metabolic pathways for differentiation-inducing factor-1 and their regulation are conserved between...
TRANSCRIPT

Differentiation (1994) 58:95-100
Differentiation Ontugcny, Neophsia and DiNerenliation Therapy
0 Springer-Verlag 1994
Metabolic pathways for differentiation-inducing factor-1 and their regulation are conserved between closely related Dictyostelium species, but not between distant members of the family Saskia van Ed, Steve Hodgkinson2, Pauline Schaap', Robert R. Kay2 'Cell Biology Unit, Institute of Molecular Plant Sciences, University of Leiden, Wassenaarseweg 64, NL-2333 AL Leiden, The Netherlands ?Medical Research Council Laboratory of Molecular Biology, Hills Road, Cambridge CB2 2QH, UK
Accepted in revised form: 2 September 1994
Abstract. There is suggestive evidence that a conserved signalling system involving differentiation inducing fac- tor-I (DIF-I) controls stalk cell differentiation in a vari- ety of slime mould species. In the standard laboratory species, Dictyostelium discoideum, DIF- 1 is first inacti- vated by dechlorination catalysed by DIF- I dechlori- nase, then by several hydroxylation events, so that even- tually about 12 metabolites are produced. If DIF-1 is used as a signal molecule in other species, they too must be able to metabolize it. We report here that the essen- tials of DIF-1 metabolism are conserved in D. mucoro- ides, the closest relative of D. discoideum. Both the de- chlorinase and hydroxylase enzymes were present in D. mucoroides, and living cells of both species produced a similar spectrum of metabolites from [-?HIDIF-1 . Fur- thermore, DIF-I dechlorinase was induced by DIF-l, as in D. discoideum, and this induction was repressed by ammonia and CAMP. DIF-1 dechlorinase could not be detected in cell extracts from D. minutum or Poly- sphondylium violaceum. However, living cells of both species are able to metabolize DIF-1; P. violaceum seems to produce a small amount of the monodechlori- nated compound, DIF-3, but all other metabolites from both species appear to be unique. Thus all investigated species can metabolize DIF- 1, but the exact route of me- tabolism is not highly conserved.
Introduction
The 50 or more species of cellular slime moulds produce a variety of stalked fruiting bodies during development. The most primitive species D. minutum, produces a tiny fruiting body consisting of a single tier of stalk cells, supporting a small spore head. D. discoideum produces a much larger fruiting body in which the stalk is stabilized by a basal disc, also consisting of stalk cells. D. mucoro- ides is closely related to D. discoideum with the major
Correspondence to: P. Schaap
difference that slugs of this species already form a stalk during migration [ 151. The more complex species of the genus Polysphondylium form secondary whorls of small fruiting bodies at regular intervals from the main stalk [ 151. Despite this variety of form, the basic mode of cell differentiation in all species appears similar: developing amoebae must differentiate either into vacuolated stalk cells or into spores with a condensed cytoplasm and a re- sistant spore coat. It is therefore possible that a relatively conserved set of signal molecules controls cell differen- tiation in all species and that structural variety arises from fine tuning of signalling in each individual species.
A number of small molecules are used as chemoat- tractants during cell aggregation: cAMP in D. discoi- deum and D. mucoroides [lo, 151, a dipeptide in P. vio- laceum [20], a pterin in D. minutum [4] and further un- identified compounds in other species. Despite this vari- ety of attractants used during aggregation, extracellular cAMP may be a universal signal coordinating postaggre- gative morphogenesis in a wide variety of species [5 , 16, 17, 19,211.
A second important molecule in the post-aggregative development of D. discoideum is a chlorinated alkyl phenone called DIF- I , which induces amoebae to differ- entiate into stalk cells [8,13J. DIF-I has been identified in D. mucoroides, the most closely related species to D. discoideum, by labelling with - ? T I . All other species in- vestigated also produce a developmentally regulated set of chlorine-containing compounds, which have not yet been positively identified as DIF-1 [9]. DIF-I induces expression of prestalk-specific mRNAs in D. discoi- deum, including the ecmB mRNA which encodes an ex- tracellular matrix protein [3, 261. A homologue of the ecmB gene has been cloned from D. minutum and can be strikingly induced by DIF-I [24].
Most details of the DIF-1 signalling system, such as the receptor and the biosynthetic route, remain unknown, but we have recently established how DIF-1 is inactived. This is done by a surprisingly complicated pathway in which the first step is a dechlorination of DIF-1 by DIF- I dechlorinase [14] followed by a hydroxylation of the

96
aliphatic side chain carried out by a P450 enzyme, DIF-3 hydroxylase (Morandini P, Offer J, Traynor D, Nayler 0, Neuhau D, Taylor GW, Kay RR unpublished results). This is followed by further oxidations of the side chain and other unknown conversions, to produce at least 12 metabolites [23]. A key feature of this metabolism is that DIF- 1 rapidly induces expression of DIF- 1 dechlorinase in responsive cells, thus producing a negative feedback loop which can regulate DIF-I levels in the aggregate
If DIF-I (or a related compound) is widely used in different slime mould species to induce stalk cell differ- entiation, we should expect that these species will also be able to metabolize, and presumably inactivate, DIF-1. To investigate this further, we have examined the metab- olism of DIF- 1 in a number of species.
~71.
Methods
DIF-I was obtained from Molecular Probes (Eugene, Oregon, USA). ['HIDIF- I (79 Ci/mmol) was synthesized by Amersham (Buckinghamshire, England, UK). DIF-3 was synthesized as de- scribed [ 121 and ['H]DIF-3 was prepared by enzymatic dechlori- nation of ['HIDIF-I [ 141 followed by high-performance liquid chromatography (HPLC) purification (Morandini et al., unpub- lished results). Thin-layer chromatography (TLC)-LK6D plates were from Whatman, and Enhance was from New England Nucle- ar (Du Pont de Nemours, s'Hertogenbosch, The Netherlands).
Growth and development of cells. D. mucoroides was grown on glucose-peptone agar. and P. violaceum and D. minutum on lac- tose-peptone agar in association with Escherichiu coli 281. Large quantities of cells were obtained by shaking about 5.10' cells/ml in 10 mM Na/K phosphate buffer, pH 6.5 (PB) at 150 rpm and 22" C with E. coli at 6.10Ycells/ml. To induce development, cells were washed free from bacteria and incubated at 22°C on PB- agar at 5x106 cells/cm2 for D. mucoroides and I? violaceum and at 106 cells/cm2 for D. minutum. D. minurum development was syn- chronized by placing 5 g activated charcoal in the lid of each petri dish.
Assay for DIF metabolism by intact cells. Aggregates or tipped ag- gregates were harvested in PB and dissociated by forcing them three times through a 23-gauge needle. Cells were resuspended at lo7 cells/ml and incubated with 50 nM ["IDIF- I . In the case of D. minurum, 3x107 cellslml and 10 nM [3H]DIF-I were used. Af- ter variable incubation periods, 1 -ml aliquots of cell suspension were centrifuged for 1 min at 15,OOO g, and pellet and supernatant fractions were stored at -20" C. Metabolites from the cell fraction were extracted with a mixture of chloroform, methanol and water [I] and from the medium by two extractions with one volume of ethyl acetate. The organic phases of both extracts were dried un- der vacuum and the residue was resuspended in 100 pI chloro- form/ethanol ( I : 1 ), containing 5 mg/ml butylated hydroxytoluene as antioxidant. Twenty-microlitre samples were loaded on a What- man LKdD TLC plate and developed in organic solvents as indi- cated in the figure legends. TLC plates were dried, sprayed with Enhance and used to expose X-ray film.
DIF-l dechlorinase assay. The DIF- I dechlorinase assay was per- formed as described by Nayler et al. [ 141. In short, lo7 cells were harvested by centrifugation and rapidly frozen to -70" C. The fro- zen cell pellets were lysed by thawing and resuspended in 200 p1 lysis buffer (50 mM Hepes pH 8.0, 50 mM KCI, I mM EDTA, I mM dithiothreitol (DTT), 10% glycerol, 5 pg/ml leupeptin, 2.5 Fglml pepstatin and 150 pg/ml benzamidine), and centrifuged at 170,OOOxg for 30 min. The amount of protein in the supernatant
was determined. Different amounts of protein (0.5, 5 and 50 pg) were used to ensure that 20%-60% of the label was converted and were incubated with 100 nM [JHIDIF-I and 5 mM glutathione for 30 min at 25" C. DIF-I and DIF-3 were extracted with 50 pl 90: I0:2 mixture of ethyl acetate/hexane/acetic acid, containing I mg/ml butylated hydroxytoluene. After extraction, 35 pI organic phase was applied to a TLC plate and developed in 60:40:2 hex- ane:ethyl acetate:acetic acid. Plates were dried, sprayed with En- hance and applied to X-ray film. DIF-I and DIF-3 bands were quantified by scraping the bands off the TLC, elution in 0.5 ml ethanol and scintillation counting.
DIF-3 hydroxylase assay. The DIF-3 hydroxylase assay in micro- soma1 fractions was performed according to Morandini et al. (un- published results). Cells were harvested and frozen as described for the DIF- 1 dechlorinase assay. Frozen cell pellets were lysed by thawing and resuspended in 100 pl lysis buffer (50 mM Na2HP0,, pH 7.8, \M)mM NaCI, 1O?b glycerol, 5 pg/ml leupeptin, 2.5 pg/ml pepstatin and 150 pg/ml benzamidine). Lysates were centri- fuged for I min at 15,000 g. The amount of protein in the superna- tant was determined and 100 pg protein was incubated in a final volume of 100 pl with 10 nM ['HIDIF-3 and 1 mM NADPH for 30 min at 23" C. DIF-3 and DIF-3 metabolites were extracted with 500 PI ethyl acetate: hexane (90: lo), containing 200 pg/ml butylated hydroxytoluene, and dried down under vacuum. The res- idue was taken up in 25 pl ch1oroform:methanol ( I : I ) containing 10 mM P-mercaptoethanol, loaded on a TLC plate and developed in hexane ethyl acetate acetic acid (20:80:2). TLC plates were fur- ther treated as described above. DIF-3 and the metabolites were quantified by measuring the radioactivity of bands scraped off the TLC plates.
Results
In vivo metabolism of DIF-I by D. mucoroides, D. minutum and f? violaceum.
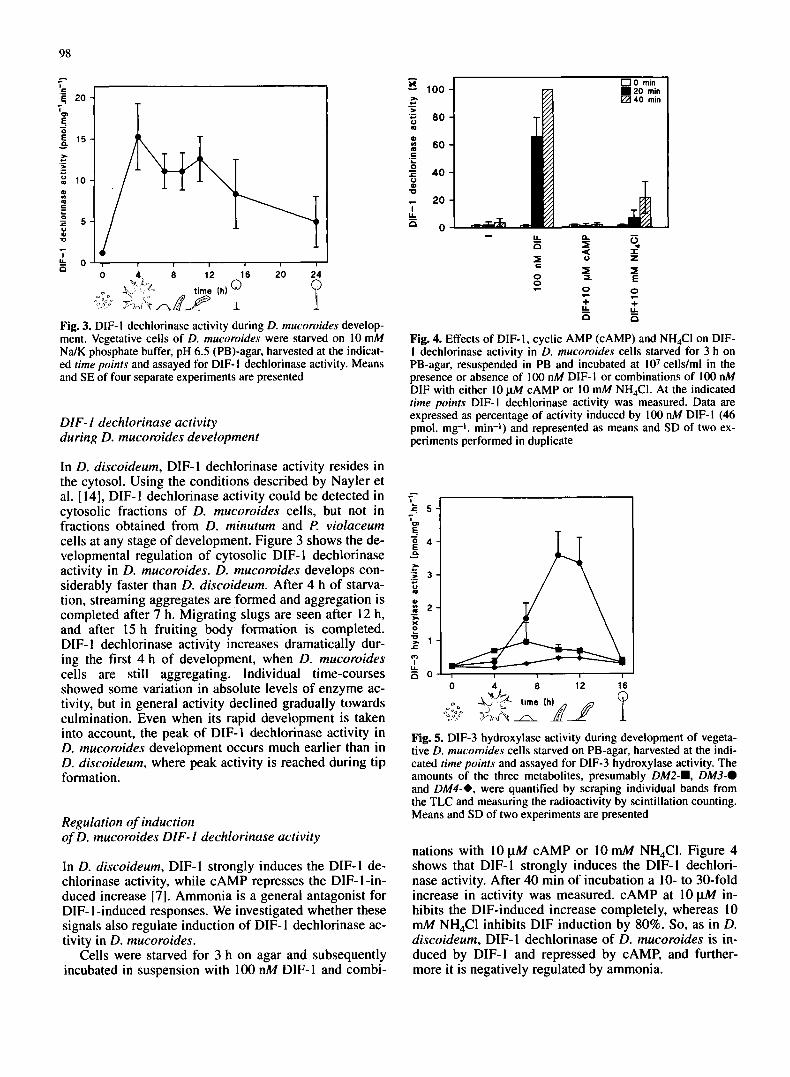
We first investigated whether the three species D. mu- coroides, D. minuturn and f? violaceum can metabolize DIF-1 in vivo. Aggregates or tipped aggregates of the three species were dissociated into single cells and incu- bated with ["IDIF-I. The metabolites that accumulated in cells and in the medium were separately extracted in chloroform/methanol/water [ 1 ] and ethyl acetate respec- tively, and analyzed by TLC. Figure 1A shows that D. mucoroides metabolizes DIF- 1 into several compounds. DIF-1 metabolism starts within 10 min of incubation and after 60 min almost all the DIF-1 is degraded. Two me- tabolites accumulate in the cells, one of which coelutes with DIF-3 in reverse-phase HPLC (data not shown), the other could not be identified. At least seven metabolites are secreted into the medium. These metabolites were compared with tritiated standards by TLC using two dif- ferent solvent systems (Fig. 2A, B). Three metabolites that were extracted from the medium at 120 min cochro- matographed with DM4, DM7 and DM8, which were previously identified in D. discoideum [23]. Using re- verse HPLC, DIF-3, DM4 and a small amount of DM3 (DM3 and DM4 are two of the three products of the DIF-3 hydroxylase in D. discoideum) were detected in these extracts by coelution with standards (data not shown). These data show that DIF-1 metabolism in D. mucoroides is rather similar to that in D. discoideum and suggest that D. mucoroides has both DIF-I dechlorinase and DIF-3 hydroxylase activity.
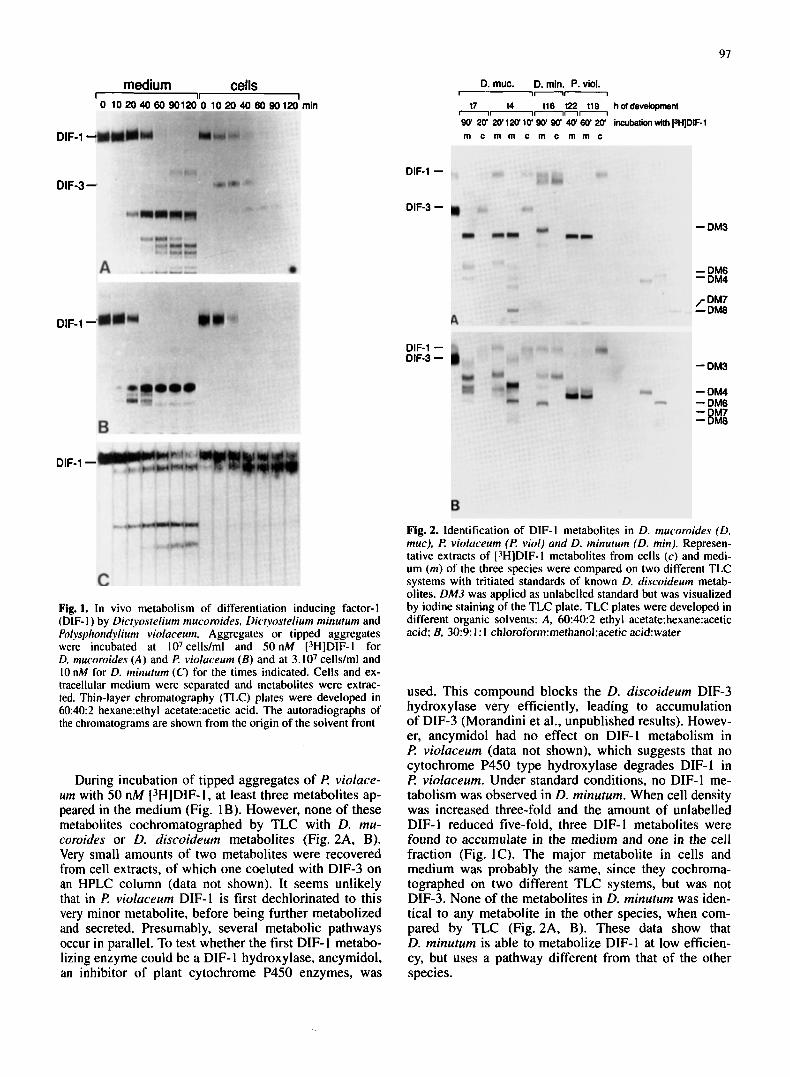
97
medium cells I 0 1020406090120
DIF-1-
Fig. 1. In vivo metabolism of differentiation inducing factor-I (DIF- I ) by Dictyostelium mucoroides, Dictyostelium minutum and Polysphondvlium viokuceum. Aggregates or tipped aggregates were incubated at lo7 cells/ml and 50 nM ['HIDIF-I for D. mucoroides ( A ) and €? violaceurn (B ) and at 3.107 cells/ml and 10 nM for D. minutum (C) for the times indicated. Cells and ex- tracellular medium were separated and metabolites were extrac- ted. Thin-layer chromatography (TLC) plates were developed in 60:40:2 hexane:ethyl acetate:acetic acid. The autoradiographs of the chromatograms are shown from the origin of the solvent front
During incubation of tipped aggregates of I? violuce- urn with 50 nM (3HlDIF-I, at least three metabolites ap- peared in the medium (Fig. IB). However, none of these metabolites cochromatographed by TLC with D. mu- coroides or D. discoideum metabolites (Fig. 2A, B). Very small amounts of two metabolites were recovered from cell extracts, of which one coeluted with DIF-3 on an HPLC column (data not shown). It seems unlikely that in F? violuceum DIF-1 is first dechlorinated to this very minor metabolite, before being further metabolized and secreted. Presumably, several metabolic pathways occur in parallel. To test whether the first DIF- 1 metabo- lizing enzyme could be a DIF- 1 hydroxylase, ancymidol, an inhibitor of plant cytochrome P450 enzymes, was
D. muc. D. rnin. P. viol. --- 17 14 116 122 t l B hotdevelopment
---n- 90' 20' 20'120 10' 90' 90' 40' 60' 20' incubation with PHJDIF-1 m c m m c m c m m c
DIF-1
DIF-3
- DM3
Fig. 2. Identification of DIF-I metabolites in D. mucoroides (D. muc), €! violaceurn (€? viol) and D. minutum (D. min). Represen- tative extracts of [3H]DIF-I metabolites from cells (c ) and medi- um (m) of the three species were compared on two different TLC systems with tritiated standards of known D. discoideum metab- olites. DM3 was applied as unlabelled standard but was visualized by iodine staining of the TLC plate. TLC plates were developed in different organic solvents: A, 60:40:2 ethyl acetate:hexane:acetic acid; B, 30:9: I : I ch1oroform:methanol:acetic acid:water
used. This compound blocks the D. discoideum DIF-3 hydroxylase very efficiently, leading to accumulation of DIF-3 (Morandini et al., unpublished results). Howev- er, ancymidol had no effect on DIF-1 metabolism in F? violuceum (data not shown), which suggests that no cytochrome P450 type hydroxylase degrades DIF-1 in I? violuceum. Under standard conditions, no DIF-I me- tabolism was observed in D. minutum. When cell density was increased three-fold and the amount of unlabelled DIF- 1 reduced five-fold, three DIF- 1 metabolites were found to accumulate in the medium and one in the cell fraction (Fig. IC). The major metabolite in cells and medium was probably the same, since they cochroma- tographed on two different TLC systems, but was not DIF-3. None of the metabolites in D. minutum was iden- tical to any metabolite in the other species, when com- pared by TLC (Fig. 2A, B). These data show that D. minutum is able to metabolize DIF-I at low efficien- cy, but uses a pathway different from that of the other species.
T
I I I I I
Fig. 3. DIF- I dechlorinase activity during D. mucoroides develop- ment. Vegetative cells of D. mucoroides were starved on 10 mM Na/K phosphate buffer, pH 6.5 (PB)-agar, harvested at the indicat- ed time points and assayed for DIF- I dechlorinase activity. Means and SE of four separate experiments are presented
DIF- I dechlorinase activity during D. mucoroides development
In D. discoideum, DIF- I dechlorinase activity resides in the cytosol. Using the conditions described by Nayler et al. [ 141, DIF-1 dechlorinase activity could be detected in cytosolic fractions of D. mucoroides cells, but not in fractions obtained from D. minutum and P. violaceum cells at any stage of development. Figure 3 shows the de- velopmental regulation of cytosolic DIF- I dechlorinase activity in D. mucoroides. D. mucoroides develops con- siderably faster than D. discoideum. After 4 h of starva- tion, streaming aggregates are formed and aggregation is completed after 7 h. Migrating slugs are seen after 12 h, and after 15 h fruiting body formation is completed. DIF- 1 dechlorinase activity increases dramatically dur- ing the first 4 h of development, when D. mucoroides cells are still aggregating. Individual time-courses showed some variation in absolute levels of enzyme ac- tivity, but in general activity declined gradually towards culmination. Even when its rapid development is taken into account, the peak of DIF-1 dechlorinase activity in D. mucoroides development occurs much earlier than in D. discoideum, where peak activity is reached during tip formation.
Regulation of induction of D. mucoroides DIF- I dechlorinase activity
In D. discoideum, DIF- 1 strongly induces the DIF- 1 de- chlorinase activity, while cAMP represses the DIF- I -in- duced increase [7]. Ammonia is a general antagonist for DIF- 1 -induced responses. We investigated whether these signals also regulate induction of DIF- 1 dechlorinase ac- tivity in D. mucoroides.
Cells were starved for 3 h on agar and subsequently incubated in suspension with 100 nM DIF-1 and combi-
100
80
60
40
20
0
+ + LL k n n
Fig. 4. Effects of DIF-I, cyclic AMP (CAMP) and NH4CI on DIF- I dechlorinase activity in D. mucoroides cells starved for 3 h on PB-agar, resuspended in PB and incubated at lo7 cellslml in the presence or absence of 100 nM DIF- I or combinations of I00 nM DIF with either 10 ph4 cAMP or 10 mM NH4CI. At the indicated time points DIF-I dechlorinase activity was measured. Data are expressed as percentage of activity induced by 100 nM DIF-I (46 pmol. mg-I. min-I) and represented as means and SD of two ex- periments performed in duplicate
I I I I
0 A 0 12 16
Fig. 5. D1F-3 hydroxylase activity during development of vegeta- tive D. mucoroides cells starved on PB-agar, harvested at the indi- cated time points and assayed for DIF-3 hydroxylase activity. The amounts of the three metabolites, presumably DM2-., DM3-0 and DM4-*, were quantified by scraping individual bands from the TLC and measuring the radioactivity by scintillation counting. Means and SD of two experiments are presented
nations with 10yM cAMP or 10mM NH,CI. Figure 4 shows that DIF-1 strongly induces the DIF-1 dechlori- nase activity. After 40 min of incubation a 10- to 30-fold increase in activity was measured. cAMP at 10 pA4 in- hibits the DIF-induced increase completely, whereas 10 mM NH,CI inhibits DIF induction by 80%. So, as in D. discoideum, DIF-1 dechlorinase of D. mucoroides is in- duced by DIF-I and repressed by CAMP, and further- more it is negatively regulated by ammonia.

99
DIF-3 hydroxylase activity during D. mucoroides development
In D. discoideum, DIF-3 hydroxylase activity was de- tected in microsomal fractions. The enzyme catalyzes hydroxylation of DIF-3 into either DM2, DM3 or DM4 (Morandini et al., unpublished results). We first tried to detect DIF-3 hydroxylase activity in different develop- mental stages of D. mucoroides, P. violaceum and D. mi- nutum. Only in microsomal fractions of D. mucoroides could activity be detected. Three metabolites were de- tected and, from their positions on TLC, it was deduced that they were most likely DM2, DM3 and DM4. This was confirmed by the identification of DM3 and DM4 by HPLC among the DIF- 1 metabolites produced in vivo (data not shown). Figure 5 shows a time-course of DIF-3 hydroxylase activity during D. mucoroides development. The most abundant product of DIF-3 hydroxylase activi- ty, DM3, is first detected when aggregation is complet- ed. It remains present during slug migration and decreas- es again during culmination. The other two products, DM2 and DM4, show maximal accumulation in mounds and slugs respectively. These data show that the second metabolic step in DIF-1 metabolism, performed by the DIF-3 hydroxylase, is also conserved in D. mucoroides.
Discussion
DIF-I is an important morphogenetic signal inducing the differentiation of stalk cells during D. discoideum devel- opment. A considerable body of evidence indicates that DIF-1 may be a general stalk-inducing signal in all cel- lular slime mold species. All investigated species pro- duce chlorinated compounds, one of which was identi- tied as DIF-1 in D. mucoroides [9]. Both P. violaceum and D. minutum secrete a compound that induces D. dis- coideum cells to differentiate as stalk cells in the DIF-1 bioassay ([2], R. Kay and S. Van Es, unpublished data). Furthermore, DIF-I induces expression of a D. minutum homologue of the ecmB prestalk gene at the same con- centrations that induce expression of this gene in D. dis- coideum [24]. Considering that all known chemical modifications strongly reduce the biological activity of DIF-1, these data suggest that both P. violaceum and D. minutum produce DIF-1, or at least a closely related compound. If DIF-I is to serve as a signal molecule, it is to be expected that all species can metabolize it in order to cause inactivation. Our results support this idea; all species tested had the capacity to metabolize DIF- 1, though the details and the rate of metabolism differed greatly. D. mucoroides and I? violaceum metabolized DIF-I at approximately the same rate as D. discoideum, but in D. minutum this rate was at least 10-20 times low- er. The metabolites produced by D. minutum and P. vio- laceum were all different from those recognized previ- ously in D. discoideum (though of course we cannot ex- clude that some minor metabolites are held in common). D. minutum metabolized DIF-1 to a number of unique metabolites that could not be further identified. All ma- jor t? violaceum metabolites differed from the ones that
are produced by D. discoideum, D. mucoroides and D. minutum. However, a minor product coeluting with DIF- 3 on HPLC was present at very low levels in cells. Ap- parently, the metabolic pathway in these species is dif- ferent from that in D. discoideum. In support of this, we were unable to detect either of the first two D. discoi- deum enzymes of DIF- 1 metabolism, DIF- 1 dechlorinase and DIF-3 hydroxylase, in D. minutum or P. violaceum.
In contrast, D. mucoroides displays a similar spec- trum of metabolites as D. discoideum and accumulates DIF-3 intracellularly, and DM3 and DM4 extracellularly. As would be expected, the first two enzymes of DIF me- tabolism, a cytosolic DIF-1 dechlorinase and a mem- brane-bound DIF-3 hydroxylase, were present in this species. Furthermore, DIF- 1 dechlorinase in D. mucoro- ides was regulated in a similar way to D. discoideum; it was induced strongly by DIF-1 and inhibited by CAMP. We also found that activity of DIF-1 dechlorinase was inhibited by ammonia in D. mucoroides. Although such an ammonia inhibition of DIF-1 dechlorinase has not been reported for D. discoideum, DIF- 1-induced stalk cell differentiation can be inhibited by ammonia in this species [6, 251. This conservation of elements of the DIF-I signalling system gives a biochemical basis for the close similarity of development in D. discoideum and D. mucoroides noted in descriptive studies.
A further possible role of metabolism is to create ac- tive compounds from DIF- 1, as happens with other lipo- philic signal molecules, such as thyroxin and retinol [ 1 1. 221. However, the observation that the three species do not produce common metabolites argues against a com- mon role for the metabolites in slime mold development. Most likely DIF-1 needs to be metabolized to prevent undue accumulation, or to create gradients of DIF- 1 that may regulate pattern formation. It is of interest that the presence of DIF-1 dechlorinase in D. discoideum and D. mucoroides is correlated with the display by these two species of an antero-posterior prestalk-prespore pattern not present in the other two species, P. violaceum and D. minutum [ 181.
The present study emphasizes the value of studies comparing multiple species for establishing the validity of putative mechanisms for pattern regulation. DIF itself is an important signal in slime mold development, but its metabolites may not function as signals themselves.
Acknowledgements. This research was supported by grant SLW- 805.17.046 from The Netherlands Organisation for Scientific Re- search and a European Union Human Capital and Mobility Grant.
References I . Bligh EG, Dyer WJ (1959) A rapid method of total lipid ex-
traction and purification. Canad J Biochem Physiol
2. Brookman JJ, Town CD, Jermyn KA, Kay RR (1982) Devel- opmental regulation of a stalk cell differentiation-inducing factor in Dicryosrelium discoideurn. Dev Biol 9 I : 19 1-1 96
3. Ceccarelli A, McRobbie SJ, Jermyn KA, Duffy K, Early A, Williams JG (1987) Structural and functional characteriza- tion of a Dicryostelium gene encoding a DIF-inducible, pre- stalk-enriched mRNA sequence. Nucleic Acids Res 157463- 7476
37:911-917

4. De Wit RJW, Konijn TM (1983) Identification of the acrasin of Dict.yostelium minutum as a derivative of folk acid. Cell Differ 12:205-2 10
5. Fukushima S, Maeda Y (1991) Whorl formation in Po1.v- sphondilium violmeurn: relevance to organization by CAMP. Dev Growth Differ 33525-533
6. Gross JD, Bradbury J, Kay RR, Pearcey MJ (1983) Intracellu- lar pH and the control of cell differentiation in Dictyostelium rliscoideum. Nature 303:244-245
7. lnsall R, Nayler 0, Kay RR (1992) DIF-I induces its own breakdown in Dictyostelium. EMBO .I I I :2849-2854
8. Kay RR, Jermyn KA (1983) A possible morphogen control- ling differentiation in Dicryostelium discoideum. Nature
9. Kay RR, Taylor GW, Jermyn KA, Traynor D (1992) Chlorine- containing compounds produced during Dictyosrelium devel- opment. Biochem J 281:155-161
10. Konijn TM, Van de Meene JGC, Bonner JT, Barkley DS (1967) The acrasin activity of adenosine-3’. S’cyclic phos- phate. Proc Natl Acad Sci USA 58: 1 152-1 154
I I . Larsen PR, Silva JE, Kaplan MM (1982) Relationships be- tween circulating and intracellular thyroid hormones: physio- logical and clinical implications. Endocr Rev 2:87-102
12. Masento MS, Morris HR, Taylor GW, Johnson SJ, Skapski AC, Kay RR ( 1988) Differentiation-inducing factor from the slime mould Dictyostelium discoideum and its analogues. Bio- chem J 256:23-28
13. Morris HR, Taylor GW, Masento MS, Jermyn KA, Kay RR ( 1987) Chemical structure of the morphogen differentiation inducing factor from Dicryostelium discoideum. Nature
14. Nayler 0, lnsall R, Kay RR (1992) Differentiation-inducing- factor dechlorinase, a novel cytosolic dechlorinating enzyme from Dictyosteliurn discoideum. Eur J Biochem 20853 1-536
15. Raper KB (1984) The Dictyostelids. Princeton University Press. Princeton, New Jersey, USA
303~242-244
328~11-814
16. Robertson A, Cohen MH ( 1972) Control of developing fields. Annu Rev Biophys Bioeng 1 :409-464
17. Schaap P, Konijn TM, Van Haastert PJM (1984) cAMP pulses coordinate morphogenetic movement during fruiting body for- mation of D. minutum. Proc Natl Acad Sci USA
18. Schaap P, Pinas JE, Wang M ( 1985) Patterns of cell differenti- ation in several cellular slime mold species. Dev Biol I 1 1:51-61
19. Schaap P, Van Lookeren Campagne M, Van Driel R, Spek W, Van Haastert, PJM, Pinas J ( 1986) Postaggregative differentia- tion induction by cAMP in Dictyostelium: intracellular trans- duction pathway and requirement for additional stimuli. Dev Biol 11852-63
20. Shimomura 0, Suthers HLB, Bonner JT (1982) Chemical identity of the acrasin of the cellular slime mold Poly- sphondylium violaceum. Proc Natl Acad Sci USA 79:7376-7379
2 1. Siegert F, Weijer CJ ( 1 992) Three-dimensional scroll waves organize Dictyostelium slugs. Proc Natl Acad Sci USA 89:6433-6437
22. Thaller C, Eichele G (1990) Isolation of 3.4-didehydroretinoic acid, a novel morphogenetic signal in the chick wing bud. Na- ture 345,8 15-8 19
23. Traynor D, Kay RR (1991) The DIF-I signaling system in Dictyostelium. J Biol Chem 266529 1-5297
24. Van Es S, Nieuwenhuijsen BW, Lenouvel F, Van Deursen EM, Schaap P (1994) Universal signals control slime mold stalk formation. Proc Natl Acad Sci (in press)
25. Wang M, Schaap P (1989) Ammonia depletion and DIF trig- ger stalk cell differentiation in intact Dictyostelium discoi- deum slugs. Development 105569-574
26. Williams JG, Ceccarelli A, McRobbie S, Mahbubani H, Kay RR, Early A, Berks M, Jermyn KA (1987) Direct induction of Dictvosrelium prestalk gene expression by DIF provides evi- dence that DIF is a morphogen. Cell 49: 185-192
8 1 :2 122-2 I26