mertk mediates intrinsic and adaptive resistance to axl ... · resistance to axl-targeting agents...
TRANSCRIPT

Small Molecule Therapeutics
MERTK Mediates Intrinsic and AdaptiveResistance to AXL-targeting AgentsNellie K. McDaniel1, Christopher T. Cummings2, Mari Iida1, Justus H€ulse3,Hannah E. Pearson1, Eleana Vasileiadi3, Rebecca E. Parker3, Rachel A. Orbuch1,Olivia J. Ondracek1, Noah B.Welke1, Grace H. Kang1, Kurtis D. Davies2,XiaodongWang4, StephenV. Frye4,5, H. Shelton Earp5,6, PaulM. Harari1, Randall J. Kimple1,Deborah DeRyckere3, Douglas K. Graham3, and Deric L.Wheeler1
Abstract
The TAM (TYRO3, AXL, MERTK) family receptor tyrosinekinases (RTK) play an important role in promoting growth,survival, andmetastatic spread of several tumor types. AXL andMERTK are overexpressed in head and neck squamous cellcarcinoma (HNSCC), triple-negative breast cancer (TNBC),and non–small cell lung cancer (NSCLC), malignancies thatare highly metastatic and lethal. AXL is the most well-charac-terized TAM receptor and mediates resistance to both conven-tional and targeted cancer therapies. AXL is highly expressed inaggressive tumor types, and patients with cancer are currentlybeing enrolled in clinical trials testing AXL inhibitors. In thisstudy, we analyzed the effects of AXL inhibition using a small-molecule AXL inhibitor, a monoclonal antibody (mAb), andsiRNA inHNSCC, TNBC, andNSCLCpreclinicalmodels. Anti-AXL–targeting strategies had limited efficacy across these dif-
ferent models that, our data suggest, could be attributed toupregulation of MERTK. MERTK expression was increased incell lines and patient-derived xenografts treated with AXLinhibitors and inhibition ofMERTK sensitizedHNSCC, TNBC,and NSCLC preclinical models to AXL inhibition. Dual target-ing of AXL and MERTK led to a more potent blockade ofdownstream signaling, synergistic inhibition of tumor cellexpansion in culture, and reduced tumor growth in vivo.Furthermore, ectopic overexpression of MERTK in AXL inhib-itor–sensitive models resulted in resistance to AXL-targetingstrategies. These observations suggest that therapeutic strate-gies cotargeting both AXL and MERTK could be highly ben-eficial in a variety of tumor types where both receptors areexpressed, leading to improved survival for patientswith lethalmalignancies. Mol Cancer Ther; 17(11); 2297–308. �2018 AACR.
IntroductionThe TAM family of receptor tyrosine kinases is comprised of
three family members: TYRO3 (SKY), AXL (ARK or UFO), andMERTK. Cognate ligand binding to TAM receptors on the cell
surface leads to receptor dimerization, kinase domain activation,and auto/trans-phosphorylation of tyrosine residues located oneach receptor's cytoplasmic tail (1). Activation of TAM receptorsstimulates PI3K/AKT, RAS/RAF/MEK/ERK (MAPK), and othersignaling cascades leading to increased tumor cell survival, pro-liferation, migration, invasion, and angiogenesis (1–4).
AXL andMERTK have been implicated in the development andprogression ofmanymalignancies including lung (3, 5–9), breast(3, 10–14), ovarian (15), colon (16), head and neck (17), thyroid(18), prostate (19), pancreatic (20), osteosarcoma (21), mela-noma (22), leukemia (23), and Kaposi sarcoma (24). With theadvancement of preclinical studies supporting the role of TAMfamily receptors in cancer, TAM family inhibitors are beingclinically developed and will likely be beneficial in specific cancersettings. Themost clinically advanced agent with AXL as a primarytarget is R428 (BGB324; ref. 10), which is currently being tested inphase Ib/II clinical trials for treatment of acute myeloid leukemia(AML), NSCLC, and melanoma (25). R428 has potent activityagainst AXL (IC50 ¼ 14 nmol/L) and 50-fold selectivity for AXLrelative to MERTK in cell-based assays (10). Similarly, UNC2025is a MERTK and FLT3 tyrosine kinase inhibitor (TKI) that haspotent activity against MERTK (IC50 ¼ 2.7 nmol/L), 40-foldselectivity for MERTK relative to AXL in cell-based assays (26),and mediates therapeutic activity in preclinical leukemia andNSCLC xenograft models (26–28). Numerous other small-mol-ecule TKIs that were developed to target other kinases have lownanomolar potency against AXL and/or MERTK, including agentsthat are FDA-approved (cabozantinb, crizotinib, and sunitinib) or
1University of Wisconsin School of Medicine and Public Health, Madison,Wisconsin. 2Department of Pediatrics, Section of Hematology, Oncology, andBone Marrow Transplantation, University of Colorado Anschutz Medical Cam-pus, Aurora, Colorado. 3Aflac Cancer and Blood Disorders Center of Children'sHealthcare of Atlanta and Department of Pediatrics, School of Medicine, EmoryUniversity, Atlanta, Georgia. 4Center for Integrative Chemical Biology and DrugDiscovery and Division of Chemical Biology and Medicinal Chemistry, EshelmanSchool of Pharmacy, University of North Carolina at Chapel Hill, Chapel Hill,North Carolina. 5Department of Medicine, UNC Lineberger ComprehensiveCancer Center, Chapel Hill, North Carolina. 6Department of Pharmacology,School of Medicine, University of North Carolina at Chapel Hill, Chapel Hill,North Carolina.
Note: Supplementary data for this article are available at Molecular CancerTherapeutics Online (http://mct.aacrjournals.org/).
N.K. McDaniel and C.T. Cummings contributed equally to this article.
Corresponding Authors: Deric L. Wheeler, University of Wisconsin School ofMedicine and Public Health, 1111 Highland Avenue, WIMR 3159, Madison, WI53705. Phone: 608-262-7837; Fax: 608-263-9947; E-mail: [email protected];and Douglas K. Graham, Emory University, 201 Downman Drive, Atlanta, GA30322. E-mail: [email protected]
doi: 10.1158/1535-7163.MCT-17-1239
�2018 American Association for Cancer Research.
MolecularCancerTherapeutics
www.aacrjournals.org 2297
on May 28, 2020. © 2018 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 9, 2018; DOI: 10.1158/1535-7163.MCT-17-1239

are being tested in clinical studies (merestinib, BMS-777607,and S49076).
Resistance to single-agent therapy is a major hurdle in cancertreatment. Cooperation of family members to overcome resis-tance is well documented (29–31). Here, we describe an intrinsicand adaptive feedback mechanism of resistance to AXL-targetingagents mediated by upregulation of MERTK. Examination of alarge panel of cell lines from differentmalignancies indicated thathigh expression of MERTK correlates with relative resistance toanti-AXL therapeutics. Targeting AXL with genetic silencing, asmall-molecule inhibitor, or an AXL-specific antibody increasedMERTK protein expression in vitro and in vivo. Furthermore,combined targeting of MERTK and AXL inhibited tumor cellexpansion in vitro and impacted tumor growth in vivo. Dualtargeting of MERTK and AXL impacted several signaling pathwaysincluding AKT, STAT6, P70S6 kinase (P70S6K), S6 ribosomalprotein (S6RP), and C-RAF. Finally, overexpression of MERTK incells that were sensitive to AXL targeting led to activation ofP70S6K, S6RP, andC-RAF andwas sufficient tomediate resistanceto AXL-targeting agents. Collectively, the data presented hereindicate a critical role for MERTK in resistance to AXL-targetedtherapy and provide rationale for clinical evaluation of strategiestargeting both AXL and MERTK for treatment of cancer.
Materials and MethodsCell lines
All cell lines were obtained from indicated sources (Supple-mentary Materials and Methods). Cell line identities were con-firmed using short tandem repeat (STR) analysis and publiclyavailable databases. Validated STR profiles were not available forthe H157 cell line, but the profile obtained did not match anyother cell line.
TAM RTK inhibitorsUNC2025 (MERTK inhibitor) was synthesized as described
previously (26) or purchased from Selleck Chemicals or Apexbio.R428 (AXL inhibitor) was purchased from Selleck Chemicals orApexbio. The mAb targeting AXL (Mab173) was kindly providedby Dr. Parkash Gill, Department of Medicine and Pathology,University of Southern California (Los Angeles, CA; ref. 24).Subconfluent cultures were treated with the indicated concentra-tions of Mab173, UNC2025, R428, or equivalent volumes ofDMSO vehicle.
siRNA transfectionNontargeting control siRNA (catalog no. D-001810-01-05)
and siRNAs targeting AXL (catalog nos. J-003104-10-0005 andJ-003104-12-0005)were purchased fromThermoFisher Scientificand utilized for transfection of NSCLC cells with final concentra-tions of 25 nmol/L siRNA and 1 mL/mL DharmaFECT1 Transfec-tion Reagent (catalog no. T-2001-02). Nontargeting control poolsiRNA (catalog no. D-001810) and SMARTpool siRNA targetingAXL (catalog no. L-003104)werepurchased fromDharmacon, Incand utilized for transfection ofHNSCC and TNBC cell lines with afinal concentration of 30 nmol/L siRNA with LipofectamineRNAiMAX (Life Technologies).
Plasmids and transfectionpDONR223-MERTK was a gift from William Hahn & David
Root (Addgene, plasmid#23900) and subcloned into theBamHI/
NOTI restriction sites of the pcDNA6.0 expression vector (LifeTechnologies). pcDNA3.1-TYRO3 was purchased fromGenScriptUSA Inc. (catalog no. OHu23055D). Transfection was performedusing Lipofectamine LTX and Opti-MEM I (Life Technologies)according to the manufacturer's instructions.
ImmunoblottingCells were washed with PBS and scraped into lysis buffer
(50 mmol/L HEPES pH 7.5, 150 mmol/L NaCl, 10 mmol/LEDTA, 10% glycerol, 1% Triton X-100, 1 mmol/L Na3VO4)supplemented with protease inhibitors (Complete Mini, RocheMolecular Biochemicals). Protein concentrations were deter-mined and immunoblotting was performed as described previ-ously (6) using antibodies shown in Supplementary Table S1. Allantibodies were used as recommended by the manufacturer'sinstructions.
TAM kinase phosphorylationCells were cultured in medium with DMSO vehicle control or
the indicated concentration of drug for 1 hour or 24 hours andthen 120 mmol/L freshly prepared pervanadate phosphataseinhibitor was added for an additional 3 minutes to stabilizephosphorylated proteins (26). Lysates were prepared, MERTKand AXL were immunoprecipitated with anti-MERTK (R&D Sys-tems, MAB8912) or anti-AXL (R&D Systems, AF154) antibodiesand Protein G beads (Invitrogen, catalog no. 10-1242) andphosphorylated and total MERTK and AXL proteins were detectedby immunoblot as described above.
Cell expansion assaysCellTiter 96 AQueous One Solution Reagent (Promega, catalog
no. G3580), Cell Counting Kit 8 (Dojindo Molecular Technolo-gies, catalog no. CK04), and crystal violet assays were used todetermine relative numbers of viable cells 72 hours after trans-fection with siRNA-targeting AXL (siAXL) or treatment with AXLand/or MERTK inhibitors. Alternatively, relative confluency wasdetermined using the Incucyte ZOOM Live Cell Imaging System(Essen Biosciences). For studies using combined treatment withsiAXLandUNC2025, cellswere transfectedwith siAXLand then24hours laterwere treatedwithUNC2025 for anadditional72hours.
Clonogenic assayH1299 NSCLC cells were cultured at low density in liquid
medium containing R428, UNC2025, R428 and UNC2025, orvehicle. Colonies were stained with crystal violet after 10 daysand counted using a GelCount automated counter (OxfordOptronix).
Xenograft modelsFemale athymic nudemice (4–6week old) were obtained from
Envigo. Animal procedures and maintenance were conducted inaccordance with institutional guidelines of University of Wiscon-sin (Madison, WI). UW-SCC1 PDX tumors, UM-SCC1, andMDAMB231 cell lines were inoculated by subcutaneous injectioninto the dorsal flank of each mouse and tumor volume wasmeasured using calipers. When tumors attained a volume of200 mm3 (for 28-day study) or 500 mm3 (for 4-day study), micewere randomized into groups and treatment was initiated. R428,UNC2025, R428þUNC2025, or an equivalent volume of vehicle(0.5% hydroxypropylmethylcellulose þ 0.1% Tween 80) wereadministered twice daily by oral gavage on 5 days per week.
McDaniel et al.
Mol Cancer Ther; 17(11) November 2018 Molecular Cancer Therapeutics2298
on May 28, 2020. © 2018 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 9, 2018; DOI: 10.1158/1535-7163.MCT-17-1239

Tumors were collected within 3 hours of the last treatment foranalysis of biochemical markers (32).
ImmunohistochemistryTumors were processed for IHC as described previously (33).
Sections were heated in 10 mmol/L citrate buffer (pH 6.0)or 10 mmol/L Tris þ 1 mmol/L EDTA buffer (pH 9.0) witha decloaking chamber and incubated overnight at 4�Cwith pS6RP Y235/236 (1:200), KI67 (1:800), or MERTK(1:50) antibodies. Bound antibodies were detected using theVECTASTAIN Universal Kit/HRP (Vector Laboratories) and3,30-diaminobenzidine substrates. Tissues were counterstainedwith Mayer hematoxylin (Thermo Fisher Scientific) and wereexamined using an Olympus BX51 microscope. Images areshown at a magnification of 20�.
Evaluation of therapeutic interactionsThe fractional productmethodwas used to evaluate therapeutic
interactions between AXL inhibitors and UNC2025 in cell cultureassays (34, 35). Thismethodutilizes relative cell density followingtreatment with each individual agent to calculate the expected (E)effect for an additive interaction as a product of the individualresponses. The observed (O) effect is the relative cell densityfollowing dual treatment. The ratio of observed to expected(O:E) values was calculated and used to indicate synergy (O:E< 1), additivity (O:E¼ 1), or antagonism (O:E > 1). Isobologramanalysis was also performed as described previously (36). Experi-ments were repeated three times in triplicate.
Statistical analysisStatistical analyses were performed using Prism 7 software
(GraphPad Software, Inc.). Differences between multiplegroups were evaluated using repeated measures ANOVA withBoneferroni post test. Student t test was used for comparisonsbetween two groups. Differences were considered significantwhen P < 0.05.
ResultsMERTK expression correlates with resistance to AXL-targetingagents
AXL has been identified as a novel molecular target in numer-ous human cancers including HNSCC, TNBC, and NSCLC (6, 11,37–40), and anti-AXL targeting strategies are now in clinicaldevelopment for these malignancies. Mechanisms of resistanceto AXL inhibitors have not been evaluated but will likely play arole in limiting the clinical efficacy of these inhibitors. To examinepotential resistance mechanisms, we assessed expression of TAMfamily receptors and sensitivity to the anti-AXL small-moleculeinhibitor R428 across a large panel of HNSCC (Fig. 1A and B),TNBC (Fig. 1C and D), and NSCLC (Fig. 1E and F) cell lines. AXL,MERTK, and TYRO3 were differentially expressed in cancer celllines and immunoblot analysis confirmed inhibition of AXLphosphorylation in response to treatment with R428 (Supple-mentary Fig. S1). R428did not inhibitMERTKphosphorylation atthe doses used (Supplementary Fig. S1C). Although all of thetumor cell lines expressed AXL to some degree, analysis of tumorcell expansion in culture revealed that only a subset of cell linesexhibited a statistically significant decrease in cell number inresponse to treatment with R428 and were scored as sensitive,whereas the majority were relatively resistant (Fig. 1B, D and F).
Interestingly, all five of the cell lines that expressed little or noMERTK were sensitive to AXL inhibition (HN30, UM-SCC38,SUM159, SUM149, H226), whereas 8 of 10 cell lines thatexpressed higher levels of MERTK had no significant response tosingle-agent R428 treatment (UM-SCC1, UM-SCC6, UM-SCC47,UM-SCC104, SUM229, BT549,MDAMB231, H157), suggesting arole for MERTK in resistance to AXL-targeting agents.
Inhibition of AXL increases MERTK protein expressionin vitro
Several studies have reported the cooperative nature of RTKs incancer and resistance to RTK-targeting strategies. For example, theHER2 and HER3 receptors can be upregulated and mediateresistance to anti-EGFR therapy (30, 41, 42). To determinewhether alternative TAM family receptors are similarly upregu-lated in response to anti-AXL therapeutics, we analyzed theexpression ofMERTK and TYRO3 inHNSCC (UM-SCC47, HN30,and UM-SCC4), TNBC (BT549, MDAMB231), NSCLC (H1299,H157), and esophageal cancer (KY270, KY410, and SKGT4) celllines after AXL inhibition mediated by genetic silencing (siAXL),small-molecule inhibition (R428), or anti-AXL mAb therapy(Mab173). First, cell lines were transfected with siAXL or a non-targeting siRNA control to examine the effects of genetic knock-down of AXL on MERTK and TYRO3 expression levels. WhileTYRO3 expression levels were unchanged in cells depleted of AXL,MERTK expression levels were increased by approximately 1.5- to15-fold (Fig. 2A, D, and H; Supplementary Fig. S2A). In contrast,expression of EGFR andC-MET, RTKs outside the TAM family, didnot change after AXL knockdown (Fig. 2A and D). Similarly,MERTK expression was increased by approximately 1.5 to 15-foldin HNSCC, TNBC, NSCLC, and esophageal cancer cells treatedwith the AXL small-molecule inhibitor R428 (Fig. 2B, E, and I;Supplementary Fig. S2B) or the anti-AXL mAb173 (Fig. 2F)compared with vehicle-treated controls. MERTK phosphorylationwas also increased in select HNSCC, TNBC, NSCLC, and esoph-ageal cell lines following treatment with R428 indicatingincreased MERTK activity along with MERTK upregulation (Fig.2C, G and J; Supplementary Fig. S2C). EGFR and cMET proteinlevels were unchanged in response to AXL inhibition in HNSCC(Fig. 2A and B) and TNBC (Fig. 2D–F) cell lines. TYRO3 proteinlevels were only slightly increased in a subset of cell lines treatedwith R428 (Fig. 2) and high levels of TYRO3did not correlate withresistance to AXL inhibition (Figs. 1 and 2). Taken together, thesedata demonstrate selective upregulation of MERTK in tumor celllines in response to AXL inhibition.
Combined inhibition of AXL and MERTK synergisticallydecreases tumor cell expansion and inhibits downstreamsignaling
Expression of MERTK correlated with resistance to AXL inhi-bitors (Fig. 1) and was upregulated in response to AXL suppres-sion (Fig. 2); thus, we hypothesized that targeting MERTK wouldenhance the efficacy of AXL inhibitors. To test this hypothesis,HNSCC (UM-SCC1, UM-SCC4, UM-SCC6, UM-SCC47, andUM-SCC104), TNBC (SUM229, BT549, and MDAMB231), andNSCLC (H1299 and H157) cell lines that coexpressed both AXLand MERTK were either treated with a combination of R428 andUNC2025 (26) for 72 hours or transfected with siAXL and thentreatedwithUNC2025 prior to determining relative cell numbers.Immunoblot analysis confirmed inhibition of MERTK phosphor-ylation in response to treatment with UNC2025 (Supplementary
MERTK Mediates Resistance to AXL-targeted Therapy
www.aacrjournals.org Mol Cancer Ther; 17(11) November 2018 2299
on May 28, 2020. © 2018 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 9, 2018; DOI: 10.1158/1535-7163.MCT-17-1239

Fig. S3). AXL phosphorylation was not affected by treatment withUNC2025 at the doses used for these studies (SupplementaryFig. S3C). Combined treatment with AXL and MERTK inhibitorssignificantly reduced tumor cell expansion relative to either agentalone in nearly all cell lines examined (Fig. 3A–F). In addition, the
interactions betweenR428 andUNC2025or siAXL andUNC2025were assessed in HNSCC, TNBC, and NSCLC cells using thefractional product method. The results demonstrated statisticallysignificant differences between the expected relative cell number(E) assuming an additive interaction and the actual value
Figure 1.
Endogenous expression of AXL and MERTK predicts sensitivity to AXL inhibition. A, C, and E, Cell lysates were prepared from HNSCC, TNBC, and NSCLCcell lines, and endogenous TAM kinase expression was determined by immunoblot. a-Tubulin is shown as loading control. B, D, and F, HNSCC, TNBC,and NSCLC cell lines were treated with vehicle or R428 for 72 hours, and relative cell numbers were determined. Mean values, SEs, and statistical analyseswere derived from 3 (HNSCC and TNBC) or 7 (NSCLC) independent experiments. � , P < 0.05; �� , P < 0.01.
McDaniel et al.
Mol Cancer Ther; 17(11) November 2018 Molecular Cancer Therapeutics2300
on May 28, 2020. © 2018 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 9, 2018; DOI: 10.1158/1535-7163.MCT-17-1239

Figure 2.
Inhibition of AXL increased MERTK protein expression and activity in vitro. A, D, H, Genetic inhibition of AXL in HNSCC, TNBC, and NSCLC cell lines. Cellswere transfected with AXL siRNA (siAXL) or nontargeting siRNA (siNT). Cell lysates were prepared 72 hours later. B, E, and I, Small-molecule inhibition ofAXL in HNSCC, TNBC, and NSCLC cell lines. Cells were treated with vehicle or R428 for 72 hours prior to preparation of cell lysates. F, Antibody inhibition ofAXL in TNBC cell lines. Cells were treated with vehicle or Mab173 for 120 hours prior to preparation of cell lysates. The indicated proteins were detected byimmunoblot.a-Tubulin is shown as a loading control. MERTK expressionwas quantitated by densitometry using ImageJ software. Mean values and SEs derived fromthree independent experiments are shown. �, P < 0.05; �� , P < 0.01; ns ¼ not significant. C, G, and J, The indicated cell lines were treated with vehicle or R428 for24 hours, and phosphorylated proteins were stabilized by treatment with pervanadate phosphatase inhibitor prior to preparation of cell lysates. MERTK wasimmunoprecipitated and phosphorylated (denoted by p-), and total MERTK proteins were detected by immunoblot. The data shown are representative oftwo or three independent experiments.
MERTK Mediates Resistance to AXL-targeted Therapy
www.aacrjournals.org Mol Cancer Ther; 17(11) November 2018 2301
on May 28, 2020. © 2018 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 9, 2018; DOI: 10.1158/1535-7163.MCT-17-1239

observed (O) indicating synergy in 6 of 10 cell lines tested(Supplementary Fig. S4A). Synergistic interactions were alsodemonstrated in UM-SCC1 and MDAMB231 cell lines usingisobologram analyses (Supplementary Fig. S4B). Similarly,reduced expansion was observed in esophageal cell lines expres-
sing both AXL and MERTK after treatment with R428 andUNC2025 combination therapy (Supplementary Fig. S4C–S4E). Collectively, these data demonstrate combined inhibitionof AXL and MERTK as a highly effective strategy to suppressexpansion of tumor cells that coexpress both receptors.
Figure 3.
Dual targeting of AXL and MERTK can effectively inhibit cell proliferation in vitro. A, C, HNSCC and TNBC cell lines were treated with vehicle (DMSO),R428, UNC2025, or R428þUNC2025, and relative cell numbers were determined after 72 hours. B, D, F, UM-SCC1, MDAMB231, and NSCLC cell lines weretransfected with AXL siRNA (siAXL, siAXL10, siAXL12) or nontargeting siRNA (siNT) and then treated with vehicle or UNC2025 for 72 hours prior todetermination of relative cell numbers. E, NSCLC cell lines were cultured at low density and treated with vehicle (DMSO), R428, UNC2025, or R428þUNC2025for 10 days. Colonies were stained and counted. � , P < 0.05; �� , P < 0.01; ��� , P < 0.001, NS ¼ not significant.
McDaniel et al.
Mol Cancer Ther; 17(11) November 2018 Molecular Cancer Therapeutics2302
on May 28, 2020. © 2018 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 9, 2018; DOI: 10.1158/1535-7163.MCT-17-1239

To investigate potential mechanisms of synergistic anti-tumoractivity mediated by the combination of R428 and UNC2025, wedetermined the status of downstream signaling pathways in thecontext of R428 and siAXL in combinationwithUNC2025. InUM-SCC1 and MDAMB231 cells treated with R428 and UNC2025,there was a substantial decrease in expression of phospho-STAT6,phospho-AKT, phospho-P70S6K, phospho-S6RP, and phospho-C-RAF (Fig. 4A and B). In both cases, signaling was decreased to agreater degree in cultures treated with R428 and UNC2025 ascompared with cultures treated with either single agent. Similarly,in H1299 NSCLC cells treated with siAXL and UNC2025, com-bined inhibition of AXL and MERTK led to more potent decreasesin phosphorylation of STAT6 andAKT (Fig. 4C). Collectively, thesedata demonstrate reduced downstream signaling in response tocombined inhibition of AXL and MERTK and suggest potentialbiochemical mechanisms for the corresponding reduction intumor cell expansion mediated by the combination treatment.
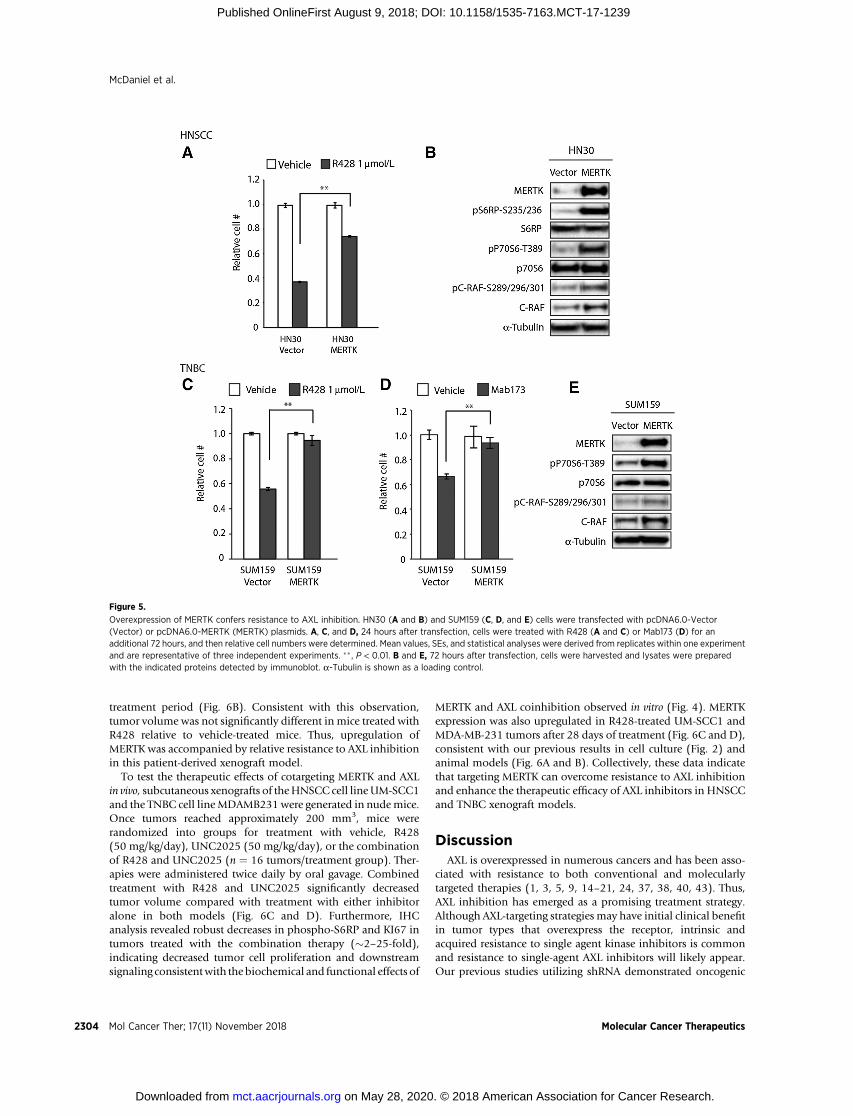
MERTK overexpression mediates resistance to AXL inhibitionTo determine whether expression of MERTK is sufficient to
mediate resistance to anti-AXL therapy, HNSCC and TNBC celllines that are sensitive to anti-AXL–targeting strategies (HN30 andSUM159) and do not express endogenous MERTK (Fig. 1A–D)were manipulated to overexpress MERTK and then treated withAXL inhibitors R428 or Mab173 for 72 hours prior to assessmentof relative cell numbers. Although tumor cell expansion wasinhibitedby45%to65% invector-expressingHN30andSUM159cells treated with R428 and Mab173, derivative cells with ectopicexpression of MERTK were relatively resistant and cell numbers
were only decreased by 3% to 23%(Fig. 5A, C, andD). In contrast,HN30 cells with ectopic expression of TYRO3 remained sensitiveto R428 treatment (Supplementary Fig. S5). Consistent with thebiochemical effects of MERTK inhibition, ectopic expression ofMERTK increased downstream signaling through phospho-P70S6K, phospho-S6rp, and phospho-C-RAF (Fig. 5B and E).Collectively, these results demonstrate a direct role for MERTK inresistance to AXL inhibitors.
MERTK-mediated resistance to AXL inhibition can be overcomeby dual targeting in vivo
The observed upregulation of MERTK in tumor cell lines inresponse to AXL inhibition (Fig. 2) and the potent antitumorresponses mediated by combined targeting of MERTK and AXLin vitro (Fig. 3) implicate coinhibition as an effective therapeuticstrategy. To further investigate this potential, we determinedwhether a similar response could be observed in xenograft mod-els. First, MDAMB231 (TNBC) xenografts were established innude mice. Once tumors reached a volume of approximately500mm3,mice were treated with vehicle or R428 (50mg/kg/day)for 4 days (n¼ 8 tumors/treatment group) and then tumors wereharvested and analyzed for MERTK expression via immunoblot-ting and IHC. In this study, MERTK expression was consistentlyincreased in MDAMB231 tumors after AXL inhibition (Fig. 6A).Similarly, a HNSCC patient-derived xenograft (PDX) wasestablished in nude mice and treated with vehicle or R428(50 mg/kg/day) for 4 days or 28 days (n ¼ 8 tumors/treatmentgroup). Again, MERTK was prominently upregulated in tumorstreated with R428 and this increase persisted during the extended
Figure 4.
Dual inhibition of AXL and MERTK reducesdownstream signaling through oncogenic pathways.A and B, UM-SCC1 and MDAMB231 cells were treatedwith vehicle (DMSO), R428, UNC2025, orR428þUNC2025 for 24 hours. C, H1299 cells weretransfected with AXL siRNA (siAXL10 or siAXL12) ornontargeting siRNA (siNT), and cells were treatedwith vehicle or UNC2025 for 1 hour. Expression of theindicated proteins was assessed in cell lysates byimmunoblot.a-Tubulin is shown as a loading control.The data shown are representative of 3independent experiments.
MERTK Mediates Resistance to AXL-targeted Therapy
www.aacrjournals.org Mol Cancer Ther; 17(11) November 2018 2303
on May 28, 2020. © 2018 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 9, 2018; DOI: 10.1158/1535-7163.MCT-17-1239
treatment period (Fig. 6B). Consistent with this observation,tumor volume was not significantly different in mice treated withR428 relative to vehicle-treated mice. Thus, upregulation ofMERTK was accompanied by relative resistance to AXL inhibitionin this patient-derived xenograft model.
To test the therapeutic effects of cotargeting MERTK and AXLin vivo, subcutaneous xenografts of theHNSCC cell lineUM-SCC1and the TNBC cell lineMDAMB231 were generated in nudemice.Once tumors reached approximately 200 mm3, mice wererandomized into groups for treatment with vehicle, R428(50 mg/kg/day), UNC2025 (50 mg/kg/day), or the combinationof R428 and UNC2025 (n ¼ 16 tumors/treatment group). Ther-apies were administered twice daily by oral gavage. Combinedtreatment with R428 and UNC2025 significantly decreasedtumor volume compared with treatment with either inhibitoralone in both models (Fig. 6C and D). Furthermore, IHCanalysis revealed robust decreases in phospho-S6RP and KI67 intumors treated with the combination therapy (�2–25-fold),indicating decreased tumor cell proliferation and downstreamsignaling consistentwith the biochemical and functional effects of
MERTK and AXL coinhibition observed in vitro (Fig. 4). MERTKexpression was also upregulated in R428-treated UM-SCC1 andMDA-MB-231 tumors after 28 days of treatment (Fig. 6C and D),consistent with our previous results in cell culture (Fig. 2) andanimal models (Fig. 6A and B). Collectively, these data indicatethat targeting MERTK can overcome resistance to AXL inhibitionand enhance the therapeutic efficacy of AXL inhibitors in HNSCCand TNBC xenograft models.
DiscussionAXL is overexpressed in numerous cancers and has been asso-
ciated with resistance to both conventional and molecularlytargeted therapies (1, 3, 5, 9, 14–21, 24, 37, 38, 40, 43). Thus,AXL inhibition has emerged as a promising treatment strategy.Although AXL-targeting strategies may have initial clinical benefitin tumor types that overexpress the receptor, intrinsic andacquired resistance to single agent kinase inhibitors is commonand resistance to single-agent AXL inhibitors will likely appear.Our previous studies utilizing shRNA demonstrated oncogenic
Figure 5.
Overexpression of MERTK confers resistance to AXL inhibition. HN30 (A and B) and SUM159 (C, D, and E) cells were transfected with pcDNA6.0-Vector(Vector) or pcDNA6.0-MERTK (MERTK) plasmids. A, C, and D, 24 hours after transfection, cells were treated with R428 (A and C) or Mab173 (D) for anadditional 72 hours, and then relative cell numbers were determined. Mean values, SEs, and statistical analyses were derived from replicates within one experimentand are representative of three independent experiments. �� , P < 0.01. B and E, 72 hours after transfection, cells were harvested and lysates were preparedwith the indicated proteins detected by immunoblot. a-Tubulin is shown as a loading control.
McDaniel et al.
Mol Cancer Ther; 17(11) November 2018 Molecular Cancer Therapeutics2304
on May 28, 2020. © 2018 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 9, 2018; DOI: 10.1158/1535-7163.MCT-17-1239

Figure 6.
Inhibition of AXL increases MERTK protein expression, and coinhibition of AXL and MERTK suppresses tumor growth in vivo. Subcutaneous xenograftswere generated in nude mice using (A and D) the TNBC MDAMB231 cell line, (B) the HNSCC UW-SCC1 patient-derived xenograft (PDX), or (C) the HNSCCUM-SCC1 cell line. Tumor-bearingmice were treated with vehicle, 25mg/kg R428, 25mg/kg UNC2025, or R428þUNC2025 administered twice daily via oral gavage.MERTK expression was assessed in tumor cell lysates by immunoblot. a-Tubulin was used as a loading control. A, MERTK expression was also detected inMDAMB231 tumors using immunohistochemistry. B–D, Tumor volume was measured (n ¼ 8 per group). C and D, pS6RP and KI67 were detected in tumorsections using IHC. Mean values and SEs are shown. � , P < 0.05; �� , P < 0.01; NS ¼ not significant.
MERTK Mediates Resistance to AXL-targeted Therapy
www.aacrjournals.org Mol Cancer Ther; 17(11) November 2018 2305
on May 28, 2020. © 2018 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 9, 2018; DOI: 10.1158/1535-7163.MCT-17-1239

roles for bothMERTKandAXL in astrocytoma andNSCLC (6, 44).Given this functional redundancy, we hypothesized a role forMERTK in resistance to AXL-targeted therapy. In the studiespresented here, single-agent anti-AXL therapy had limited efficacyand the TAM family receptor MERTKwas upregulated in responseto AXL suppression in HNSCC, TNBC, and NSCLC preclinicalmodels. Moreover, ectopic expression of MERTK was sufficient tomediate resistance to AXL-targeting strategies, and combinedinhibition of both AXL and MERTK using a variety of differentapproaches provided potent antitumor activity inHNSCC, TNBC,and NSCLC cell culture and animal models. Collectively, thesedata demonstrate the importance of MERTK in resistance to AXLinhibitors in several tumor types and provide rationale for cotar-geting AXL and MERTK in tumors that express both receptors.
Several preclinical studies have described similar adaptiveresponses to single-agent RTK-targeting strategies. For example,HER2 and HER3 are upregulated in response to EGFR inhibition(29–31, 41), and knockdown of the RTK RON results in upre-gulation of its close family member cMET. Cotargeting of RTKfamily members has been highly beneficial to overcome theseadaptive responses in several tumor models (29–31, 41, 42, 45,46). Additional studies have demonstrated activation of RTKsoutside the immediate family of the targeted kinase, such as theinduction of AXL expression in response to treatment with EGFRinhibitors (5, 11, 17, 43). Although genetic mechanisms such asmutations, polymorphisms, or copy number variations are likelyto mediate or contribute to kinase inhibitor resistance in somecases, these findings illustrate the importance of bypass signalingas a mechanism of resistance and advocate the utility of thera-peutic strategies targeting multiple receptors to prevent develop-ment of resistance consistent with the data presented here.
Regulation of numerous signaling pathways known to playroles in tumorigenesis has been demonstrated downstream ofTAM-family kinases, including RAF/MEK/ERK, JAK/STAT, PI3K/AKT, and mTOR/p70S6K (47). Previous studies showed activa-tion of AKT/mTOR via GAS6 stimulation of MERTK in NSCLCcells (6) and activation of PI3K/P70S6K via a chimeric MERTKreceptor (48). Similarly, data presented here reveal activation ofP70S6K and C-RAF in response to MERTK overexpression (Fig. 5)and potent inhibition of P70S6K, C-RAF, and AKT in response toMERTK and AXL combination therapy (Fig. 4). We have alsorecently found that AXL can mediate EGFR activation throughC-RAF/ERK/C-JUN signaling (43). The therapeutic importance ofsome of these oncogenic pathways associated with TAM familysignaling has been investigated further. Recently, Swick andcolleagues demonstrated that suppression of both PI3K/AKT/mTORC signaling and RAS/RAF/MAPK1 signaling resulted indramatic growth suppression in HNSCC (49). Previous studiesalso associated P70S6K overexpression with increased risk oflocoregional recurrence in early breast cancer patients (50), andPF-4708671, a selective P70S6K inhibitor (51), has been utilizedto reduce local relapse in breast cancer models (52) and inhibitNSCLC tumorigenesis (53). Thus, our observation that combinedtargeting of AXL and MERTK can synergistically inhibit theseoncogenic signaling pathways suggests possible mechanisms forthe synergistic antitumor activity described here. Given the ther-apeutic impact of these signaling pathways, inhibition of thesepathways via combined targeting of AXL andMERTK underscoresthe clinical benefit that this treatment strategy may provide.
Small-molecule inhibitors, mAbs, and soluble decoy receptorstargeting AXL are currently being tested in early-phase clinical
trials for treatment of several different malignancies (Clinical-Trials.gov). R428 (BGB324) is in phase I/II clinical trials enrollingpatients with NSCLC, TNBC, AML, myelodysplastic syndromes,and melanoma. Our data suggest that MERTK upregulation is acommon occurrence in at least the broad group of epithelialmalignancies tested here. Whether increased by AXL inhibitionor present constitutively, MERTK mediated resistance to AXL-targeting agents in functional assays. In addition, cell lines thatexpressed very little or noMERTKproteinwere invariably sensitivetoAXL inhibition and80%of cell lines that expressedhigher levelsof MERTK were resistant to AXL inhibition (Fig. 1). AlthoughMERTKexpression levels alonedidnot always predict resistance toAXL inhibition (e.g.,H1299,UM-SCC4), these data suggest that inmany cases MERTK expression could be useful as a predictivebiomarker to identify tumors that are less likely to respond tosingle-agent AXL therapy. Targetingof a nonmutant RTKalonehasnot been very successful; however, we are learning through mul-tiple examples (e.g., mutant EGFR and ALK) that patients canbenefit from second- and third-generation inhibitors and combi-nations of RTK inhibitors that reduce tumor cell survival signalingand other oncogenic pathways. Indeed, inhibition of MERTKresensitized tumors to AXL inhibition mediated by R428 inpreclinical models, providing strong rationale for clinical devel-opment of therapeutic strategies targeting both AXL and MERTK.
Disclosure of Potential Conflicts of InterestD.K. Graham is founder andmember of the Board of Directors at Meryx and
has ownership interest (including stocks and patents) in Meryx.Inc. S.V. Frye isPresident and CEO at Meryx and has ownership interest (including stocks andpatents) inMeryx.Inc. H.S. Earp is founder and boardmember at Meryx and hasownership interest (including stocks and patents) in Meryx.Inc. X. Wang andD. DeRyckere have ownership interest (including stocks and patents) in Meryx,Inc. No potential conflicts of interest were disclosed by the other authors.
DisclaimerThe content is solely the responsibility of the authors and does not neces-
sarily represent the official views of the NIH.
Authors' ContributionsConception and design: N.K. McDaniel, C.T. Cummings, X. Wang, H.S. Earp,R.J. Kimple, D. DeRyckere, D.K. Graham, D.L. WheelerDevelopment of methodology: N.K. McDaniel, C.T. Cummings, M. Iida,D.L. WheelerAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): N.K. McDaniel, C.T. Cummings, J. H€ulse,H.E. Pearson, E. Vasileiadi, R.E. Parker, R.A. Orbuch, N.B. Welke, K.D. Davies,P.M. Harari, R.J. KimpleAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): N.K. McDaniel, C.T. Cummings, M. Iida, J. H€ulse,E. Vasileiadi, R.A. Orbuch, O.J. Ondracek, K.D. Davies, R.J. Kimple,D. DeRyckere, D.K. Graham. D.L. WheelerWriting, review, and/or revision of the manuscript: N.K. McDaniel,C.T. Cummings, M. Iida, H.E. Pearson, R.A. Orbuch, X. Wang, S.V. Frye,H.S. Earp, P.M. Harari, R.J. Kimple, D. DeRyckere, D.K. Graham, D.L. WheelerAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): N.K. McDaniel, M. Iida, G.H. Kang, X. Wang,D.L. WheelerStudy supervision: N.K. McDaniel, R.J. Kimple, D. DeRyckere, D.L. WheelerOther (supervised design of MerTK inhibitors used in this study): S.V. Frye
AcknowledgmentsWe thank Parkash Gill at the University of Southern California for use of the
AXL antibody (Mab173) and the Daniel Horyn Foundation for the gift ofesophageal cell lines. The authors also thankMasami Glines for expert technicalassistance, and the University of Wisconsin Carbone Cancer Center Support
Mol Cancer Ther; 17(11) November 2018 Molecular Cancer Therapeutics2306
McDaniel et al.
on May 28, 2020. © 2018 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 9, 2018; DOI: 10.1158/1535-7163.MCT-17-1239

(grant no. P30 CA014520). Research reported in this publication wassupported by the Wisconsin Head & Neck Cancer SPORE (grant no. P50DE026787, to D.L. Wheeler, R.J. Kimple, and P.M. Harari), Vilas FacultyEarly Career Investigator Award (grant no. MSN189325, to D.L. Wheeler), anNIH SPORE in Lung Cancer (grant no. P50CA058187, to K.D. Davies,), theLung Cancer Colorado Fund (to K.D. Davies), and an NIH grant fromCellular and Molecular Pathology Graduate Training Program (grant no.5 T32 GM81061-7, to N.K. McDaniel).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received December 22, 2017; revised May 23, 2018; accepted July 31, 2018;published first August 9, 2018.
References1. Linger RM, Keating AK, Earp HS, Graham DK. TAM receptor tyrosine
kinases: biologic functions, signaling, and potential therapeutic targetingin human cancer. Adv Cancer Res 2008;100:35–83.
2. Verma A, Warner SL, Vankayalapati H, Bearss DJ, Sharma S. Targeting Axland Mer kinases in cancer. Mol Cancer Ther 2011;10:1763–73.
3. Li Y, Ye X, Tan C, Hongo JA, Zha J, Liu J, et al. Axl as a potential therapeutictarget in cancer: role of Axl in tumor growth, metastasis and angiogenesis.Oncogene 2009;28:3442–55.
4. Linger RM, Keating AK, Earp HS, Graham DK. Taking aim at Mer and Axlreceptor tyrosine kinases as novel therapeutic targets in solid tumors. ExpertOpin Ther Targets 2010;14:1073–90.
5. Zhang Z, Lee JC, Lin L,Olivas V, AuV, LaFramboise T, et al. Activation of theAXL kinase causes resistance to EGFR-targeted therapy in lung cancer. NatGenet 2012;44:852–60.
6. Linger RM, Cohen RA, Cummings CT, Sather S, Migdall-Wilson J,Middleton DH, et al. Mer or Axl receptor tyrosine kinase inhibitionpromotes apoptosis, blocks growth and enhances chemosensitivity ofhuman non-small cell lung cancer. Oncogene 2013;32:3420–31.
7. Byers LA, Diao L, Wang J, Saintigny P, Girard L, Peyton M, et al. Anepithelial-mesenchymal transition gene signature predicts resistance toEGFR and PI3K inhibitors and identifies Axl as a therapeutic target forovercoming EGFR inhibitor resistance. Clin Cancer Res 2013;19:279–90.
8. Ye X, Li Y, Stawicki S, Couto S, Eastham-Anderson J, Kallop D, et al. Ananti-Axl monoclonal antibody attenuates xenograft tumor growth andenhances the effect of multiple anticancer therapies. Oncogene 2010;29:5254–64.
9. Shieh YS, Lai CY, Kao YR, Shiah SG, Chu YW, Lee HS, et al. Expression ofaxl in lung adenocarcinoma and correlation with tumor progression.Neoplasia 2005;7:1058–64.
10. Holland SJ, Pan A, Franci C, Hu Y, Chang B, Li W, et al. R428, a selectivesmall molecule inhibitor of Axl kinase, blocks tumor spread and prolongssurvival in models of metastatic breast cancer. Cancer Res 2010;70:1544–54.
11. Meyer AS, Miller MA, Gertler FB, Lauffenburger DA. The receptor AXLdiversifies EGFR signaling and limits the response to EGFR-targeted inhi-bitors in triple-negative breast cancer cells. Sci Signal 2013;6:ra66.
12. Zhang YX, Knyazev PG, Cheburkin YV, Sharma K, Knyazev YP, Orfi L, et al.AXL is a potential target for therapeutic intervention in breast cancerprogression. Cancer Res 2008;68:1905–15.
13. Asiedu MK, Beauchamp-Perez FD, Ingle JN, Behrens MD, Radisky DC,Knutson KL. AXL induces epithelial-to-mesenchymal transition andregulates the function of breast cancer stem cells. Oncogene 2014;33:1316–24.
14. D'Alfonso TM, Hannah J, Chen Z, Liu Y, Zhou P, Shin SJ. Axl receptortyrosine kinase expression in breast cancer. J Clin Pathol 2014;67:690–6.
15. Rankin EB, Fuh KC, Taylor TE, Krieg AJ, Musser M, Yuan J, et al. AXL is anessential factor and therapeutic target formetastatic ovarian cancer. CancerRes 2010;70:7570–9.
16. DunnePD,McArtDG, Blayney JK, KalimuthoM,Greer S,Wang T, et al. AXLis a key regulator of inherent and chemotherapy-induced invasion andpredicts a poor clinical outcome in early-stage colon cancer. Clin CancerRes 2014;20:164–75.
17. Giles KM, Kalinowski FC, Candy PA, EpisMR, Zhang PM, Redfern AD, et al.Axl mediates acquired resistance of head and neck cancer cells to theepidermal growth factor receptor inhibitor erlotinib. Mol Cancer Ther2013;12:2541–58.
18. Avilla E, Guarino V, Visciano C, Liotti F, SveltoM, Krishnamoorthy G, et al.Activation of TYRO3/AXL tyrosine kinase receptors in thyroid cancer.Cancer Res 2011;71:1792–804.
19. Paccez JD, Vasques GJ, Correa RG, Vasconcellos JF, Duncan K, Gu X, et al.The receptor tyrosine kinase Axl is an essential regulator of prostate cancerproliferation and tumor growth and represents a new therapeutic target.Oncogene 2013;32:689–98.
20. Song X, Wang H, Logsdon CD, Rashid A, Fleming JB, Abbruzzese JL, et al.Overexpression of receptor tyrosine kinase Axl promotes tumor cell inva-sion and survival in pancreatic ductal adenocarcinoma. Cancer2011;117:734–43.
21. Han J, Tian R, Yong B, Luo C, Tan P, Shen J, et al. Gas6/Axl mediates tumorcell apoptosis, migration and invasion and predicts the clinical outcomeof osteosarcoma patients. Biochem Biophys Res Commun 2013;435:493–500.
22. Muller J, Krijgsman O, Tsoi J, Robert L, Hugo W, Song C, et al. Low MITF/AXL ratio predicts early resistance tomultiple targeted drugs inmelanoma.Nat Commun 2014;5:5712.
23. Huey MG, Minson KA, Earp HS, DeRyckere D, Graham DK. Targeting theTAM receptors in leukemia. Cancers 2016;8:101.
24. Liu R, GongM, Li X, Zhou Y, GaoW, Tulpule A, et al. Induction, regulation,and biologic function of Axl receptor tyrosine kinase in Kaposi sarcoma.Blood 2010;116:297–305.
25. BerGenBio. Pipeline: BGB324. Available from: http://www.bergenbio.com/pipeline/bgb324/.
26. Zhang W, DeRyckere D, Hunter D, Liu J, Stashko MA, Minson KA, et al.UNC2025, a potent and orally bioavailable MER/FLT3 dual inhibitor.J Med Chem 2014;57:7031–41.
27. DeRyckere D, Lee-Sherick AB, Huey MG, Hill AA, Tyner JW, Jacobsen KM,et al. UNC2025, a MERTK small-molecule inhibitor, is therapeuticallyeffective alone and in combinationwithmethotrexate in leukemiamodels.Clin Cancer Res 2017;23:1481–92.
28. Cummings CT, ZhangW, Davies KD, Kirkpatrick GD, Zhang D, DeRyckereD, et al. Small molecule inhibition of MERTK is efficacious in non-smallcell lung cancermodels independent of driver oncogene status.Mol CancerTher 2015;14:2014–22.
29. Chen FL, Xia W, Spector NL. Acquired resistance to small molecule ErbB2tyrosine kinase inhibitors. Clin Cancer Res 2008;14:6730–4.
30. Mancini M, Gaborit N, LindzenM, Salame TM, Dall'Ora M, Sevilla-SharonM, et al. Combining three antibodies nullifies feedback-mediated resis-tance to erlotinib in lung cancer. Sci Signal 2015;8:ra53.
31. Yamaguchi H, Chang SS, Hsu JL, Hung MC. Signaling cross-talk in theresistance to HER family receptor targeted therapy. Oncogene2014;33:1073–81.
32. Iida M, Brand TM, Starr MM, Li C, Huppert EJ, Luthar N, et al. Sym004,a novel EGFR antibody mixture, can overcome acquired resistance tocetuximab. Neoplasia 2013;15:1196–206.
33. Li C, Brand TM, Iida M, Huang S, Armstrong EA, van der Kogel A, et al.Human epidermal growth factor receptor 3 (HER3) blockade withU3-1287/AMG888 enhances the efficacy of radiation therapy in lung andhead and neck carcinoma. Discov Med 2013;16:79–92.
34. Chou TC.Drug combination studies and their synergy quantification usingthe Chou-Talalay method. Cancer Res 2010;70:440–6.
35. Chou TC, Talalay P. Quantitative analysis of dose-effect relationships: thecombined effects of multiple drugs or enzyme inhibitors. Adv EnzymeRegul 1984;22:27–55.
36. Irwin ME, Mueller KL, Bohin N, Ge Y, Boerner JL. Lipid raft localization ofEGFR alters the response of cancer cells to the EGFR tyrosine kinaseinhibitor gefitinib. J Cell Physiol 2011;226:2316–28.
37. von Massenhausen A, Bragelmann J, Billig H, Thewes B, Queisser A, VogelW, et al. Implication of the receptor tyrosine kinase AXL in head and neckcancer progression. Int J Mol Sci 2016;18.
MERTK Mediates Resistance to AXL-targeted Therapy
www.aacrjournals.org Mol Cancer Ther; 17(11) November 2018 2307
on May 28, 2020. © 2018 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 9, 2018; DOI: 10.1158/1535-7163.MCT-17-1239

38. Brand TM, Iida M, Stein AP, Corrigan KL, Braverman CM, Coan JP, et al.AXL is a logical molecular target in head and neck squamous cell carci-noma. Clin Cancer Res 2015;21:2601–12.
39. Leconet W, Chentouf M, du Manoir S, Chevalier C, Sirvent A, Ait-Arsa I,et al. Therapeutic activity of anti-AXL antibody against triple-negativebreast cancer patient-derived xenografts and metastasis. Clin Cancer Res2017;23:2806–16.
40. WuX,MaW, ZhouQ, YanH, LimZF,HuangM, et al. AXL-GAS6 expressioncan predict for adverse prognosis in non-small cell lung cancer with brainmetastases. J Cancer Res Clin Oncol 2017;143:1947–57.
41. Wheeler DL, Huang S, Kruser TJ, Nechrebecki MM, Armstrong EA,Benavente S, et al. Mechanisms of acquired resistance to cetux-imab: role of HER (ErbB) family members. Oncogene 2008;27:3944–56.
42. Iida M, Bahrar H, Brand TM, Pearson HE, Coan JP, Orbuch RA, et al.Targeting theHER family with Pan-HER effectively overcomes resistance tocetuximab. Mol Cancer Ther 2016;15:2175–86.
43. Brand TM, Iida M, Stein AP, Corrigan KL, Braverman CM, Luthar N, et al.AXL mediates resistance to cetuximab therapy. Cancer Res 2014;74:5152–64.
44. Keating AK, Kim GK, Jones AE, Donson AM, Ware K, Mulcahy JM, et al.Inhibition of Mer and Axl receptor tyrosine kinases in astrocytoma cellsleads to increased apoptosis and improved chemosensitivity. Mol CancerTher 2010;9:1298–307.
45. Iida M, Brand TM, Starr MM, Huppert EJ, Luthar N, Bahrar H, et al.Overcoming acquired resistance to cetuximab by dual targeting HER
family receptors with antibody-based therapy. Mol Cancer 2014;13:242.
46. Kawada I, Hasina R, Arif Q, Mueller J, Smithberger E, Husain AN, et al.Dramatic antitumor effects of the dual MET/RON small-molecule inhib-itor LY2801653 in non-small cell lung cancer. Cancer Res 2014;74:884–95.
47. Graham DK, DeRyckere D, Davies KD, Earp HS. The TAM family: phos-phatidylserine sensing receptor tyrosine kinases gone awry in cancer. NatRev Cancer 2014;14:769–85.
48. Ling L, Kung HJ. Mitogenic signals and transforming potential of Nyk, anewly identified neural cell adhesion molecule-related receptor tyrosinekinase. Mol Cell Biol 1995;15:6582–92.
49. Swick AD, Prabakaran PJ, Miller MC, Javaid AM, Fisher MM, Sampene E,et al. Cotargeting mTORC and EGFR signaling as a therapeutic strategy inHNSCC. Mol Cancer Ther 2017;16:1257–68.
50. van der Hage JA, van den Broek LJ, Legrand C, Clahsen PC, Bosch CJ,Robanus-Maandag EC, et al. Overexpression of P70 S6 kinase protein isassociated with increased risk of locoregional recurrence in node-negativepremenopausal early breast cancer patients. Br J Cancer 2004;90:1543–50.
51. Pearce LR, Alton GR, Richter DT, Kath JC, Lingardo L, Chapman J, et al.Characterization of PF-4708671, a novel and highly specific inhibitor ofp70 ribosomal S6 kinase (S6K1). Biochem J 2010;431:245–55.
52. Segatto I, Berton S, Sonego M, Massarut S, D'Andrea S, Perin T, et al.Inhibition of breast cancer local relapse by targeting p70S6 kinase activity.J Mol Cell Biol 2013;5:428–31.
53. Qiu ZX, SunRF,MoXM, LiWM. The p70S6K specific inhibitor PF-4708671impedes non-small cell lung cancer growth. PLoSOne 2016;11:e0147185.
Mol Cancer Ther; 17(11) November 2018 Molecular Cancer Therapeutics2308
McDaniel et al.
on May 28, 2020. © 2018 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 9, 2018; DOI: 10.1158/1535-7163.MCT-17-1239

2018;17:2297-2308. Published OnlineFirst August 9, 2018.Mol Cancer Ther Nellie K. McDaniel, Christopher T. Cummings, Mari Iida, et al. AXL-targeting AgentsMERTK Mediates Intrinsic and Adaptive Resistance to
Updated version
10.1158/1535-7163.MCT-17-1239doi:
Access the most recent version of this article at:
Material
Supplementary
http://mct.aacrjournals.org/content/suppl/2018/08/09/1535-7163.MCT-17-1239.DC1
Access the most recent supplemental material at:
Cited articles
http://mct.aacrjournals.org/content/17/11/2297.full#ref-list-1
This article cites 51 articles, 25 of which you can access for free at:
Citing articles
http://mct.aacrjournals.org/content/17/11/2297.full#related-urls
This article has been cited by 2 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mct.aacrjournals.org/content/17/11/2297To request permission to re-use all or part of this article, use this link
on May 28, 2020. © 2018 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 9, 2018; DOI: 10.1158/1535-7163.MCT-17-1239