measurement and characteristics of …jcs.biologists.org/content/joces/45/1/169.full.pdfmeasurement...
TRANSCRIPT

J. Cell Sci. 45, 169-186 (1980)
Printed in Great Britain © Company of Biologists Limited 1080
MEASUREMENT AND CHARACTERISTICS OF
FUSION OF ISOLATED MEMBRANE
FRACTIONS FROM MAIZE ROOT TIPS
ELIAS A-H. BAYDOUN AND D. H. NORTHCOTEDepartment of Biochemistry, University of Cambridge, Cambridge CBz iQW, England.
SUMMARY
Maize root tips were incubated in vivo with radioactive glucose, choline or diazotized8ulphanilic acid. Membrane fractions were prepared from radioactive and non-radioactiveroots. The transfer of radioactivity between mixed membrane fractions has enabled a quanti-tative system to be developed to study in vitro membrane fusion between Golgi apparatus andplasma membrane-rich fractions. Membrane fusion was found to be dependent on time,temperature, Mn1+ and Ca1+. Mn*+ was as effective as Ca'+, other divalent cations had amoderate or no effect. The effect of various substances, including inhibitors of microtubularand microfilament assembly and blocking reagents for sulphydryl group on membrane fusionhas been investigated. The process appears to be dependent on the membrane proteins orglycoproteins; trypsinization of mixed membranes prior to the addition of Ca1+ inhibitedsignificantly the fusion process. SDS-polyacrylamide gel electrophoresis of trypsin-treatedmembranes revealed the selective loss of one particular polypeptide which could play a role inmembrane fusion.
INTRODUCTION
Morris & Northcote (1977) showed that the secretion of substances from sycamoresuspension cells was controlled not only by the rate of their synthesis, but also bythe membrane fusion responsible for their export from the cell. Membrane fusion is amajor event in a large number of cellular and subcellular activities (Roberts & North-cote, 1970, 1971; Poste & Allison, 1973; Satir, Schooley & Satir, 1973; Plattner, 1974;Satir, 1974; Berger, Dahl & Meissner, 1975; Steinman, Brodie & Cohn, 1976; Allen &Wolf, 1979; Horwitz, Wight & Knudsen, 1979). Membrane fusion is regarded as oneof the processes which maintains the dynamic state of the endomembrane system ineukaryotic cells (Northcote, 1979).
The fluid mosaic model of the structure of biological membranes (Singer &Nicolson, 1972), has contributed a great deal to the establishment of several schemesdescribing the ultrastructural events taking place during membrane fusion. Theseschemes are mainly based on freeze-fracture and thin-section electron microscopy ofseveral types of cells active in secretion (Satir et al. 1973; Burwen & Satir, 1977;Da Silva & Nogueira, 1977; Orci, Perrelet & Friend, 1977; Kalderon & Gilula,
1979)-Several agents have been used experimentally to initiate fusion, including Sendai
Please address all correspondence to Professor D. H. Northcote at the above address.12 CEL 45

170 E. A-H. Baydoun and D. H. Northcote
virus (Knutton, 1977), isoproterenol (De Camilli, Peluchetti & Meldolesi, 1976), poly-ethylene glycol (Kao & Michayluk, 1974; Svoboda, 1978; Knutton, 1979), glycerol,fatty acids, and lysolecithin (Ahkong, Fisher, Tampion & Lucy, 1973, 1975) andn-alkyl bromides (Manson, Hladky & Haydon, 1979). But Ca2+ is considered as themost important fusogenic reagent and it has been studied in detail with both biological(Dahl & Gratzl, 1976; Gratzl & Dahl, 1976; Milutinovic, Argent, Schulz & Sachs,1977; Volsky & Loyter, 1977; Gratzl & Dahl, 1978) and artificial membranes(Papahadjopoulos, Poste, Schaeffer & Vail, 1974; Breisblatt & Ohki, 1976; Koter,De Kruijff & Van Deenen, 1978; Liao & Prestegard, 1979).
Although roles for phospholipids (Ahkong et al. 1975; Lemay, Collyn-D'Hooghe &Torpier, 1977), proteins and glycoproteins (Gratzl & Dahl, 1978; Dahl, Schudt &Gratzl, 1978) in membrane fusion have been proposed, the molecular mechanism ofthe process is not well understood. Most of the information on the molecular mecha-nism of membrane fusion comes from investigations using artificial phospholipidbilayer vesicles (Papahadjopoulos, Vail, Pangborn & Poste, 1976; Koter et al. 1978;Liao & Prestegard, 1979; Portis, Newton, Pangborn & Papahadjopoulos, 1979;Puskin & Martin, 1979). The results obtained from these investigations are difficultto relate to the in vivo condition, because the role of proteins, a major component ofbiological membranes, is excluded.
The aim of our work was to develop a quantitative system to measure membranefusion in vitro. For this purpose several radioactive probes were used to label membranefractions of maize root tips in vivo. The incorporation and transfer of this radioactivityenabled a study of the in vitro fusion between mixed Golgi apparatus- and plasmamembrane-rich fractions to be made. The effect of various additives on the transferof radioactivity between mixed membrane fractions has been investigated.
MATERIALS AND METHODS
Acid phosphatase (Grade 1; specific activity at 25 °C, pH 5-6 = 60 U./mg enzyme protein)was obtained from Boehringer, Mannheim, FRG, dialysed to remove ammonium sulphate,and freeze-dried. The enzyme was assayed spectrophotometrically at 405 nm by following thehydrolysis of £-nitrophenyl phosphate (A 185 mM"1 cm"1) at 30 °C in the presence of 10 mMTris-HCl, pH 7-4, o-i mM MgCla, 1 mM EDTA and 34 % (w/w) sucrose solution. Under theseconditions the specific activity was 15-5 U./mg enzyme protein. Recovery of activity afterdialysis and freeze drying was 80 %. The enzyme showed no phosphodiesterase activity in theassay of Koerner & Sinsheimer (1957).
Trypsin (sp. act. 33 U./mg protein) was obtained from. Boehringer Mannheim, FRG.Trypsin inhibitor from soybean was obtained from Sigma Chemical Co., St Louis, Mo., USA.Trypsin was assayed spectrophotometrically at 253 nm by following the hydrolysis of N-benzoyl-L-arginine ethyl ester at 20 °C in the buffer described for acid phosphatase. Underthese conditions trypsin (32/ig ml"1) caused an increase in Ali3 of 0-35 min"1. The activitywas completely and instantaneously abolished by the inhibitor at all ratios tested (1:1, 1:1-5,1:2, (w/w), trypsin: trypsin inhibitor).
Growth of maize seedlings, incubation of roots with radioactive precursors in vivo, homo-genization, membrane fractionation, determination of radioactivity and SDS-polyacrylamidegel electrophoresis were carried out as previously described (Baydoun & Northcote, 1980).

Measurement of membrane fusion in vitro 171
Transfer of radioactivity between membrane fractions mixed in vitro
Membrane fractions were prepared from radioactive and non-radioactive roots. RadioactiveGolgi apparatus-rich fraction (05 ml) was mixed with an equal volume of non-radioactiveplasma membrane-rich fraction at 30 °C for 40 min in the presence of 1 mM Cat+ (final appa-rent free concentration) in a final volume of 104 ml. EDTA was then added to 6 mil, Themixed membranes were cooled on ice and the volume was made up to 2-5 ml by addition of10 mM of a solution of Tris-HCl, pH 7'4- They were then immediately refractionated bycentrifugation at IOOOOOJ* and 4 CC for 90 min in a discontinuous sucrose density gradientprepared in a 5'5-ml cellulose nitrate tube by layering in succession 45 % (w/w), 0-5 ml, 34 %(w/w), 1-25 ml, and 25 % (w/w), 1 ml, sucrose solutions. After recentrifugation, bands at the25~34 % interface (Golgi apparatus-rich), 34-45 % interface (plasma membrane-rich) and allthe material left in the tube (remainder fraction) were collected and pelleted (iooooog,4 °C, 30 min) in the presence of non-radioactive homogenate as carrier. The pellets weresuspended in 18 % (w/v) trichloroacetic acid, washed and assayed for radioactivity.
The effect of various additions on the transfer of radioactivity between mixed membranefractions was investigated. Tris-HCl (pH 7-4, 10 mM) was used as solvent for all the additivesexcept where otherwise stated.
RESULTS
Transfer of radioactivity between mixed membrane fractions using various radioactiveprecursors
Roots were incubated in vivo with D-[U-14C]glucose, diazotized [U-3H]sulphanilicacid, or [Me-14C]choline chloride (Baydoun & Northcote, 1980). Radioactive mem-brane fractions, either alone or mixed with non-radioactive membrane fractions,were maintained at 30 °C for 40 min in the presence of 1 mM Ca2+. Tables r-3 showthat a similar percentage of the radioactivity was transferred between mixed membranefractions with all 3 radioactive probes. Therefore, it seems likely that all the majorcomponents of the membrane are participating in the transfer of radioactivity. Inpreliminary experiments mixed membrane fractions were layered on top of adiscontinuous gradient prepared in a 5-5-1111 cellulose nitrate tube by layering insuccession 45 % (w/w), 0-5 ml, and 1 ml each of 39, 34 and 25 % (w/w) sucrosesolutions (Table 1). Low amounts of radioactivity were recovered on the 39-45%interface, especially when radioactive Golgi apparatus-rich fraction was mixed withnon-radioactive plasma membrane-rich fraction; thus, the 39% step was omittedand recentrifugation was carried out in gradients made up as specified underMaterials and methods.
Time course of the transfer of radioactivity between mixed membrane fractions and effectofCa*+
Radioactive Golgi apparatus-rich fraction was mixed with non-radioactive plasmamembrane-rich fraction and maintained at 30 °C. Table 4 shows that in the absenceof Ca2+ no transfer of radioactivity occurred between membrane fractions over aperiod of 45 min. In another experiment, when Ca2+ was present at 1 mM (Table 5),radioactivity was transferred to the plasma membrane-rich fraction. The plasmamembrane-rich fraction accounted for 34% of the total radioactivity after 10 min.The amount of radioactivity transferred approached a maximum after 40 min.

172 E. A-H. Baydoun and D. H. Northcote
Effect of temperature on the transfer of radioactivity between mixed membrane fractions
Radioactive Golgi apparatus-rich fraction was mixed with non-radioactive plasmamembrane-rich fraction and maintained for 40 min in the presence of o-i mM Ca2+
at various temperatures. A marked increase in the transfer of radioactivity was observedas the temperature was increased (Table 6). This increase probably resulted from theeffect of temperature on membrane fluidity. The radioactivity found in the plasmamembrane-rich fraction at o °C was not in excess of that found after mixing and
Table 1. Transfer of radioactivity between membrane fractions one of which was non-radioactive and the other made radioactive with D-[U-14C]glucose
FractionGolgi apparatus-rich (GA)Plasma membrane-rich (PM)Mitochondria-richRemainder fractionTotal cpm obtained from
all the fractionsRecovery*
Radioactivity (%) fromrefractionated membranes after mixing
A
GA* with PM°
47-9494
0 9i -8
14950
90-0
GA* alone
89-18-30 9
1-7
13650
83 0
GA° with PM*
3 2 8
47-81 0 7
8-77180
948
PM* alone
4 0
75-1I2-O
8-96350
83-8
• Recovery (%) was calculated by comparing total cpm with the initial cpm obtained bypelleting a radioactive fraction.Roots were incubated with D-[U-14C]glucose iw vivo. Membrane fractions were prepared
from radioactive roots (•) and from non-radioactive roots (°). These were mixed at 30 °C for40 min in the presence of 1 mM Ca*+ and then refractionated by centrifugation. The distribu-tion of radioactivity in the fractions was determined.
Table 2. Transfer of radioactivity between membrane fractions one of which was non-radioactive and the other made radioactive with diazotized [U-*H]sulphaniIic acid
FractionGolgi apparatus-richPlasma membrane-richRemainder fractionTotal cpm obtained fromall the fractions
Recovery
Radioactivity (%) fromrefractionated membranes after mixing
GA* with PM°
5 5 142-1
2-82730
93-8
GA* alone
88-68-i
3'32660
9 1 2
GA° with PM*
40-856-7
2-5397°
999
PM* alone
2-9
93 43 7
3780
95-iRoots were incubated with diazotized [U-'H]sulphanilic acid in vivo. Membrane fractions
were prepared from radioactive roots (•) and from non-radioactive roots (°). These weremixed at 30 °C for 40 min in the presence of 1 mM Ca*+ and then refractionated by centrifuga-tion. The distribution of radioactivity in the fractions was determined.

Measurement of membrane fusion in vitro 173
immediate refractionation of the membranes (Table 5) or when radioactive Golgiapparatus-rich fraction was recentrifuged alone (Tables 1-3).
Effect of cations on the transfer of radioactivity between mixed membrane fractions
It was assumed in this work that the endogenous concentration of the divalentcations was negligible. Since the homogenization and fractionation media containedabout 0-9 mM excess EDTA (i.e. o-i mM MgCl2 and i-o mM EDTA), the solutionsused in this study contained 0-9 mM of the divalent cation in excess of the concentrationstated. Thus, cation concentrations are expressed as final apparent free concentration.
Radioactive Golgi apparatus-rich fraction was mixed with non-radioactive plasma
Table 3. Transfer of radioactivity between membrane fractions one of which was non-radioactive and the other made radioactive with [Me-^C^holine chloride
FractionGolgi apparatus-richPlasma membrane-richRemainder fractionTotal cpm obtained from
all the fractionsRecovery
GA# with
48-146-3
5 *18880
9 0 0
Radioactivity (%) fromrefractionated membranes after mixing
A
PM° GA# alone
83 01 0 6
6 418700
9 8 1
GA° with PM»
3 2 853-31 3 9
8520
978
PM* alone
6 78 3 1IO-2
8lSO
9 3 5
Roots were incubated with [Me-14C]choline chloride in vivo. Membrane fractions wereprepared from radioactive roots (•) and from non-radioactive roots (°). These were mixed at30 °C for 40 min in the presence of 1 mM Ca'+ and then refractionated by centrifugation. Thedistribution of radioactivity in the fractions was determined.
Table 4. Effect of time on the transfer of radioactivity between mixed membrane fractionsin the absence of Ca?+
Tim*1 nfmixing, min. . .
FractionGolgi apparatus-richPlasma membrane-richRemainder fractionTotal cpm obtained fromall the fractions
Recovery
o
88-88 0
3 210650
92-1
Radioactivity (%) fromrefractionated membranes after
IS
88-47 73 9
10480
90-6
A
3O
88-o8 04 0
10940
946
mixing
45
88-57-83 7
10560
0 1 3
Roots were incubated with D-[U-uC]glucose in vivo. Golgi apparatus-rich fraction wasprepared from radioactive roots. Plasma membrane-rich fraction was prepared from non-radioactive roots. These were mixed at 30 °C, maintained for various periods of time and thenrefractionated by centrifugation. The distribution of radioactivity in the fractions was deter-mined.

174 E. A-H. Baydoun and D. H. Northcote
membrane-rich fraction and maintained at 30 °C for 40 min in the presence of various
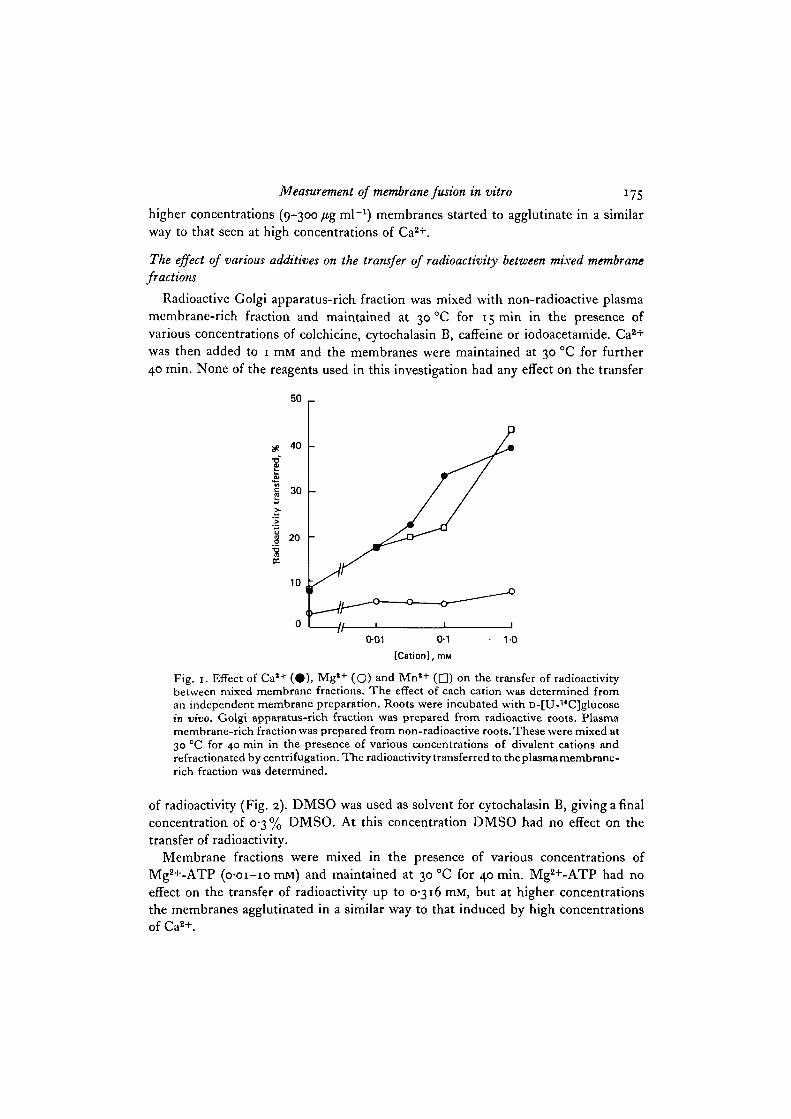
concentrations of Ca2+, Mg2+ and Mn2+ (Table 7 and Fig. 1). The transfer of radio-
activity was dependent on the concentration of Ca2+ or Mn2+ present in the mixing
medium. Mg2+ at the same concentrations showed little or no effect. Ba2+ and Sr2+
Table 5. Effect of time on the transfer of radioactivity between mixed membrane fractions
in the presence of Ca2+
Time of mixing, min.
FractionGolgi apparatus-richPlasma membrane-richRemainder fractionTotal cpm obtained from
all the fractionsRecovery
0
848io-o5-2
7600
9 1 8
Radioactivity (%) fromrefractionated membranes after
1 0
6 1 73 4 °
4'3734O
88-6
2 0
5 6 1
4 ° 53'4
7810
9 4 3
30
4 9 0476
3'47790
9 4 0
mixing
4 0
45'45 0 1
4'57480
9 0 3
Roots were incubated with D-[U-14C]glucose in vivo. Golgi apparatus-rich fraction wasprepared from radioactive roots. Plasma membrane-rich fraction was prepared from non-radioactive roots. These were mixed at 30 °C in the presence of 1 mM Ca1+, maintained forvarious periods of time and then refractionated by centrifugation. The distribution of radio-activity in the fractions was determined.
Table 6. Effect of temperature on the transfer of radioactivity between mixed membrane
fractions
Radioactivity (%) fromrefractionated membranes after mixing
Temperatureof mixing, °C. . .
FractionGolgi apparatus-richPlasma membrane-richRemainder fractionTotal cpm obtained fromall the fractions
Recovery
0
83-5I I - I
5'47230
8 7 3
1 0
8 1 9
1 4 53-6
7000
84-5
A
2 0
75-22 0 4
4'47290
8 8 0
3 0
6 6 23 1 0
2 86810
82-2
4 0
48-347-1
4 67230
87-3
Roots were incubated with D-[U-14C]glucose in vivo. Golgi apparatus-rich fraction was pre-pared from radioactive roots. Plasma membrane-rich fraction was prepared from non-radio-active roots. These were mixed for 40 min in the presence of o-i mM Ca1+, maintained atdifferent temperatures and refractionated by centrifugation. The distribution of radioactivityin the fractions was determined.
at i-omM showed a moderate effect (Table 8). When Ca2+ was used at or above
3 mM the membranes agglutinated and were isolated as a single band on the gradient,
precluding any refractionation even in the presence of high concentrations of EDTA.
La3+ caused membrane agglutination even at very low concentrations (o-oi-
i-o mM). The polyvalent cation poly-L-lysine had no effect up to 3 fig ml"1, but at
Measurement of membrane fusion in vitro 175
higher concentrations (9-300 /tg ml"1) membranes started to agglutinate in a similarway to that seen at high concentrations of Ca2+.
The effect of various additives on the transfer of radioactivity between mixed membranefractions
Radioactive Golgi apparatus-rich fraction was mixed with non-radioactive plasmamembrane-rich fraction and maintained at 30 °C for 15 mm in the presence ofvarious concentrations of colchicine, cytochalasin B, caffeine or iodoacetamide. Ca2+
was then added to 1 rail and the membranes were maintained at 30 °C for further40 min. None of the reagents used in this investigation had any effect on the transfer
50 _
0-01 0-1
[Cation], mM
Fig. 1. Effect of Cal+ ( • ) , Mg*+ (O) and Mn'+ ( • ) on the transfer of radioactivitybetween mixed membrane fractions. The effect of each cation was determined froman independent membrane preparation. Roots were incubated with D-[U-14C]glucosein vivo. Golgi apparatus-rich fraction was prepared from radioactive roots. Plasmamembrane-rich fraction was prepared from non-radioactive roots. These were mixed at30 °C for 40 min in the presence of various concentrations of divalent cations andrefractionated by centrifugation. The radioactivity transferred to the plasma membrane-rich fraction was determined.
of radioactivity (Fig. 2). DMSO was used as solvent for cytochalasin B, giving a finalconcentration of 0-3 % DMSO. At this concentration DMSO had no effect on thetransfer of radioactivity.
Membrane fractions were mixed in the presence of various concentrations ofMg2+-ATP (O-OI-IO mM) and maintained at 30 °C for 40 min. Mg2+-ATP had noeffect on the transfer of radioactivity up to 0-316 mM, but at higher concentrationsthe membranes agglutinated in a similar way to that induced by high concentrationsof Ca2+.
176 E. A-H. Baydoun and D. H. Northcote
Enzymic modification of the membrane fractions and its effect on the transfer of radio-activity
Trypsin. Trypsin treatment of the individual membrane fractions did not influencetheir behaviour upon recentrifugation (Table 9). Radioactive Golgi apparatus-richfraction was mixed with non-radioactive plasma membrane-rich fraction at o°C for30 min in the presence of various concentrations of trypsin, and trypsin inhibitor(1:1-5, w/w, trypsin: trypsin inhibitor) was then added. After thorough mixing, Ca2+
was added to 1 mM and the membranes were maintained at 30 °C for a further 40 min.The transfer of radioactivity was markedly inhibited by trypsin pre-treatment (Table 1 o).
Table 7. Effect of Ca2+ on the transfer of radioactivity between mixed membrane fractions
Radioactivity (%) fromrefractionated membranes after mixing
[Ca1+], mM. . .
FractionGolgi apparatus-richPlasma membrane-richRemainder fractionTotal cpm obtainedfrom all the fractions
Recovery
0
8857 93 6
I353O
87-9
o-oi
77'5i7'55°
13900
90-3
0-0316
74-522-8
2 7
I442O
9 3 7
O' l
6 2 6
33'44 0
14700
955
1
5763 9 5
2-914540
94'5
Roots were incubated with D-[U-14C]glucose in vivo. Golgi apparatus-rich fraction was pre-pared from radioactive roots. Plasma membrane-rich fraction was prepared from non-radio-active roots. These were mixed at 30 °C for 40 min in the presence of various concentrationsof Ca1+ and refractionated by centrifugation. The distribution of radioactivity in the fractionswas determined.
Table 8. Effect of divalent cations on the transfer of radioactivity between mixedmembrane fractions
Radioactivity (%) fromrefractionated membranes after mixing
Cation (1 mM) added. . .
FractionGolgi apparatus-richPlasma membrane-richRemainder fractionTotal cpm obtained fromall the fractions
Recovery
None
9 1 2
7'4i - 4
8210
90-4
Ba»+
8 4 2
I4-3i '5
8610
9 4 9
Mn« +
5 6 142-7
1-2
8890
9 7 9
Sr» +
7 9 41 8 9
1-78470
9 3 3
Roots were incubated with D-[U-14C]glucose in vivo. Golgi apparatus-rich fraction wasprepared from radioactive roots. Plasma membrane-rich fraction was prepared from non-radioactive roots. These were mixed at 30 °C for 40 min in the presence of various divalentcations (1 mM final apparent free concentration) and refractionated by centrifugation. Thedistribution of radioactivity in the fractions was determined.

Measurement of membrane fusion in vitro 177
Trypsin treatment had 2 other effects in parallel: (a) it caused a net loss of membraneprotein (Table 11); and (b) it selectively eliminated one particular polypeptide(mol. wt approx. 36000) as shown in Fig. 3; a trypsin concentration of 3i-6/*g ml"1
was optimal for all 3 effects.
50
40
50
40
0
-11-001 01
[Colchicine], ITIM
10
2-5 5 7-5Cytochalasin B, fig ml"1
10
50 1-
40
0
40 ,
0-05 0-1Caffeine, %
0-15
30 r
0001 0-1 1 0
[lodoacetamide], mM
Fig. 2. Effect of colchicine, cytochalasin B, caffeine and iodoacetamide on the transferof radioactivity between mixed membrane fractions. The effect of each additive wasdetermined from an independent membrane preparation. Roots were incubated withD-[U-14C]glucose in vivo. Golgi apparatus-rich fraction was prepared from radioactiveroots. Plasma membrane-rich fraction was prepared from non-radioactive roots. Thesewere mixed at 30 CC for 15 min in the presence of various concentrations of colchicine,cytochalasin B, caffeine or iodoacetamide. Caa+ (1 mM) was then added and mem-branes were further mixed for 40 min and refractionated by centrifugation. Theradioactivity transferred to the plasma membrane-rich fraction was determined.
Add phosphatase. Radioactive Golgi apparatus-rich fraction was mixed with non-radioactive plasma membrane-rich fraction in the presence of various concentrationsof acid phosphatase and maintained at 30 °C for 60 min without added Ca2+. Theenzyme had no effect on the transfer of radioactivity (Fig. 4). In another experiment,the membranes were mixed at 30 °C for 60 min in the presence of acid phosphataseand absence of Ca2+, and then maintained in the presence of 1 mM Ca2+ for a further40 min. Acid phosphatase did not interfere with the action of Ca2+ on the transfer ofradioactivity (Fig. 4).
178 E. A-H. Baydoun and D. H. Northcote
DISCUSSION
The present experiments have demonstrated the transfer of radioactivity betweenmixed Golgi apparatus- and plasma membrane-rich fractions in the presence of Ca2+.Little or no transfer of radioactivity occurred in the absence of Ca2+.
The transfer of radioactivity between membrane fractions labelled with D-[U-14C]-glucose could be due to exchange of radioactivity contained within the membrane
Table 9. Effect of trypsin treatment on individual membrane fractions
FractionGolgi apparatus-richPlasma membrane-richRemainder fractionTotal cpm obtained from
all the fractionsRecovery
Radioactivity (%) fromrefractionated membranes after trypsin
GA aftertrypsin
treatment
8859-22-3
5980
644
Control
8998-3i-8
8240
88-7
PM aftertrypsin
treatment
8-6894
2 - 0
4590
69-2
treatment
Control
7'39 1 5
1-2
58OO
87H
Roots were incubated with D-[U-l4C]glucose in vivo. Golgi apparatus- and plasma membrane-rich fractions were prepared from radioactive roots. These were treated individually withtrypsin (ioo fig ml"1) at o °C. After 30 min, trypsin inhibitor was added (1:1-5, w/w, trypsin:trypsin inhibitor) and membrane3 were then refractionated by centrifugation. The distribution ofradioactivity in the fractions was determined. In the control, heat-inactivated enzyme was used.
Table 10. Effect of trypsin on the transfer of radioactivity betweenmixed membrane fractions
Trypsin, fig ml"1. . .
FractionGolgi apparatus-richPlasma membrane-richRemainder fractionTotal cpm obtainedfrom all the fractions
Recovery
0
5 i - i462
2-710310
947
Radioactivity (%)refractionated membranes
A
I IO
53-o 73-i440 24-4
3 0 2-59910 8580
91-0 78-9
fromafter mixing
3 1 6
77-92O-2
1-986lO
7 9 1
IOO
77-S2O'O
2-57580
696
Roots were incubated with D-[U-l4C]glucose in vivo. Golgi apparatus-rich fraction was pre-pared from radioactive roots. Plasma membrane-rich fraction was prepared from non-radio-active roots. These were mixed at o °C for 30 min in the presence of various concentrations oftrypsin; trypsin inhibitor (1:1-5, w/w, trypsin: trypsin inhibitor) was then added. Afterthorough mixing Ca1+ was added to 1 mM and the membranes were maintained at 30 °C for40 min and refractionated by centrifugation. The distribution of radioactivity in the fractionswas determined. In the control, heat-inactivated enzyme (100 fig ml"1) was used.

Measurement of membrane fusion in vitro 179
vesicles or it could be due to transfer of part of the membrane components. Thesecond suggestion is more likely since a similar transfer was obtained when diazotizedradioactive sulphanilic acid was used. This reagent labels proteins and some phos-pholipids (e.g. phosphatidylethanolamine and phosphatidylserine) by forming
M T 1 2 3 4 5Mol. wt
68000
40000
I
1430012400
+ve
Fig. 3. SDS-polyacrylamide gel electrophoresis of the trypsin-treated mixed membranefractions. Track 1: mixed membranes with no trypsin added. Track 2: mixed mem-branes in the presence of 1 /tg ml"1 trypsin. Track 3: mixed membranes in thepresence of 10 fig nil"1 trypsin. Track 4: mixed membranes in the presence of31-6/tgml"1 trypsin. Track 5: mixed membranes in the presence of 100 fig ml"1
trypsin. Approx. 100 fig of proteins were applied per track. Track M: proteinmarkers with molecular weights as indicated. Track T: trypsin and trypsin inhibitor(10 fig each). Golgi apparatus- and plasma membrane-rich fractions were preparedfrom maize roots and mixed at o °C for 30 min in the presence of various concentra-tions of trypsin. Trypsin inhibitor was then added (1:1-5, w/w, trypsin: trypsininhibitor) and the membranes were washed twice by sequential centrifugation andresuspension in 10 mM Tris-HCl, pH 7-4. The final pellets were analysed on a 15 %,gel at 200 V for approx. 5 h. Staining was with Coomassie brilliant blue.

180 E. A-H. Baydoun and D. H. Northcote
covalent bonds with them. In addition when radioactive choline, which was incor-porated only into phosphatidylcholine (Baydoun & Northcote, 1980), was used tolabel the membranes a similar transfer was obtained.
The transfer of radioactivity seems therefore to be due to membrane fusion.
However, other possibilities such as membrane aggregation, diffusion of some mem-
Table 11. Amount of protein in trypsin-treated mixed membrane fractions
[Trypsin], Protein (mg), remain-ing in membranes
0 0-461 042
10 034316 0-33
100 0-33
Golgi apparatus- and plasma membrane-rich fractions prepared from maize roots weremixed at o °C for 30 min in the presence of various concentrations of trypsin. Trypsin inhibitorwas then added (1:1-5, w/w> trypsin: trypsin inhibitor) and the membranes were washed twiceby sequential centrifugation and resuspension in 10 mM Tris-HCl, pH 7-4. Protein in the finalpellet was estimated by the method of Lowry, Rosebrough, Farr & Randall (1951).
50
40 •
"S
I 30
« 20 -
•oDC
10 .
y
•
I
O
1 1 1
8 16 24
Acid phojphatase, M9 ml~'
32
Fig. 4. Effect of acid phosphatase on the transfer of radioactivity between mixed mem-brane fractions in the presence (O) or absence (#) of Ca1+. The effect of each treat-ment wa3 determined from an independent membrane preparation. Roots were incu-bated with D-[U-l4C]glucose in vivo. Golgi apparatus-rich fraction was prepared fromradioactive roots. Plasma membrane-rich fraction was prepared from non-radioactiveroots. These were mixed in the presence of various concentrations of acid phosphataseand maintained at 30 °C for 60 min without added Ca1+. In another experiment, themembranes were mixed at 30 °C for 60 min in the presence of acid phosphatase andabsence of Ca1+, and then maintained in the presence of 1 mM Ca1+ for a further 40min. Mixed membranes were refractionated by centrifugation. The radioactivitytransferred to the plasma membrane-rich fraction was determined. In the control, heat-inactivated enzyme (32 fig ml"1) was used.

Measurement of membrane fusion in vitro 181
brane components or molecular exchange of membrane components cannot be com-pletely excluded. The addition of excess EDTA before refractionation chelated Ca2+
and ensured disaggregation of non-fused vesicles. Transfer of radioactivity haspreviously been proposed as a method of studying the fusion of phospholipid vesicles(Papahadjopoulos et al. 1974). The transfer of phospholipids and glycolipids betweencells has been reported by several workers (Peterson & Rubin, 1970; Huang, 1976,1977; Collard, De Wildt & Inbar, 1978); this transfer is believed to be due to mem-brane fusion.
Vesicles of intermediate densities might be expected to be formed as a result ofmembrane fusion. This, however, would not be apparent from our experimentsbecause refractionation of mixed membranes was carried out by using the samesucrose interfaces used to isolate the original fractions. Any newly formed membranevesicle of intermediate density would find its way to one of the sucrose interfaces(either 25-34% or 34-45% interface) during the recentrifugation. If the transfer ofradioactivity is due to exchange of small pieces of membrane components, the totaldensity of the membrane fractions would not be expected to change.
The extent of the membrane fusion was dependent on the concentration of Ca2+
up to a final concentration of i-o mM. This Ca2+ concentration range is similar to thatfound for other biological systems used to study membrane fusion in vitro (Gratzl,Dahl, Russell & Thorn, 1977; Gratzl & Dahl, 1978). Although membrane fusionbetween isolated secretory vesicles from rat liver was reported to be Ca2+-specific(Gratzl & Dahl, 1978), our results showed that other divalent cations were as effectiveas Ca2+ or had a moderate effect. This is in good agreement with the results of Dahlet al. (1978) on the fusion of isolated myoblast plasma membranes and those ofHolz & Stratford (1979) who found that Mn2+ was the most effective ion in inducingfusion between lipid vesicles. Ca2+ ions are important and widely used for the induc-tion of fusion between artificial lipid membranes. However, the concentrationsrequired are much higher than those needed for biological membranes (Holz &Stratford, 1979).
The failure, in our work, of similar concentrations of Mg2+ to mimic the effectsshown by Ca2+ could be due to the differences in their chemistry. Ca2+ has a coordina-tion number of 7 or 8 and its coordination geometry is irregular in both bond angleand bond length, which is quite different from Mg2+. Mg2+ maintains 6-coordinationin a closely regular octahedron. The structural role of the 2 ions in biology is thereforequite different. Mg2+ is weaker in its ability to bind to the irregular geometries ofcoordination sites of biological molecules such as proteins (Williams, 1976). Higherconcentrations of Mg2+ as compared to Ca2+ were able to induce fusion in variousbiological systems (Howell & Tyhurst, 1977; Dahl et al. 1978).
Evidence is increasing for the function of Ca2+ in membrane fusion. Most of thisevidence is obtained indirectly from observations on the factors that control secretionin animal (Cochrane & Douglas, 1974; Zabucchi, Soranzo, Rossi & Romeo, 1975;Baker & Knight, 1978; Matt, Bilinski & Plattner, 1978), and plant cells (Morris &Northcote, 1977). The mechanism by which Ca2+ ions affect membrane fusion ispoorly understood. It has been suggested that Ca2+ ions decrease the electrical

182 E. A-H. Baydoun and D. H. Northcote
repulsion between negatively charged membranes, resulting in their close approach.This will lead to membrane contact by van der Waals forces and finally membranefusion (Dean, 1975; Howell&Tyhurst, 1977). Close contact between fusing membranesforming pentalaminar layers has been revealed by electron microscopy (Lagunoff,1973), but further events in membrane fusion were difficult to investigate. In ourwork, although membrane fusion was time-dependent, electron microscopy did notreveal any of its mechanism. The events leading to fusion are believed to occur inmilliseconds. This short life-time, as well as the possible instability of any inter-mediate structures, could account for the difficulties in their detection (Lagunoff,1973). In our electron-microscope studies the heterogeneity in membrane vesicle sizeprecluded any observation of vesicle enlargement due to fusion.
Using artificial bilayer lipid membranes Papahadjopoulos et al. (1977) suggestedthat Ca2+ induced fusion by a phase change which resulted in a transient destabiliza-tion of the bilayer membranes making them susceptible to fusion. It was also suggestedthat Ca2+ affected every step in the events leading to membrane fusion, includingincrease in membrane fluidity (Cullis & Hope, 1978).
Membrane fluidity is probably very important for fusion. The biochemical changesaccompanying membrane fusion are greatly facilitated by the presence of a dynamicmembrane structure. These changes include the perturbation of the lipid bilayer toproduce protein-free regions which are able to fuse (Ahkong et al. 1975; Lemay et al.1977). Raising the temperature is a factor that increases membrane fluidity and givesrise to structural instability leading to membrane fusion (Breisblatt & Ohki, 1976;Prives & Shinitzky, 1977). In the present work, the in vitro membrane fusion betweenGolgi apparatus- and plasma membrane-rich fractions was temperature-dependent.This result is in good agreement with those obtained for membrane fusion betweensecretory vesicles of rat liver (Gratzl & Dahl, 1978) and isolated myoblast plasmamembranes (Dahl et al. 1978).
Cytochalasin B, a drug disrupting microfilaments, had no effect in our study on themembrane fusion between Golgi apparatus- and plasma membrane-rich fractionsin vitro. This agrees with the findings of Dahl et al. (1978) on the fusion of isolatedmyoblast plasma membranes. Cytochalasin B is known to affect secretion in plants byinhibiting the movement of secretory vesicles from the site of production at the Golgiapparatus to the site of fusion at the cell surface (Mollenhauer & Morre, 1976;Pope, Thorpe, Al-Azzawi & Hall, 1979). Our results are different from those ofMiyake, Kim & Okada (1978), who reported an inhibitory effect of cytochalasin D onfusion of cells mediated by Sendai virus. Colchicine, a substance capable of disruptingmicrotubules, also affects the movement of vesicles; it did not show any effect on themembrane fusion in the present in vitro investigation. This disagrees with the resultsof Gratzl & Dahl (1978) who reported inhibitory effect of colchicine on membranefusion in vitro between secretory vesicles of rat liver. In their system, as in ours,membranes were isolated in the cold: a condition resulting in depolymerization ofmicrotubules within the cytoplasm.
Caffeine has been shown to inhibit cytokinesis in several plants by interfering withthe arrangement and fusion of membrane vesicles destined to form the cell plate

Measurement of membrane fusion in vitro 183
(Becerra, 1977; Jones & Payne, 1978). In the present work, membrane fusion was notaffected by caffeine even at concentrations higher than those shown to inhibit cyto-kinesis in vivo. Recently, it was reported that caffeine destroys or disorganizes themicrotubules associated with the formation of secondary cell walls (Juniper & Lawton,1979). This could explain the lack of effect shown by caffeine in our in vitro system.
In our investigation, iodoacetamide, a sulphydryl blocking reagent had no effecton membrane fusion. This disagrees with the results of Gratzl & Dahl (1978) andthose of Lalazar, Michaeli & Loyter (1979) who reported that sulphydryl-blockingreagents inhibit membrane fusion. In their investigation only sulphydryl-blockingreagents containing Hg showed an inhibitory effect. Other reagents lacking Hg wereineffective. Thus, it seems likely that the inhibition reported by these workers wasnot due to blocking the sulphydryl groups of the membranes but resulted from otherchanges in the membranes induced by the Hg atom.
It has been suggested that Ca2+ affected biological membranes by specific bindingto proteins on the membrane surface (McLaughlin, Grathwohl & McLaughlin, 1978).To investigate the role of proteins in membrane fusion, mixed membrane fractionswere treated with trypsin at o °C before the addition of Ca2+. The ability of themembranes to fuse was significantly inhibited by this treatment. This is in goodagreement with the results obtained from other systems in which membrane fusionhas been studied in vitro (Milutinovic et al. 1977; Gratzl & Dahl, 1978; Dahl et al.1978). Our results indicated that Ca2+ could mediate the formation of bridges betweenmixed membrane vesicles through the negatively charged sites on the surface proteinsor glycoproteins. Such bridges would bring membranes in close contact, a pre-requisite for membrane fusion. The selective loss of one of the polypeptides of themixed membrane fractions after trypsin treatment could indicate a possible relation-ship between this specific peptide and the sites of Ca2+ action on membranes.
It seems unlikely that Ca2+ could bind to membrane vesicles through phosphategroups attached to proteins, as treatment of mixed membranes with acid phosphatasedid not affect membrane fusion. It has been reported that the major part of the surfacecharge originates from the phosphate groups at the cell surface of some plant proto-plasts (Nagata & Melchers, 1978). In the present experiments, trypsin treatment did notcompletely abolish membrane fusion. This could be due to the presence of certainproteins which were not removed by trypsin under the experimental conditionsused; alternatively, it could result from Ca2+ binding to acidic phospholipids tobring membranes in contact. The latter suggestion seems to be true for artificiallipid membranes (Papahadjopoulos et al. 1977; Portis et al. 1979).
REFERENCES
AHKONG, Q. F., FISHER, D., TAMPION, W. & LUCY, J. A. (1973). The fusion of erythrocytesby fatty acids, esters, retinol and a-tocopherol. Biochem.J. 136, 147-155.
AHKONG, Q. F., FISHER, D., TAMPION, W. & LUCY, J. A. (1975). Mechanisms of cell fusion.Nature, Lond. 353, 194-195.
ALLEN, R. D. & WOLF, R. W. (1979). Membrane recycling at the cytoproct of Tetrahymena.J. Cell Set. 35, 217-227.

184 E. A-H. Baydoun and D. H. Northcote
BAKER, P. F. & KNIGHT, D. E. (1978). Calcium-dependent exocytosis in bovine adrenalmedullary cells with leaky plasma membrane. Nature, Lond. 276, 620-622.
BAYDOUN, E. A-H. & NORTHCOTE, D. H. (1980). Isolation and characterization of membranesfrom the cells of maize root tips. jf. Cell Set. 45, 147-167.
BECERRA, J. (1977). Calcium and magnesium in plant cytokinesis and their antagonism withcaffeine. Experientia 33, 1318—1319.
BERGER, W., DAHL, G. & MEISSNER, H. P. (1975). Structural and functional alterations infused membranes of secretory granules during exocytosis in pancreatic islet cells of the mouse.Cytobiologie 12, 119-139.
BREISBLATT, W. & OHKI, S. (1976). Fusion of phospholipid spherical membranes. II. Effectof cholesterol, divalent ions and pH. J. Membrane Biol. 29, 127-146.
BURWEN, S. J. & SATIR, B. (1977). A freeze-fracture study of early membrane events duringmast cell secretion. J. Cell Biol. 73, 660-671.
COCHRANE, D. E. & DOUGLAS, W. W. (1974). Calcium-induced extrusion of secretory granules(exocytosis) in mast cells exposed to 48/80 or the ionophores A-23187 and X-537A. Proc.natn. Acad. Sci. U.S.A. 71, 408-412.
COLLARD, J. C, DE WILDT, A. & INBAR, M. (1978). Translocation of a fluorescent lipid probebetween contacting cells. Evidence for membrane lipid interactions. FEBS Letters, Amster-dam 90, 24-28.
CULLIS, P. R. & HOPE, M. J. (1978). Effects of fusogenic agent on membrane structure oferythrocyte ghosts and the mechanism of membrane fusion. Nature, Lond. 271, 672-674.
DAHL, G. & GRATZL, M. (1976). Calcium-induced fusion of isolated secretory vesicles from theislet of Langerhans. Cytobiologie 12, 344-355.
DAHL, G., SCHUDT, C. & GRATZL, M. (1978). Fusion of isolated myoblast plasma membranes.An approach to the mechanism. Biochim. biophys. Acta 514, 105-116.
DA SILVA, P. P. & NOGUEIRA, M. L. (1977). Membrane fusion during secretion. A hypothesisbased on electron microscope observation of Phytophthora palmivora zoopores during encyst-ment. J. CellBiol. 73, 161-181.
DE CAMILLI, P., PELUCHETTI, D. & MELDOLESI, J. (1976). Dynamic changes of the luminalpi asmalemma in stimulated parotid acinar cells. A freeze-fracture study. J. Cell Biol. 70,59—74.
DEAN, P. M. (1975). Exocytosis modelling: an electrostatic function for calcium in stimulus-secretion coupling. J. theor. Biol. 54, 289-308.
GRATZL, M. & DAHL, G. (1976). Cat+-induced fusion of Golgi-derived secretory vesiclesisolated from rat liver. FEBS Letters, Amsterdam 62, 142-145.
GRATZL, M. & DAHL, G. (1978). Fusion of secretory vesicles isolated from rat liver. J. Mem-brane Biol. 40, 343-364.
GRATZL, M., DAHL, G., RUSSELL, J. T. & THORN, N. A. (1977). Fusion of neurohypophysealmembranes in vitro. Biochim. biopkys. Acta 470, 45-57.
HARWOOD, J. L. (1976). Biosynthesis of phosphatidylcholine and phosphatidylethanolamineby germinating soya bean. Biochem. Soc. Trans. 4, 50-52.
HOLZ, R. W. & STRATFORD, C. A. (1979). Effects of divalent ions on vesicle-vesicle fusionstudied by a new luminescence assay for fusion. J. Membrane Biol. 46, 331—358.
HORWITZ, A. F., WIGHT, A. & KNUDSEN, K. (1979). A role for lipid in myoblast fusion.Biochem. biophys. Res. Commun. 86, 514-521.
HOWELL, S. L. & TYHURST, M. (1977). Distribution of anionic sites on surface of B cellgranule and plasma membranes: a study using cationic ferritin. J. Cell Sci. 27, 289—301.
HUANG, R. T. C. (1976). Labelling of animal cells with fluorescent dansyl cerebroside. Z.Naturforsch. 31 c, 737-740.
HUANG, R. T. C. (1977). Transfer of glycolipid between membranes of tissue culture cells,using dansylcerebroside as a model. Z. Naturforsch. 32 c, 379-383.
JONES, M. G. K. & PAYNE, H. L. (1978). Cytokinesis in Jmpatiens balsamina and the effect ofcaffeine. Cytobios 20, 79-91.
JUNIPER, B. E. & LAWTON, J. R. (1979). The effect of caffeine, different fixation regimes andlow temperature on microtubules in the cells of higher plants. Evidence for diversity intheir response to chemical and physical treatments. Planta 145, 411-416.
KALDERON, N. & GILULA, N. B. (1979). Membrane events involved in myoblast fusion. J. CellBiol. 81, 411-425.

Measurement of membrane fusion in vitro 185
KAO, K. N. & MICHAYLUK, M. R. (1974). A method for high-frequency intergeneric fusionof plant protoplasts. Planta 115, 355-367.
KOERNER, J. F. & SINSHEIMER, R. L. (1957). A deoxyribonuclease from calf spleen. I. Purifi-cation and properties. J. biol. Chem. 228, 1039-1048.
KOTER, M., D E KRUIJFF, B. & VAN DEENEN, L. L. M. (1978). Calcium-induced aggregationand fusion of mixed phosphatidylcholine-phosphatidic acid vesicles as studied by 31P NMR.Biochim. biophys. Ada 514, 255-263.
KNUTTON, S. (1977). Studies of membrane fusion. II. Fusion of human erythrocytes bySendai virus. J. Cell Sci. 28, 189-210.
KNUTTON, S. (1979). Studies of membrane fusion. III. Fusion of erythrocytes with poly-ethylene glycoLJ. Cell Sci. 36, 61-72.
LAGUNOFF, D. (1973). Membrane fusion during mast cell secretion. J. Cell Biol. 57, 252-259.LALAZAR, A., MICHAELI, D. & LOYTER, A. (1977). Restoration of the fusion capacity of human
erythrocyte ghosts by the blocking reagents. Expl Cell Res. 107, 79—88.LEMAY, P., COLLYN-D'HOOGHE, M. & TORPIER, G. (1977). XC-cell fusion induced by murine
plasmocytoma cells. IV. A dynamic analysis by microcinematography and freeze-fracture.Expl Cell Res. 109, 70-86.
LIAO, M. J. & PRESTEGARD, J. H. (1979). Fusion of phosphatidic acid-phosphatidylcholinemixed lipid vesicles. Biochim. biophys. Ada 550, 157-173.
LOWRY, O. H., ROSEBROUGH, N. J., FARR, A. L. & RANDALL, R. J. (1951). Protein measure-ment with the Folin phenol reagent. J. biol. Chem. 193, 265-275.
MANEON, W. T., HLADKY, S. B. & HAYDON, D. A. (1979). Fusion of photoreceptor membranevesicles. J. Membrane Biol. 46, 171-181.
MATT, H., BILINSKI, M. & PLATTNER, H. (1978). Adenosine triphosphate, calcium and tem-perature requirements for the final steps of exocytosis in Parameciirm cells. J. Cell Sci. 32,67-86.
MCLAUGHLIN, A., GRATHVVOHL, C. & MCLAUGHLIN, S. (1978). The adsorption of divalentcations to phosphatidylcholine bilayer membranes. Biochim. biophys. Ada 513, 338—357.
MILUTINOVIC, S., ARGENT, B. E., SCHULZ, I. & SACHS, G. (1977). Studies on isolated subcellu-lar components of cat pancreas. J. Membrane Biol. 36, 281-295.
MIYAKE, Y., KIM, J. & OKADA, Y. (1978). Effects of cytochalasin D on fusion of cells by HVJ(Sendai virus). Cell-cell fusion is separable from cell-virus fusion. Expl Cell Res. 116,167-178.
MOLLENHAUER, H. H. & MORRE, D. J. (1976). Cytochalasin B, but not colchicine, inhibitsmigration of secretory vesicles in root tips of maize. Protoplasma 87, 39-48.
MORRIS, M. R. & NORTHCOTE, D. H. (1977). Influence of cations at the plasma membrane incontrolling polysaccharide secretion from sycamore suspension cells. Biochem. J. 166,603-618.
NAGATA, T. & MELCHERS, G. (1978). Surface charges of protoplasts and their significance incell-cell interaction. Planta 142, 235-238.
NORTHCOTE, D. H. (1979). The involvement of the Golgi apparatus in the biosynthesis andsecretion of glycoproteins and polysaccharides. Biomembranes 10, 51-76.
ORCI, L., PERRELET, A. & FRIEND, D. S. (1977). Freeze-fracture of membrane fusions duringexocytosis in pancreatic B-cells. J. Cell Biol. 75, 23-30.
PAPAHADJOPOULOS, D., POSTE, G., SCHAEFFER, B. E. & VAIL, W. J. (1974). Membrane fusionand molecular segregation in phospholipid vesicles. Biochim. biophys. Ada 352, 10-28.
PAPAHADJOPOULOS, D., VAIL, W. J., NEWTON, C , NIR, S., JACOBSON, K., POSTE, G. & LAZO, R.
(i977)- Studies on membrane fusion. III. The role of calcium-induced phase changes.Biochim. biophys. Ada 465, 579-598.
PAPAHADJOPOULOS, D., VAIL, W. J., PANGBORN, W. A. & POSTE, G. (1976). Studies on mem-brane fusion. II. Induction of fusion in pure phospholipid membranes by calcium ions andother divalent metals. Biochim. biophys. Ada 448, 265-283.
PETERSON, J. A. & RUBIN, H. (1970). The exchange of phospholipids between cultured chickembryo fibroblasts as observed by autoradiography. Expl Cell Res. 60, 383-392.
PLATTNER, H. (1974). Intramembraneous changes on cationophore-triggered exocytosis inParamecium. Nature, Lond. 252, 722-724.
13 CEL 45

186 E. A-H. Baydoun and D. H. Northcote
POPE, D. G., THORPE, J. R., AL-AZZAWI, M. J. & HALL, J. L. (1979). The effect of cytochalasinB on the rate of growth and ultrastructure of wheat coleoptiles and maize roots. Planta 144,373-383-
PORTIS, A., NEWTON, C, PANGBORN, W. & PAPAHADJOPOULOS, D. (1979). Studies on themechanism of membrane fusion: evidence for an intermembrane Ca2+-phospholipid com-plex, synergism with Mg1+ and inhibition by spectrin. Biochemistry, N.Y. 18, 780-790.
POSTE, G. & ALLISON, A. C. (1973). Membrane fusion. Biochim. biophys. Acta 300, 421-465.PRIVES, J. & SHINITZKY, M. (1977). Increased membrane fluidity precedes fusion of muscle
cells. Nature, Lond. 268, 761-763.PUSKIN, J. S. & MARTIN, T. (1979). Divalent cation binding to phospholipid vesicles. Depen-
dence on temperature and lipid fluidity. Biochim. biophys. Acta 552, 53-65.ROBERTS, K. & NORTHCOTE, D. H. (1970). The structure of sycamore callus cells during division
in a partially synchronized suspension culture. J. Cell Sci. 6, 299-321.ROBERTS, K. & NORTHCOTE, D. H. (1971). Ultrastructure of the nuclear envelope; structural
aspects of the interphase nucleus of sycamore suspension culture cells. Microscopica Acta 71,102-120.
SATIR, B. (1974). Ultrastructural aspects of membrane fusion. J. supramolec. Struct. 2, 529-537.SATIR, B., SCHOOLEY, C. & SATIR, P. (1973). Membrane fusion in a model system. Mucocyst
secretion in Tetrahymena. J. Cell Biol. 56, 153-176.SINGER, S. J. & NICOLSON, G. L. (1972). The fluid mosaic model of the structure of cell
membranes. Science, N.Y. 175, 720-731.STEINMAN, R. M., BRODIE, S. E. & COHN, Z. A. (1976). Membrane flow during pinocytosis.
A stereologic analysis. J. Cell Biol. 68, 665-687.SVOBODA, A. (1978). Fusion of yeast protoplasts induced by polyethylene glycol. J. gen.
Microbiol. 109, 169-175.VOLSKY, D. & LOYTER, A. (1977). Rearrangement of intramembranous particles and fusion
promoted in chicken erythrocytes by intracellular Cai+. Biochim. biophys. Acta 471, 243-259.WILLIAMS, R. J. P. (1976). Calcium chemistry and its relation to biological function. In
Calcium in Biological Systems (ed. C. J. Duncan), pp. 1-17. Cambridge: Cambridge Univer-sity Press.
YANOVSKY, A. & LOYTER, A. (1972). The mechanism of cell fusion. I. Energy requirementsfor virus-induced fusion of Ehrlich ascite3 tumor cells. J. biol. Chem. 247, 4021-4028.
ZABUCCHI, G., SORANZO, M. R., ROSSI, F. & ROMEO, D. (1975). Exocytosis in human poly-morphonuclear leukocytes induced by A 23187 and calcium. FEBS Letters, Amsterdam 54,44-48.
(Received 20 November 1979)