metabolism medium chain dicarboxylic acids in the … · in situ metabolism of 1,o medium chain...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1988 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 263, No. 24. Issue of 'August 25, pp. 11664-11674,1988 Printed in U.S.A.
In Situ Metabolism of 1,o Medium Chain Dicarboxylic Acids in the Liver of Intact Rats as Detected by 13C and 'H NMR*
(Received for publication, February 8, 1988)
Sebastian CerdanSS, Basil Kiinnecke, Andreas Dollell, and Joachim Seelig From the Biocenter of the University of Basel, Klingelbergstrasse 70, CH-4056 Basel, Switzerland
The hepatic metabolism of 1,~dodecanedioic acid, a physiologically relevant representative of the medium- chain dicarboxylic acid family, has been studied by a combination of in vivo and in vitro "C and 'H NMR spectroscopic techniques. Rats in different nutritional or hormonal situations were infused with [1,12-'3C2]- or [1,2,1 1,12-'3C4]dodecanedioic acid, and the kinetics of "C label appearance as well as the final relative concentrations of metabolic products were measured noninvasively in the liver of the intact rat by "C NMR spectroscopy. Perchloric acid and chloroform/metha- no1 extracts of liver biopsies obtained at the end of the infusion period were further analyzed by high resolu- tion I3C NMR and one-dimensional and two-dimen- sional COSY and J-resolved 'H NMR.
[ l-'3C]- and [ 1,2-"C2]adipic acids were the main end products of the in vivo metabolism of [1,12-'3C2]- or [ 1,2,11, 12-13C4]dodecanedioic acids, respectively, in- dicating that the@-oxidation pathway of medium-chain dicarboxylic acids proceeds in situ monodirectionally. [ l-13C]Adipic acid, the main product of peroxisomal 8- oxidation, could also be detected in situ. This finding, together with the in vivo and in vitro absence of signals characteristic of intramitochondrial oxidation of [ 1-
Clacetyl-coenzyme A, provide a strong evidence sup- porting a predominant contribution of the peroxisomal @-oxidation system to the overall oxidation of these compounds in vivo.
Homonuclear two-dimensional COSY 'H NMR spec- tra of acid extracts from rat liver provided a conven- ient method of analyzing the metabolic repercussions of dicarboxylic acid accumulation, revealing a de- crease in the hepatic concentration of 8-hydroxybutyr- ate and an accumulation of adipic acid and the amino acid L-lysine.
13
In 1934 Verkade and van der Lee (1, 2) described the presence of 1,w medium-chain and short-chain dicarboxylic acids for the first time in urine samples. They proposed that these substances were formed in vivo by the w-oxidation of monocarboxylic fatty acids of the corresponding chain length to allow further bidirectional @-oxidation, a process which was suggested to be more efficient than conventional mono-
* This work was supported by the Swiss National Science Foun- dation Grant 4.889.85.18 (to J. S.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "advertisement" in accord- ance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Supported in part by an EMBO long-term Fellowship ALTF 44- 1986 and Grant 87/1687 from the Fondo de Investigaciones Sanitarias de la Seguridad Social (Spain).
§ To whom correspondence should be addressed. 1 Supported in part by the Deutscher Akademischer Austausch-
dienst and the Dr. Carl Duisberg Stiftung.
directional 8-oxidation. Interest in dicarboxylic acid metabo- lism was later enhanced by the observation of elevated tissue levels (3), as well as increased urinary excretion of medium- chain and short-chain dicarboxylic acids in situations of im- paired or stimulated free fatty acid metabolism such as the Jamaican vomiting syndrome (4, inherited @-oxidation de- fects (5, 6), high fat diets (7), or alloxan- and streptozotocin- induced diabetes (8, 9). In contrast to the very early studies (1, 2), it was shown later that short-chain dicarboxylic acids excreted in the urine were formed preferentially by the w- oxidation of medium chain monocarboxylic acids, followed by one or several @-oxidation steps (10). Moreover, it was dem- onstrated by in vitro experiments that the @-oxidation of dicarboxylic acids could occur both in the mitochondria and in the peroxisomes of rat liver (11).
Several important aspects of medium-chain dicarboxylic acid metabolism still remain unclear, primarily (i) the precise mechanism of their 8-oxidation, (ii) the contribution of the peroxisomal @-oxidation system to the overall oxidative me- tabolism in situ, and (iii) the cause and the metabolic conse- quences of the accumulation of dicarboxylic acids in the liver under different physiological or pathological conditions.
Recently, a variety of novel NMR techniques have made it possible to follow metabolic events noninvasively in situ, either in whole animals or even in human beings. We have previously shown that it is possible to obtain relevant bio- chemical information about the metabolism of short-chain and medium-chain monocarboxylic acids in the intact rat by infusing 13C-labeled fatty acids and following the fate of the 13C label by I3C NMR techniques (12-14). In this report we address some aspects of the metabolism of medium-chain dicarboxylic acids. We have also expanded our previous pro- tocol which involved exclusively 13C NMR spectroscopy with the use of two-dimensional homonuclear J-resolved and chemical-shift correlated 'H NMR. The latter spectra were obtained from perchloric acid extracts of rat liver and pro- vided a wealth of additional metabolic information.
In the following it will be shown that (i) the in situ metab- olism of medium-chain dicarboxylic acids proceeds via a mon- odirectional @-oxidation mechanism, and (ii) the peroxisomes are the predominant site of 8-oxidation for these compounds in vivo. It will also be shown, that the accumulation of dicarboxylic acids observed during fasting and diabetes is due, in part, to a reduced oxidation capacity of the liver for 1,w- dicarboxylic acids. Accumulation of dicarboxylic acids in the liver not only results in a decrease in the 3-hydroxybutyrate concentration (7, 15) but also in an inhibition of the degra- dation of the amino acid L-lysine.
EXPERIMENTAL PROCEDURES
Animals and Infusion Techniques-Adult male Sprague-Dawley rats (250-300 g) were used in all experiments. Well-nourished animals received a standard laboratory rat chow ad libitum; fasted animals
11664

In vivo NMR of 1,w Medium Chain Dicarboxylic Acids 11665
were deprived of food for 48 h prior to their use. Experimental diabetes was induced through a daily intraperitoneal injection of alloxan (100 mg/kg body weight). Animals were used after four days of treatment and had blood glucose concentrations above 20 mM (Hemoglucotest, Boehringer Mannheim GmbH, West Germany) but only traces of ketone bodies in the urine (Ketostix, Miles Laboratories, Ames Di- vision, England).
For the infusion experiments, rats were anesthetized with Nem- butal (25 mg/kg body weight), and the right jugular vein was dissected and cannulated, Shortly after cannulation the rat was placed on a specially designed plexiglass holder such that the surface coil was located directly underneath the liver (13) and the whole arrangement positioned in the center of the homogeneous magnetic field. 0.1 M [1,12-13C2]dodecanedioic acid or [1,2,11,12-'3C4]dodecanedioic acid solutions in saline were then infused at a rate of 4 pmol/min/100 g body weight. This delivery rate was markedly smaller than the max- imal capacity of the liver for the CoA activation of dicarboxylic acids (2 pmols/min/g of liver) (16). In uiuo 13C NMR spectroscopy was performed during the infusion period as described below. At the end of the infusion, the rat abdomen was opened, a small lobe of the liver was excised, and the 13C NMR spectrum of this tissue was recorded immediately. The remaining hepatic tissue was frozen between alu- minum tongs (precooled in liquid nitrogen) and kept a t -80 'C for generally no longer than 24 h until the acid-soluble extracts and the lipid extracts were prepared. The former were obtained from frozen pulverized liver biopsies using the conventional perchloric acid tech- nique (17). Lipids were extracted from liver tissue using the chloro- form/methanol procedure described in Ref. 18. Liver extracts were freeze dried and resuspended in D20 or CDCl&D30D 2 1 prior to high resolution NMR analysis.
In Vivo and in Vitro I3C NMR Spectroscopy-In uiuo I3C NMR spectroscopy (at 20 MHz) was performed in the intact rat using a Bruker Bio-Spec NMR spectrometer equipped with a 1.9 T Oxford Instruments superconducting magnet with a horizontal clear bore of 24 cm. A four-turn surface coil with a diameter of 1.5 cm was used for I3C NMR. The magnetic field was shimmed on the water resonance by tuning the I3C surface coil to the 'H frequency (80 MHz) through the use of an external impedance transformer (12). Signals from subcutaneous fat were minimized by applying a 180" pulse (18 ps) at the center of the coil which nulled signals from the surface tissues and simultaneously allowed the observation of internal tissues. In vivo I 3 C NMR spectra were collected with 4K computer memory, 5400 Hz spectral width (0.371-s acquisition time) and a 1-s recycle time. This recycle time did not allow full relaxation of the carboxyl reso- nances but provided a significant reduction in the overall acquisition time required to obtain an appropriate signal-to-noise ratio as de- scribed in Ref. 13. Normally, 12 blocks of 600 scans were acquired during the infusion period, preceded by one or two blocks of the same duration in the preinfusion interval. The time resolution was there- fore 10 min. 'H decoupling was achieved with a WALTZ-8 pulse sequence using a rat-imaging whole body probe (20). Changes in the intensity of the carboxyl signals were quantitated by comparison with the broad liver carboxyl resonance centered at 176.2 ppm. In vivo 13C chemical shifts were assigned by using the methylene signals of natural abundance fatty acids at 31.0 ppm as an internal standard.
In vitro 13C NMR spectroscopy of liver extracts was performed on a Bruker CXP-300 NMR spectrometer operating at 75.46 MHz. 13C NMR spectra were obtained at 20 'C, pH 7.4, using a broad band gated decoupling sequence and D20 or CDC13 as lock signal. Unless otherwise stated acquisition conditions were: 45" pulse, 16 KHz spectral width, 16K computer memory (0.492-s acquisition time) and 10-s total recycle time. This repetition rate did not induce a significant saturation of the carboxyl resonances of the dicarboxylic acids with respect to the aliphatic ones, as shown in the inset of Fig. 2. Assign- ments were made by addition of internal standards. Chemical shifts were referred to tetramethylsilane at 0 ppm and were calibrated with a coaxial capillary of dioxane at 67.4 ppm.
High Resolution One- and Two-dimensional 'H NMR Spectros- copy-Conventional 'H NMR high resolution spectra (300 MHz) were acquired in D,O at 20 "C, pH 7.4 (uncorrected pH reading), using the same CXP-300 NMR spectrometer. Conditions were: 90" pulses, 3,600 Hz spectral width, 16K data points, and 3-5 recycle time. The residual HDO signal was eliminated by a DANTE presaturating pulse train consisting of 4,000 10" pulses with a 400-ps interpulse delay.
Homonuclear J-resolved two-dimensional spectroscopy was per- formed using the (90" - tl - 180" - tl - acquire(&)), sequence (n = 64) (21), with a sweep width of 27 Hz and 3600 Hz in the tl and t2
dimensions, respectively. A 64.8192 points time domain matrix, resulting in a digital resolution of 0.42 and 0.44 Hz/point in the tl and tz directions of the frequency domain matrix, was used in all cases. In Figs. 5 and 6B, the resulting two-dimensional frequency domain matrix was projected on the F2 dimension resulting in a broad band proton decoupled 'H NMR spectrum (21). Homonuclear chemical shift correlated two-dimensional 'H NMR spectroscopy was performed using a (90" - tl - 45" - acquire(t2))Jn = 256) sequence (COSY 45) (22). Spectra were collected using a 256.1024 points time domain matrix over 3600 Hz along the tl and t2 directions. In both types of two-dimensional spectroscopy, the signal was averaged over a 16-step phase cycle for every t l value, and the residual solvent resonance suppressed as indicated above. Sine-bell window functions were used along the tl and t 2 directions prior to two-dimensional Fourier transformation, the final two-dimensional frequency domain spectrum being further improved by triangular symmetrization. J- Resolved frequency domain matrices were tilted 45" before symme- trization.
Resonance assignments were made by comparing the spectra of the extracts with those of model solutions and were confirmed by internal standard additions. Two-dimensional J-resolved and COSY 'H NMR spectra were recorded before and after the addition of the appropriate metabolite standards at three different pH values of 1, 7.4, and 12 in order to verify the coresonance between the added standard and the original metabolite. Literature data were used as initial estimates for the assignment process (23-25). TSP was added as an internal standard.
Synthesis of '3C-Labeled Dicarboxylic Acids-90% '%-enriched [1,12-'3C~]dodecanedioic acid was synthesized from 1,lO-dibromo- decane by a simultaneous bilateral chain elongation step with Na'3CN/Me2S0 with a 55% yield based on the starting Na13CN. A similar procedure was used for the synthesis of (1,2,11,1213C4]dode- canedioic acid. Briefly, 1,8-dibromooctane was subjected to simulta- neous bilateral carboxylation yielding [l,10-'3C2]sebacic acid. The latter was reduced to 1,lO-decanediol (26) and brominated with PBr3 to [l,10-13C2]dibromodecane. Further simultaneous chain elongation steps with Na'3CN/Me2S0 yielded tetralabeled [1,2,11,12-13C4]dode- canedioic acid with 30% yield based on the initial NaWN. The chemical structures of the synthesized compounds were confirmed by I3C and 'H NMR. Both '%-labeled dodecanedioic acid preparations were chemically pure when analyzed by gas-chromatography, however the [1,2,11,12-'3C4]dodecanedioic preparation contained a 10% con- tamination of [1,12-13C2]dodecanedioic acid as determined by 13C NMR.
Reagents-Metabolite standards were purchased from Sigma Chemie GmbH (Deisenhofen, West Germany) or Aldrich (Steinheim, West Germany). Deuterated solvents and NaI3CN were purchased from CEA (Gif-sur-Yvette, France). TSP was obtained from CIBA- GEIGY AG (Basel, Switzerland). All reagents were of the highest purity available.
RESULTS
In Situ Metabolism of Medium Chain Dicarboxylic Acids as Followed by in Vivo 13C NMR Spectroscopy-Fig. 1 shows representative in vivo I 3 C NMR spectra of the liver of a fasted rat before (A) and during ( B ) the infusion of [1,2-'3C2]do- decanedioic acid. The essential difference between the two spectra is the appearance of two new resonances in the car- boxyl region during the infusion (inset B). The first resonance, centered at 184.7 ppm, is relatively broad (approximately 29- Hz linewidth at half-height) but sufficiently intense to be monitored with a 10-min time resolution. The second reso- nance (-7-Hz linewidth at half-height) appears at 184.1 ppm (cf. arrow in Fig. lB, inset) and can be detected only with longer accumulation times due to its lower intensity. The chemical shift of the two resonances is characteristic of the carboxyl groups of free fatty acids. Esterified fatty acids can be excluded since their carboxyl resonances appear at lower frequencies, in the range of 165-180 ppm. In fact the reso- nances centered at 176.2 and 172.9 in the inset as well as the majority of the remaining resonances of Fig. 1 arise primarily from the natural abundance 13C nuclei of highly mobile tri- glycerides present in the fasted liver (12, 13,27).

11666
A
In vivo NMR of 1,w Medium Chain Dicarboxylic Acids
1 ' 1 ' 1 ' ' 1 ' 1 ' ' l ' l ' l ' l ' l ~ ~ ' l ' l ' ~ ' l ~ l ' ~ ~ '
PPM 17010 14d.O 110.0 80.0 50.0 20.0
B
PPM
1 ' 1 ' 1 ' ' ~ ' " 1 ' " " I ' " " I ' " " 1 ' " " 1 "
PPM
FIG. 1. In vivo "C NMR spectra of a fasted rat liver before (A) and during (B) the infusion of [1,12-1SCz]dodecanedioic acid. In vivo 'H-decoupled 13C NMR spectra were acquired from a 48-h starved Sprague-Dawley rat as indicated under "Experimental Procedures." The substrate was infused via the jugular vein over a period of 120 min. A, spectral acquisition for the period 10 min before to 10 min after the start of the infusion. B, spectral acquisition for the infusion period 20-120 min. Inset, expanded scale of the carboxyl region showing the presence of the adipic acid singlet (arrow) at high field of the 13C signals from the broader composite dicarboxylic acid resonance. 4K free induction decays were zero filled to 8K prior to Fourier transformation. Artificial line broadening A, 3 Hz, B, 1 Hz.
17d.O 140.0 110.0 80.0 50.0 20.0
The new resonances observed in Fig. 1B are directly related to the 13C-enriched infusate and are not derived from natural abundance 13C-esterified fatty acids. This can be demon- strated by repeating the experiment with [ 1,2,11,12-'3C4]do- decanedioic acid. Fig. 2 shows the in uiuo 13C NMR spectrum of the liver of a fasted rat infused with [1,2,11,12-'3C4]do- decanedioic acid acquired under conditions identical to those of Fig. 1B. Fig. 2 also shows, as an inset, the proton-decoupled high resolution 13C NMR spectrum of [1,2,11,12-'3C4]dodec- anedioic acid in saline (pH 7.4, 22 'C) which displayed two doublets centered at 184.7 and 38.0 ppm. These two doublets can be assigned to the C1, C12 and C2, C11 13C-enriched carbons, respectively, and are characterized by a 13C-'3C ho- monuclear coupling constant of 50.8 Hz. This coupling arises from the scalar magnetic interaction between two neighboring 13C nuclei and is not observed in natural abundance 13C fatty acids. Therefore, the observation of 13C-13C homonuclear
184.7 h
500 Hz h-rrrl
1 ' " " 1 " ' " 1 ' " " 1 " ' " I ' " " I " ' " ~ ' ' 170.0 140.0 110.0 80.0 50.0 20.0
PPM FIG. 2. In vivo proton-decoupled "C NMR spectrum (20.1
MHz) of the liver of a fasted rat infused with [l,2,11,12-1sC.] dodecanedioic acid. Acquisition conditions and spectral processing were identical to those of Fig. 1. Insets, high resolution proton- decoupled 13C NMR spectra (75.46 MHz) of a 10-mM solution of [1,2,11,12-'3C4]dodecanedioic acid in saline (22 "C, pH 7.4). Doublets a t 184.7 and 38.0 ppm correspond to the C1, C12 and C2, C11 carbons, respectively. Specific conditions were: 512 scans, 10-s recycle delay, and 2 Hz artificial line broadening.
couplings in the in situ 13C NMR spectrum of the liver, as reflected by the indicated doublets in the carboxyl as well as in the methylene region, provides unambiguous evidence that the new signals observed in Fig. 1B are directly derived from the 13C-enriched infusate.
Further analysis of the in uiuo 13C NMR signals shown in Figs. 1 and 2 was facilitated by high resolution 13C NMR spectra (at 75.46 MHz) of the perchloric acid extracts obtained from the excised livers. Two basic results were obtained (i) the sharp singlet centered at 184.1 ppm in the in uiuo spectrum (labeled by an arrow in Fig. 1B) could be assigned to the carboxyl groups of adipic acid. (ii) The broader resonance centered at 184.7 ppm was shown to have a composite nature, containing not only contributions from the infused dodec- anedioic acid but also from its @-oxidation products, i.e. se- bacic acid and suberic acid. Fig. 3A shows the 13C NMR spectrum of the carboxyl region of a perchloric acid extract obtained from the excised liver of a fasted rat after 120 min of infusion of [1,12-'3C2]dodecanedioic acid (corresponding to Fig. 1). In agreement with the results obtained in vivo, no other signals from low molecular weight, acid-soluble metab- olites were observed in the remaining portion of the spectrum (not shown). The resonance at 184.06 ppm, when recorded as proton-coupled 13C spectrum (not shown), was characterized by two long range coupling constants ('Jc.H, = 5.1 Hz and 3JC.HB = 3.1 Hz) identical to those observed in solutions of authentic adipic acid. Furthermore, this signal coresonated and cotitered (pK,' = 4.75) with an adipic acid internal standard. The remaining signals of Fig. 3 a t 184.60, 184.50, and 184.36 ppm, coresonated and cotitered (pK,' = 4.75) with internal standards of dodecanedioic acid, sebacic acid, and suberic acid, respectively. The increased intensity of the adi- pic acid signal as compared to the resonances of the dicarbox- ylic acids of longer chain length is caused by the higher solubility of adipic acid in perchloric acid solutions. In con- trast when a biopsy of the same liver tissue was extracted

In vivo NMR of 1,w Medium Chain Dicarboxylic Acids 11667
I“L1 *- B
18410 1
PPM FIG. 3. High resolution “C NMR spectra (75.46 MHz) of
perchloric acid extracts obtained from rat livers after infu- sion of [1,12-1SC~]dodecanedioic acid (A) or [ l , 2 , l 1 , l2 -”C4] dodecanedioic acids (B) . Only the carboxyl region of the spectra is shown. Rat livers were excised immediately after the infusion and freeze-clamped between aluminum tongs precooled in liquid nitrogen. Neutralized perchloric acid extracts were prepared as described under “Experimental Procedures.” NMR parameters: 5000 scans, 10-s re- cycle delay, and 2 Hz artificial line broadening.
with chloroform/methanol, the 13C NMR spectrum showed only the singlet characteristic of [1,12-’3C2]dodecanedioic acid. The latter result also excludes the formation of signifi- cant amounts of 13C-labeled triglycerides. It can be concluded, therefore, that the resonance centered at 184.7 ppm in the in vivo spectra is primarily derived from the 13C-labeled infusate with small contributions from sebacic acid and suberic acid. The 184.7 ppm signal in vivo appears as a broadened singlet because of the different chemical shifts of these carboxyl groups.
Similar results were obtained for acid extracts prepared from rat livers infused with [1,2,11,12-’3C4]dodecanedioic acid. Fig. 3B shows the proton-decoupled 13C NMR spectrum of the perchloric acid extract corresponding to the in vivo measurement of Fig. 2. An intense doublet centered at 184.06 ppm (‘Jc.~ = 50.8 Hz), arising from 1,2-13C21abeled adipic acid, together with doublets of smaller intensity derived from con- tiguously I3C-labeled sebacic acid and suberic acid are also observed. These results demonstrate that sebacic acid, suberic acid, and adipic acid are derived from the ‘3C-tetralabeled precursor by a chain-shortening mechanism which conserves part of the original carbon skeleton.
Fig. 4A summarizes the kinetics of 13C label appearance for the dicarboxylic acid signal (at 184.7 ppm) in the liver of a fasted rat infused with [1,12-13C2]dodecanedioic acid. The same experiment was performed with a well-nourished and an alloxan-diabetic rat, and a comparison of the final relative intensities is shown in Fig. 4B. Two more infusions using [1,2,11,12-’3C4]dodecanedioic acid were carried out on a fasted and an alloxan-diabetic animal, respectively (not shown). The kinetics of 13C label appearance were similar in the fed, fasted, and diabetic animals. However, the intensities at the end of the infusion period (Fig. 4B) were appreciably higher in fasted and diabetic animals than in well-nourished rats, indicating a higher accumulation of medium-chain dicarboxylic acids in the liver of fasted and diabetic animals. Similar intensity ratios were observed in I3C NMR spectra of the liver biopsies obtained at the end of the infusion experiments, thereby confirming the values obtained in vivo.
Finally, the present results may be compared with previous investigations on the in situ metabolism of 1,3-l3Cz-labeled short chain monocarboxylic acids under similar experimental conditions (12, 14). In this case no accumulation of the free substrate was observed and fatty acids with shorter chains could not be detected, neither in vivo, nor in the perchloric acid extracts. Instead the in vivo 13C NMR spectra provided
L >
i W a
20 40 60 80 100 120 MINUTES OF lNFUSlON
‘ . , ~ # - m v , m q
CONDITION
FIG. 4. A , time course of the relative intensity of the composite dicarboxylic acid resonance during infusion of [1,12-13C2]dodecane- dioic acid into a fasted rat. Male Sprague-Dawley rats (200-250 g body weight) were infused with a solution of [1,12-’3C~]dodecanedioic acid in saline through the jugular vein (4 pmol/min/100 g body weight) and the in vivo proton-decoupled NMR spectra of the liver recorded in 10-min intervals. E , Relative intensities of the composite dicarboxylic acid resonance at the end of the infusion of [1,12-13C~]dodecanedioic acid for various nutritional and hormonal states. Infusion conditions were those of A. Relative intensities were calculated as indicated under “Experimental Procedures” and repre- sent the mean of the last three spectra corresponding to the period 90-120 min.
11668 I n vivo NMR of 1,w Medium Chain Dicarboxylic Acids
evidence for an intense intramitochondrial [l-'3C]acetyl-CoA' production since the 13C label appeared primarily in [1-"C] acetoacetate, [ l-'3C]3-hydroxybutyrate, [5-'3C]glutamate, [5- '3C]glutamine, and [3,4-'3C]glucose.
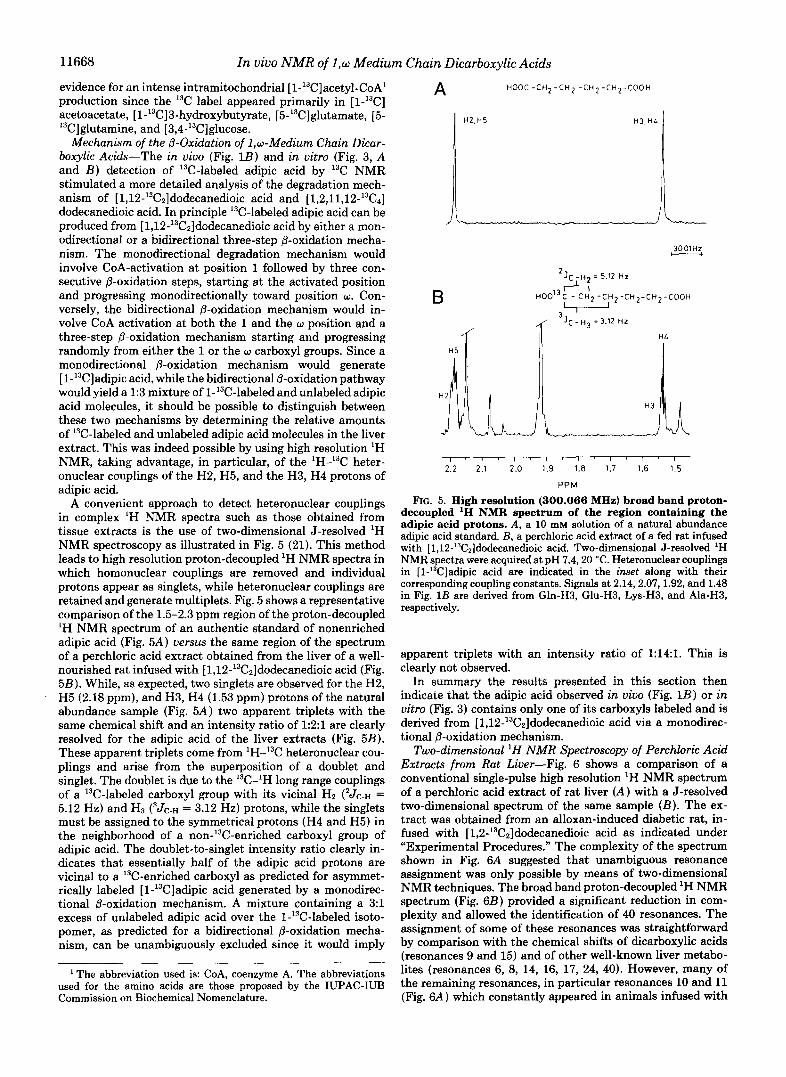
Mechanism of the @-Oxidation of 1,w-Medium Chain Dicar- boxylic Acids-The in oiuo (Fig. 1B) and in vitro (Fig. 3, A and B ) detection of '%-labeled adipic acid by 13C NMR stimulated a more detailed analysis of the degradation mech- anism of [1,12-'3C2]dodecanedioic acid and [1,2,11,12-'3C4] dodecanedioic acid. In principle I3C-labeled adipic acid can be produced from [1,12-'3C2]dodecanedioic acid by either a mon- odirectional or a bidirectional three-step @-oxidation mecha- nism. The monodirectional degradation mechanism would involve CoA-activation at position 1 followed by three con- secutive @-oxidation steps, starting at the activated position and progressing monodirectionally toward position w. Con- versely, the bidirectional @-oxidation mechanism would in- volve CoA activation at both the 1 and the w position and a three-step @-oxidation mechanism starting and progressing randomly from either the 1 or the w carboxyl groups. Since a monodirectional @-oxidation mechanism would generate [ l-I3C]adipic acid, while the bidirectional @-oxidation pathway would yield a 1:3 mixture of l-13C-labeled and unlabeled adipic acid molecules, it should be possible to distinguish between these two mechanisms by determining the relative amounts of 13C-labeled and unlabeled adipic acid molecules in the liver extract. This was indeed possible by using high resolution 'H NMR, taking advantage, in particular, of the 'H-13C heter- onuclear couplings of the H2, H5, and the H3, H4 protons of adipic acid.
A convenient approach to detect heteronuclear couplings in complex 'H NMR spectra such as those obtained from tissue extracts is the use of two-dimensional J-resolved 'H NMR spectroscopy as illustrated in Fig. 5 (21). This method leads to high resolution proton-decoupled 'H NMR spectra in which homonuclear couplings are removed and individual protons appear as singlets, while heteronuclear couplings are retained and generate multiplets. Fig. 5 shows a representative comparison of the 1.5-2.3 ppm region of the proton-decoupled 'H NMR spectrum of an authentic standard of nonenriched adipic acid (Fig. 5A) uersus the same region of the spectrum of a perchloric acid extract obtained from the liver of a well- nourished rat infused with [1,12-'3C2]dodecanedioic acid (Fig. 58). While, as expected, two singlets are observed for the H2, H5 (2.18 ppm), and H3, H4 (1.53 ppm) protons of the natural abundance sample (Fig. 5A) two apparent triplets with the same chemical shift and an intensity ratio of 1:2:1 are clearly resolved for the adipic acid of the liver extracts (Fig. 5B). These apparent triplets come from 'H-I3C heteronuclear cou- plings and arise from the superposition of a doublet and singlet. The doublet is due to the I3C-'H long range couplings of a 13C-labeled carboxyl group with its vicinal Hz ('Jc-H = 5.12 Hz) and H3 ( 3 J ~ . ~ = 3.12 Hz) protons, while the singlets must be assigned to the symmetrical protons (H4 and H5) in the neighborhood of a non-13C-enriched carboxyl group of adipic acid. The doublet-to-singlet intensity ratio clearly in- dicates that essentially half of the adipic acid protons are vicinal to a I3C-enriched carboxyl as predicted for asymmet- rically labeled [l-'3C]adipic acid generated by a monodirec- tional @-oxidation mechanism. A mixture containing a 3:l excess of unlabeled adipic acid over the l-'3C-labeled isoto- pomer, as predicted for a bidirectional @-oxidation mecha- nism, can be unambiguously excluded since it would imply
' The abbreviation used is: CoA, coenzyme A. The abbreviations used for the amino acids are those proposed by the IUPAC-IUB Commission on Biochemical Nomenclature.
A H O O C - C H z - C H z - C H * - C H * - C O O H
HZ,H5 H3. HA
3001Hz - HL
l ~ l ~ ~ ~ l ' l ~ l ~ l ~ l
2.2 2.1 2.0 1.9 1.8 1.7 1.6 1.5
PPM
FIG. 5. High resolution (300.066 MHz) broad band proton- decoupled 'H NMR spectrum of the region containing the adipic acid protons. A, a 10 mM solution of a natural abundance adipic acid standard. B, a perchloric acid extract of a fed rat infused with [1,12-'3C2]dodecanedioic acid. Two-dimensional J-resolved 'H NMR spectra were acquired at pH 7.420 "C. Heteronuclear couplings in [l-'3C]adipic acid are indicated in the inset along with their corresponding coupling constants. Signals at 2.14,2.07,1.92, and 1.48 in Fig. 1B are derived from Gln-H3, Glu-H3, Lys-H3, and Ala-H3, respectively.
apparent triplets with an intensity ratio of 1:14:1. This is clearly not observed.
In summary the results presented in this section then indicate that the adipic acid observed in viuo (Fig. 1B) or in vitro (Fig. 3) contains only one of its carboxyls labeled and is derived from [1,12-'3C2]dodecanedioic acid via a monodirec- tional @-oxidation mechanism.
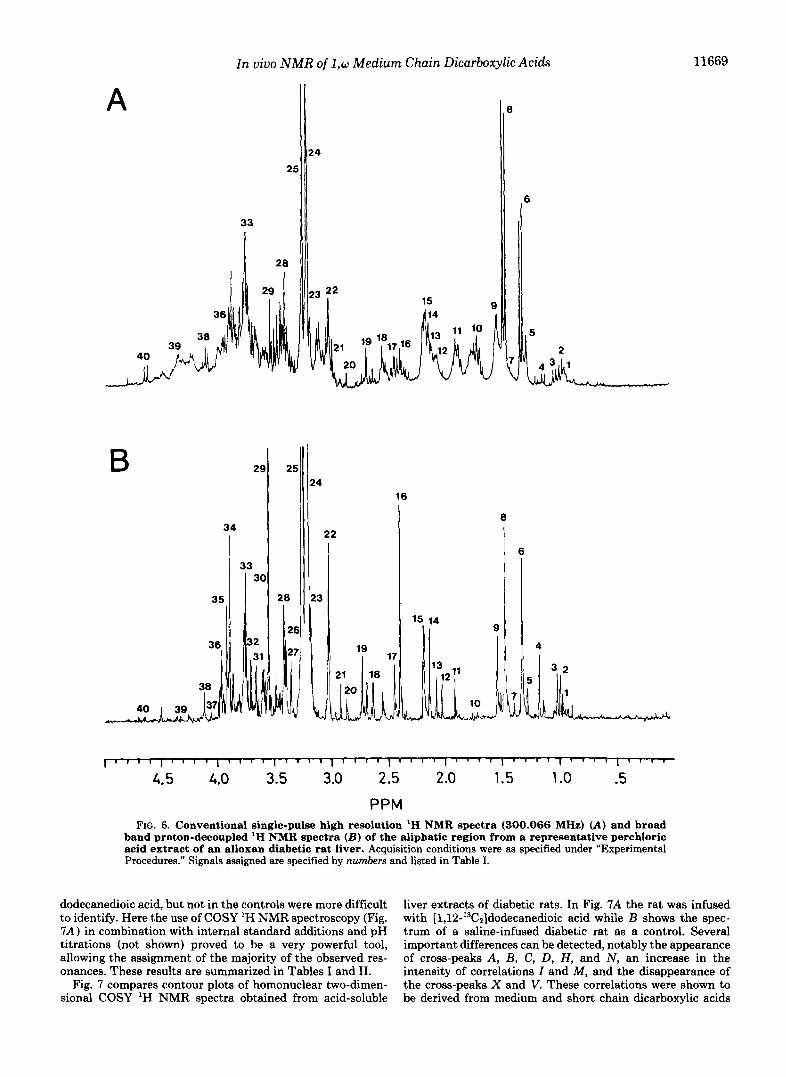
Two-dimensional ' H NMR Spectroscopy of Perchloric Acid Extracts from Rat Liver-Fig. 6 shows a comparison of a conventional single-pulse high resolution 'H NMR spectrum of a perchloric acid extract of rat liver (A) with a J-resolved two-dimensional spectrum of the same sample ( B ) . The ex- tract was obtained from an alloxan-induced diabetic rat, in- fused with [1,2-'3C2]dodecanedioic acid as indicated under "Experimental Procedures." The complexity of the spectrum shown in Fig. 6A suggested that unambiguous resonance assignment was only possible by means of two-dimensional NMR techniques. The broad band proton-decoupled 'H NMR spectrum (Fig. 6B) provided a significant reduction in com- plexity and allowed the identification of 40 resonances. The assignment of some of these resonances was straightforward by comparison with the chemical shifts of dicarboxylic acids (resonances 9 and 15) and of other well-known liver metabo- lites (resonances 6, 8, 14, 16, 17, 24, 40). However, many of the remaining resonances, in particular resonances 10 and 11 (Fig. 6A) which constantly appeared in animals infused with
In vivo NMR of 1,o Medium Chain Dicarboxylic Acids 11669
A 24
25 '
33
6 29
34
33 I
25 I 2
22
I
4 16
8
1 6
6
1 " " 1 1 ' " 1 1 " " " ' " " " ' ' " " " ' " ' ' ' ~ 1 ' ' '
4.5 4.0 3.5 3 .O 2.5 2 .o 1.5 1 .o .5 PPM
FIG. 6. Conventional single-pulse high resolution '€3 NMR spectra (300.066 MHz) (A) and broad band proton-decoupled 'H NMR spectra (B) of the aliphatic region from a representative perchloric acid extract of an alloxan diabetic rat liver. Acquisition conditions were as specified under "Experimental Procedures." Signals assigned are specified by numbers and listed in Table I.
dodecanedioic acid, but not in the controls were more difficult to identify. Here the use of COSY 'H NMR spectroscopy (Fig. 7A) in combination with internal standard additions and pH titrations (not shown) proved to be a very powerful tool, allowing the assignment of the majority of the observed res- onances. These results are summarized in Tables I and 11.
Fig. 7 compares contour plots of homonuclear two-dimen- sional COSY 'H NMR spectra obtained from acid-soluble
liver extracts of diabetic rats. In Fig. 7A the rat was infused with [1,12-'3C2]dodecanedioic acid while B shows the spec- trum of a saline-infused diabetic rat as a control. Several important differences can be detected, notably the appearance of cross-peaks A, B, C, D, H, and N , an increase in the intensity of correlations I and M, and the disappearance of the cross-peaks X and V. These correlations were shown to be derived from medium and short chain dicarboxylic acids

11670 In vivo NMR of 1,w Medium Chain Dicarboxylic Acids
I " " " " ' l " " " " ' l " " " " ' 1 " " " " ' 1 " " " " ' 5.0 4.0 3.0 2.0 1 .o
PPM
6 r 4 .H 1.0
2.0
1 3.0
1 4.0
- 5.0 PPM
FIG. 7. Contour plots of two-dimensional chemical shift correlated spectra (COSY 45) of perchloric acid extracts of alloxan diabetic rat livers. A, diabetic rat liver infused with [1,12-'3C2]dodecanedioic acid (same extract as Fig. 6). B, saline infused control. Specific correlations are noted with capital letters. Assignments are listed in Table 11.
(correlations A and D), 3-hydroxybutyrate (correlations X and V ) , and the amino acid L-lysine (correlations B, C, I, and M ) as indicated below:
D A v x *I-+ +* OH I B C M
O O C " C H 4 H r C H - R C H & , H 4 H 4 O O -
l-%i-%l-+& +NHr-CH?-CHr-CHrCH&H4OO-
&Ha
The identification of lysine which must be assigned to resonances 9 and 10 (Fig. 7A) was confirmed by excluding other hepatic metabolites of similar structure such as the amino acids ornithine, &hydroxylysine or a-aminoadipic acid, the ketoacid a-oxoadipic acid, or the polyamines spermine and spermidine (cf. Table 111). Although some proton chemical shifts of these compounds are similar to those of L-lysine (see Table 111), L-lysine and L-saccharopine are the only hepatic metabolites which exhibit the characteristic of H3-H4-H5 connectivity at 1.90-1.45-1.72 ppm. However, the HZ-proton resonance of the glutamate moiety in saccharopine is shifted 0.15 ppm upfield to that of the HZ-proton of free glutamate. Therefore, the H2-H3 correlation of the glutamate in sac- charopine (3.61-2.08 ppm) can be distinguished quite readily from that of free glutamate H2-H3 (3.76-2.05 ppm) and free lysine (3.75-1.90) in a two-dimensional COSY spectra. This correlation which is characteristic of saccharopine was not observed in the present study. Hence, it can be concluded that correlations B, C, I, and M are derived from the amino acid L-lysine. Correlations E, Q , and U could not be assigned.
No difference between the infused and control one-dimen- sional and two-dimensional COSY 45 spectra were detected in the aromatic region which showed mainly resonances from the adenine (H2, H8) and ribose (Hl') protons of the adenine
nucleotides (not shown). In particular no increase in the tryptophan concentration of the dodecanedioic acid-infused livers as reflected by the appearance of indole-ring signals (H4 at 7.72 ppm, H7 at 7.54 ppm, H2 at 7.31 ppm, and H5- H6 at 7.25 ppm) could be demonstrated.
Finally, similar comparisons with that shown in Fig. 7 between contour plots of COSY 45 spectra of infused and control animals, were performed with fasted and well-nour- ished animals and the results are summarized in Table IV. Some cross-peaks which appear with low intensities in Fig. 7 were not consistently found in all spectra of dodecanedioic acid-infused animals or were present also in the remaining controls (cf. peak H). Analysis of their potential physiological significance was not further pursued. In general infusion of 13C-bi- or tetralabeled dodecanedioic acid resulted in the appearance of intense cross-peaks from adipic acid and lysine and the disappearance of the 3-hydroxybutyrate correlations.
DISCUSSION
Accumulation of 1,w-Dicarboxylic Acids in Rat Liver-The present experiments were designed to gain in situ information on the metabolism of medium-chain 1,w-dicarboxylic acids in rat liver. Previous experimental protocols involved the oral administration of different diets, of normal, or of radioactively labeled dicarboxylic acids (7,10,15), followed by the detection of metabolic products in the urine by either gas chromatog- raphy-mass spectrometry or scintillation counting. Here we have applied a different method, i.e. a continuous intravenous infusion of '3C-multilabeled dodecanedioic acid, and have followed noninvasively the hepatic metabolism of this com- pound in different hormonal or nutritional situations.
In contrast to our previous studies on the in situ metabolism of short-chain and medium-chain monocarboxylic acids (12- 14), the infusion of dodecanedioic acid led to an intrahepatic accumulation of medium-chain and short-chain dicarboxylic acids, which was independent of the nutritional or hormonal

In vivo NMR of 1,o Medium Chain Dicarboxylic Acids 11671
TABLE I Chemical shifts and multiplicities of some 'H NMR obseruable
diabetic rats infused with 11,12-'3Culdodecaned~~c acid as determined metabolites in perchloric acid extracts obtained from the liuer of
from two-dimensional homonuclear J-resolved spectroscopy Resonance numbers correspond to those shown in Fig. 6. Chemical
shifts were determined versas internal TSP at 20 "C, pH 7.4, in the broad band proton decoupled 'H NMR spectrum shown in Fig. 6B. Assignments were performed as indicated under Experimental Pro- cedures. (a), singlet; (d), doublet; (t), triplet; (dd), doublet of doubiets; (q), quartet; (m), multiplet.
Resonance no.
Chemical Shlft
(multiplicity) Assignment
1 0.97 (d) Leu-H5 2 3 4
6 5
7 8
10 9
11
13 12
14
15 16 17 18
20 19
21 22
23 24
25 26 27
28 29 30b
31 32
33'
34
35
36d
37 38
39
40
0.99 (d) 1.01 (d)
1.27 (m) 1.17 (d)
1.32 (d) 1.45 (m)
1.53 (t)" 1.47 (d)
1.72-1.78 (m)
. ~ . ~~,
1.90 (m) 2.02 (m) 2.05 (m) 2.12 (m)
2.18 (t)" 2.39 (m) 2.44 (m) 2.63 (t)
2.69 (dd) 2.86 (dd) 2.92 (dd) 3.01-3.05 (t)
3.18 (t) 3.20-3.23 (8) + (m)
3.25 ( 8 )
3.28 (t) 3.35 (t)
3.43 (t) 3.55 (8)
3.58-3.60 (d)
3.69 (m) 3.70 (m)
3.14-3.76-3.78 (dd) + (t) +
3.89 (m) (q)
3.94 (m)
3.96-3.97-3.98
4.05 (m) (m)
4.11 (9) + (m)
4.15-4.32 (m)
Leu-H5', VaLH4 Val-H4' 3-Hydroxybutyrate H4 Dicarboxylic acid H4 Lac-H3, Thr-H4
Ala-H3 Adipic acid H3, H4 Lys-H5,1,4-Butanediamine
moiety in spermine and spermidine H2, H3
Lys-H4
Lys-HB Pro-H3
Gln-H3,3-propylamine G b H 3
moiety in spermine and
Adipic acid H2, H5 spermidine HZ
Gln-H4 2-Mercaptoethylamine from
Asp-HB Asp-HB' Cys in GSSG H3 3-Propylamine moiety H1,
GIu-H~
COA (HS-CHz-)
H3 and 1,4-butanediamine moiety H1, H4 in sperm- ine and spermidine, Lys- H6
Ethanolamine in GPE HZ (MeIaN'- in choline, phos-
phorylcholine and GPC, Cys in GSSG H3' Trp-HI, glucose pH2
Tau N-CH2- (Me),N+- in carnitine
2-Mercaptoethanolamine
Tau S-CH2 from CoA (NH,-CHz-)
GLv-H~ Thr-HZ, glycerol moiety in
Glycerol in GPC H2 Choline moiety in GPC N-
GPE H2
Glu-H2, Gln HZ, Lys-H2, CHz-
Ala-Hz
Choline moiety P-O-CH2 and glycerol moiety H3 in GPC
Ethanolamine moiety in
Cys-HZ, glycerol in GPC and GPE P-0-CHZ
GPE H3 Tv-HZ Lac-HZ, 3-Hydroxybutyrate
H3 Glycerol in GPC and GPE
Thr-H3 H1
4.65 id) . , ~~ .~
Glucose pH1 "Heteronuclear coupling triplet originated from the superposition of a doub-
let arising from 'J and 'J 13C-'H heteronuclear couplings and the corresponding singlet where homonuclear coupling has been removed and no heteronuclear coupling is detectable ('C'H moiety).
Four resolved resonances, two unassigned. 'Five resolved resonances, one unassigned.
Five resolved resonances, two unassigned.
situation, This was remarkable since the concentration of the infusate was only half of that previously used for the mono- carboyxlic fatty acids. It can be concluded, therefore, that the in situ @-oxidation of medium-chain and short-chain dicar- boxylic acids proceeds at a slower rate than that of short- chain and medium-chain monocarboxylic fatty acids. One possible explanation for this difference could be related to different mitochondrial transport rates for the two types of fatty acids. Medium-chain and short-chain monocarboxylic fatty acids can enter the inner mitochondrial space quite readily, bypassing the carnitine transferase step. In contrast dodecanedioic acid must first be activated with CoA (16) and then esterified with carnitine (28) before it can be transported into the mitochondrial matrix. These processes, which are thought to play an important regulatory role and are probably the rate-limiting step in the hepatic degradation of monocar- boxylic fatty acids (29), have been shown for 1,w-dicarboxylic acids to proceed only at 10% of the rate found for the corre- sponding monocarboxylic acids (28). Alternatively, one must consider the possibility that &oxidation of dicarboxylic acids occurs primarily not in the hepatic mitochondria but in a different cellular organelle. Evidence for a peroxisomal B- oxidation of 1,w-dodecanedioic acid in situ will be presented below.
As described in Fig. 4, the hepatic concentration of dicar- boxylic acids at the end of the infusion as reflected in the intensity of the composite dicarboxylic acid resonance was found to be higher in the fasted and diabetic animals than in the well-nourished rats. These differences between different hormonal or nutritional situations parallel those observed in Ref. 7 and 10 and could be caused by (i) an enhancement of w-oxidation during fasting and diabetes (30-32), (ii) a decrease in the @-oxidation rate for dicarboxylic acids, or (iii) a com- bination of both processes. The identical infusion conditions used with the different nutritional or hormonal situations analyzed in this study and the fact that the accumulated dicarboxylic acids were '3C-Iabeled (Figs. 4 and 5 ) and there- fore derived from the infusate, eliminate the possibility of different delivery rates or of an increased production of dicar- boxylic acids by w-oxidation of endogenous monocarboxylic fatty acids. Thus, it can be concluded that the increased accumulation of 1,w-dicarboxylic acids observed in situ during fasting and diabetes (Fig. 423) is due to a decrease in the p- oxidation capacity of the liver for these substances.
It is generally assumed that 1,w-dicarboxylic acids are me- tabolized in the liver by the same or at least by a very similar &oxidation enzyme system as used for monocarboxylic acids, regardless of the actual intracellular &oxidation site. How- ever, the kinetic constants for several key @-oxidation en- zymes like the microsomal fatty acyl-CoA synthase (33) and dicarboxylyl-CoA synthase (16) or the mitochondrial carni- tine activation and transport system (28) are more favorable for the 8-oxidation of monocarboxylic fatty acids than for that of dicarboxylic fatty acids. During fasting and diabetes a drastic increase in the plasma concentration of monocar- boyxlic fatty acids is observed which is due to an increased lipolysis in the adipose tissue (34). Therefore, the competition between mono- and dicarboxylic acids for the same p-oxida- tion system could then lead to the observed accumulation of the less-favored dicarboxylic acids.
In Situ Metabolic Compartmentatwn and Mechanism of the @-Oxdation of 1,w-Medium Chain Dicarboxylic Acids-Short- chain monocarboxylic fatty acids are oxidized exclusively in the liver mitochondria. Previous 13C NMR experiments with infused 1,3-13Cz-labeled butyrate or octanoate have demon- strated that the main Y!-labeled products observable in uiuo

11672 In vivo NMR of 1,w Medium Chain Dicarboxylic Acids
TABLE I1 Assignments of the correlations observed in two-dimensional homonuclear chemical shift-correlated 'H NMR
spectra of perchloric acid extracts from diabetic rats infused with [l,12-13CJdodecaned~ic acid or saline Cross-peaks are identified by the capital letters used in Fig. 7, A and B. Numbers refer to those of resonances
from Fig. 6, A and B. The assignments were performed as described in Table I. Chemical shifts were measured versus internal TSP, at 20 "C, pH 7.4. Cross-peak Chemical
identifi- shifts cation correlated
Peak no. in Fig. 6 Assignment
A 1.53-1.27 9-5 Dicarboxylic H3-H4 B 1.45-1.72 7-10 LYS H4-H5 C 1.45-1.90 7-1 1 LYS H4-H3 D 1.53-2.18 9-15 Dicarboxylic H3-H2 E 1.47-3.23 F G H I
8-24 3.78-1.47 33-8 Ala H2-H3
Lac H2-H3 4.11-1.32 38-6 4.32-1.32 39-6 Thr H3-H4 1.78-3.02 10-22 Lys H5-H6, 1,4-Butanediamine
moiety in spermine and spermidine H2-H1, H3-H4
J 2.05, 2.12-2.39, 13, 14-16, 17 Glu H3-H4, Gln H3-H4 2.44
K 2.12-3.05 14-22 3-Propylamine moiety in spermine and spermidine HZ-HI, H2-H3
L 2.05, 2.12-3.76, 13,14-33 Glu H3-H2, Gln H3-H2
M N
3.77 3.75-1.90 33-11 LYS H2-H3 2.63-3.35 18-27 2"erca~toethvlamine from
2.92-3.23 3.18-3.94 3.21-4.11 3.23-4.65 4.18-3.58
4.32-3.69
4.32-4.05 4.11-1.17 4.11-2.35
21-24 23-35 24-38 24-40 39-30
39-31
39-37
38" 38-4
CoA Hl-H2 Cys H3-H3' in GSSG Ethanolamine in GPE Hl-H2
Glucose PH2-pH1 Glycerol moiety in GPE H1-
Glycerol moiety in GPC H1- H2
H2
3-Hydroxybutyrate H3-H4 3-Hydroxybutyrate H3-H2
The signal from 3-hydroxybutyrate H2 (2.35 ppm) is not present in the neutralized perchloric acid extract livers of animals infused with dodecanedioic acid.
TABLE I11 'H NMR chemical shifts and multiplet structures of L-lysine and structurally related compounds of rat liver
Chemical shifts were determined in 50 mM solutions of the appropriate authentic compounds in 0.1 M NaCI/ DzO, at 25 "C, pH = 7.4, and are referred to internal TSP at 0 ppm. a, axial; e, equatorial; t, triplet; q, quartet; dd, doublet of doublets; m, multiplet.
Proton chemical shift (multiplicity) Compound
H1 H2 H3 H4 H5 H6
L-LYS L-Pipecolic acid
L-Saccharopine Glu moiety Lys moiety
L-6-Hydroxylysine
L-Orn Spermine and spermidine
1,4-Butanediamine 3-Pro~vlamine moietv
3.75t 3.57(.,"
3.61' 3.76' 3.80q
3.78'
3.11-3.16" 1.78" 3.11-3.16" 2.12"
1.90" 1.55-1.73(,)" 2.20C)"
2.08q 1.92" 2.04" 1.93" 1.93"
1.78" 3.11-3.16"
1.45" 1.72" 3.02' 1.55-1.73(.)" 1.55-1.73(.)" 3.02(.)" 1.84-1.90(.)" 1.84-1.90(,)" 3.40(.,"
2.40t 1.49" 1.77" 3.06'
2.93" 1.46-1.76" 3.88" 3.16dd
1.81" 3.06'
3.11-3.16"
are either those related to the Krebs cycle such as glutamate, glutamine, and C03H- (fasted rats) or the ketone bodies acetoacetate and 3-hydroxybutyrate (diabetic rats) (12-14). In both cases intramitochondrially generated [l-'3C]acetyl- CoA from the fatty acid @-oxidation is the key intermediate entering either the tricarboxylic acid cycle or the ketogenesis pathway.
@-Oxidation of [l,w-13Cz]dicarboxylic acids also produces
[l-'3C]acetyl-CoA. However, no 13C labeling in the Krebs cycle-related metabolites nor of the ketone bodies could be detected either in vivo or in vitro in the present experiments. This finding provides a first clue indicating that the in situ @-oxidation of dicarboxylic acids takes place primarily in the extramitochondrial space. It has been shown that @-oxidation of mono- and dicarboxylic acids in rat liver can occur not only in the mitochondria but also in the peroxisomes (11, 35).

In vivo NMR of 1,w Medium Chin Dicarboxylic Acids 11673
TABLE IV Summary of cross-peaks observed in the contour plots of homonuclear two-dimensional COSY 45 'H N M R spectra
of perchloric acid extracts from rat livers obtained from saline and dodecanedioic acid infused animals under different nutritional conditions
Two-dimensional COSY 45 spectra were acquired from neutralized perchloric acid extracts of fed and fasted rat livers of animals infused with [1,12-'3C2]dodecanedioic acid under the conditions specified under "Experimental Procedures" and compared with those of saline infused control rats as illustrated in Fig. 7 A and B. (+) indicates the presence of a specific cross-peak in the spectrum and (-) indicates its absence. The capital letter notation is the same as in Fig. 7 and Table 11.
Condition/Cross-peak A B C D E F G H I J K L M N 0 P Q R S T U V X Fed rat dioic . . . . . . . . . . . . . . . . . . . . . . Fed rat control Fasted rat dioic
+ + + + + + + + + - + + + + + + + + + + + + + + + + - + + + + + + + - + + - + + "
Fasted rat control - - - - + + + + + + + + + - + + + + - + + + + ""
Recently, the apparent kinetic constants and the chain length dependence of &oxidation for dicarboxylic acids in isolated liver mitochondria and purified peroxisomal preparations have been determined (36). It was demonstrated that 1,o- dodecanedioic acid can be oxidized to succinyl-CoA if the p- oxidation occurs in the mitochondria, but only to adipic acid if it takes place in the peroxisomes. The same study also showed that intact isolated rat liver mitochondria could not oxidize exogenous adipic acid appreciably, probably because an inability to transport this substrate through the inner mitochondrial membrane. In the present study [l-13C]adipic acid, the final product of the peroxisomal @-oxidation of [1,12-'3C2]dodecanedioic acid was the shortest chain dicarbox- ylic acid detectable in situ by 13C NMR. This observation provides further support for a major contribution of the peroxisomal @-oxidation system in situ since (i) in vivo NMR signals are known to be derived from cytosolic metabolites and (ii) [l-13C]adipic acid produced by peroxisomal @-oxida- tion would be expected to accumulate in the cytosol because of the inability of hepatic mitochondria to metabolize exoge- nous adipic acid.
The results presented in Fig. 5 also provide new and direct information about the mechanism of @-oxidation of dicarbox- ylic acids. The 13C-labeling pattern can only be explained by a monolateral &oxidation mechanism and a significant con- tribution from a bilateral @-oxidation pathway, as initially proposed by Verkade (2) can be excluded. A similar conclusion was derived indirectly in Ref. 15 from calculations based on the 14C labeling of plasma glucose after the administration of l-14C or uniformly labeled [14C]palmitic acids to rats. How- ever, in these calculations it was assumed that the 14C labeling of plasma glucose directly reflected the labeling of a unique pool of oxaloacetate and that palmitic acid was exclusively and completely oxidized in the mitochondria to eight acetyl- CoA units. Both assumptions are difficult to reconcile with our present knowledge of kinetically different oxaloacetate pools in rat liver (37) or with the incomplete oxidation of ['*CC]palmitic acid by isolated rat hepatocytes (38) and per- oxisomal preparations (35). Thus, the direct estimates of 13C labeling in short-chain dicarboxylic acids described in this study should provide a more reliable procedure to differentiate between the two alternative @-oxidation mechanisms.
Metabolic Consequences of the Accumulation of Medium- chain and Short-chain Dicarboxylic Acids in Rat Liver as Revealed by Two-dimensional NMR Spectroscopy-Few stud- ies have reported two-dimensional spectra of tissue extracts (24, 39-41), and the possibility of obtaining metabolic infor- mation from this type of spectroscopy has not been explored in detail. This may be due, in part, to difficulties found in the spectral interpretations because of incomplete assignments. Therefore, an important aspect of the present study has been
DIETARY INTAKE PROTEOLYSIS
I \ L- LYSINE
WDECANEDIOIC INFUSION
ACID
1 SACCHAROPINE
L-PIPECOLIC ACID SEBACIC ACID
1 E-SEMIALDEHIDE a-AMINOADIPIC SUBERIC ACID
1 1 AMlNOADlPlC
ACID L-TRYPTOPHANE
h.
\a-OXOADIPIC ACID
1 GLUTARYL-CoA
1-0- GLUTARYL-COA DEHYDROGENASE ~~
I DEFICIENCY
3-HYDROXYBUTYRYL-CoA - 3-HYDROXYBUTYRATE 1 1
1 2 ACETYL-CoA
FIG. 8. Scheme of the possible metabolic interactions be- tween medium chain dicarboxylic acid &oxidation and the hepatic pathway of lysine degradation. Potential inhibitory sites for adipic acid are indicated by the e sign.
to analyze systematically the 'H J-resolved and COSY 45 spectra of perchloric acid extracts of rat liver, to provide reliable assignments, and to analyze the metabolic informa- tion obtainable from this kind of spectroscopy.
Fig. 8 provides the framework in which to discuss the metabolic interactions caused by the infusion of a medium- chain dicarboxylic acid. As indicated in Fig. 7 and Tables I1 and IV the most characteristic effect of [ 1,12-'3C2]dodecane- dioic acid infusion is a decrease in the concentration of 3- hydroxybutyrate and an increase in the hepatic concentra- tions of adipic acid and lysine. As shown in the metabolic chart of Fig. 8, L-lysine is degraded in rat liver primarily through the saccharopine pathway (42) to glutaryl-CoA and further to two molecules of acetyl-coA (43). This pathway involves the formation of a-aminoadipic semialdehyde, a- aminoadipic acid, and a-oxoadipic acid as key intermediates prior to glutaryl-CoA formation. The alternative pipecolate pathway has been found to be of very minor importance in rat liver (42).
Since L-lysine is an essential amino acid only produced in the mammalian liver by proteolysis or dietary intake, its accumulation after infusion of a medium-chain dicarboxylic acid can only be due to an inhibition of its degradation caused

11674 In vivo NMR of 1,w Medium Chain Dicarboxylic Acids
by the infused dicarboxylic acid. Since the concentration of 3-hydroxybutyrate, one of the products of lysine metabolism decreases, and L-tryptophan, which enters the same pathway at the level of a-oxoadipic acid does not accumulate, it can be concluded that the inhibitory interaction(s) should take place at a step prior to the formation of 3-hydroxybutyrate and a- oxoadipic acid (see Fig. 8). A competitive inhibition of adipic acid at the level of a-aminoadipic acid or a-aminoadipic semialdehyde is proposed in Fig. 8.
No information is available on the rate-limiting steps or close to equilibrium segments of hepatic lysine degradation. The present results together with those of Przyrembel et al. (44) and Swetmann et al. (45) who also found an accumulation of lysine in patients with glutaryl-CoA dehydrogenase type I1 deficiency, can provide some insight into this question. Inhi- bition of one or several steps of the lysine degradation path- way prior to glutaryl CoA formation results in an accumula- tion of lysine, with no detectable presence of any of the metabolic intermediates between lysine and glutaryl-CoA. This strongly suggests that this segment of lysine metabolism is reversible and close to equilibrium in uiuo.
Concluding Remarks-In uiuo I3C NMR spectroscopy of rat liver as described in this and earlier studies (12-14) has demonstrated distinct differences between the metabolism of monocarboxylic acids and dicarboxylic fatty acids in situ. 13C- Labeled short-chain monocarboxylic acids are primarily oxi- dized in the mitochondria with [l-'3C]acetyl-CoA as the key intermediate. Depending on the hormonal and physiological situation [1-'3C]acetyl-CoA is either directed into the Krebs cycle (fasted rats) or into the ketogenic pathway (diabetic rats). Accordingly, metabolites of the Krebs cycle are detected for fasted rats, whereas 13C-labeled ketone bodies are predom- inant in diabetic rats. In contrast the primary oxidation site for dicarboxylic acids in situ are the liver peroxisomes. The main degradation products of infused 13C-labeled dodecane- dioic acid are dioic acids of shorter chain length, with adipic acid being the minimum chain length detected. 13C-Label was not detected in other metabolites. @-Oxidation of dicarboxylic acids gave very similar results for fasted and diabetic animals, but the rate of oxidation was reduced compared to well-fed rats.
The use of multilabeled dicarboxylic acids in combination with two-dimensional J-resolved 'H NMR spectroscopy pro- vides unambiguous evidence that the p-oxidation of these compounds in situ follows a unidirectional mechanism. Fi- nally, the infusion of dicarboxylic acids also influences the concentration of other liver metabolites. In particular the analysis of COSY 45 two-dimensional NMR spectra clearly shows an increase in the L-lysine and a decrease of 3-hydrox- ybutyrate concentrations.
Further experiments on the in situ metabolism of long- chain monocarboxylic acids constitute a natural extension of these studies.
Acknowledgment-We are deeply indebted to P. Ganz for the competent synthesis of T!-labeled dicarboxylic acids.
1.
2. 3. 4. 5.
REFERENCES Verkade, P. E., and van der Lee, J. (1934) Biochemical J. 28,31-
Verkade, P. E. (1938) Chem. Ind. 5 7 , 704-711 Mortensen, P. B. (1986) Biochim. Biophys. Acta 878, 14-19 Tanaka, K. (1972) J. Biol. Chem. 247,7465-7478 Gregersen, N., Kolvraa, S., Rasmussen, K., Mortensen, P. B.,
Divry, P., David, M., and Hobolth, N. (1983) Clin. Chim. Acta
40
132,181-191
6. Gregersen, N. (1985) Scad. J. Clin. Lab. Invest. 45 (Suppl. 174)
7. Mortensen, P. B. (1981) Biochim. Biophys. Acta 6 6 4 , 335-348 8. Mortensen, P. B. (1980) Biochim. Biophys. Acta 6 2 0 , 177-185 9. Mortensen, P. B. (1981) Biochim. Biophys. Acta 664,349-355 10. Mortensen, P. B., and Gregersen, N. (1981) Biochim. Biophys.
11. Mortensen, P. B., Kdvraa, S., Gregersen, N., and Rasmussen, K.
12. Cross, T. A., Pahl, C., Oberhiinsli, R., Aue, W. P., Keller, U., and
13. Pahl-Wostl, C., and Seelig, J. (1986) BiochemistTy 25,6799-6807 14. Pahl-Wostl, C., and Seelig, J. (1987) Biol. Chem. Hoppe-Seyler
15. Wada, F., and Usami, M. (1977) Biochim. Biophys. Acta 487,
16. Vameca. J.. De Hoffmann. E.. and Van Hoof, F. (1985) Biochem.
1-60
Acta 666,394-404
(1982) Biochim. Biophys. Acta 713,393-397
Seelig, J. (1984) Biochemistry 23,6398-6402
368,205-214
261-268
J. 230,683-693 . .
17. LOWN. 0. H.. and Passonneau. J. (1972) in A Flexible System of Eniymatic Analysis, Academic Press, New York
18. Kates, M. (1972) in Techniques of Lipidology, Isolation, Analysis and Identification of &ids, PP. 349-350, North Holland Pub- . " lishing Co:, New York -
19. Shaka, A. J., Keeler, J., and Freeman, R. (1983) J. Magn. Res.
20. Cross, T. A., Muller, S., and Aue, W. P. (1985) J. Magn. Res. 6 2 , 53,313-340
87-98 21. Aue, W. P., Karhan, J., and Ernst, R. R. (1976) J. Chem. Phys.
22. Bax, A., and Freeman, R. (1981) J. Magn. Res. 44,542-561 23. Wuthrich, K. (1976) in NMR in Biological Research: Peptides and
Proteins, pp. 42-50, North Holland/American Elsevier, New York
24. Fan, T. W.-M., Higashi, R. M., Lane, A. N., and Jardetzky, 0. (1986) Biochim. Biophys. Acta 8 8 2 , 154-167
25. Cerdan. S.. Parrilla, R., Santoro, J., and Rico, M. (1985) FEBS
64,4226-4227
Lett. '187, 167-172 . .
26. Nystrom, R. F., and Brown, W. G. (1947) J. Am. Chem. SOC. 6 9 ,
27. Canioni, P., Alger, J. R., and Shulman, R. G. (1983) Biochemistry
28. McGarry, J. D., and Foster, D. W. (1980) Annu. Reu. Biochem.
29. Pettersen. J. E. (1973) Biochim. BioDhvs. Acta 306. 1-14
2548-2549
22,4974-4980
49,395-420
30. Nilsson, A., Arey, H., Pedersen, J: I.; and Christiansen, E. N.
32. Bjorkhem, I. (1976) J. Bwl. Chem. 251,5259-5266 31. Bjorkhem, I. (1973) Eur. J. Biochem. 40,415-422
33. Tanaka, T., Hosaka, K., Hoshimaru, M., Numa, S. (1979) Eur.
34. Newsholme, E. A., and Leech, A. R. (1983) in Biochemistry for the Medical Sciences, pp. 274-293, John Wiley & Sons, New York
35. Lazarow, P. B., and de Duve, C. (1976) Proc. Natl. Acad. Sci. U.
36. Kolvraa, S., and Gregersen, N. (1986) Biochim. Biophys. Acta
37. Katz, J., and Grunnet, N. (1979) in Techniques in Metabolic Research (Kornberrr. H. L.. ed), part I, B208, pp. 1-18, Elsevier/
. .
(1986) Biochim. Bwphys. Acta 879,209-214
J. Biochem. 98,165-172
S. A. 73,2043-2046
876,515-525
North Holland, New York . . ..
38. KondruD. J.. and Lazarow, P. B. (1982) Annu. N. Y. Acad. sci. 386, hO4-405
39. Evanochko, W. T., Sakai, T. T., Thian, CNG., Rama Krishna, N., Hyun, D. K., Zeidler, R. B., Ghanta, V. K., Wallace- Brockman, R., Schiffer, L. M., Braunschweiger, P. G., and Glickson, J. D. (1984) Biochim. Biophys. Acta 806,104-116
40. Arus, C., Westler, W. M., Barany, M., and Markley, J. (1986) Biochemistry 25,3346-3351
41. Cross, K. J., Holmes, K. T., Mountford, C. E., and Wright, P. E. (1984) Biochemistry 23,5895-5897
42. Boulanger, P., and Mizon, J. (1976) Biochim. Biophys. Acta 451,
43. Vamecq, J., and Van Hoof, F. (1984) Bwchem. J. 221,203-211 44. Przyrembel, H., Wendel, U., Becker, K., Bremer, H. J., Bruinvis,
L., Ketting, D., and Wadman, S. K. (1976) Clin. Chim. Acta
45. Sweetman, L., Nyhan, W. L., Trauner, D. A., Allen Merritt, T.,
. .
161-169
66,227-239
and Singh, M. (1980) J. Pediatr. 9 6 , 1020-1026