melatonin inhibits apoptosis during early b-cell development in mouse bone marrow
TRANSCRIPT

J. Pineal Res. 2000; 29:86–93
Melatonin inhibits apoptosis during earlyB-cell development in mouse bone marrow
Yu Q, Miller SC, Osmond DG. Melatonin inhibits apoptosis duringearly B-cell development in mouse bone marrow. J. Pineal Res. 2000;2:86–93. © Munksgaard, Copenhagen
Abstract: The pineal secretory product, melatonin, exerts a variety ofeffects on the immune system. Administration of melatonin stimulatescell-mediated immunity, particularly by inhibiting apoptosis among Tlymphocytes in the thymus and inducing production of T-cell-derivedcytokines. However, its possible effects on the humoral immune systemare unclear. In the present study, we have examined whether melatoninmay influence the in vivo development of B lymphocytes in mousebone marrow, a process in which apoptosis is normally a prominentfeature. Double immunofluorescence labeling and flow cytometry wereused to quantitate phenotypically defined precursor B-cell and matureB-cell populations and their apoptotic rates in bone marrow of micefed either melatonin-containing or control diet for 16 days from 9 wkof age. In short-term bone marrow cultures, the incidence of apoptosisamong large pre-B cells, including cells expressing the l5 component ofpre-B-cell receptor, was markedly reduced in melatonin-treated mice,associated with an increase in the absolute number of large pre-B cellsin bone marrow. In contrast, apoptosis of earlier precursor B cells andmature B lymphocytes did not differ from control values. The resultsindicate that orally administered melatonin can substantially promotethe survival of precursor B cells in mouse bone marrow. Melatonintreatment may thus boost the survival of newly formed B cellsmediating humoral immunity.
Qingnan Yu, Sandra C. Miller andDennis G. OsmondDepartment of Anatomy and Cell Biology,McGill University, Montreal, Quebec, Canada
Key words: apoptosis – B cell – bonemarrow – melatonin – pre-B cell
Address reprint requests to Dr. D.G. Os-mond, Department of Anatomy and Cell Biol-ogy, McGill University, 3640 UniversityStreet, Montreal, Quebec, H3A 2B2 Canada.E-mail: [email protected]
Received July 13, 1999;accepted October 18, 1999.
Introduction
Melatonin is produced by the pineal gland mainlyduring hours of darkness to coordinate circadianbiological rhythms [Nelson and Demas, 1996;Mazzoccoli et al., 1997]. Among its many activi-ties, melatonin appears to be capable of playing animmunoregulatory role [Guerrero and Reiter,1992; Liebmann et al., 1997], especially by exertingan anti-apoptotic effect on cells of the immuneand hemopoietic systems [Maestroni et al., 1994a;Sainz et al., 1995; Maestroni and Conti, 1996;Provinciali et al., 1996; Maestroni, 1998].There areclear-cut effects on T and natural killer (NK) cellsconcerned with cell-mediated immunity and onphagocyte cell systems in the bone marrow [Gon-zalez-Haba et al., 1995; Sainz et al., 1995;Maestroni and Conti, 1996; Provinciali et al.,1996; Champney et al., 1997; Pozo et al., 1997;Demas and Nelson, 1998; N. L. Currier, L. Sun, S.C. Miller, unpublished observations]. However,the possible role of melatonin in modulating theB-cell lineage and the humoral immune system
remains unclear [Champney et al., 1998; Demasand Nelson, 1998]. Melatonin receptor (Mel1a)mRNA is expressed by B lymphocytes in ratspleen and thymus [Pozo et al., 1997], and specificbinding sites for melatonin have been reported inthe avian bursa of Fabricius [Calvo et al., 1995],but melatonin treatment of mice and hamsters hasproduced no change in immunoglobulin M (IgM)or IgG antibody levels [Champney et al., 1997,1998; Demas and Nelson, 1998]. The melatoninreceptor, found on both cell and nuclear mem-branes, may play a role in mediating anti-apop-totic effects. On the other hand, melatonin itself isan effective free radical scavenger and anti-oxidant[Hardeland et al., 1993; Inserra et al., 1998], sothat, at least in part, its anti-apoptotic effects maybe receptor independent. The possible actions ofmelatonin on cell development and survival duringB lymphopoiesis in bone marrow are unknown.
B lymphocytes are produced throughout lifefrom precursor cells located in mouse bone mar-row [Osmond, 1990; Rolink and Melchers, 1993;Osmond et al., 1994]. The process is marked by
86 Printed in Ireland—all rights reser7ed.

Melatonin inhibits apoptosis of pre-B cells
large-scale apoptotic cell death [Lu and Osmond,1997], constituting an important quality controlmechanism to eliminate functionless or poten-tially harmful cells and to regulate the numberof newly formed B cells entering the immunesystem [Melchers et al., 1995; Rajewsky, 1996].At the earliest defined (pro-B cell) stage of devel-opment, actively proliferating B lineage cells un-dergo rearrangement of gene segments toconstitute functional genes encoding the m heavy(H) chain of IgM [Oettinger et al., 1990]. Thismolecule forms part of the B-cell antigen recep-tor (BCR) that characterizes all mature Blymphocytes. Most of the resulting m chains areat first retained within the cytoplasm (cm), defin-ing the pre-B-cell stage, while some appear at thecell surface bound to a surrogate light (L) chainas part of a pre-B-cell receptor complex (pre-BCR) [Karasuyama et al., 1994; Osmond et al.,1998]. Many cells, however, which either sustainerrors of DNA recombination or fail to makeproductive IgH gene rearrangements and expresspre-BCR, become diverted into an apoptotic celldeath pathway [Melchers et al., 1995; Rajewsky,1996]. At this pro-B/pre-B checkpoint, as manyas 70% of developing B cells normally undergoapoptosis [Opstelten and Osmond, 1983; Lu etal., 1999]. Surviving cells cease dividing, formingsmall pre-B cells that then rearrange L chaingene segments and synthesize L chains to form Blymphocytes expressing complete IgM moleculesat the cell surface (sIgM). A second apoptoticcheckpoint at this stage deletes many cells ex-pressing potentially self-reactive IgM [Melcherset al. 1995]. B lymphopoiesis and apoptosis inbone marrow are influenced by a variety of sys-temic factors [Kincade, 1994; Medina and Kin-cade, 1994; Fauteux and Osmond, 1996] andlocal microenvironmental factors, notably stro-mal cell-derived interleukin-7 (IL-7) [Henney,1989; Hayashi et al., 1990; Kincade, 1991; Valen-zona et al., 1998], but the possible contributionsof melatonin to B-cell homeostasis and survivalin bone marrow have not been determined.
In the present work, using double immu-nofluorescence labeling, flow cytometry, and sen-sitive assays of apoptotic activity, we examinewhether orally administered melatonin may mod-ulate the in vivo development and selection ofprecursor B cells in mouse bone marrow. In thebone marrow of melatonin-treated and controlmice, we have quantitated phenotypically definedpopulations of precursor B cells and assessedtheir apoptotic frequency and cell cycle charac-teristics by DNA content, both ex vivo and inshort-term culture.
Methods and materials
Mice
Female C3H/HeJ mice (The Jackson Laboratory,Bar Harbor, ME) were housed in microisolatorcages subject to a 12-hr light/dark cycle (06:00–18:00 hr). Mice were allowed to settle into ourfacility for 1 wk before use.
Melatonin administration
Melatonin (Schiff Products Inc., Salt Lake City,UT) was administered in the diet, ground homo-geneously into pulverized standard mouse chow.Mice were housed four per cage and were pro-vided every evening (17:00 hr) with the groundchow with or without (control) melatonin (14mg/mouse/day) starting at 9 wk of age. Micewere killed after 16 days. Although melatonin-containing chow was available throughout the24-hr period, the food was consumed almost ex-clusively during the night time. The mice re-mained in the rest/sleep phase during the day.The dose of melatonin was based on an averagedaily food consumption per mouse of 5.5 g/24hr, as previously established [N. L. Currier, L.Sun, S. C. Miller, unpublished observations]. Thedose of melatonin/mouse was selected from dose/body weight studies in adult mice of otherstrains [Maestroni et al., 1994a; Demas and Nel-son, 1998]. Since inter-cage difference in foodconsumption was insignificant during any 24-hrperiod, it is assumed that during the long periodof melatonin administration, minor fluctuationsin daily consumption of the agent by individualmice would have been canceled out.
Bone marrow cell suspension and short-term culture
Mice were killed by cervical dislocation andfemoral marrow cells were flushed out, as previ-ously described [Lu and Osmond, 1997]. Afterwashing with newborn calf serum (NCS; LifeTechnologies, Grand Island, NY), nucleated cellswere counted using an electronic particle counter(Coulter Corp., Burlington, Ontario, Canada).Cell suspensions were adjusted to 40×106 cells/mL in minimum essential medium (MEM) withL-glutamine (Life Technologies) supplementedwith 10% (v/v) NCS. Aliquots of cell suspensions(100 mL) were either assayed immediately or in-cubated in MEM-NCS for 4 hr at 37°C with 5%CO2, as previously described [Lu and Osmond,1997].
87

Yu et al.
Immunophenotypic staining of B lineage cells
For phenotypic study, bicolor immunolabelingwas carried out paired as B220:total m and sur-face IgM:total m. For apoptotic study, single-color immunolabeling was carried out beforeDNA labeling. To label surface determinants,bone marrow cell samples were incubated withfluorescein isothiocyanate (FITC) -conjugatedand R-phycoerythrin (PE) -conjugated anti-mouse B220 monoclonal antibody (mAb) (RA3-6B2; Pharmingen, San Diego, CA) andFITC-conjugated and PE-conjugated goat anti-mouse IgM (m heavy chain specific) (SouthernBiotechnology, Birmingham, AL), respectively(15 min, 4°C, in dark), as previously described[Lu and Osmond, 1997]. To examine total m ex-pression, including both cytoplasmic m chainsand the m chains of surface IgM molecules,ethanol-fixed bone marrow cells were incubatedwith FITC-conjugated goat anti-mouse IgM(Pharmingen) mAb for 1 hr. Samples stainedwith isotype-matched mAbs of irrelevant specific-ity were used as negative controls.
DNA content staining
After immunofluorescent phenotypic staining,cell samples were washed with NCS and Ca2+-and Mg2+-free phosphate buffered saline (PBS;Life Technologies), fixed dropwise with ice-cold70% ethanol (50 min, 4°C), and washed exten-sively with PBS. Samples were suspended in 50mg/mL RNase (Boehringer Mannheim Corp.,Mannheim, Germany) and 50 mg/mL propidiumiodide (PI; Sigma Chemical Co., St. Louis, MO)in PBS and kept on ice in dark until analyzed,as previously described [Lu and Osmond, 1997].
Flow-cytometric analysis of B-cell phenotypes and apoptosis
After phenotypic and PI staining, single-cell sus-pensions were analyzed using a FACScan flowcytometer (Becton Dickinson, Mountain View,CA) equipped with a doublet discriminationmodule. FITC, PE, and PI were excited with the480 nm line of an argon laser and emissionswere detected at 550 and 650 nm, respectively. Aminimum of 10,000 events was collected for eachsample, gated on forward scatter versus sidescatter to exclude debris and cell clumps. Datawere acquired and analyzed using FACScanLYSYS II software (Becton Dickinson). Pheno-typic populations were identified by positivestaining of each marker, large and small cellsbeing identified by gating on forward scatter ver-
sus side scatter on the dot plots. Apoptotic cellswere identified in the hypodiploid region ofDNA content profiles, as previously described[Lu and Osmond, 1997].
Results
Precursor B-cell populations in the bone marrow ofmelatonin-treated mice
The total number of nucleated cells recoveredfrom the bone marrow of mice given dietarymelatonin for 16 days was not significantly dif-ferent from that of age- and sex-matched controlmice (10.690.7 versus 10.290.5×106 cells/fe-mur). Cells expressing the B lineage-associatedsurface glycoprotein, B220, showed a small butstatistically non-significant increase in meannumber of cells per femur (25.490.9 versus23.391.2×105 cells/femur). In individual experi-ments, however, B220+ cells consistently showedsmall increases in absolute number, comparedwith their respective control values. To examinethe possible effects of melatonin at particularpoints during the process of B lymphopoiesis,double immunofluorescence labeling and flow cy-tometry were used to define three successivestages of B-cell differentiation, as shown in Fig.1. Pro-B cells express B220 in the absence of mchains; pre-B cells express cm but not sIgM, andthey include both large dividing pre-B cells andsmall postmitotic pre-B cells; B cells expresssIgM [Osmond et al., 1994, 1998]. The numberof B lineage cells in melatonin-treated miceshowed consistent but statistically non-significantincreases in the early stages of B-cell develop-ment (Fig. 2).
Fig. 1. Scheme of B lymphocyte development in normalmouse bone marrow, showing phenotypic differentiationmarkers defining the developmental stages of B lineage cellsand the stages at which apoptotic cell death is particularlyprominent.
88

Melatonin inhibits apoptosis of pre-B cells
Fig. 2. Number of B lineage cells of various differentiationstages (phenotypes) in the bone marrow of melatonin-treatedmice and control mice. Mean9S.E. derived from eight sepa-rate experiments.
Inhibited apoptosis of pre-B-cell receptor-bearing cells inmelatonin-treated mice
Pre-BCR+ cells, detected by immunofluorescencelabeling of the l5 component of the surrogatelight chain at the cell surface, totaled 1.590.3% ofall nucleated cells in control bone marrow, repre-senting 20% of the total pre-B-cell population,consistent with previous findings [Karasuyama etal., 1994]. The apoptotic rate of pre-BCR+ cells,normally one-third of that of the large pre-B-cellpopulation as a whole (Figs. 4 and 5), was furtherreduced to only half control levels in melatonin-treated mice (Fig. 5), but with no significantchange in population size (Fig. 5).
Decreased apoptosis among pre-B cells of melatonin-treatedmice
We have previously shown that dissociated bonemarrow cell suspensions in short-term culture un-dergo apoptosis at a linear rate, the apoptotic cellsaccumulating in intact form without being in-gested by macrophages, thus forming a sensitiveassay by which to compare rates of spontaneousapoptosis between bone marrow samples [Lu andOsmond, 1997; Lu et al., 1999]. Double immu-nofluorescence labeling of DNA in conjunctionwith B lineage phenotypic markers after 4 hr invitro incubation of bone marrow suspensions frommelatonin-treated mice and control mice has re-vealed the incidence of hypodiploid cells (Fig. 3),previously shown to be a reliable criterion ofapoptosis among mouse bone marrow cells [Luand Osmond, 1997].
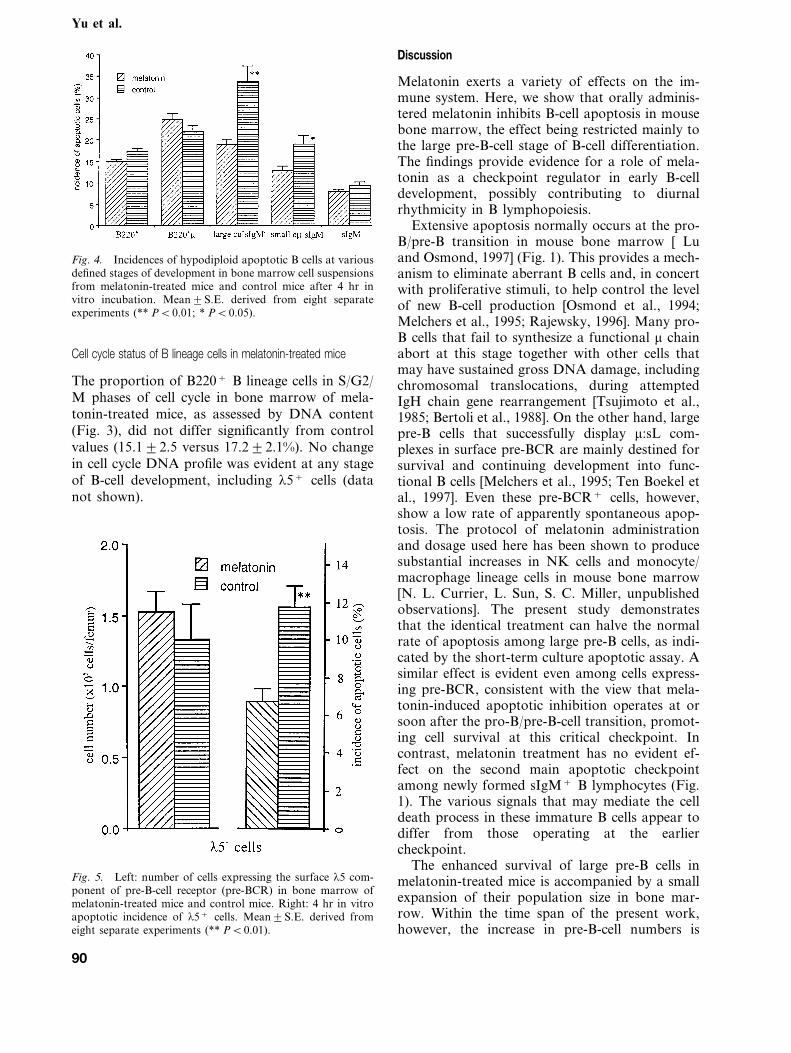
In individual experiments, the incidence ofapoptotic cells among B lineage cells was lower inmelatonin-treated mice than control mice (Fig. 3).Calculation of the apoptotic incidences amongsuccessive precursor B-cell populations revealedthat this reduction in apoptotic rate resulted froma marked suppression of apoptosis among pre-Bcells, the effect being restricted mainly to the pop-ulation of large, mitotically active pre-B cells (PB0.01) (Fig. 4). Whereas the hypodiploid incidenceof large pre-B cells from control mice increasedrapidly during incubation to reach 33.297.5% by4 hr (Fig. 4), apoptosis of large pre-B cells frommelatonin-treated mice only reached just over halfthe normal values (19.193.1%) (Fig. 4). In con-trast, the numbers of apoptotic pro-B cells andsIgM+ B lymphocytes did not differ significantlyfrom normal values (Fig. 4).
Fig. 3. Representative histograms showing hypodiploidapoptotic cells among B lineage cells in bone marrow suspen-sions from melatonin-treated mice and control mice after 4 hrin vitro incubation. B220+ cells include all identifiable Blineage cells. Total m-bearing (tm+) cells include pre-B cellsand B cells. sIgM+ cells include immature and mature B cells.Hypodiploid incidences are indicated.
89
Yu et al.
Fig. 4. Incidences of hypodiploid apoptotic B cells at variousdefined stages of development in bone marrow cell suspensionsfrom melatonin-treated mice and control mice after 4 hr invitro incubation. Mean9S.E. derived from eight separateexperiments (** PB0.01; * PB0.05).
Discussion
Melatonin exerts a variety of effects on the im-mune system. Here, we show that orally adminis-tered melatonin inhibits B-cell apoptosis in mousebone marrow, the effect being restricted mainly tothe large pre-B-cell stage of B-cell differentiation.The findings provide evidence for a role of mela-tonin as a checkpoint regulator in early B-celldevelopment, possibly contributing to diurnalrhythmicity in B lymphopoiesis.
Extensive apoptosis normally occurs at the pro-B/pre-B transition in mouse bone marrow [ Luand Osmond, 1997] (Fig. 1). This provides a mech-anism to eliminate aberrant B cells and, in concertwith proliferative stimuli, to help control the levelof new B-cell production [Osmond et al., 1994;Melchers et al., 1995; Rajewsky, 1996]. Many pro-B cells that fail to synthesize a functional m chainabort at this stage together with other cells thatmay have sustained gross DNA damage, includingchromosomal translocations, during attemptedIgH chain gene rearrangement [Tsujimoto et al.,1985; Bertoli et al., 1988]. On the other hand, largepre-B cells that successfully display m:sL com-plexes in surface pre-BCR are mainly destined forsurvival and continuing development into func-tional B cells [Melchers et al., 1995; Ten Boekel etal., 1997]. Even these pre-BCR+ cells, however,show a low rate of apparently spontaneous apop-tosis. The protocol of melatonin administrationand dosage used here has been shown to producesubstantial increases in NK cells and monocyte/macrophage lineage cells in mouse bone marrow[N. L. Currier, L. Sun, S. C. Miller, unpublishedobservations]. The present study demonstratesthat the identical treatment can halve the normalrate of apoptosis among large pre-B cells, as indi-cated by the short-term culture apoptotic assay. Asimilar effect is evident even among cells express-ing pre-BCR, consistent with the view that mela-tonin-induced apoptotic inhibition operates at orsoon after the pro-B/pre-B-cell transition, promot-ing cell survival at this critical checkpoint. Incontrast, melatonin treatment has no evident ef-fect on the second main apoptotic checkpointamong newly formed sIgM+ B lymphocytes (Fig.1). The various signals that may mediate the celldeath process in these immature B cells appear todiffer from those operating at the earliercheckpoint.
The enhanced survival of large pre-B cells inmelatonin-treated mice is accompanied by a smallexpansion of their population size in bone mar-row. Within the time span of the present work,however, the increase in pre-B-cell numbers is
Cell cycle status of B lineage cells in melatonin-treated mice
The proportion of B220+ B lineage cells in S/G2/M phases of cell cycle in bone marrow of mela-tonin-treated mice, as assessed by DNA content(Fig. 3), did not differ significantly from controlvalues (15.192.5 versus 17.292.1%). No changein cell cycle DNA profile was evident at any stageof B-cell development, including l5+ cells (datanot shown).
Fig. 5. Left: number of cells expressing the surface l5 com-ponent of pre-B-cell receptor (pre-BCR) in bone marrow ofmelatonin-treated mice and control mice. Right: 4 hr in vitroapoptotic incidence of l5+ cells. Mean9S.E. derived fromeight separate experiments (** PB0.01).
90

Melatonin inhibits apoptosis of pre-B cells
proportionately less than the degree of apoptoticsuppression and no increase in the number of Bcells at later differentiation stages is evident. Possi-bly, after an initial lag period required for themelatonin treatment to become apoptotically effec-tive, an elevated steady-state level of B lympho-poiesis has yet to be fully achieved. Ananti-proliferative effect has been proposed to con-tribute to the anti-apoptotic effect of melatonin[Mayo et al., 1998]. However, DNA content analy-sis showed no effect of melatonin treatment on thecell cycle of precursor B cells at any defined stageof development in the present work.
The anti-apoptotic activity of melatonin treat-ment on B cells in bone marrow resembles in somerespects its action in preventing physiologic apop-tosis of T-cell precursors in the thymus [Sainz et al.,1995; Provinciali et al., 1996; Pozo et al., 1997].However, in contrast to the stage-restricted effecton B lymphopoiesis, melatonin appears to act on Tlineage cells throughout all stages of their develop-ment into mature T-helper and T-cytotoxic cells.
Melatonin may exert apoptosis-suppressive ef-fects by both direct and indirect mechanisms. Act-ing via specific receptors, melatonin can directlystimulate CD4+ T-helper cells to produce a varietyof cytokines and neuropeptides (kappa-opioid cy-tokine peptides) [Maestroni et al., 1996; Garcia-Maurino et al., 1997]. On the other hand, thehematopoietic effects of melatonin treatment ap-pear to result indirectly from a complex cytokinecascade. Hematopoietic apoptotic protection inbone marrow induced by melatonin is both granu-locyte macrophage-colony stimulating factor (GM-CSF) dependent and T-cell dependent, and, inaddition, requires interactions between local stro-mal cells and hematopoietic cells [Maestroni et al.,1994a,b]. Bone marrow stromal cells express opioidreceptors [Maestroni and Conti, 1996; Boranic etal., 1997] that can bind kappa-opioids with varyingdegrees of affinity [Maestroni, 1998]. Hematopoi-etic apoptosis protection has therefore been pro-posed to result from an indirect mechanism,whereby bone marrow stromal cells are induced tosecrete cell survival-promoting GM-CSF in re-sponse to novel kappa-opioids produced by mela-tonin-stimulated CD4+ T-helper cells. A similarchain of events could modify B-cell apoptosis inbone marrow by stimulating the production ofB-cell-supportive stromal cell-derived factors.
Precursor B cells in vivo form intimate associa-tions with bone marrow stromal cells, whichprovide molecular adhesion and short rangegrowth factors essential for B-cell development[Jacobsen and Osmond, 1990; Kincade, 1991; Ja-cobsen et al., 1992, 1996]. In particular, the stromal
cell-derived cytokine, IL-7, activates receptors ex-pressed on pro-B cells and large pre-B cells. Thisstimulates pre-B-cell proliferation and promotessurvival of pre-B cells by increasing their intracellu-lar Bcl-2/Bax protein ratio [Cumano et al., 1990;Valenzona et al., 1996, 1998; Lu et al., 1999].Overexpression of IL-7 in IL-7 transgenic miceinhibits apoptosis at the pre-B-cell stage of B-celldevelopment but has no such effect on sIgM+ Bcells [Lu et al., 1999], closely resembling our presentfindings in melatonin-treated mice. Possibly, likethe melatonin-induced myeloid protection medi-ated by GM-CSF, the melatonin-induced protec-tion of pre-B cells from apoptosis may be mediatedindirectly by IL-7 released from bone marrowstromal cells as a result of stimulation by kappa-opioids derived from the T cells that recirculatecontinuously between the blood stream and bonemarrow [Osmond, 1980].
The suppression of pre-B-cell apoptosis by mela-tonin treatment may have important physiologicaland pharmacological implications. Fluctuating cir-culatory levels of melatonin during the normallight/dark cycle could influence the survival frac-tion of early B-cell precursors and thus producediurnal fluctuations in the supply of new B cells tothe immune system. Melatonin treatment wouldtherefore be expected to boost the numbers ofnewly formed B cells. This could have a desirableshort-term effect, potentially promoting humoralimmunity. An ability of melatonin to enhancehumoral immunity, by indirect means, has beendescribed [Maestroni et al., 1986]. The presentfindings are consistent with the enhancement ofhumoral immunity, as reflected by IgG levels, pro-duced by housing mice under short-day conditionscompared with long-day-exposed mice; short daysresult in increased secretion of endogenous mela-tonin by the pineal gland [Demas and Nelson, 1996;Nelson and Demas, 1997]. Speculatively, however,excessive melatonin-induced suppression of pre-B-cell apoptosis could permit some genetically aber-rant B cells to evade the normal quality controlcheckpoint and to enter the peripheral lymphoidsystem as viable pre-neoplastic cells.
Acknowledgments
This work was supported by grants from the MedicalResearch Council of Canada and the National CancerInstitute of Canada with funds from the Cancer ResearchSociety, Inc.
Literature cited
BERTOLI, L.F., H. KUBAGAWA, G.V. BORZILLO, P.D. BUR-
ROWS, M.T. SCHREEDER, A.J. CARROLL, M.D. COOPER
91

Yu et al.
(1988) Bone marrow origin of a B cell lymphoma. Blood72:94–101.
BORANIC, M., L. KRIZANAC-BENGEZ, J. GABRILOVAC, T.MARROTTI, D. BRELJAK (1997) Enkephalins in hemato-poiesis. Biomed. Pharmacother. 51:29–37.
CALVO, J.R., M. RAFII-EL-IDRISSI, D. POZO, J.M. GUER-
RERO (1995) Immunomodulatory role of melatonin: specificbinding sites in human and rodent lymphoid cells. J. PinealRes. 18:119–126.
CHAMPNEY, T.H., J. PRADO, T. YOUNGBLOOD, K. APPEL,D.N. MCMURRAY (1997) Immune responsiveness of spleno-cytes after chronic daily melatonin administration in maleSyrian hamsters. Immunol. Lett. 58:95–100.
CHAMPNEY, T.H., G.C. ALLEN, M. ZANNELLI, L.A. BEAU-
SANG (1998) Time-dependent effects of melatonin on im-mune measurements in male Syrian hamsters. J Pineal Res.25:142–146.
CUMANO, A., K. DORSHKIND, S. GILLIS, C.J. PAIGE (1990)The influence of S17 stromal cells and interleukin-7 on B celldevelopment. Eur. J. Immunol. 20:2183–2189.
DEMAS, G.E., R.J. NELSON (1996) Photoperiod and tempera-ture interact to affect immune parameters in adult male deermice (Peromyscus maniculatus). J. Biol. Rhythms 11:94–102.
DEMAS, G.E., R.J. NELSON (1998) Exogenous melatonin en-hances cell-mediated, but not humoral, immune function inadult male deer mice (Peromyscus maniculatus). J. Biol.Rhythms 13:245–252.
FAUTEUX, L.J., D.G. OSMOND (1996) Interleukin-1 as a sys-temic stimulant of B lymphopoiesis: IL-1a binds to stromalcells and sinusoid endothelium in bone marrow and perturbsprecursor B cell dynamics. J. Immunol. 156:2376–2383.
GARCIA-MAURINO, S., M.G. GONZALEZ-HABA, J.R. CALVO,M. RAFII-EL-IDRISSI, V. SANCHEZ-MARGALET, R. GOB-
ERNA, J.M. GUERRERO (1997) Melatonin enhances IL-2,IL-6, and IFN-gamma production by human circulatingCD4+ cells: a possible nuclear receptor-mediated mecha-nism involving T helper type 1 lymphocytes and monocytes.J. Immunol. 15:574–581.
GONZALEZ-HABA, M.G., S. GARCIA-MAURINO, J.R. CALVO,R. GOBERNA, J.M. GUERRARO (1995) High-affinity bindingof melatonin by human circulating T lymphocytes (CD4+).FASEB J. 9:1331–1335.
GUERRERO, J.M., R.J. REITER (1992) A brief survey of pinealgland–immune system interrelationships. Endocr. Res.18:91–113.
HARDELAND, R., R.J. REITER, B. POEGGLER, D.X. TAN
(1993) The significance of the metabolism of the neurohor-mone melatonin: Antioxidative protection and formation ofbioactive substances. Neurosci. Behav. Rev. 17:347–357.
HAYASHI, S., T. KUNISADA, M. OGAWA, T. SUDO, H. DO-
DAMA, T. SUDA, S. NISHIKAWA, S. NISHIKAWA (1990) Step-wise progression of B lineage differentiation supported byinterleukin-7 and other stromal cell molecules. J. Exp. Med.171:1683–1695.
HENNEY, C.S. (1989) Interleukin-7: Effects on early events onlymphopoiesis. Immunol. Today 10:170–173.
INSERRA, P., Z. ZHANG, S.K. ARDESTANI, M. ARAGHI-NIKNAM, B. LIANG, S. JIANG, D. SHAW, M. MOLITOR, K.ELLIOT, R.R. WATSON (1998) Modulation of cytokine pro-duction by dehydroepiandrosterone (DHEA) plus melatonin(MLT) supplementation of old mice. Proc. Soc. Exp. Biol.Med. 218(1):76–82.
JACOBSEN, K., D.G. OSMOND (1990) Microenvironmental or-ganisation and stromal cell associations of B lymphocyteprecursor cells in mouse bone marrow. Eur. J. Immunol.20:2395–2404.
JACOBSEN, K., D. MIYAKE, P.W. KINCADE, D.G. OSMOND
(1992) Highly restricted expression of a stromal cell determi-nant in mouse bone marrow in vivo. J. Exp. Med. 176:927–935.
JACOBSEN, K.A., J. KRAVITZ, P.W. KINCADE, D.G. OSMOND
(1996) Adhesion receptors on bone marrow stromal cells: Invivo expression of vascular cell adhesion molecule-1 by retic-ular cells and sinusoidal endothelium in normal and g-irradi-ated mice. Blood 87:73–82.
KARASUYAMA, H., A. ROLINK, Y. SHIKAI, F. YOUNG, F.W.ALT, F. MELCHERS (1994) The expression of VpreB/l5surrogate light chain in early bone marrow precursor B cellsof normal and B-cell deficient mutant mice. Cell 77:133–143.
KINCADE, P.W. (1991) Molecular interactions between stro-mal cells and B lymphocyte precursors. Semin. Immunol.3:379–390.
KINCADE, P.W. (1994) B lymphopoiesis: Global factors, localcontrol. Proc. Natl. Acad. Sci. USA 91:2888–2889.
LIEBMANN, P.M., A. WOLFLER, P. FELSNER, D. HOFER, K.SCHAUENSTEIN (1997) Melatonin and the immune system.Int. Arch. Allergy Immunol. 112:203–211.
LU, L., D.G. OSMOND (1997) Apoptosis during B lympho-poiesis in mouse bone marrow. J. Immunol 158:5136–5145.
LU, L., P. CHAUDHURY, D.G. OSMOND (1999) Regulation ofcell survival during B lymphopoiesis: Apoptosis and Bcl-2/Bax content of precursor B cells in bone marrow of micewith altered expression of IL-7 and recombinase-activatinggene-2. J. Immunol. 162:1931–1940.
MAESTRONI, G.J. (1998) Kappa-opioid receptors in marrowstroma mediate the hematopoietic effects of melatonin-in-duced opioid cytokines. Ann. N.Y. Acad. Sci. 840:411–419.
MAESTRONI, G.J., A. CONTI (1996) Melatonin and the im-mune-hematopoietic system: Therapeutic and adverse phar-macological correlates. Neuroimmunomodulation3:325–332.
MAESTRONI, G.J., A. CONTI, W. PIERPAOLI (1986) Role ofthe pineal gland in immunity. Circadian synthesis and releaseof melatonin modulates the antibody response and antago-nizes the immunosuppressive effect of corticosterone. J. Neu-roimmunol. 13:19–30.
MAESTRONI, G.J., V. COVACCI, A. CONTI (1994a) Hemato-poietic rescue via T-cell-dependent, endogenous granulocyte-macrophage colony-stimulating factor induced by the pinealneurohormone melatonin in tumor-bearing mice. CancerRes. 54:2429–2432.
MAESTRONI, G.J., A. CONTI, P. LISSONI (1994b) Colony-stim-ulating activity and hematopoietic rescue from cancerchemotherapy compounds are induced by melatonin via en-dogenous interleukin-4. Cancer Res. 54:4740–4743.
MAESTRONI, G.J., E. HERTENS, P. GALLI, A. CONTI, E.PEDRINIS (1996) Melatonin-induced T-helper cell hemato-poietic cytokines resembling both interleukin-4 and dynor-phin. J. Pineal Res. 21:131–139.
MAYO, J.C., R.M. SAINZ, H. URIA, I. ANTOLIN, M.M. ESTE-
BAN (1998) Inhibition of cell proliferation: a mechanismlikely to mediate the prevention of neuronal cell death bymelatonin. J. Pineal Res. 25:12–18.
MAZZOCCOLI, G., M. CORRERA, G. BIANCO, A. DE CATA,M. BALZAMELLI, A. GIULIANI, R. TARGUINI (1997) Age-re-lated changes of neuro-endocrine-immune interactions inhealthy humans. J. Biol. Regul. Homeost. Agents 11:143–147.
MEDINA, K.L., P.W. KINCADE (1994) Pregnancy-relatedsteroids are potential negative regulators of B lymphopoiesis.Proc. Natl. Acad. Sci. USA 91:5382–5386.
MELCHERS, F., A. ROLINK, U. GRAWUNDER, T.H. WIN-
KLER, H. KARASUYAMA, P. GHIA, J. ANDERSSON (1995)
92

Melatonin inhibits apoptosis of pre-B cells
Positive and negative selection events during B lympho-poiesis. Curr. Opin. Immunol. 7:214–227.
NELSON, R.J., G.E. DEMAS (1996) Seasonal changes in im-mune function. Q. Rev. Biol. 71:511–548.
NELSON, R.J., G.E. DEMAS (1997) Role of melatonin inmediating seasonal energetic and immunologic adaptations.Brain Res. Bull. 44:423–430.
OETTINGER, M.A., D.G. SCHATZ, C. GORKA, D. BALTIMORE
(1990) RAG-1 and RAG-2, adjacent genes that synergisti-cally activate V(D)J recombination. Science 248:1517–1523.
OPSTELTEN, D., D.G. OSMOND (1983) Pre-B cells in mousebone marrow: Immunofluorescence stathmokinetic studies ofproliferation of cytoplasmic m-chain-bearing cells in normalmice. J. Immunol. 131:2635–2640.
OSMOND, D.G. (1980) The contribution of bone marrow tothe economy of the lymphoid system. Monogr. Allergy16:157–172.
OSMOND, D.G. (1990) B cell development in the bone mar-row. Semin. Immunol. 2:173–180.
OSMOND, D.G., S. RICO-VARGAS, H. VALENZONA, L. FAU-
TEUX, L. LIU, R. JANANI, L. LU, K. JACOBSEN (1994)Apoptosis and macrophage-mediated cell deletion in theregulation of B lymphopoiesis in mouse bone marrow. Im-munol. Rev. 142:209–230.
OSMOND, D.G., A. ROLINK, F. MELCHERS (1998) Murine Blymphopoiesis: Towards a unified model. Immunol. Today19:65–68.
POZO, D., M. DELGADO, J.M. FERNANDEZ-SANTOS, J.R.CALVO, R.P. GOMARIZ, I. MARTIN-LACAVE, G.G. ORTIZ,J.M. GUERRERO (1997) Expression of the Mel1a-melatoninreceptor mRNA in T and B subsets of lymphocytes from ratthymus and spleen. FASEB J. 11:466–473.
PROVINCIALI, M., G. DI STEFANO, D. BULIAN, A. TIBALDI,N. FABRIS (1996) Effect of melatonin and pineal grafting onthymocyte apoptosis in aging mice. Mech. Aging Dev. 90:1–19.
RAJEWSKY, K. (1996) Clonal selection and learning in theantibody system. Nature 381:751–758.
ROLINK, A., F. MELCHERS (1993) Generation and regenera-tion of cells of the B lymphocyte lineage. Curr. Opin. Im-munol. 5:207–217.
SAINZ, R.M., J.C. MAYO, H. URIA, M. KOTLER, I. ANTOLIN,C. RODRIGUEZ, A. MENENDEZ-PELAEZ (1995) The pinealneurohormone melatonin prevents in vivo and in vitro apop-tosis in thymocytes. J. Pineal Res. 19:178–188.
TEN BOEKEL, E., F. MELCHERS, A.G. ROLINK (1997)Changes in the VH gene repertoire of developing precursor Blymphocytes in mouse bone marrow mediated by the pre-Bcell receptor. Immunity 7:357–368.
TSUJIMOTO, Y., J. GORHAM, J. COSSMAN, E. JAFFEE, C.M.CROCE (1985) The t(14;18) chromosome translocations in-volved in B cell neoplasms result from mistakes in VDJjoining. Science 229:1390–1393.
VALENZONA, H.O., R. POINTER, R. CEREDIG, D.G. OSMOND
(1996) Prelymphomatous B cell hyperplasia in bone marrowof interleukin-7 transgenic mice: precursor B cell dynamics,microenvironmental organization and osteolysis. Exp. He-matol. 24:1522–1530.
VALENZONA, H.O., S. DHANOA, F.D. FINKELMAN, A. NA-
MEN, D.G. OSMOND (1998) Exogenous IL-7 as a prolifera-tive stimulant of early precursor B cells in bone marrow:Comparative efficacy of IL-7 injection, IL-7 infusion andIL-7 complexes. Cytokine 10:404–412.
93