mecanismos de regulación post transcripcional
TRANSCRIPT

Universidad Central del EcuadorFacultad de Medicina
Biología Molecular

El ARN mensajero obtenido después de la transcripción se
conoce como transcrito primarioo ARN precursor (pre-ARN), que en la mayoría de los casos no se
libera del complejo de transcripción en forma totalmente
activa, sino que ha de sufrir modificaciones antes de ejercer su
función (procesamiento o maduración del ARN).

Maduración
Procariotas: Si el ARN formado es ARNm, no hay maduración; si se trata de ARNr o ARNt, hay un transcrito primario que sufre un proceso de 'cortes y empalmes.
Eucariotas: Maduración se produce en el núcleo y la hace un enzima llamada RNPpn, que elimina los nuevos intrones (I) formados.



Comienza a modificarse poco después de la trascripción
Se trata de una modificación covalente que implica 3 tipos de enzimas
Las enzimas son la fosfatasa, guanililtranferasa, y la metilasa
Esta modificación conduce a la incorporación singular de un
nucleótido protector sobre el NPT en la posición 5’ terminal del ARN
Este nucleótido es normalmente purínico,
ATP, el mismo que actuó como cebador en la transcripción


Se da en 2 fasesa) Una desfosforilación previab) guanililación a costa del GTP

En la segunda reacción se transfiere un grupo guanililo
El conjunto de estas 2 reacciones es la incorporación al ARNm de tan solo un
residuo de guanosina
La estructura terminal resultante se conoce como
caperuza de guanina o caperuza 5’
Esta caperuza tiene una estructura singular que no se la encuentra en
ningún otro nucleótido
Además ejerce un efecto protector de la molécula frente a las exonucleasas, marca el pre ARNm para su empleo como
sustrato en otras reacciones de procesamiento del núcleo y sirve como punto de union del ARNm a los ribosomas al
iniciar la síntesis proteica


Los 3 primeros nucleótidos de la 5’ son modificados por las
metiltranferasas
Estas enzimas emplean el S- adenosil – L - metionina como coenzima donadora
del grupo metilo
La estructura terminal resultante se conoce como
caperuza de guanina o caperuza 5’
En el 2’- OH de la ribosa adyacente forma la
caperuza 1 la reacción la cataliza la ARNm
(nucleosido -2’-O)metiltranferasa
En el n7 de la guanosina terminal para formar la caperuza 0, la reacción
la cataliza la ARNm (guanina – N7-
)metiltranferasa
En el 2’ OH de la ribosa siguiente reacción
forma la caperuza 2 la cataliza la ARNm (nucleosido -2’-
O)metiltranferasa
Esta metilacion puede tener lugar sobre varias posiciones



0Otra peculiaridad del ARNm eucariotico es que el extremo 3’ del ARNm no corresponde a la posición donde se termina la transcripción si no que deriva de la molécula de pre – ARNm o hetero ARN



Sobre el extremo 3’-OH resultante del corte por
endonucleasa actúa una ARN polimerasa especial que no usa molde y acepta como sustrato
solo al ATP POLI (A) POLIMERASA
Como consecuencia de esto se añade al ARN un gran número de residuos de
adelinato formando la cola de poliadenilato o cola poli (A)
Esta estructura que permanece intacta hasta la formación final
del ARN
Participa en procesos como en el transporte del ARNm al citoplasma y la determinación y la estabilidad de la
vida media del ARNm

Poliadenilación

Poliadenilación del extremo 3’
• Sobre el extremo 3’ OH delcorte por la endonucleasaactúa, poli (A) polimerasa,que no usa molde y usa comosustrato al ATP.
• Como consecuencia, se añadeal ARNm un gran número deresiduos de adenilato (AMP),tomando una “cola” depoliadenilato o poli (A), delongitud típica entre 40 y 250nt (como excepción los ARNmde histonas carecen de estacola).
• Esta estructura permaneceintacta hasta la formaciónfinal de ARNm
• Puede participar enprocesos de tan gransignificado como eltransporte de ARNm alcitoplasma y ladeterminación, en parte,de la estabilidad y vidamedia del ARNm

SITIOS DE POLIADENILACIÓN ALTERNATIVA
0 Puede ocurrir algo similar a la ocurrencia de sitios alternativos de inicio de la transcripción pero en el extremo 3’.
0 En este caso hallándose sobre un mismo gen varios sitios susceptibles de ser poliadenilados como es el caso del gen para la cadena pesada m de inmunoglobulina.
0Dependiendo del sitio 3’ poliadenilado, la proteína resultante puede anclarse a membrana o ser secretada, de acuerdo al estadío de desarrollo en que se encuentra la célula.

Eliminación de intrones y empalme de exones
(SPLICING)

• La etapa inicial de la maduración postranscripcionaldel transcrito primario hasta dar el ARNm eucarióticoconsiste en la eliminación de los intrones y la unión oempalme de los exones.
• En el núcleo de las células eucariotas existenpartículas ribonucleoproteicas, los espliceosomas,formados por ARN pequeño nuclear (ARNsn =”smallnuclear”) y proteínas, cuyas moléculas de ARNcatalizan el corte de intrones y el empalme de exones.

Magnitud de los intrones
• Lo mas frecuente es que la longitud total de losintrones supere ampliamente a la suma de los exones,por lo que el tamaño del gen depende fuertemente delnúmero y longitud de sus intrones.
• Aunque las cifras son variables, se habla de hasta 40intrones por gen y de una longitud de hasta 20 kb porintron. Los exones son de tamaño mucho máspequeño, típicamente inferior a 1 kb.

Secuencias que delimitan el intrón
• La eliminación de un intrón está determinada por susecuencia, y no por su tamaño.
• Solo intervienen 2 secuencias específicas, que definenlos extremos del intrón, y otra de su interior,conocidas en su conjunto como sitios o centros decorte y empalme o de splicing.
• Todos los sitios de corte y empalme parecen serequivalentes, mientras que la secuencia del resto delintrón no tiene trascendencia sobre el proceso.

SPLICING ALTERNATIVO
0Es un mecanismo muy difundido, que permite que un solo gen pueda codificar para más de una proteína.
0 Los cortes sobre el RNAhn deben producirse con absoluta precisión
0 Puede ocurrir que uno más exones sean removidos (dando una proteína más corta), o que uno o más intrones no sean removidos (dando una proteína más larga).
0 Una familia de 6 proteínas llamadas ASF (factores de splicing alternativo) responsables de reconocer y seleccionar los lugares para los cortes alternativos, contienen serina y arginina por lo que se las llama también proteinasSR.

Secuencias que delimitan el intrón
• Un cambio en el sitio de corte (por ejemplo, pormutación de uno de los nucleótidos consensoesenciales) daría lugar a un ajuste erróneo: seutilizaría para el corte y empalme la siguientesecuencia consenso, si la hubiera, con lo que el intróno parte de él quedaría incluido en el ARNm.
• Esto conduciría a la síntesis de un polipéptidoalterado, con una inserción grande en su secuencia deaminoácidos, o incluso con cambio de tosa lasecuencia a partir del punto de alteración debido a undesplazamiento del marco de lectura.

0 Posteriormente las ARN ligasas empalman los exones (E) y forman el ARNm

Maduración ARNr y ARNtPROCARIOTAS
Transcrito primario
Reacciones de corte (RNAsas)
-3 RNAr
-1 o varios RNAt
1.Adición de grupos metilos
2.Modificación de nucleótidos.

Post-Transcripción
Silenciado de genes asociado al ARN

Vías de silenciado asociado al ARN
Interfiere un elemento básico (ARN de interferecnia que es un ARN báscio de unos 21-27 nucleótidos:
Se distinguen:
Los micro ARNs (miRNAs)
Los RNA interferentes pequeños (siRNAs)
Los RNA interferentes asociados a repeticiones (rasiRNAs)

Micro RNAs (miRNAs)0 Son transcitos por la RNA Polimerasa II como mirARNas
primarios (pri-mirARNas).

Micro RNAs humanos

miRNAs y siRNAs
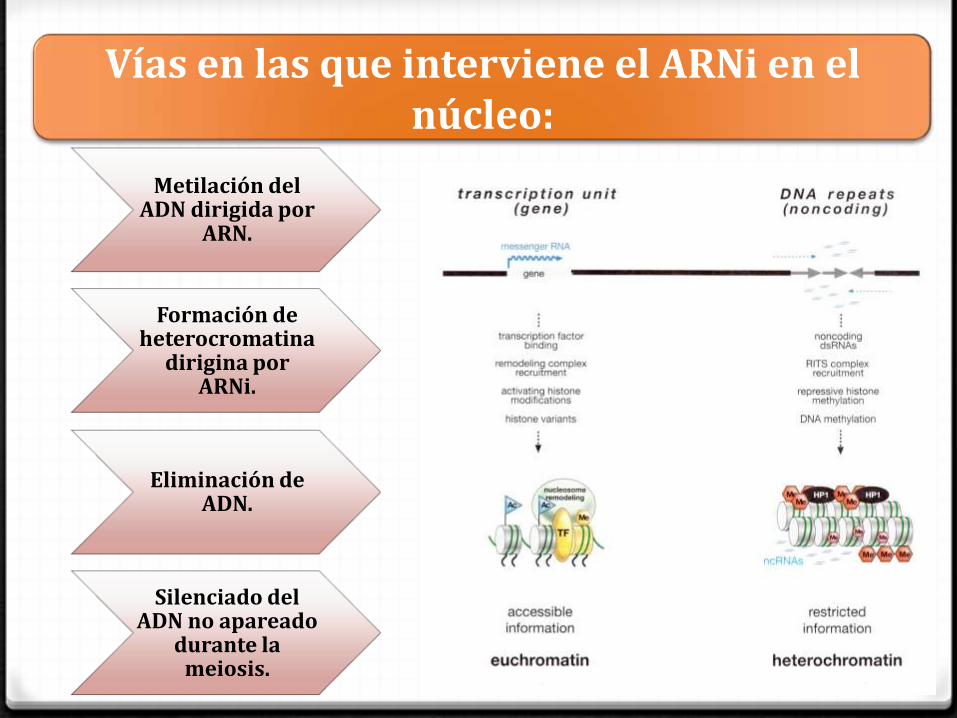
Vías en las que interviene el ARNi en el núcleo:
Metilación del ADN dirigida por
ARN.
Formación de heterocromatina
dirigina por ARNi.
Eliminación de ADN.
Silenciado del ADN no apareado
durante la meiosis.