriaeis a novel TLR4 ligand. J. Immunol. 181:6702– 6706.
27. Ofek I, Doyle RJ. 1994. Bacterial adhesion to cells and tissues, p 536 –546.Chapman and Hall, New York, NY.
28. Rendón MA, et al. 2007. Commensal and pathogenic Escherichia coli usea common pilus adherence factor for epithelial cell colonization. Proc.Natl. Acad. Sci. U. S. A. 104:10637–10642.
29. Rolhion N, Darfeuille-Michaud A. 2007. Adherent-invasive Escherichiacoli in inflammatory bowel disease. Inflamm. Bowel Dis. 13:1277–1283.
30. Sasaki M, et al. 2007. Invasive Escherichia coli are a feature of Crohn’sdisease. Lab. Invest. 87:1042–1054.
31. Schembri MA, Pallesen L, Connell H, Hasty DL, Klemm P. 1996. Linkerinsertion analysis of the FimH adhesin of type 1 fimbriae in an Escherichiacoli fimH-null background. FEMS Microbiol. Lett. 137:257–263.
32. Seksik P, et al. 2006. The role of bacteria in onset and perpetuation ofinflammatory bowel disease. Aliment. Pharmacol. Ther. 24:11–18.
33. Sokurenko EV, et al. 1998. Pathogenic adaptation of Escherichia coli bynatural variation of the FimH adhesin. Proc. Natl. Acad. Sci. U. S. A.95:8922– 8926.
34. Sokurenko EV, Courtney HS, Maslow J, Siitonen A, Hasty DL. 1995.
Iebba et al.
1416 iai.asm.org Infection and Immunity
on April 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

Quantitative differences in adhesiveness of type 1 fimbriated Escherichiacoli due to structural differences in fimH genes. J. Bacteriol. 177:3680 –3686.
35. Sokurenko EV, et al. 2004. Selection footprint in the FimH adhesin showspathoadaptive niche differentiation in Escherichia coli. Mol. Biol. Evol.21:1373–1383.
36. Subramanian S, Campbell BJ, Rhodes JM. 2006. Bacteria in the patho-genesis of inflammatory bowel disease. Curr. Opin. Infect. Dis. 19:475–484.
37. Swidsinski A, et al. 2002. Mucosal flora in inflammatory bowel disease.Gastroenterology 122:44 –54.
38. Tanaka T, Nei M. 1989. Positive Darwinian selection observed at thevariable-region genes of immunoglobulins. Mol. Biol. Evol. 6:447– 459.
39. Tchesnokova V, et al. 2011. Type 1 fimbrial adhesin FimH elicits an
immune response that enhances cell adhesion of Escherichia coli. Infect.Immun. 79:3895–3904.
40. Tchesnokova V, et al. 2008. Integrin-like allosteric properties of the catchbond-forming FimH adhesin of Escherichia coli. J. Biol. Chem. 283:7823–7833.
41. Thomas WE, Trintchina E, Forero M, Vogel V, Sokurenko EV. 2002.Bacterial adhesion to target cells enhanced by shear force. Cell 109:913–923.
42. Thompson-Chagoyán OC, Maldonado J, Gil A. 2005. Aetiology of in-flammatory bowel disease (IBD): role of intestinal microbiota and gut-associated lymphoid tissue immune response. Clin. Nutr. 24:339 –352.
43. Weissman SJ, et al. 2007. Differential stability and trade-off effects ofpathoadaptive mutations in the Escherichia coli FimH adhesin. Infect. Im-mun. 75:3548 –3555.
Microevolution of fimH Gene in IBD Pediatric Patients
April 2012 Volume 80 Number 4 iai.asm.org 1417
on April 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from