microcystis aeruginosa toxin: cell culture toxicity, hemolysis, and
TRANSCRIPT

APPLIED AND ENVIRONMENTAL MICROBIOLOGY. June 1982, p. 1425-14330099-2240/82/061425-09$02.00/0
Vol. 43, No. 6
Microcystis aeruginosa Toxin: Cell Culture Toxicity,Hemolysis, and Mutagenicity Assays
W. 0. K. GRABOW,l* W. C. Du RANDT,1 O. W. PROZESKY,2 AND W. E. SCOTT1
National Institute for Water Research, Council for Scientific and Industrial Research, P.O. Box 395, Pretoria0001,1 and National Institute for Virology, Johannesburg,2 South Africa
Received 9 November 1981/Accepted 22 February 1982
Crude toxin was prepared by lyophilization and extraction of toxic Microcystisaeruginosa from four natural sources and a unicellular laboratory culture. Theresponses of cultures of liver (Mahlavu and PLC/PRF/5), lung (MRC-5), cervix(HeLa), ovary (CHO-Kl), and kidney (BGM, MA-104, and Vero) cell lines tothese preparations did not differ significantly from one another, indicating thattoxicity was not specific for liver cells. The results of a trypan blue staining testshowed that the toxin disrupted cell membrane permeability within a few minutes.Human, mouse, rat, sheep, and Muscovy duck erythrocytes were also lysedwithin a few minutes. Hemolysis was temperature dependent, and the reactionseemed to follow first-order kinetics. Escherichia coli, Streptococcus faecalis,and Tetrahymena pyriformis were not significantly affected by the toxin. Thetoxin yielded negative results in Ames/Salmonella mutagenicity assays. Micro-titer cell culture, trypan blue, and hemolysis assays for Microcvstis toxin aredescribed. The effect of the toxin on mammalian cell cultures was characterizedby extensive disintegration of cells and was distinguishable from the effects of E.coli enterotoxin, toxic chemicals, and pesticides. A possible reason for the acutelethal effect of Microcystis toxin, based on cytolytic activity, is discussed.
Some cellular components of various speciesof algae have toxic, carcinogenic, or mutageniceffects (7, 11, 23, 31, 32, 34, 37). Neither thechemical nor the toxicological properties ofthese compounds have been fully established (6,7, 11). Certain strains of the blue-green alga(cyanobacterium) Microcystis aeruginosa Kutzemend Elenkin produce toxins which may con-sist of different combinations of a number ofpeptide or peptide-containing toxins of unde-fined structure with hepatotoxic or neurotoxicactivities (7, 11, 22). The M. aeruginosa toxinknown as microcystin or fast-death factor,which kills white mice in 1 to 3 h, may consist ofmore than one type of cyclic polypeptide whichmay contain up to 16 amino acids and have amolecular weight ranging from 654 to 19,400 (7,11, 28, 37). Death of mice and vervet monkeysinjected with Microcystis toxin has been as-cribed to circulatory failure as an indirect conse-quence of extensive liver damage (10, 37, 38).However, the short survival time suggests thatliver damage may not be the primary cause ofdeath, and indications that neurotoxic activitylocated in the same fraction as the hepatotoxinor in a different fraction is the cause of deathhave been observed (11). In white rat and mousetoxicological studies in which algal toxin prepa-rations that probably included microcystin butpossibly also other toxins have been used, a
number of disturbances in enzymatic systemsand metabolic processes have been observed,and it has been concluded that the toxins of blue-green algae cause degenerative changes in par-enchymatous organs and brain cells (22). Toxinsof blue-green algae have been reported to agglu-tinate erythrocytes (6) and to be cytopathogenicfor cultures of mammalian cells (22). Primaryfibroblast cultures of human and rat embryocells have proved particularly sensitive, fol-lowed, in decreasing order of sensitivity, by a ratfibroblast cell line and the HeLa, A-8, and HEp-2 cell lines (22).
This study deals with the response of previ-ously untested cells to Microcystis toxin, thedevelopment and evaluation of biological assaysfor Microcystis toxin, and the testing of toxin formutagenic activity by means of the Ames/Sal-monella microsome mutagenicity assay (Amestest). The results contribute to the characteriza-tion of Microcystis toxin, the establishment ofpractical methods for research on the toxin, andthe understanding of its mechanism of action.
MATERIALS AND METHODS
Microcystis toxin. Harvests consisting predominantlyof toxic M. aeruginosa were collected in South Africafrom the Hartbeespoort Dam (near Pretoria) in 1978,the Vaal Dam (near Johannesburg) in 1977 and 1980,and the Roodeplaat Dam (near Pretoria) in 1978 (35,
1425

APPL. ENVIRON. MICROBIOL.
37). Similar harvests of nontoxic Microcystis orga-nisms collected at the Hartbeespoort Dam in 1974 and1979 and the Rietvlei Dam (near Pretoria) in 1975served as negative controls. A unicellular laboratoryculture of a toxic strain of M. aeruginosa designatedWR70 (35) was propagated as described elsewhere (35;W. E. Scott, D. J. Barlow, and J. H. Hauman, Pro-ceedings ofthe International Conference on the WaterEnvironment: Algal Toxins and Health, in press).Freeze-dried samples were thoroughly ground with apestle and mortar, suspended in 0.1 M ammoniumbicarbonate (1 g/100 ml), stirred overnight at 10°C, andcentrifuged (10,000 x g for 30 min); the toxin-contain-ing supernatant was then freeze-dried. This materialwas suspended in distilled water (4 mg/ml) and thenfiltered (Sartorius SM/S13400 [diameter, 47 mm; poresize, 5 ,m] followed by Sartorius SM/N11306 [diame-ter, 25 mm; pore size, 0.45 pLm]) for testing.
Escherichia coli enterotoxin. E. coli strains 18, B7A,and H10407, which all produce both heat-labile andheat-stable enterotoxins (8, 9, 15, 33), were suppliedby A. S. Greeff (Institute of Pathology, Pretoria).Cultures were grown in Oxoid Nutrient Broth no. 2 for48 h at 35°C without shaking and centrifuged (10,000 xg for 30 min); then the supernatant was filteredthrough a membrane (pore size, 0.45 pm) for toxicitytesting.
Cell cultures. The human hepatoma cell lines PLC/PRF/5 (hepatitis B surface antigen positive) (passage91) and Mahlavu (hepatitis B surface antigen negative)(passage 102) (1, 29, 30) were obtained from J. J.Alexander, R. Saunders, and J. A. Pienaar (NationalInstitute for Virology, Johannesburg). The Chinesehamster ovary cell line CHO-Kl (passage 14) waspurchased from Flow Laboratories, Inc., and the fetalrhesus monkey kidney cell line MA-104 and the humanembryo lung cell line MRC-5 (passage 27) were pur-chased from M. A. Bioproducts. Cultures of the Vero(African green monkey kidney) and HeLa (humancervical carcinoma) cell lines were provided by E. M.Bey (National Institute for Virology) and J. A. Pien-aar. The BGM (African green monkey kidney celllines) and primary vervet kidney cells have beendescribed elsewhere (14). Hepatoma, HeLa, MA-104,Vero, BGM, and primary vervet kidney cells weregrown in Eagle minimal essential medium with Earlesalts EMEM (Auto-Pow; Flow Laboratories) supple-mented (per milliliter) with 100 U of penicillin, 100 pgof streptomycin, 50 ptg of neomycin, 292 pg of L-glutamine, and 2 mg of sodium bicarbonate. Gentamy-cin (50 ,ug/ml) and tylosin (8 p.g/ml) were occasionallyadded to control mycoplasma contamination in PLC/PRF/5 cultures. CHO-Kl cells were cultured in Nutri-ent Mixture F12 Ham (Bio-Cult; GIBCO Laboratories)containing (per milliliter) penicillin (100 U), streptomy-cin (100 ,ug), and sodium bicarbonate (1,176 ,ug). E199medium with L-glutamine and Earle salts (Flow Labo-ratories), supplemented (per milliliter) with penicillin(100 U), streptomycin (100 p.g), and sodium bicarbon-ate (2.2 mg), was used for MRC-5 cell cultivation.Each medium was supplemented with 10% heat-inacti-vated fetal calf serum (State Vaccine Institute, CapeTown, South Africa) for the propagation of cells at37°C and with 2% serum for the maintenance ofcultures at 33°C (in 75- or 25-cm2 plastic tissue cultureflasks [Costar]). Equal volumes of two solutions, onecontaining (per milliliter) 2.5 mg of Trypsin 1:250(Difco Laboratories), 100 pg of penicillin, and 100 jig
of streptomycin and the other containing (per millili-ter) 1 mg each of EDTA (Titriplex III; E. Merck) andglucose in sodium chloride (8 mg/ml), 0.2 mg ofpotassium chloride, 2.89 mg of disodium hydrogenphosphate 12 hydrate, and 0.2 mg of potassium hydro-gen phosphate (calcium- and magnesium-free phos-phate-buffered saline), were mixed together immedi-ately before being used for the trypsinization of thecells.
Microtiter cell culture assay. The mammalian cellcultures were trypsinized, and the cells were suspend-ed in appropriate volumes of growth medium supple-mented with 2 to 10% serum (the harvest from conflu-ent growth in 25-cm2 flasks was suspended in 10 to 25ml of medium), and 0.1-ml volumes of these suspen-sions were seeded into wells of microtiter plates(Titertek plates with 96 flat-bottom wells; Flow Labo-ratories). Dilutions of test suspensions (0.1 ml) wereadded immediately or after 24 h. Sterile distilled waterand similar preparations of nontoxic strains of M.aeruginosa were used in negative controls. After incu-bation at 37°C in an atmosphere of 5% C02, the cellswere studied for cytopathogenic effects.
Hemagglutination and hemolysis tests. Sheep, Mus-covy duck, mouse, rat, and human (group A, RH+)erythrocytes were washed three times with 0.85%sodium chloride and resuspended in sodium chloride.In hemagglutination tests, 1% suspensions (0.1 ml) oferythrocytes were seeded into wells of microtiterplates (Titertek plates with 96 V-shaped wells; FlowLaboratories), and dilutions of the test suspensions(0.1 ml) were added immediately. Saline and similarpreparations of nontoxic strains of M. aeruginosawere used in negative controls. The plates were cov-ered and allowed to stand for 3 to 4 h at roomtemperature. Results were recorded as ++ (no cellssettling at the bottom, erythrocyte agglutination), + -(partially positive: about half of the cells settling at thebottom, the rest remaining diffused), or - - (negative:well-defined spot of erythrocytes settled at the bottomof the well and clear solution at the top), as describedelsewhere (6). In hemolysis tests, equal volumes oferythrocytes in saline (10% [vol/vol]) and test materialsuspended in saline were mixed and kept at 35°C for 60min. Reaction mixtures were then macrohematocritcentrifuged in Wintrobe tubes (3,000 x g for 3 min atroom temperature or 2,260 x g for 30 min at 5°C) (41)or microhematocrit centrifuged by the method de-scribed previously (41) or in 1.5-ml Eppendorff tubeswith a Heraeus-Christ type 00912 microhematocritcentrifuge for 5 min at room temperature. Saline orsaline preparations of nontoxic strains of M. aerugin-osa were mixed with erythrocyte suspensions in nega-tive controls. Test and negative-control suspensionswere centrifuged simultaneously. After centrifugation,the supernatants were assayed, and any reduction inpacked-erythrocyte volume and concomitant increasein visible hemoglobin concentration relative to thenegative-control levels was recorded as a positivehemolysis reaction (41).Tetrahymena pyriformis. A culture of T. pyriformis W
was supplied by J. L. Slabbert (National Institute forWater Research, Pretoria). Propagation of the orga-nisms and preparation of washed cell suspensions ingrowth medium or balanced Osterhout salt solutionwere done as described elsewhere (J. L. Slabbert andW. S. G. Morgan, Water Res., in press). Appropriatemixtures of cell suspensions and suspensions of test
1426 GRABOW ET AL.

M. AERUGINOSA TOXIN 1427
material in water were incubated at 27°C. Osterhoutsolution, saline, or distilled water was used in negativecontrols. Reaction mixtures contained about 3 x 104cells per ml. The effect of test suspensions on themotility and morphology of organisms was evaluatedwith a hemacytometer by comparison with the effectof negative controls. Cell membrane permeability wasstudied by using the trypan blue test.Mouse toxicity test. White mice weighing about 20 g
each were inoculated intraperitoneally with the testsuspensions (7, 18, 28, 37).
Test for E. coli heat-labile enterotoxin. Suspensionsof CHO-Kl cells were subcultured in 0.1-ml quantitiesinto wells of flat-bottom microtiter plates, and dilu-tions of enterotoxin preparations (0.1 ml) were addedimmediately. Similar preparations of non-enterotoxi-genic strains of E. coli were used in negative controls.After incubation for 24 h at 37°C in an atmosphere of5% C02, the morphology of the cells was investigated.The percentage of elongated cells (clearly bipolar,length three times greater than diameter) was deter-mined (15, 33), and the results were recorded as + +(more than 15% elongated cells), + - (ca. 15% elongat-ed cells), or - - (less than 15% elongated cells). Thisprocedure was used in all studies of the effect of E. colienterotoxin and Microcystis toxin on the morphologyof mammalian cells.Trypan blue test of cell membrane permeability. The
confluent growth of cell cultures in 25-cm2 flasks wastrypsinized, and the cells were suspended in appropri-ate volumes (5 to 10 ml) of growth medium supple-mented with 10% serum. A 1.0-ml volume of each cellsuspension was mixed with 1.0 ml of test suspensionand incubated at 35°C in an atmosphere of 5% CO2- Innegative controls, the test suspension was replaced byphosphate-buffered saline and similar preparations ofnontoxic strains of M. aeruginosa. After 60 min at35°C, 0.1 ml each was removed from the test andcontrol suspensions and mixed with an equal volumeof membrane-filtered 0.5% trypan blue solution inphosphate-buffered saline (17), and the percentage ofinactivated cells was evaluated with a hemacytometer(improved Neubauer, Spencer Bright-line; AmericanOptical Corp.) as described by Hoskins (17).
Bacteria. Strains of E. coli and Streptococcus faeca-lis were isolated from wastewater and identified byGram staining and API 20E and API 10 Strep testsystems (12). Inactivation of stationary-phase bacteriawas evaluated by adding 0.1-ml overnight broth cul-ture (35°C) samples diluted to 10-2 in saline to 4 ml ofsaline containing 2 mg of lyophilized Microcystis ex-tract per ml and counting viable bacteria before andafter incubation for 5 h at 35°C. The effect of toxin onthe multiplication of bacteria was evaluated by inocu-lating 0.1-ml overnight broth culture samples diluted10' in saline into 4 ml of nutrient broth (Difco)containing 2 mg of lyophilized Microcystis extract perml and counting viable bacteria before and after incu-bation for 24 h at 35°C. In negative controls, toxin wasreplaced by similar extracts of nontoxic Microcystisharvests. We counted bacteria by membrane filtration,using m-Endo LES agar for E. coli and m-Enterococ-cus agar for S. faecalis (12). Lower counts in testmixtures than in negative controls were taken asevidence of bacterial inactivation or inhibition of mul-tiplication.Ames tests. Plate incorporation assays and spot tests
in which tester Salmonella sp. strains TA98 and
TA100 were used were performed as described previ-ously (3, 13). Tests were done on the toxin prepara-tions described above as well as on slurries of toxic M.aeruginosa in dimethyl sulfoxide (10 mg/ml), whichwere tested without filtration.
Chemical toxicants and pesticides. Solutions of anhy-drous copper sulfate, mercuric chloride, ammoniumsulfate, lead chloride, cadmium chloride, and phenolwere prepared as described previously (21). Ethylalcohol was used to facilitate the solution of thepesticides parathion, chlordane, and pentachlorophe-nol (26) in water. All solutions were decontaminatedby membrane filtration.
RESULTSResponse of mammalian cell cultures to Micro-
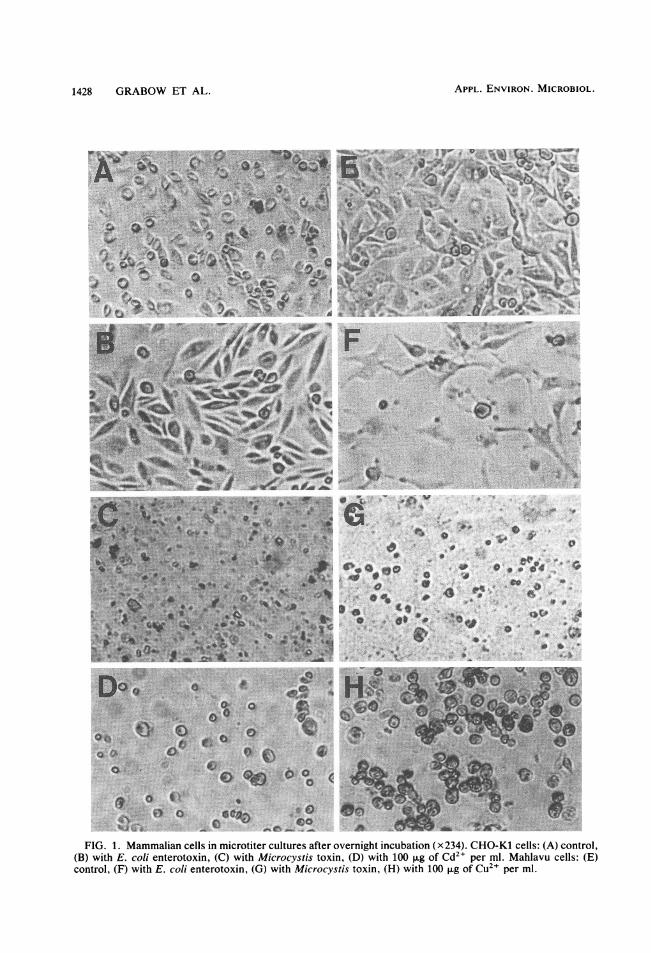
cystis toxin. Cells of all cultures were severelydamaged or disintegrated after overnight incuba-tion in the presence of toxin (Fig. 1C and G). Allcell lines underwent the same type of cell disin-tegration, which resulted in cell debris of differ-ent sizes and granular material (Fig. 1C and G).The disintegration of cells was not preceded byelongation or associated morphological changes.Cytopathogenic effects were detectable withinabout 6 h in cells seeded in the presence ofundiluted toxin preparations and about 8 h afterthe preparations were added to established 24-hcultures. Results of toxin titrations were usuallyread after overnight incubation (16 to 24 h), butthe lowest concentrations of toxin which exerteda cytopathogenic effect were generally detect-able only after 2 to 5 days of incubation. Serumhad no effect on the action of toxin: serum addedconcomitantly with toxin or 4 h after toxin wasadded to washed cells in serum-free growthmedium did not detectably alter the cytopatho-genic effect.The results of a typical toxin titration (Table 1)
illustrate that the sensitivities of the cell lines didnot differ significantly from one another. Similarresults were recorded for all toxin preparations.CHO-Kl and Mahlavu were marginally moresensitive than were the other cell lines, andBGM and Vero cell lines were the least sensi-tive. Primary vervet kidney cells were slightlymore sensitive than were the cell lines. Onedisadvantage of primary cells for toxicity testingwas that they had to be incubated for relativelylong periods before yielding established cellgrowth suitable for testing. The optimum nutri-ent concentration in the growth medium andnumber of cells seeded in the wells had to beestablished for each cell line. Tests in whichcells were seeded in the presence of toxin weregenerally more sensitive than were tests inwhich toxin was added to established cell layers.CHO-Kl and Mahlavu cells proved to be themost practical for the detection of toxicity by amicrotiter assay in which cells are seeded in thepresence of test suspensions. These cell lineswere easy to keep for instant use over several
VOL. 43, 1982
1428 GRABOW ET AL.
En-_
FIG. 1. Mammalian cells in microtiter cultures after overnight incubation (x234). CHO-Kl cells: (A) control,(B) with E. coli enterotoxin, (C) with Microcystis toxin, (D) with 100 p.g of Cd21 per ml. Mahlavu cells: (E)control, (F) with E. coli enterotoxin, (G) with Microcystis toxin, (H) with 100 [Lg Of CU21 per ml.
APPL. ENVIRON. MICROBIOL.

M. AERUGINOSA TOXIN 1429
TABLE 1. Microtiter cell culture, mouse,hemolysis, and trypan blue tests for toxin derived
from a bloom of toxic M. aeruginosa inHartbeespoort Dam
Response to indicated concn (mg/ml) ofTest toxin'
1.0 0.5 0.25 0.125 0.063 0.032
Cell cultureCHO-Kl ++ ++ ++ +.-- __Mahlavu + + + + + + - - -
HeLa. + + + - + - .. _ _ _ _PLC/PRF/5 ++ +.MRC-5 .... ++ +-MA-104 .... ++ +Vero.+-. +.BGM +.- +-
Mouse ....+.+ ++ +
Trypan blue .. ++ _
Hemolysis ++ ++ ++ I_++ ++ ++a Toxin preparation and test procedures are de-
scribed in text. The cell culture results after 24 h areexpressed as follows: + +, distinct cytopathogeniceffect; +-, indications of cytopathogenic effect; --,no effect. The mouse test results after 5 h are ex-pressed as follows: ++, dead; +-, convulsions, ab-normal breathing, pallor; - -, no effect. The results ofthe trypan blue test of Mahlavu cells after 60 min at35°C are expressed as follows: + +, percentage ofstained cells higher than that in negative controls; - -,percentage of stained cells not higher than that innegative controls. The results of the hemolysis testafter 60 min at 35°C are expressed as follows: + +,hematocrit volume of packed erythrocytes smallerthan that in negative controls.
weeks simply by incubation at 33°C in mainte-nance medium that was changed twice a week.The microtiter cell culture assay yielded repro-ducible results, had a fairly distinct endpoint,and proved suitable for titrating toxin derivedfrom a variety of sources.
In the case of the toxin obtained from Hart-beespoort Dam, cell culture assays proved to bemore sensitive than the mouse toxicity test, andthe hemolysis test was the most sensitive (Table1). In similar comparative tests of M. aerlugin-osa WR70 toxin and the toxins obtained fromVaal and Roodeplaat Dams, the cell cultureassays were also more sensitive than the mousetoxicity test; in contrast, the hemolysis test wasless sensitive than both. None of the tests re-sponded to negative controls.To evaluate possible interference by other
compounds which may occur in water and havecytopathogenic effects, we performed microtitercell culture assays of membrane-filtered samplesof wastewater collected at the Daspoort sewage
purification works in Pretoria (12), water fromthe Hartbeespoort Dam, and solutions of toxiccompounds in distilled water. The results of atypical comparative experiment show that rawand treated wastewater and river and dam waterlacking an algal bloom yielded negative results incell culture assays in which Mahlavu or CHO-Kl cells were used (Table 2). Similar resultswere recorded for samples collected on 8 differ-ent days and assays in which BGM, Vero, MA-104, HeLa, or PLC/PRF/5 cells were used. Evenmembrane-filtered samples of water with abloom of toxic M. aeruginosa or laboratorycultures of M. aeruginosa WR70 generally yield-ed negative results. Cell toxicity was detectableonly after the release of toxin from the toxicalgae by lyophilization or sonication. The cyto-pathogenic effect of the chemicals and pesticides(Table 2) was characterized by the rounding ofcells and by death without instant disintegration(Fig. 1D and H) and was distinguishable fromthe effect of Microcystis toxin.Response of cell cultures to E. coli enterotoxin.
In microtiter cell culture assays, CHO-Kl cellsexposed to E. coli enterotoxin displayed thetypical cell elongation (Fig. 1B) described previ-ously (15). Mahlavu cells elongated similarly(Fig. 1F), but PLC/PRF/5 cells did not. The cellelongation induced by the E. coli enterotoxinwas distinguishable from the cytopathogenic ef-fect of Microcystis toxin (Fig. 1).Mouse toxicity test. The mouse toxicity test
was less sensitive than microtiter cell cultureassays to all Microcystis toxin preparations.Mice did not die within 5 h after the inoculationof wastewater, river and dam water, E. colienterotoxin, or relatively high concentrations ofpotentially toxic compounds (Table 2). Themouse toxicity test has the advantage of givingresults within a few hours, and death is a well-defined endpoint for titration purposes.Trypan blue test. Disruption of membrane
permeability was clearly visible in mammaliancells about 5 min after the addition of Microcys-tis toxin. Control (unaffected) cells were round,shining, and colorless, whereas affected cellswere flat, dull, and clearly blue as a result of thepenetrating stain. The responses of CHO-Kl,Mahlavu, PLC/PRF/5, HeLa, and monkey kid-ney cells to Microcystis toxin did not differsignificantly from one another. As an assay forthe toxin, the trypan blue test was not particular-ly sensitive (Table 1). Tests in which CHO-Klcells were used and incubation was for 60 min at35°C proved practical and had a well-defined andreproducible endpoint. The assay was not affect-ed by wastewater, river and dam water, E. colienterotoxin, or relatively high concentrations oftoxic compounds (Table 2).
Hemagglutination and hemolysis tests. Micro-
VOL. 43, 1982

TABLE 2. Microtiter cell culture, mouse, trypan blue, and hemolysis tests for various water samples andtoxic compound solutions"
Sample
Test Stl Activated ApiesrHabeespoort |E co/i LT Cu2+ at (igIml): Hg>+ at (pg/ml): Ammonia (as N)Stld sludge River HatesortE boiL at (~LgIml):sewage effluent water Dam water enterotoxin -
100 10 1 100 10 1 1,000 100 10
Cell cultureCHO-Kl __ __ __ __ E ++ ++ -- ++ ++ ++ +++-Mahlavu - E ++ ----+++++- ++---
Mouse +- + + + +- +- ND ND +- ND ND +- ND ND
Trypan blue -- ++
Hemolysis __ _ _ _-_- _- _- __ _ ++_ _-__ ++.a E, More than 15% of the cells were elongated; ND, not done; -- (hemolysis test), hematocrit volume of
packed erythrocytes not smaller than that in negative controls. The other symbols are explained in footnote a ofTable 1.
b LT, Heat labile.
titer hemagglutination tests of Microcystis toxinyielded results resembling the agglutination oferythrocytes. However, hemolysis tests showedthat the apparent diffusion of erythrocytes andlack of settling were mainly, probably exclusive-ly, due to the lysis of cells. In tests of dilutions oftoxin preparations, no evidence was found thatagglutination occurred in addition to lysis. Bothreactions were temperature dependent. Whenmoderate concentrations of toxin were used,both tests yielded negative results at 4°C, evenafter 16 h. At 10°C, both tests yielded marginallypositive results within 5 h, whereas at 35°C,reactions had reached an endpoint within 5 h.Reactions at different temperatures also failed toindicate agglutination in addition to lysis. Nodifference was detected among the responses ofthe five types of erythrocytes. The results oftitrations indicated that the lysis of erythrocytesby Microcystis toxin was a first-order kineticreaction. Similar results were recorded for alltoxin preparations.Even though the sensitivities of the hemolysis
test to various toxin preparations differed fromthose of microtiter cell culture and mouse toxici-ty tests, no difference was detected among thehemolytic reactions to the toxin preparations. Intests of 4-mg/ml samples of Hartbeespoort DamMicrocystis toxin, microscopic observation re-
vealed that the lysis of erythrocytes startedimmediately and that more than 95% of the cellswere lysed within 2 to 8 min. The detection oflow concentrations of toxin required prolongedincubation. Mixtures of erythrocyte and testsuspensions were successfully incubated in avariety of tubes or vials and centrifuged directlyfor screening purposes or analyzed accuratelyafter hematocrit centrifugation. For microscopicdetection of erythrocyte lysis with a hemacy-
tometer or ordinary microscope slide, a reactionmixture consisting of 0.1 ml of erythrocyte sus-pension (50 x 105 erythrocytes per ml) and 0.1ml of test suspension was sufficient and wasconveniently prepared in wells of microtiterplates. The hemolysis tests were simple andeconomical and yielded results with a clearlydefined endpoint within a short period. Forinstant availability, erythrocyte suspensionswere stored at 4°C for several weeks. Decon-tamination of test suspensions was generally notnecessary, but it was important to have anisotonic reaction mixture to avoid the autolysisof erythrocytes. Tests of ordinary samples ofwater therefore required the addition of sodiumchloride to a final concentration of 0.85%. Un-der these conditions, negative results were re-corded for wastewater, river and dam water, E.coli enterotoxin, and relatively high concentra-tions of toxic compounds (Table 2).Response of T. pyriformis to Microcystis toxin.
No significant difference between suspensionswith and without toxin in viability, motility, ormorphology of T. pyriformis was detected, evenafter 16 h at 27°C. As far as could be establishedby trypan blue tests, the toxin also had no effecton the membrane permeability. These resultsshow that toxin in concentrations highly cytolyt-ic to mammalian cells and lethal to mice (Table1) had no significant effect on T. pyriformis.Response of bacteria to Microcystis toxin. In
saline suspensions containing toxin, the averagecount of E. coli cells increased from 38 x 103 to71 x 106/ml in 5 h at 35°C, and in salinesuspensions without toxin, the count declinedfrom 35 x 103 to 23 x 103/ml. In similar tests onS. faecalis, the count in saline suspensions withtoxin increased from 20 x 103 to 85 x 105/ml,and in negative controls, the average count
APPL. ENVIRON. MICROBIOL.1430 GRABOW ET AL.

M. AERUGINOSA TOXIN 1431
TABLE 2-ContinuedSample
Pb2+ at (,ug/ml): Cd2+ at (pg/ml): Phenol at (pg/ml): Parathion at Chlordane at (±g/ Pentachlorophenol(pg/mi): ml): at (,ug/ml):100 10 1 100 10 1 1,000 100 10 20 10 1 100 10 1 100 10 1
+- ND ND +- ND ND +- ND ND +-ND ND +- ND ND ND +- ND
declined from 21 x 103 to 9 x 103/ml. Theseresults show that both E. coli and S. faecalisactually thrived on the toxin preparations. Thetoxin had no detectable effect on the multiplica-tion of the bacteria in broth for 24 h at 35°C. Inbroth with or without toxin, the average count ofE. coli increased from 4 x 102 to 20 x 108/ml,and that of S. faecalis increased from 2.8 x 102to 90 x 107/ml. Similar results were recorded inexperiments in which preparations of toxic M.aeruginosa were replaced by homologous prepa-rations of nontoxic strains. These results showthat toxin in concentrations highly cytolytic tomammalian cells and lethal to mice (Table 1) hadno detectable effect on E. coli and S. faecalis.Ames tests. No mutagenic activity was detect-
ed in assays of any of the test strains in thepresence or absence of activating liver enzymes.
DISCUSSIONThe absence of a significant difference among
the sensitivities of liver, lung, cervix, ovary, andkidney cells shows that the Microcystis toxin didnot specifically attack liver cells. Histologicaldamage in animals is possibly first detected inthe liver (6, 10, 38) because toxins administeredorally or intraperitoneally are directly transport-ed to the liver, and, as a result of their metabolicactivity, liver cells may absorb most of the toxiceffect. Epithelial cells (CHO-Kl, Mahlavu,HeLa, PLC/PRF/5) are possibly slightly moresensitive than fibroblast cells (MRC-5, Vero,MA-104, BGM) (Table 1). However, it should bekept in mind that these tests were done in vitroon transformed cells and that untransformedcells under in vivo conditions may behave differ-ently. This is illustrated by the primary vervetkidney cells, which were more sensitive thantransformed kidney cells or any of the othertransformed cells. The observation that untrans-formed cells are more sensitive than trans-formed cells has been noted before (22).
The results of the trypan blue and hemolysistests show that one effect of the toxin was toinactivate the cell membrane permeability barri-er almost instantaneously. As in the case of thesensitivities of the cell cultures, there was hardlyany difference among the sensitivities of thedifferent cells to this effect. Damage to cellmembranes substantiates the observation thatdestruction of liver cells is the earliest histologi-cal sign of a toxic effect on animals. Liverdamage is characterized by the breakdown ofsinusoidal endothelia, the disappearance of thespaces of Disse between hepatocytes and sinu-soids, irregular-sized inter- and intralobular con-nective tissue focus proliferation, leukocyte in-filtration, and progressive disruption of tissuestructure (6, 10, 38). In vivo liver cell destruc-tion, the damage to in vitro cell cultures charac-terized by almost complete cell disintegration(Fig. 1C and G), and the penetration of trypanblue into cells that had been exposed for a fewminutes to toxin in suspension cultures indicatethat cell membrane destruction is probably theprimary, possibly only, mechanism of action ofMicrocystis toxin. Destruction of the liver cellpermeability barrier would explain the earlyincrease in serum levels of liver enzymes (10).The failure of disrupted hepatocytes and theliver structure to dispose of the influx of circu-lating blood may account for the toxin poisoningsymptoms of accumulation of blood in the liver(10, 18, 37), pallor (18, 37), and death as aconsequence of circulatory collapse. The instan-taneous disruption of membrane permeabilityand hemolysis indicate that protein synthesis isnot involved in the cytolytic action of the toxin,which may explain the sudden onset of symp-toms of toxin poisoning in vivo and demonstratethat the toxin has no cytotoxic effect.The absence of cell elongation or related mor-
phological changes in all of the cell types afterexposure to toxin shows that the mechanism ofaction of the toxin differs from that of E. coli
VOL. 43, 1982

1432 GRABOW ET AL.
heat-labile enterotoxin. The latter induces typi-cal cell elongation in CHO-Kl (Fig. 1B), Mah-lavu (Fig. 1F), and various other cell lines suchas HeLa and Vero by stimulation of the adenylcyclase enzyme (15). The typical pattern of rapidand extensive cellular disintegration by Micro-cystis toxin (Fig. 1C and G) may prove useful inthe identification and characterization of thetoxin: the pattern was also distinguishable fromthat of heavy metal and pesticide toxin effects(Table 2), which included the gradual roundingof cells, death, and no early disintegration (Fig.1D and H).The failure of Microcystis toxin to significant-
ly affect the protozoan T. pyriformis, the gram-negative bacterium E. coli, and the gram-posi-tive bacterium S. faecalis indicates that thetoxin may specifically attack cell membranes ofwarm-blooded animals. This is substantiated bythe temperature dependence of erythrocyte ly-sis. The resistance of T. pyriformis shows thatthe toxin may be similar to toxins of blue-greenalgae such as Anabaena flos-aquae, which donot affect the protozoan Paramecium caudatum,and may differ from toxins such as those ofFischerella epiphytica and Gloeotricha echinu-lata, which inactivate P. caudatum (31).The erythrocyte lysis results and the lack of
evidence of agglutination suggest that unlessdifferent compounds were involved, the hemag-glutination reported in an earlier study (6) mayhave been mistaken for hemolysis. This indi-cates that the mechanism of action of Microcys-tis toxin differs from that of hemagglutinatingtoxic lectins of higher plants (6, 19, 20, 36) andhemagglutinating compounds of certain marinealgae (5, 6). The molecular weights of plantlectins (in the range of 100,000) are also muchhigher than that of Microcystis toxin (6, 11), andthe survival time of 2 to 4 days after acutedosages of toxic lectins is much longer than thatobserved after acute dosages of Microcystistoxin. It appears that the mechanism of action ofMicrocystis toxin may be more similar to that ofbacterial cytolytic toxins (2, 24) or even snakevenom cardiotoxins (25). The negative Amestest results show that the toxin differs frommutagenic and carcinogenic compounds of cer-tain marine algae (23), fungi (4, 39), and higherplants (40).The cell assays described in this study should
prove useful in research on the characterizationand mechanism of action of Microcystis toxinand related toxins. The observation that thesensitivity of hemolysis tests to preparations ofdifferent harvests of toxic M. aeruginosa did notcorrelate with those of mouse toxicity and cellculture tests suggests that these three tests mea-sure activities of different components and thatthe ratio in which these components are pro-
duced is not the same for all strains of toxic M.aeruginosa. The presence of various compoundswith different molecular weights, physical prop-erties, and amino acid compositions in Micro-cystis extracts has been established (11, 28, 37),and the data on the hemolytic activities of differ-ent preparations are in agreement with the hem-agglutination test results which did not correlatequantitatively with data on the lethal doses ofdifferent toxin preparations for mice (6).The in vitro procedures described in this study
make it possible to investigate the effect oftoxins on the morphology and chemical compo-sition of membranes of selected cells undercontrolled conditions and to distinguish betweencytolytic and cytotoxic activities. The negativeresults recorded in multiple tests of settled andtreated wastewater, river and dam water lackinga toxic algal bloom, and solutions of toxic chem-icals and pesticides (Table 2) in concentrationswhich are rarely exceeded in natural bodies ofwater (16) indicate that false-positive results ofthe cell assays are unlikely to be due to theconventional pollutants in the test samples. Interms of analytical methods, the microtiter cellculture assay was more sensitive than the con-ventional mouse toxicity test to all preparations,and the hemolysis test was the most sensitive tothe Hartbeespoort Dam toxin. The in vitro cellassays may also prove useful for the generalscreening of water quality. Although they areless sensitive to toxicants than more sophisticat-ed biological toxicity assays (21, 26, 27), theymay be used for the detection of shock loads of avariety of potentially acute toxicants because ofbeing simple, quick, and economical.Even though the results of our studies of toxic
M. aeruginosa from five sources were in closeagreement with one another, comparison of ourfindings with those of other studies should bedone with caution because M. aeruginosa, aswell as other blue-green algae, produces a vari-ety of toxic compounds with different proper-ties, toxic effects, and mechanisms of action (6,7, 11, 18, 22, 28, 34, 37). The toxicity noted inthis study may be the result of combined orsynergistic effects of different compounds (2,24), and the effects of other compounds mayhave been antagonized or obscured. Similarstudies on chemically purified and characterizedfractions are therefore in progress.
ACKNOWLEDGMENTS
We thank D. P. Botes and J. Alexander for advice and J. A.Pienaar and H. Kruger for technical assistance.
LITERATURE CITED
1. Alexander, J., G. Macnab, and R. Saunders. 1978. Studieson in vitro production of hepatitis B surface antigen by ahuman hepatoma cell line. Perspect. Virol. 10:103-120.
2. Alouf, J. E. 1977. Cell membranes and cytolytic bacterial
APPL. ENVIRON. MICROBIOL.

M. AERUGINOSA TOXIN 1433
toxins, p. 219-270. In P. Cuatrecasas (ed.), The specificityand action of animal, bacterial and plant toxins. Chapmanand Hall, London.
3. Ames, B. N., J. McCann, and E. Yamasaki. 1975. Methodsfor detecting carcinogens and mutagens with the Salmio-nella/mammalian-microsome mutagenicity test. Mutat.Res. 31:347-364.
4. Bjeldanes, L. F., G. W. Chang, and S. V. Thomson. 1978.Detection of mutagens produced by fungi with the Salmo-nella typhimuriium assay. Appl. Environ. Microbiol.35:1150-1154.
5. Blunden, G., D. J. Rogers, and W. F. Farnham. 1975.Survey of British seaweeds for hemagglutinins. Lloydia38:162-168.
6. Carmichael, W. W., and P. E. Bent. 1981. Hemagglutina-tion method for detection of freshwater cyanobacteria(blue-green algae) toxins. Appl. Environ. Microbiol.41:1383-1388.
7. Collins, M. 1978. Algal toxins. Microbiol. Rev. 42:725-746.
8. DuPont, H. L., S. B. Formal, R. B. Hornick, M. J. Sny-der, J. P. Libonati, D. G. Sheahan, E. H. LaBrec, andJ. P. Kalas. 1971. Pathogenesis of Escherichia coli diar-rhea. N. Engl. J. Med. 285:1-9.
9. Evans, D. J., Jr., D. G. Evans, and S. L. Gorbach. 1973.Production of vascular permeability factor by enterotoxi-genic Escherichia coli isolated from man. Infect. Immun.8:725-730.
10. Falconer, I. R., A. R. B. Jackson, J. Langley, and M. T.Runnegar. 1981. Liver pathology in mice in poisoning bythe blue-green alga Mic rocvstis aeru4ginosa. Aust. J. Biol.Sci. 34:179-187.
11. Gorham, P. R., and W. W. Carmichael. 1979. Phycotox-ins from blue-green algae. Pure Appl. Chem. 52:165-174.
12. Grabow, W. 0. K., J. S. Burger, and E. M. Nupen. 1980.Evaluation of acid-fast bacteria, Candida albicans, enter-ic viruses and conventional indicators for monitoringwastewater reclamation systems. Prog. Water Technol.12:803-817.
13. Grabow, W. 0. K., R. Denkhaus, and P. G. van Rossum.1980. Detection of mutagens in wastewater, a pollutedriver and drinking-water by means of the Ames Salmonel-la/microsome assay. S. Afr. J. Sci. 76:118-123.
14. Grabow, W. 0. K., and E. M. Nupen. 1981. Comparisonof primary kidney cells with the BGM cell line for theenumeration of enteric viruses in water by means of a tubedilution technique, p. 253-256. In M. Goddard and M.Butler (ed.), Viruses and wastewater treatment. Perga-mon Press, Oxford, England.
15. Guerrant, R. L., L. L. Brunton, T. C. Schnaitman, L. I.Rubhun, and A. G. Gilman. 1974. Cyclic adenosine mono-phosphate and alteration of chinese hamster ovary cellmorphology: a rapid, sensitive in vitro assay for theenterotoxins of Vibrio cholerae and Escherichia coli.Infect. Immun. 10:320-327.
16. Hattingh, W. H. J. 1977. Reclaimed water: a health haz-ard? Water S A (Pretoria) 3:104-112.
17. Hoskins, J. M. 1967. Virological procedures, p. 25-319.Butterworths Publishers, Inc., London.
18. Hughes, E. O., P. R. Gorham, and A. Zehnder. 1958.Toxicity of a unialgal culture of Microcvstis aeruiginosa.Can. J. Microbiol. 4:225-236.
19. Ikegwuonu, F. I., and 0. Bassir. 1977. Effects of phytohe-magglutinins from immature legume seeds on the functionand enzyme activities of the liver, and on the histopatho-logical changes of some organs of the rat. Toxicol. Appl.Pharmacol. 40:217-226.
20. Kakade, M. L., K. K. Keahey, C. K. Whitehair, and R. J.Evans. 1965. Morphological changes in rats fed Navybeans. Proc. Soc. Exp. Biol. Med. 119:934-937.
21. Kfir, R., and 0. W. Prozesky. 1981. Detection of toxic
substances in water by means of a mammalian cell culturetechnique. Water Res. 15:553-559.
22. Kirpenko, Y. A., V. V. Stankevich, V. M. Orlovskiy, N. I.Kirpenko, A. V. Bokov, and T. F. Karpenko. 1979. Acomparative assessment of the toxic effect of biologicallyactive substances of blue-green algae at the cellular andorganismic levels. Hydrobiol. J. 15:83-86.
23. Leary, J. V., R. Kfir, J. J. Sims, and D. W. Fulbright.1979. The mutagenicity of natural products from marinealgae. Mutat. Res. 68:301-305.
24. Le Vine, H., and P. Cuatrecasas. 1981. An overview oftoxin-receptor interactions. Pharmacol. Ther. 12:167-207.
25. Louw, A. I., and L. Visser. 1977. Kinetics of erythrocytelysis by snake venom cardiotoxins. Biochim. Biophys.Acta 498:143-153.
26. Morgan, W. S. G. 1975. Monitoring pesticides by meansof changes in electric potential caused by fish opercularrhythms. Prog. Water Technol. 7:33-40.
27. Morgan, W. S. G. 1977. An electronic system to monitorthe effects of changes in water quality on fish opercularrhythms, p. 38-55. In J. Cairns, Jr., K. L. Dickson, andG. F. Westlake (ed.), Biological monitoring of water andeffluent quality, ASTM STP 607. American Society forTesting and Materials, Philadelphia.
28. Murthy, J. R., and J. B. Capindale. 1970. A new isolationand structure for the endotoxin from Microcvstis aeriugin-osa NRC-1. Can. J. Biochem. 48:508-510.
29. Oefinger, P. E., D. L. Bronson, and G. R. Dreesman.1981. Induction of hepatitis B surface antigen in humanhepatoma-derived cell lines. J. Gen. Virol. 53:105-113.
30. Prozesky, 0. W., C. Brits, and W. 0. K. Grabow. 1973. Invitro culture of cell lines from Australia antigen positiveand negative hepatoma patients, p. 358-360. In S. J.Saunders and J. Terblanche (ed.), Liver. Pitman MedicalBooks, London.
31. Ransom, R. E., T. A. Nerad, and P. G. Meier. 1978.Acute toxicity of some bluegreen algae to the protozoanParamecilum caudatum. J. Phycol. 14:114-116.
32. Reynolds, C. S. 1980. Cattle deaths and blue-green algae:a possible instance from Cheshire, England. J. Inst. WaterEng. Sci. 34:74-76.
33. Schoub, B. D., A. S. Greeff, G. Lecatsas, 0. W. Prozesky,I. T. Hay, J. G. Prinsloo, and R. C. Ballard. 1977. Amicrobiological investigation of acute summer gastroen-teritis in black South African infants. J. Hyg. 78:377-385.
34. Schwimmer, M., and D. Schwimmer. 1968. Medical as-pects of phycology, p. 279-358. In D. F. Jackson (ed.),Algae, man and the environment. Syracuse UniversityPress, Syracuse.
35. Scott, W. E. 1974. The isolation of Microcystis. S. Afr. J.Sci. 70:179.
36. Sharon, N., and H. Lis. 1972. Lectins: cell-agglutinatingand sugar-specific proteins. Science 177:949-959.
37. Toerien, D. F., W. E. Scott, and M. J. Pitout. 1976. Mi-crocvstis toxins: isolation, identification, implications.Water S A (Pretoria) 2:160-162.
38. Tustin, R. C., S. J. van Rensburg, and J. N. Eloff. 1973.Hepatic damage in the primate following ingestion of toxicalgae, p. 383-385. In S. J. Saunders and J. Terbianche(ed.), Liver. Pitman Medical Books, London.
39. Wehner, F. C., P. G. Thiel, and R. Vleggaar. 1979. Mu-tagenicity of 5,6-dimethoxysterigmatocystin, a metabolitefrom Aspergillus multicolor, in the Salmonella/microsomesystem. Appl. Environ. Microbiol. 38:1015-1017.
40. Weisburger, E. K. 1979. Natural carcinogenic products.Environ. Sci. Technol. 13:278-281.
41. Wintrobe, M. M., G. R. Lee, D. R. Boggs, T. C. Bithell,J. W. Athens, and J. Foerster. 1974. Clinical hematology,7th ed., p. 110-117. Lea & Febiger, Philadelphia.
VOL. 43, 1982