maternal high fat diet alters the transcriptional … · board games, brunch and dinner nights,...
TRANSCRIPT

Maternal High Fat Diet Alters the Transcriptional Response to
Glucocorticoid and Immune Challenge in the Amygdala and
Hippocampus of Adult Rat Offspring
by
Christine Min Wei Lum
A thesis submitted in conformity with the requirements
for the degree of Master of Science
Graduate Department of Cell & Systems Biology
University of Toronto
© Copyright by Christine Min Wei Lum 2016

ii
Maternal High Fat Diet Alters the Transcriptional Response to Glucocorticoid and Immune
Challenge in the Amygdala and Hippocampus of Adult Rat Offspring
Master of Science 2016, Christine Min Wei Lum, Cell & Systems Biology, University of
Toronto
Abstract
Maternal overnutrition during pre- and post-(peri)natal periods of development has previously
been suggested to increase offspring risk for developing psychiatric disorders. The mental
impacts are often accompanied by altered expressions of glucocorticoid receptors (GR), which
regulate the inflammatory response to stress. In this study, rats were acutely injected with
corticosterone (CORT) and/or lipopolysaccharide (LPS) in adulthood after perinatal exposure to
maternal high-fat diet (HFD) to examine its effects on responding to stress. Gene transcript
analyses in the amygdala and hippocampus revealed higher immunosuppressive activity and
altered abundance of genes associated with anxiety- and depressive-like behavior compared to
rats perinatally exposed to standard diets. These effects were sex and brain region-specific, and
appeared to coincide with levels of GR. Epigenetic analyses suggest impairments in the DNA
methylation machinery to be related to these diet differences. Hence, perinatal HFD incurs
differential immune responses to stress in the rodent brain.

iii
Acknowledgements
I would like to thank all my friends back home, and at UTSC for offering me their kind, listening
ear, emotional support, comedic relief, and place of refuge. Thank you all for reminding me that
even though no one's perfect, and that everyone has needs which may conflict with our own,
most people want to help and empathize, and that every small gesture counts.
I would also like to thank my family for providing me a steady supply of energy to keep me
going through each and every day, and for always trying to adapt to my needs. I want to give a
special shout-out to my sister Jocelyn for showing me that I will always be loved no matter
where I am, and who I become as I keep trying to move forward.
Finally, I want to thank my lab mates for the good times we've had together ex. the road trips,
board games, brunch and dinner nights, Lego movie - and for the rare times in my life where I
have truly, deeply been candid with another person, and have felt that I was known and cherished
in return...whatever comes next.

iv
Contributions
I am grateful to the students and faculty at UTSC for their help with the following:
Professor Patrick McGowan - Supervision, mentorship, provision of great ideas and direction,
sacrifice of rats and brain extractions.
Wilfred de Vega, PhD Candidate - Sacrifice of rats and brain extractions, brain microdissections,
portion of qPCRs for LPS condition, evaluation of housekeeping gene stability with geNorm (R-
package), general assistance.
Sameera Abuaish, MSc - Brain microdissections.
Shathvee Sivanathan, Atif Hussain, Patrick Ng, BSc - Handling and weighing of rats, maternal
behavioral coding, drug injections.
Ben Hing, PhD; Professors Mauricio Terebiznik, Joanne Nash - Excellent research advice and
provision of critical thought.
Aya Sasaki, MSc - Sacrifice of rats and brain extractions, teaching brain microdissections.
Pauline Pan and Sophie St-Cyr, MSc - Technical advice on DNA cleanup.

v
Table of Contents
Acknowledgments..........................................................................................................................iii
Contributions..................................................................................................................................iv
Table of Contents.............................................................................................................................v
List of Tables..................................................................................................................................ix
List of Figures..................................................................................................................................x
List of Supplementary Tables.........................................................................................................xi
List of Abbreviations.....................................................................................................................xii
Chapter 1 Introduction.....................................................................................................................1
1.1 Effects of Maternal Obesity and Overnutrition on Offspring physical and mental
health....................................................................................................................................2
1.2 Perinatal Programming of the HPA-axis: Effect on Mental Health..............................4
1.2.1 Effects of GR and MR on Anxiety and Depression........................................7
1.2.2 Overexposure to GCs alters GR and MR expression......................................8
1.3 Mediation of CORT and LPS-induced Inflammatory Changes Through GR and
MR..................................................... .................................................................................8
1.3.1 The General Immunosuppressive Action of GCs...........................................9
1.3.2 Inflammatory Actions of GCs in the Central Nervous System (CNS).........11

vi
1.4 Neuroinflammation, GR, and Mental Disorders: Implications of Maternal
Overnutrition......................................................................................................................13
1.4.1 How Cytokines in the CNS Promote Psychopathology................................14
1.4.2 Caveats to Consider with Maternal Overnutrition Studies...........................16
1.4.3 Effects of Perinatal HFD on Inflammation...................................................16
1.4.4 Effects of Perinatal HFD on the HPA-axis as a Consequence of
Inflammation..........................................................................................................18
1.5 Role of Epigenetic Regulation in Incurring Lifelong Effects of Maternal Over
Nutrition ............................................................................................................................19
Box 1. Regulators of DNA methylation................................................................20
1.6 Role of Epigenetic Regulation in Responding to Stress..............................................21
1.6.1 Gene-specific Functions................................................................................21
1.6.2 Mediation of Global Changes.......................................................................22
1.7 The Effects of Prior Experiences on Responses to Stress............................................22
1.7.1 Studies inspecting HPA-axis Activity..........................................................23
1.7.2 Studies examining Transcriptional and Epigenetic effects...........................24
1.7.3 Other studies with Inflammatory Pathways..................................................25
1.8 Research Hypotheses...................................................................................................25

vii
Chapter 2 Materials & Methods.....................................................................................................28
2.1 Animals........................................................................................................................28
2.2 Diets.............................................................................................................................28
2.3 Subjects and general procedures..................................................................................29
2.4 Drug challenges...........................................................................................................29
2.5 Brain tissue preparation...............................................................................................30
2.6 Analyses of transcript abundance by quantitative real-time reverse transcriptase-
polymerase chain reaction (qRT-PCR)..............................................................................31
2.7 Primer specifications....................................................................................................33
2.8 Measurement of DNMT enzyme activity and abundance in the amygdala.................34
2.9 Global CpG methylation analysis in the amygdala with Luminometric methylation
assay (LUMA) ..................................................................................................................34
2.10 Statistical analyses.....................................................................................................35
Chapter 3 Results...........................................................................................................................36
3.1 Perinatal-HFD offspring display differences in body weight throughout
development.......................................................................................................................36
3.2 Perinatal-HFD exposed animals display exacerbated anti-inflammatory transcriptional
responses to CORT challenge in the amygdala.................................................................37

viii
3.3 Perinatal-HFD exposure incurs similar, but sex-dependent transcriptional responses to
CORT in the hippocampus.................................................................................................41
3.4 Perinatal HFD-exposed animals display exacerbated anti-inflammatory transcriptional
responses to LPS challenge in the amygdala.....................................................................45
3.5 Perinatal-HFD exposed animals display altered pro-inflammatory transcriptional
responses to CORT/LPS challenge in the amygdala.........................................................48
3.6 Diet-dependent transcriptional responses cluster with transcript levels of epigenetic
regulators in the amygdala.................................................................................................52
3.7 DNMT activity reduces in response to CORT and CORT/LPS for Perinatal HFD-
exposed males, but not CORT/LPS females in the amygdala...........................................53
3.8 DNMT activity correlates with global CpG methylation in CORT-injected male
groups in the amygdala......................................................................................................56
Chapter 4 Discussion.....................................................................................................................57
4.1 Interpretation of results from CORT challenge...........................................................57
4.1.1 Diet x Sex differences in the Amygdala.......................................................57
4.1.2 Females' distinct response in the Hippocampus...........................................61
4.2 Results from LPS challenge in the amygdala..............................................................62
4.3 Results from CORT/LPS challenge in the amygdala..................................................63
4.4 Comparison of results with Sasaki et al. (2013) Maternal HFD Study.......................65

ix
4.5 Maternal HFD transmission of CORT as possible mechanism for differential GC
sensitivity...........................................................................................................................67
4.6 Role of Epigenetic Regulators: MeCP2 as potential hub for manifestation of diet- and
sex- differences..................................................................................................................68
4.7 Explaining the effects of DNMT activity on Global CpG methylation levels............70
4.8 Conclusion...................................................................................................................71
References.....................................................................................................................................71
Appendix......................................................................................................................................101
List of Tables
Table 1 Number of adult offspring in each challenge condition...................................................30
Table 2 List of genes analyzed for transcript abundance with qPCR............................................32
Table 3 Primer sequences and catalogue numbers for genes analyzed with qPCR.......................33
Table 4 Transcriptional response to CORT challenge in the amygdala........................................40
Table 5 Transcriptional response to CORT challenge in the hippocampus...................................44
Table 6 Transcriptional response to LPS challenge in the amygdala............................................47
Table 7 Transcriptional response to CORT/LPS challenge in the amygdala................................51

x
Table 8 Transcriptional response to CORT and CORT/LPS in the amygdala grouped by post-hoc
comparisons within drug................................................................................................................52
Table 9 Spearman's Correlation Tests Between DNMT transcript abundance and Total DNMT
activity............................................................................................................................................55
List of Figures
Figure 1 Summary of the HPA-axis from Smith and Vale (2006) ...............................................6
Figure 2 Simplified diagram of the general CORT/LPS pathway.................................................10
Figure 3 Perinatal-HFD regimen used in this study.......................................................................29
Figure 4 Body weights of offspring...............................................................................................36
Figure 5 Transcript level data for CORT-challenged males in the amygdala...............................38
Figure 6 Transcript level data for CORT-challenged females in the amygdala............................39
Figure 7 Transcript level data for CORT-challenged males in the hippocampus..........................42
Figure 8 Transcript level data for CORT-challenged females in the hippocampus......................43
Figure 9 Transcript level data for LPS-challenged males in the amygdala...................................45
Figure 10 Transcript level data for LPS-challenged females in the amygdala..............................46
Figure 11 Increases in IL-6/IL-10 ratios between LPS and CORT/LPS.......................................48
Figure 12. Transcript level data for CORT/LPS-challenged males in the amygdala....................49

xi
Figure 13 Transcript level data for CORT/LPS-challenged females in the amygdala..................50
Figure 14 DNMT Activity and Protein Assays.............................................................................54
Figure 15 Global CpG methylation measured by LUMA in the amygdala of CORT-injected
males..............................................................................................................................................56
Figure 16 Comparison of IL-6/IL-10 ratios between CORT-challenged males and females in the
amygdala........................................................................................................................................59
Figure 17 Simplified diagram of the effect of Perinatal HFD on the general CORT/LPS
pathway..........................................................................................................................................66
List of Supplementary Tables
Supplementary Table 1
Transcriptional response to CORT challenge in the amygdala (post-hocs)................................101
Supplementary Table 2
Transcriptional response to CORT challenge in the hippocampus (post-hocs)...........................102
Supplementary Table 3
Transcriptional response to LPS challenge in the amygdala (post-hocs)....................................103
Supplementary Table 4
Transcriptional response to CORT/LPS challenge in the amygdala (post-hocs)........................104

xii
List of Abbreviations
11-β-HSD2 11-Beta hydroxysteroid dehydrogenase 2
ActB Beta-actin
ACTH Adrenocorticotropic hormone
ANOVA Analysis of variance
BMI Body mass index
CBG Corticosteroid-binding globulin
CHD Control House chow diet
ChIP Chromatin immunoprecipitation
CNS Central nervous system
Co-IP Co-immunoprecipitation
CORT Cortisol/corticosterone
CRP C-reactive protein
Fisher's PLSD Fisher's protected least significant difference
FKBP5 FK506 binding protein 5
Gapdh Glyceraldehyde 3-phosphate dehydrogenase
GC Glucocorticoid

xiii
HFD High-fat diet
HPA-axis Hypothalamic-pituitary-adrenal axis
IL-1β Interleukin-1 beta
LPS Lipopolysaccharide
LUMA Luminometric methylation assay
MAPK Mitogen-activated protein kinase
PI-3K Phosphoinositide 3-kinase
OD Optical density
PND Postnatal day
qRT-PCR/qPCR Quantitative real-time reverse transcriptase-polymerase chain reaction
rRNA Ribosomal RNA
Tet Ten-eleven translocation methylcytosine dioxygenase
TLR4 Toll-like receptor 4
TNF-α Tumor necrosis factor alpha
Ubc Ubiquitin C
Ywhaz 14-3-3 protein zeta/delta
Refer to Table 2 (pg.32) for genes analyzed with qPCR.

1
Chapter 1
1 Introduction
There is accumulating literature on the psychological effects that maternal obesity and
overnutrition, the predominant intake of high-fat foods, incurs on offspring of mammals,
including humans. At least two prevailing explanations exist to account for these effects: 1.
Differences in development of the hypothalamic-pituitary-adrenal (HPA)-axis during pre- and
post-(peri)natal periods of life, and 2. Inflammatory changes in the brain resulting from perinatal
exposure to saturated high fat diets (HFD) - the most commonly used paradigm to drive
overnutrition. These systems are not mutually exclusive: The HPA-axis regulates inflammation
through glucocorticoid and mineralocorticoid receptors (GR/MR), and cytokines, mediators of
inflammation, also regulate the HPA-axis. However, both can affect mental health through
separate mechanisms. Together, with the detrimental effects that overexposure to saturated fats
may have on brain development, makes offspring perinatally exposed manifest unique mental
and behavioral profiles during their lifespan compared to other early-life stressors or diet
manipulations. Engrained defects in activity of epigenetic regulators due to perinatal stress are
known to affect both systems, as they determine the extent that HPA-axis- and inflammatory-
related genes are transcribed. What is not clear, is how these genes may become differentially
regulated when the HPA-axis is activated by stressors experienced during adulthood. In this
study, we studied the GR/MR-mediated inflammatory pathway in limbic brain areas - regions
where the HPA-axis stress response is (dys)regulated by GR/MR. Here, we expected to see local
alterations in transcript abundance as a consequence of perinatal HFD exposure that would
indicate amplified stress and sickness-induced anxious and depressive-like behaviors, as well as

2
cognitive changes in rodents. Further, we expected to see diet-related differences in the activity
of epigenetic mediators, as well as on the regions of DNA that they target.
We will begin by reviewing literature that provides context for our findings - starting
with a brief overview regarding the societal impacts of maternal obesity and overnutrition.
Pertinent to the thesis, the remainder will be focused on the effects on anxiety- and depressive-
like behavioral responses in rodents, and how long-lasting alterations on the HPA-axis and
neuroinflammatory state could predispose individuals to these responses.
1.1 Effects of Maternal Obesity and Overnutrition on Offspring Physical and Mental
Health
68% of adults in the United States are now overweight or obese, including many women of
childbearing age (Flegal et al., 2010; Sullivan et al., 2010). Maternal obesity incurs health risks
for both the mother and their children, some of which manifest later in life. Indeed,
epidemiological studies often suggest that exposure to maternal obesity during early
development is a risk factor for cardiovascular disease, diabetes, and metabolic syndrome
(Godfrey Gluckman, and Hanson, 2010). Rodent and human studies further show heightened
incidences of eating disorders, including offspring preferences for fat-rich foods (Ong and
Muhlhausler, 2011; Rivera et al., 2015). Likely as a consequence, in addition the permanent
changes in metabolism, children of obese women tend to exhibit higher body mass index (BMI)
and body fat percentage (Simmons, 2008; Freeman, 2010). This suggests that the health risks can
be perpetuated through successive generations, calling into attention the need for more biological
research on how a mother's medical condition and dietary intake impacts development.

3
Studies in humans further suggest that offspring of obese mothers, and those that
consume high fat diets and drive weight gain during pregnancy, show an increased risk for
psychiatric ailments including autism, attention-deficit/hyperactivity disorder, schizophrenia,
cognitive impairments, anxiety, and depression (Sullivan et al., 2015; Krakowiak et al., 2012;
Grissom and Reyes, 2013; Rivera et al., 2015). The increased risk is in part a consequence of the
tendency for maternal obesity and weight gain to impair placental function and development, and
result in low or high birth weights, because the latter alone also poses risk for these diseases
(Grissom and Reyes, 2013; Rivera et al., 2015). In at least three studies, links between maternal
obesity and the development of anxiety and depression in children and adolescents have been
demonstrated: High maternal pre-pregnancy BMI has predicted reports of disrupted emotions in
children, such as fear and sadness, and increased internalizing behaviors which associate with
depression (Rodriguez, 2010; Van Lieshout et al., 2013). Another study correlated maternal
obesity with high or low birth weight, which coincided with the development of anxiety and
depression in adolescents (Coleman et al., 2012).
Several studies modelling maternal obesity with non-human primates as well as rodents,
through overnutrition with high fat diets (HFD), have also shown increased anxiety-like and fear
responses in the presence of novelty in adulthood during a variety of behavioral tests, which
frequently depended on the sex of the animals (Sullivan et al., 2010; Bilbo and Tsang, 2010;
Peleg-Raibstein, Luca, and Wolfrum, 2012; Sasaki et al., 2013). Furthermore, depressive-like
behavior has been reported in two studies with male offspring, where less time was spent
swimming and climbing in a Porsolt Forced Swim test (Can et al., 2012; Giriko et al., 2013).
These psychiatric illnesses in humans, and affect-related behavioral responses in rodents
are likely caused by malfunctions in at least two particular systems: The hypothalamic-pituitary-

4
adrenal (HPA)-axis, and inflammation of the central nervous system (CNS). Below, we describe
the comprehensive literature available on perinatal stressors in general, and what is known with
regards to maternal overnutrition, which we will establish as another type of 'stress'.
1.2 Perinatal Programming of the HPA-axis: Effect on Mental Health
Apart from maternal obesity or overnutrition, each of the psychiatric or behavioral
alterations have often been suggested to be due least in part by exposures to a variety of stressful
experiences which lead to abnormal brain development during the perinatal period. This includes
childhood maltreatment and trauma (Teicher and Samson, 2013; Shapero et al., 2014; Varese et
al., 2012) and maternal prenatal anxiety (O'Connor et al., 2002; van den Bergh et al., 2006), and
presence of stressors such as viral infection and adverse life events (Schmitt et al., 2014;
Markham and Koenig, 2011). Such experiences have been extensively shown to permanently
modify the activity of the developing hypothalamic-pituitary-adrenal (HPA) axis, which is a
major component of the neuroendocrine system that mediates response to stress (van Winkel et
al., 2008; Heim et al., 2000; MacKenzie et al., 2007; Weinstock, 2008). The HPA-axis regulates
processes including digestion, metabolism, behaviour, and the immune system according to
magnitude of perceived threat in the environment, in part by controlling the secretion of the
glucocorticoid, cortisol (corticosterone, in rodents) (Smith and Vale, 2006). Among other
upstream hormones such as corticotropin-releasing hormone (CRH), cortisol is circulated
throughout the body to signal physiological changes in tissues (Smith and Vale, 2006). For
instance, it increases levels of glucose in the blood to prepare for metabolic demands during
intense exercise (Robson et al., 1999).

5
Changes in gene expression of the receptors that bind cortisol, glucocorticoid receptors
(GRs) and mineralocorticoid receptors (MR), have often been reported following exposures to
pre and/or postnatal stress. In particular, the changes occur within the amygdala and
hippocampus, where both receptors are abundant, and specifically serve the purpose of
regulating the HPA-axis (Welberg and Seckl, 2000; Brunton, 2010; McGowan et al., 2009;
Weinstock, 2005). These brain regions are a part of the limbic system which regulates processes
including memory, emotion, behavior, and motivation. In the amygdala, the receptors are
involved in activating the HPA-axis, while negative feedback is initiated by receptors in the
hippocampus. Both are accomplished by enhancing or reducing the secretion of cortisol
respectively (Smith and Vale, 2006; Anacker et al., 2011; Qin et al., 2003). Since GR has a lower
affinity to cortisol than MR, it is only active during stress responses where levels increase, and
MR is saturated. On the other hand, MR is constitutively active (Korte, 2001). (Figure 1)
summarizes the relationships between each component of the HPA-axis.

6
Figure 1. Summary of the HPA-axis from Smith and Vale (2006). Neurons in the amygdala are
activated following exposures to stress. The signal is passed through projections innervating the
paraventricular nucleus of the hypothalamus, which releases corticotropin-releasing
factor/hormone (CRH). Receptors in the pituitary gland bind CRH, inducing the release of
adrenocorticotropic hormone (ACTH) into the bloodstream. The adrenal cortex binds ACTH,
causing the synthesis and release of cortisol/corticosterone (CORT), which then binds
glucocorticoid receptors (GR) and mineralocorticoid receptors (MR) to carry out its systemic
metabolic, immune, behavioural, and cardiovascular functions. GR and MR in the hippocampus
bind cortisol to reduce the secretion of CORT upstream, thus gradually terminating the stress
response. Under high magnitude of stress, persistent circulation of CORT can further stimulate
the amygdala and maintain HPA-axis activity (Makino et al., 2002). Credit for the Diagram:
Hyman (2009).

7
1.2.1 Effects of GR and MR on Anxiety and Depression
A body of work has supported the role of limbic GR and MR in mediating anxiety and
depression, although the effects generally appears to depend on a number of factors, including
sex, brain region, and context. Limbic GR has frequently been connected to anxiety-like
behavior, typically increasing its expression (Welberg et al., 2000; Sarrazin et al., 2009;
Chmielarz et al., 2013; Durairaj and Koilmani, 2014). Its activity has also been shown to be
involved in the development and maintenance of major depression (Ströhle and Holsboer, 2003;
Anacker et al., 2011; Juruena et al., 2013) or depressive-like behavior (Chmielarz et al., 2013).
Several studies have suggested limbic MR to have a protective function against anxiety in
males, in the amygdala (Mitra et al., 2009), hippocampus (Herrero et al., 2006), and forebrain
(Lai et al., 2007; Rozeboom et al., 2007). On the other hand, one study suggests that
hippocampal MR, rather than GR, regulates the expression of anxiety in males (Smythe et al.,
1997). In contrast, a recent study suggests that forebrain MR, a part of the brain that includes the
limbic system, does not affect anxiety-like behavior in female mice (Kanatsou et al., 2015) -
conflicting with the study by (Rozeboom et al., 2007) which suggested that MR was also
protective for females.
A protective role of limbic MR also seems to exist for depression. In the 1990s, many
studies have shown that treatment of rats with antidepressant drugs increases the gene expression
and binding capacity of MR in limbic brain areas, including the hippocampus (Reul et al., 2000).
Hippocampal MR, but not GR has also been found decreased in patients with major depression
(Medina et al., 2013). Furthermore, a common haplotype of MR has been discovered to protect
against depression in women (Klok et al., 2011). Malfunctional activity of MR on HPA-axis

8
regulation have also been found in depressed patients (Juruena et al., 2013). In contrast, one
study suggests that patients with major depression maintain high functional activity of the MR
system (Young et al., 2003).
1.2.2 Overexposure to GCs alters GR and MR expression
Overall, limbic GR appears to increase anxiety and depressive behaviors, while limbic MR tends
to be protective. At least one way that altered gene expression of the receptors specifically occurs
during development is through excess fetal exposure to maternal or synthetic glucocorticoids
(GCs) (Welberg et al., 2000; Weinstock, 2005; Levitt et al., 1996), which has been shown to
occur in reaction to maternal prenatal anxiety (O'Donnell et al., 2011). More specifically,
increased exposure to GCs apparently increases mRNA levels of CRH in the amygdala, which
when individually administered during the last week of gestation, causes reductions in levels of
GR and MR in the hippocampus (Welberg et al., 2001). There is evidence to suggest that
cytokines activated by maternal overnutrition also results in fetal overexposure to GCs, which
will be discussed in more detail in Section 1.4 (Bellisario et al., 2015; Kossintseva et al., 2006).
This, with the fact that cytokines activate and regulate the HPA-axis (Kronfol, 2003), makes
maternal overnutrition another form of perinatal stress. Therefore, perinatal stressors each cause
similar mental health outcomes, in part because of how they increase fetal exposure to GCs.
1.3 Mediation of CORT- and LPS-induced Inflammatory Changes Through GR and MR
In addition to regulating the HPA-axis, GR mediates the expression of pro- and anti-
inflammatory molecules depending on the level and sensitivity to CORT - thereby contributing
to the role of the HPA-axis in controlling immune responses. Some studies also suggest the
involvement of MR, where it seems to potentiate inflammation in peripheral and central tissue

9
(Duan et al., 2013; Changtong et al., 2012). There is extensive evidence to indicate that central
and peripheral inflammation can result in psychiatric illnesses and anxiety- and depressive-like
behaviors, which is reviewed on Section 1.4. Therefore, one way that GR and MR modulates
anxiety and depression, among others, is through modifications of inflammatory state.
1.3.1 The General Immunosuppressive Action of GCs.
The involvement of GR in mediating immune responses has been well-documented (Sorrells et
al., 2009; Rhen and Cidlowski, 2005) . In most instances, it serves to suppress the activity of a
key pro-inflammatory transcription factor, Nuclear Factor Kappa Light Chain Enhancer of
Activated B cells (NFkB), upon recognition of an immune stressor. Classically,
lipopolysaccharide (LPS), a component in the cell membranes of gram-negative bacteria, is used
to activate NFkB. It binds to toll-like receptor 4 (TLR4) on the host cell membrane, and signals
Mitogen-activated protein kinase/Phosphoinositide 3-kinase (MAPK/PI-3K) pathways to activate
NFkB. It does this by causing Nuclear factor of kappa light polypeptide gene enhancer in B-
cells inhibitor, alpha (IkBa) to dissociate from NFkB, which is an anti-inflammatory protein that
inhibits its activity. NFkB is then capable of translocating to the nucleus to activate the
transcription of pro-inflammatory cytokines necessary for initiating an inflammatory response,
one of which is Interleukin-6 (IL-6) (Libermann and Baltimore, 1990).
Over time, the GC CORT is secreted, dissociates from corticosteroid-binding globulin
(CBG), and enters the cell. In the cytoplasm, it can either bind to MR, GR, or 11β-HSD, which
will degrade CORT. If it binds and activates GR, this causes the release of co-chaperones from
GR, including heat shock proteins 70 and 90, and FK506 binding protein 5 (FKBP5), while
expression of the latter is induced by GR over time to desensitize receptors to CORT (Binder,

10
2009). Activated GR forms a dimerized complex with CORT and translocates to the nucleus,
where it activates the expression of anti-inflammatory molecules, including IkBa, and MAP
kinase phosphatase-1 (MKP-1), which is a key molecule that is necessary to ameliorate the
immune response. The molecules translocate to the nucleus, where IkBa rebinds NFkB, and
MKP-1 inhibits the MAPK pathway. This mechanism of action is referred to as transactivation.
GR additionally suppresses inflammation by preventing NFkB from binding to its consensus
sites, kB RE. This indirect mechanism of action is known as transrepression. NFkB also begins
to activate the expression of anti-inflammatory cytokines such as Interleukin-10 (IL-10), which
inhibits NFkB (Saraiva and O'Garra, 2010; Driessler et al., 2004). A simplified diagram of this
process is illustrated in Figure 2.

11
Figure 2. Simplified diagram of the general CORT/LPS pathway. In the presence of LPS,
transcription factor NFkB is activated, allowing it to translocate to the nucleus to induce gene
expression of IL-6 and other pro-inflammatory cytokines. Later in the immune response, CORT
is secreted, and binds GR in the cytoplasm. GR is activated, and inhibits NFkB activity through
transactivation of anti-inflammatory molecules, IkBa and MKP-1, and direct transrepression of
NFkB. NFkB also increases expression of anti-inflammatory cytokine, IL-10 over time.
It is important to note, however, that basal CORT levels are needed to mount the initial
immune response against LPS, and that secretion of CORT stimulates the immune response to
acute, low-to-moderate physiological stress. This could be from the action of MR, but has not
been tested extensively. One study has indicated that in microglia, the resident immune cells in
the brain, MR potentiates the activation of NFkB, and expression of pro-inflammatory cytokines,
IL-6 and TNF-α, after treatments with low-to-moderate concentrations of CORT - supporting
this idea (Changtong et al., 2012). Therefore, the effect of GCs is only immunosuppressive when
it is: (1) secreted/administered after LPS, and (2) when they are administered chronically, or with
acute high-doses. ‘Acute’ refers to single-time stressors or injections of CORT, while ‘chronic’
refers to repeated stressors or injections given over a period of weeks or months, In fact, if either
acute or chronic CORT is administered prior to LPS, it facilitates the cell's pro-inflammatory
response. This also holds true in the central nervous system (CNS), but not for all regions of the
brain (Sorrells et al., 2009).
1.3.2 Inflammatory actions of GCs in the Central Nervous System (CNS).
In the CNS, acute stressors also stimulate inflammation, in one way by increasing NFkB activity.
In addition, higher basal CORT levels are associated with a greater accumulation of pro-

12
inflammatory mediators even in the absence of stress - likely via MR. The LPS-induced pro-
inflammatory actions of GCs in the CNS are known to at least in part be mediated by GR. In
spite of reductions in protein levels, GR is necessary to increase LPS-induced NFkB activation in
the hippocampus and cortex - regions abundant in GR, but not in the hypothalamus after acute or
chronic doses of CORT (Frank et al., 2009; Munhoz et al., 2006; de Pablos et al., 2006). As in
the periphery, CORT suppresses the immune response if given after LPS. One way that CORT is
known to mediate pro-inflammatory responses in the CNS is through GR-mediated increases in
proliferation and activation of microglia, the resident macrophage-like immune cells of the brain
(Nair and Bonneau, 2006; Frank et al., 2006), and another is through decreases in expression of
MKP-1 and IkBa (Munhoz et al., 2010). The reason GCs can sometimes activate NFkB in the
CNS, and not other times, is because it is protective in neurons (e.g. when facilitating
hippocampal synaptic plasticity; Albensi and Mattson, 2000), while being damaging to
microglia.
It must be noted, however, that GCs are not always pro-inflammatory in GR-abundant
brain regions. Reductions in NFkB signaling have also been found in the mouse hippocampus
following exposures to chronic psychosocial stress (Feldker et al., 2006). Thus, chronic stress
seems to be immunosuppressive in the CNS in the absence of an immune challenge, as in the
periphery. Acute, high-doses of CORT could also mimic the effects of chronic stress, as
observed by Mitra and Sapolsky (2008). In their study, they observed morphological changes
and inductions of anxiety of similar magnitude to those from chronically stressed male rats.
Similar indications have been made by Dhabhar (2000). Below, we begin to describe the
psychiatric and behavioral effects that microglial activity and inflammation has on the human
and rodent brain – with and without the involvement of GR and MR.

13
1.4. Neuroinflammation, GR, and Mental Disorders: Implications for Maternal
Overnutrition
When an individual or animal contracts a sickness, or is under high stress, what often occurs is
the development of depressive-like symptoms, such as anhedonia (reduced ability to experience
pleasure), reductions in food and water intake, reduced exploration, social, and physical activity.
These behavioral changes can be replicated by injections with pro-inflammatory cytokines,
indicating that it is not the pathogen itself, but the host immune response that causes these
symptoms (Najjar et al., 2013). Beginning in the periphery, many autoimmune illnesses and
medical conditions associated with chronic inflammation, such as systemic lupus erythematosus,
sepsis, obesity, diabetes, rheumatoid arthritis, or chronic fatigue syndrome, are often comorbid
with, or increase the risks for depression (Najjar et al., 2013; Zjaja, 2013; Afari and Buchward,
2003). In lupus and sepsis, the psychiatric symptoms may additionally include anxiety and
psychotic disturbances (Postal et al., 2011; Zjaja, 2013). Furthermore, patients with major
depressive disorder can be distinguished from healthy controls by immunological biomarkers in
their serum, including elevated plasma concentrations of pro-inflammatory cytokine, IL-6,
relative to anti-inflammatory cytokine, IL-10 - which indicates the net state of inflammation
(Dhabhar et al., 2009; Fredericks et al., 2009; Voorhees et al., 2013). Severe psychiatric illnesses
such as schizophrenia, have also been associated with increased serum levels of inflammatory
molecules in human birth cohorts (Ratnayake et al., 2013), having prior hospitalization for
serious infection, and having an autoimmune disease (Benros et al., 2011). As previously
mentioned in Section 1.2, perinatal exposures to maternal, or fetal infection can result in the
development of a milieu of psychiatric disorders or affect-related behaviors (Bilbo and Schwarz,

14
2009; Schmitt et al., 2014; Markham and Koenig, 2011), thereby providing evidence of a partial
involvement of cytokines in mediating psychopathology throughout the lifespan.
There are three processes by which circulation of cytokines in the periphery could
proceed to affect the limbic system (i.e. amygdala and hippocampus, which regulate affect and
affect-related behavior) within the CNS. The first is through cytokine receptors that line the
vagus nerve, which extends from the brainstem to multiple organs, including the esophagus,
heart, lungs, and gastrointestinal tract (Berthoud and Neuhuber, 2000). Second, "glymphatic"
vessels have also recently been discovered to provide immune cells from cerebrospinal fluid a
passage within, and out of the brain (Louveau et al., 2015). Third is a well-characterized passage
through the blood, where cytokines pass through the median eminence of the hypothalamus,
where the blood-brain-barrier is relatively weak (Kronfol, 2003). Here, they bind to receptors
including TLR4, and induce the production of soluble mediators that can cross into the brain and
activate microglial cells, which secrete cytokines through pathways including the NFkB system.
1.4.1 How Cytokines in the CNS Promote Psychopathology
The activation of microglia is crucial for the development of sickness behaviors during infection:
Selectively blocking inflammatory activity in the CNS can block these behaviour changes (Maier
and Watkins, 2012). Cytokines in the brain can modulate affect and behavior in several ways.
First, cytokines within the brain are exclusively involved in stimulating noradrenergic and
serotonergic systems, increasing the synthesis of tryptophan, and activating the HPA-axis by
increasing CORT circulation - all of which mediate affect and affect-related behavioral processes
(Kronfol, 2003). Further, since the brain is particularly vulnerable to oxidative stress, which is
provoked by neuroinflammation, the anti-inflammatory action of microglia are important for

15
maintaining the integrity of neural tissue and the blood-brain-barrier - both which are also
reported in psychiatric illnesses (Najjar et al., 2013). Similarly, depression has been suggested to
be caused at least in part by impairments in synaptic and structural plasticity due to elevations in
cytokine levels (Datson et al., 2013). Finally, cytokines also play a crucial role in the brain
during development, as they mediate the migration of neuronal and glial cells, differentiation,
and synaptic maturation (Nawa and Takei, 2006), while microglia are involved in processes such
as angiogenesis, axonal growth, and apoptosis. Thereby, one could imagine how the microglia in
the CNS can become sensitized to repeated or severe exposures to infections, or other forms of
stress that activate GR. During adulthood, this could enable transient sickness behaviors to
manifest as psychiatric illnesses. On the other hand, exposures during the perinatal period could
increase susceptibility to developing these illnesses throughout life.
There is extensive literature on the effects of cytokine expression in the developing
hippocampus during particular time-points, and its resultant positive and negative impacts on
memory, learning, and affect during adulthood, which has been reviewed by Bilbo and Schwarz
(2009). In essence, early immune challenges "prime" microglia to be more constitutively active,
pro-inflammatory, and reactive to stress during adulthood - either impairing or facilitating
performances during learning tasks, for example. Pertaining to maternal overnutrition, we will
now describe how perinatal exposures can alter cytokine profiles in the rodent brain, how this
can cause alterations in HPA-axis activity, and collectively affect behaviour and responses to
stress.

16
1.4.2 Caveats to Consider with Maternal Overnutrition Studies
Before we proceed, it must be noted that studies on maternal overnutrition tend to vary in the
type of diet used, and the duration of exposure. Most incorporate diet manipulations throughout
the perinatal period, and use diets that are high in fat. High fat diet (HFD), then, will be used
interchangeably with overnutrition. The composition of fats can differ between saturated,
unsaturated, and trans fats, and have varying proportions of protein and carbohydrate content.
'Cafeteria' diets are also sometimes used. Through the rest of the thesis, the focus will be on diets
high in saturated fat, the most common fat used to drive overnutrition.
1.4.3 Effects of Perinatal HFD on Inflammation
Potentially, what makes maternal saturated HFD unique compared to other perinatal stressors is
the ability of saturated fats to cross the blood-brain-barrier as well as the placental barrier
(Bolton and Bilbo, 2014). The implications of this process is two-fold: (1) saturated fats will be
the primary fat type integrated into the fetal brain, which is predominantly composed of fats, and
affect development, and (2) saturated (and trans) fats can access and bind to TLR4 within the
brain and dictate the distribution of local cytokines (Milanski et al., 2009). Because, in both
humans and rodents, the proportion of saturated fatty acids taken into the brain peak, then
continue decreasing through perinatal development to favour incorporation of other fat types
(Svennerholm and Vanier, 1978; Bourre et al., 1979), brain development could become restricted
by the deficits of other types of fats included in the diet. In humans, there is also evidence that
the mother's inflammatory state from ingesting saturated fats can be transmitted to the offspring
through releases of pro-inflammatory cytokines, which can cross into the fetal component
(Zaretsky et al. ,2004), as well as through the breast milk during postnatal development and

17
further activate TLR4 (Fagerås et al., 1999). Therefore, much like in the presence of infection,
HFD acts as an early-life 'challenge' that induces neuroinflammation in the developing fetus, and
sets the immunological template to reacting to future infections or stressful events.
In support of this, maternal HFD-exposed rodent offspring display increased circulating
levels of pro-inflammatory markers C-reactive protein (CRP) and IL-6 during birth, which
resemble those apparent after exposures to maternal infection (Williamson et al., 2011). In
humans, the restricted delivery of nutrients to the fetus (placental insufficiency) that often
happens as a result of maternal obesity also tends to be accompanied by pro-inflammatory
cytokine responses in the placenta (Wang et al., 2003; Bartha et al., 2003) – recalling that growth
restriction tends to be accompanied by future psychiatric illnesses (Section 1.1). During
adulthood, rodent models of placental insufficiency exhibit anxiety-like behavior and reduced
expressions of GABA and serotonin receptors in the hippocampus - indicating altered activity of
cytokines within the brain (Mikaelsson et al., 2013). Indeed, another maternal HFD rodent study
found increased brain levels of IL-1β, TNF-α, and microglial activation in female offspring
which associated with increased anxiety and decreased sociability (i.e. depression). Most
interestingly, replacing the maternal diets with standard chow during lactation was able to
alleviate these effects (Kang et al., 2014). Similarly, perinatal HFD males display heightened
levels of IL-6, microglial activation, and oxidative stress in the brain, which associated with
cognitive decline in the Morris water maze spatial learning task (White et al., 2009). In
summary, maternal HFD directly increases neuroinflammation, and indirectly by incurring
placental insufficiency - resulting in anxious- and depressive-like behavior and cognitive deficits
in learning tasks during adulthood.

18
1.4.4 Effects of Perinatal HFD on the HPA-axis as a Consequence of Inflammation.
Pro-inflammatory cytokines secreted due to exposures to saturated fats can also decrease the
activity of 11-β-HSD2, an enzyme that inactivates maternal GCs before it crosses the fetal
component (Kossintseva et al., 2006). This results in increased fetal exposure to CORT, and
should mimic the effects of physiological/psychological stressors on the expression of limbic GR
and MR. The resultant impact on GR and MR would then further impact the activity of microglia
and distribution of cytokines, and drive affect and behavioral abnormalities experienced with or
without stressful life experiences. Hence, saturated fats separately expose offspring to impaired
brain development, GR/MR, and pro-inflammatory cytokine expression during development,
which all have downstream consequences on affect and affect-related behaviors that at times
overlap - making it a distinct from other perinatal stressors.
To our knowledge, the effect of perinatal HFD on the expression of GR and MR in adults
has only been examined in our recent study (Sasaki et al., 2013), although some literature exists
on chronic consumptions of HFD during adulthood - with similar results (Sasaki et al., 2015).
We found that exposure to maternal saturated HFD in rodents increases the basal transcript levels
of GR and/or MR in the amygdala mostly for females (recalling that it activates the HPA-axis),
and such changes are associated with alterations in abundance of target pro- and anti-
inflammatory molecules, including those shown in (Figure 2). In HFD females, subtle decreases
in anti-inflammatory transcript abundance were also apparent in the hippocampus. Coinciding
with the changes in transcripts were enhanced anxiety responses to the open field, elevated plus
maze, and light-dark transition tests. A hyperactive HPA-axis response was also apparent in the
HFD females, such that CORT levels remained elevated above the control chow diet (CHD)
females during the recovery period from restraint stress. This was not observed in the males. Our

19
study further suggested that maternal HFD may preferentially alter GR/MR, and inflammatory
responses in the amygdala compared to the hippocampus, bringing into attention the need for
more studies on maternal HFD in the former limbic region.
Epigenetic processes, which dynamically mediate the expression of genes depending on
cues taken from the environment, is known to be permanently altered by maternal HFD. Systems
affected by perinatal stress-induced changes in epigenetic regulation include those of the HPA-
axis, and those that mediate immune responses. Therefore, alterations of the two systems are in
fact manifestations of underlying differences in epigenetic regulation - making this an additional
layer that has to be considered. We will now discuss how DNA methylation, one means of
epigenetic regulation, causes permanent changes in expression of the related genes in response to
perinatal stressors, including maternal HFD. Pertinent to this thesis, we will also describe the
involvement of epigenetic mechanisms in responding to stress challenges during adulthood. In
essence, some studies indicate that impaired stress responses can occur from exposures to prior
stressors, and that this could be due in part by dysregulated expression of molecules that mediate
epigenetic processes.
1.5 Role of Epigenetic Regulation in Incurring Lifelong Effects of Maternal Overnutrition
To begin, a number of commonly-cited studies examining lifelong effects of postnatal
experiences such as quality of maternal care in rodents, or exposures to childhood abuse and
neglect in humans have found changes in expression of limbic GR and MR to be the result of
alterations in DNA methylation at their promoters (Weaver et al., 2004; Weaver et al., 2007;
McGowan et al., 2009). DNA methylation is an epigenetic mechanism generally indicated to
regulate gene expression by either directly preventing the binding of repressive or activating

20
components of the transcription machinery, or signaling to configure the surrounding chromatin
into making the DNA more or less accessible to the transcription machinery (Jones, 2012;
Fagiolini et al., 2009). Altered promoter methylation of GR has also been observed with prenatal
exposure to maternal depression or chronic variable stress in humans and rodents respectively
(Oberlander et al., 2008; Mueller and Bale, 2008). In the liver, reduced DNA methylation of GR,
and increased expression has been reported following maternal protein-restricted diets during
pregnancy (Lillycrop et al., 2007). Similarly,
in response to increased quality and quantity
of maternal care, the DNA methylation of
IL-10, an anti-inflammatory cytokine that
also regulates the HPA-axis (Smith et al.,
1999), has been found reduced specifically in
the microglia, and related to increases in
expression (Schwarz et al., 2011).
In addition to the alterations in
methylation state resulting from perinatal
stress, the activity and expression of
molecules which regulate DNA methylation
has been suggested to become impaired,
particularly in response to perinatal diet.
McKee and colleagues (2014) have found
that maternal saturated HFD (60% to 20%
protein and carbohydrates) results in reduced DNA methyltransferase (DNMT) enzymatic
Box 1. Regulators of DNA methylation.
DNMTs are the enzymes responsible for
appending methylation marks to DNA.
MeCP2 is a chromatin-associated protein that
binds methylated DNA to sterically block
binding of the transcription machinery, and
interact with histone deacetylases (HDACs)
to alter the chromatin state (Theisen et al.,
2013). The isoforms of DNMTs, maintenance
methyltransferases DNMT1 which sustain
existing methylation, and de novo
methyltransferases DNMT3a and DNMT3b
which add novel methyl groups (Jeltsch and
Jurkowska, 2014), are known to interact with
one another and MeCP2 to carry out their
specific activities (Fatemi et al., 2002;
Hseieh, 2005; Kimura and Shiota, 2003). On
the other hand, Gadd45b and Tet are factors
and enzymes invovled in actively
demethylating DNA (Niehrs and Schäfer,
2012; Kohli and Zhang, 2013).

21
activity in the prefrontal cortex that coincided with decreased interactions of DNMT1 isoforms
with Methyl-CpG-binding Protein 2 (MeCP2) (Box 1). The impaired DNMT activity co-
occurred with global hypomethylation of promoters, and overall increased gene expression.
Unexpectedly, the gene and protein levels of DNMT1 were overexpressed in this study,
suggesting a compensatory mechanism for the reduced function. Other studies indicate
reductions in global and gene-specific DNA methylation, including GR, in response to
nutritional deficits during gestation (Doherty et al., 2000; Lillycrop et al., 2008), associating with
decreased mRNA levels of DNMT1 in newborn (Pham et al., 2003) and adult rodents (Lillycrop
et al., 2007). The other epigenetic mechanisms, histone modifications and non-coding
microRNAs, have also been found to be affected by exposures to nutritional factors during
perinatal development (Canani et al., 2011).
1.6 Role of Epigenetic Regulation in Responding to Stress
1.6.1 Gene-specific Functions
Epigenetic mechanisms are known to play a role in mediating the transcriptional response to
stress. When bound to CORT, activated GR normally regulates transcription by associating with
epigenetic regulators, with one example involving interactions with DNMTs, MeCP2, and
HDAC1 in the repression of hypothalamic CRH. This serves as a means to initiate negative
feedback on the HPA-axis (Sharma et al., 2013). To terminate the stress response, GR is also
downregulated over time in order to reduce sensitivity to CORT (Chiba et al., 2012; Gehring et
al., 1984), and this is also mediated epigenetically. In one mechanism, activated GR binds to its
own negative consensus GR-binding site (nGRE) at a distal regulatory region, then recruits
HDAC3 to the promoter-proximal region of GR to form a repression complex and inhibit

22
transcription (Ramamoorthy and Cidlowski, 2013). Pertaining most to this thesis, MeCP2 is
known to repress the expression of downstream targets in response to CORT and Tumour
Necrosis Factor alpha (TNF-α), an immunological stressor. This includes FKBP5 and IL-6
respectively, in the microglia (Cronk et al., 2015).
1.6.2 Mediation of Global Changes
The involvement of epigenetic processes in responding to GCs is by no means limited to a select
number of genes. Rapid changes in histone marks occur in the hippocampus during responses to
acute stressors such as forced swim and novelty tests, which would result in genome-wise
changes in gene transcription through altered global repatterning of chromatin state (Datson et
al., 2013). During late stages of fetal development, a natural GC surge occurs in order to
facilitate the maturation of several organ systems through genome-wide changes in gene
expression (Crudo et al., 2012). This surge has been observed to be accompanied by global
changes in CpG methylation in a number of peripheral tissues that persist into adulthood (Crudo
et al., 2012). CpG refers to the dinucleotide sequence where DNA methylation most often
occurs, and serves as the binding site for MeCP2 (Jones and Takai, 2001). They are commonly
found in promoters of mammalian genes, and tend to be found unmethylated when the genes are
expressed. The converse is true for those residing within gene coding regions. This forms the
basis for a number of epigenetic techniques developed to approximate the amount of DNA
methylation in particular regions or genomic samples (Suzuki and Adrian, 2008).
1.7 The Effects of Prior Experiences on Responses to Stress
Since early-life stressors such as maternal HFD appear to have a 'priming' effect on epigenetic
regulators such that its enzymatic activity is permanently altered, then its activity and interaction

23
with GR may also be impaired during responses to stress. It is currently unclear how
inflammatory pathways in the brain, thereby affect and behaviour, would be affected by the
presence of a perceived threat after prior stress, because studies that examine its consequences
upon re-induction of stress have generally been scarce. Here, we review the existing literature on
the effects that prior chronic stress has on the HPA-axis activity, inflammation, epigenetic
regulation, and transcriptional response to acute stressors.
1.7.1 Studies inspecting HPA-axis Activity
Previous exposures to chronic social isolation have been reported to lead to decreased secretion
of CORT and increased NFkB signaling in response to acute restraint stress in the liver
(Djordjevic et al., 2010). Prior exposures to a single prolonged stress paradigm have also been
shown to impair the regulation of of GR and MR mRNA expression during recovery to an acute
stressor in the hippocampus (Liberzon et al., 1999): Initially, both downregulate expression, but
MR remains downregulated 1-2 weeks later, while GR reaches higher than prestress levels. On
the other hand, animals that did not display altered negative feedback responses after the initial
prolonged stressor were able to normalize GR/MR levels.
Similarly, there is evidence that maternally separated neonatal rats can maintain adequate
or enhanced HPA-axis activity during baseline, while displaying prolonged activity in response
to acute stressors coinciding with differential GR/MR gene expression in the hippocampus (Ladd
et al., 2004). Comparing between rats with shorter periods of maternal separation, ACTH and
CORT levels were sustained for longer times in response to an airpuff startle stimulus. And, in
response to a dexamethasone suppression test which is used to gauge negative feedback
responses to exogenous CORT, ACTH and CORT levels returned to baseline levels sooner. It

24
was found that hippocampal MR mRNA density had initially been enhanced in these rats, while
hippocampal GR was reduced - explaining the impairments in negative feedback. This study
suggested that some effects of prior chronic stress may only be apparent when GR is activated,
such as if the chronic stressor is of moderate intensity.
In contrast to these psychological stressors, rats exposed to bacterial infections early in
life showed faster recovery from depressive symptoms induced by CORT, and blunted HPA-axis
responses (i.e. reduced serum CORT) during and after the stressor - although the depressive
symptoms were more pronounced (Bilbo et al., 2008). Hence, there is preliminary evidence to
support that perinatal stress affects the responsiveness of the HPA-axis during further activation,
but that the outcome appears to vary according to the initial type of stress. Therefore, it is again
pertinent to note the distinctiveness of maternal saturated HFD from other types of early-life
stressors.
1.7.2 Studies examining Transcriptional and Epigenetic effects
Along similar lines as with Ladd et al (2004), effects of prior chronic stress in the rodent
hippocampus have only been unmasked after GR was activated with acute, high-dose
corticosterone (CORT) injections (Datson et al., 2013). Compared to animals with no previous
exposure to chronic restraint stress, about 60% of genes affected by CORT were differentially
expressed (p < 0.05). This included several genes involved in chromatin structure and epigenetic
processes, which supported that the changes in expression may have been partially causing the
differential transcriptional response to CORT. In this study, genes from the TNF-α/NFkB
signaling pathway were exclusively affected by the presence of prior restraint after administration
of acute CORT - noting that the population isolated was predominantly neuronal cells. The

25
directionality, however, was not specified. Connections to epigenetic regulation have also been
implicated in a recent study using restraint stress and LPS in the stress paradigm. It was
discovered that the gene expression of a number of epigenetic mediators failed to upregulate in
the hypothalamus of maternal HFD adult rats, which showed overall reduced transcriptional
responses in a panel of 26 genes (Grissom et al., 2015).
1.7.3 Other studies with Inflammatory Pathways
To our knowledge, there has only been one study with maternal HFD (saturated and trans fat)
that examined changes in cytokine expression in response to LPS by Bilbo and Tsang (2010).
They discovered that the protein levels of pro-inflammatory cytokine, IL-1β in the rodent adult
hippocampus were strikingly increased both basally, and post-LPS, and this co-occurred with
higher basal and LPS-induced microglial activation - suggesting a chronic priming effect. The
HFD, non-challenged offspring also displayed increased anxiety, but enhanced performance in a
spatial learning task, the Morris Water Maze. This was perhaps due to the elevations of IL-β,
which is crucial for normal learning (Williamson et al., 2011).
1.8 Research Hypotheses
To our knowledge, the majority of research on maternal HFD, including epigenetic studies, has
focused on the consequences on peripheral tissues. These studies also tend to exclude females
offspring due to the effects of the stage of estrus on measurements such as behavior. Moreover,
measurements have typically been taken with unstressed animals, while more studies have begun
to unmask effects of prior chronic stress during GR activation (Ladd et al., 2004; Datson et al.,
2013). Given the prior evidence that genes such as GR and MR would be differentially expressed
(Sasaki et al., 2013), we were interested in examining effects of maternal saturated HFD on the

26
limbic brain areas of both sexes after 'challenges' with different types of stressors: Acute stress
with high-dose CORT, and/or bacterial infection with injections of LPS. In this study, we chose
to focus on differences in transcript levels of components from the established inflammatory
pathway targeted by GR and MR, as outlined on Section 1.3 because of our interest in
psychiatric illnesses, and the fact that we had seen diet-related differences in our previous study
(Sasaki et al., 2013). We expected diet-related differences to exist in the amygdala and
hippocampus, given that they are the regions where the HPA-axis is (dys)regulated by GR/MR,
and where local inflammatory changes would result in anxiety/depressive behaviors.
We hypothesized that:
1) Perinatal HFD-exposed animals will display aberrant transcriptional inflammatory
responses to each challenge. In response to the acute, high-dose of CORT used, we
predicted its immunosuppressive properties would become enhanced due to GR/MR
being permanently conditioned by the prior exposure to chronic stress. In contrast, we
expect animals to show heightened pro-inflammatory responses to LPS, as observed by
Bilbo and Tsang (2010), due to the perinatally sensitized state of existing microglia. With
simultaneous injections of CORT/LPS, we then expected that the perinatal overexposure
to CORT would result in similar pro-inflammatory changes as when CORT is given
hours before LPS. If this is the case, we should observe increased transcript abundance of
CD11b, a marker for microglia.
2) Given the study by Bilbo and colleagues (2008), and the fact that anxious and depressive
behaviors are already apparent in non-stressed animals, we expected perinatal HFD-
exposed rats to also carry a higher propensity for reacting to stress with anxious- and
depressive-behavior than CHD rats. Therefore, we predicted that the expression of genes

27
associated with anxiety and depression would become elevated in the HFD groups. This
includes CRH in the amygdala, associated with anxiety (McGill et al., 2006; Samaco et
al., 2012); and MKP-1 and IL-6/IL-10, associated with depression (Duric et al. 2010,
Dhabhar et al., 2009; Fredericks et al., 2009; Voorhees et al., 2013).
3) We hypothesized that DNMT transcript levels and activity will be altered in perinatal
HFD-exposed animals to partially explain the differential transcript responses we might
observe. Since the maternal HFD study by McKee et al. (2014) found reduced DNMT
activity in non-stressed animals, with apparent compensatory increases in DNMT1
expression, we also expect to see this with our HFD groups before and during the stress
challenges.
4) If DNMT activity is reduced, then global or gene-specific differences in proportion of
DNA methylation should also accompany the transcript differences. Given that DNMTs
methylate DNA, we expected global CpG methylation to overall be reduced in limbic
brain areas. In response to CORT, which at least up- and down-regulates an even number
of genes in the hippocampus (Datson et al., 2013), the net outcome of reduced DNMT
activity should be further reductions in global CpG methylation. .

28
Chapter 2
2 Materials & Methods
2.1 Animals
Adult male and female Long Evans rats (7 week) used were obtained from Charles River Canada
(St. Constant, QC). Rats were housed in same-sex pairs, and maintained on a 12:12-h light-dark
cycle (lights on from 7:00 am - 7:00 pm) with ad libitum access to food and water. Experimental
protocols were approved by the Local Animal Care Committee at the University of Toronto,
Scarborough, and were in accordance with the guidelines of the Canadian Council on Animal
Care.
2.2 Diets
Female breeders were placed on one of two diets: A HFD containing 60% saturated fat, 20%
carbohydrate, and 20% protein (n = 15), or control house chow diet (CHD) with 58%
carbohydrate, 28.5% protein, 13.5% saturated fat (n = 14). The 5.24-kcal/g HFD was obtained
from Research Diets, Inc. (New Brunswick, NJ: cat. no. D12492), and the 3.02-kcal/g CHD was
obtained from Purina Lab Diets (St. Louis, MO: cat. no. 5001). Similar formulations have
previously been compared to examine diet-induced obesity (El-Haschimi et al., 2000; de Souza
et al., 2005; Dunn and Bale, 2009; Tamashiro et al., 2009; Purcell et al., 2011). The females
remained on the diet for 4 weeks prior to mating, and throughout pregnancy and lactation. Upon
weaning at Postnatal day (PND) 21, offspring were maintained on CHD throughout adulthood. A
timeline for the diet regimen is illustrated in Figure 3.

29
Figure 3. Perinatal-HFD regimen used in this study. The animals were injected with CORT,
LPS, or CORT/LPS, and brain regions were collected 3 hours later for downstream applications
such as qPCR for gene expression i.e. transcript level analysis.
2.3 Subjects and general procedures
Female breeders were housed individually after mating, with no significant differences in litter
size or sex ratio among the diet groups. Except for weighing during weekly cage changes,
offspring remained undisturbed until weaning at PND 21, when they were housed in same-sex
pairs. Prior to the drug challenges, body weights of the offspring were measured (n = 167).
2.4 Drug challenges
During PND 90, the adult offspring were injected with either an acute, subcutaneous dose of
CORT dissolved in propylene glycol (10 mg/kg, Sigma-Aldrich, cat. no. 27840), a low
intraperitoneal dose of LPS from Escherichia coli dissolved in saline (25 ug/kg, Sigma-Aldrich,
cat no. L2630), or a simultaneous high-dose CORT/LPS injection (10 mg/kg, 50 ug/kg) (n = 6
per diet and sex). To determine diet x drug effects, a subset evenly divided by sex and diet were
left non-injected (n = 24). The dose of CORT has been shown to result in CORT levels
comparable to that of several hours of high physiological stress (Stein-Behrens et al., 1994), and
is sufficient to induce anxiety and dendritic hypertrophy on the amygdala of similar magnitude to

30
those from chronic treatments (Mitra and Sapolsky, 2008). The 25yg/kg dose used for LPS has
been used in an extensive body of studies on neonatal infection cited in Williamson et al. (2011),
where cognitive impairments and altered cytokine profiles were observed in the hippocampus
after exposures to further stress and immune challenges. For the 50 ug/kg dose of LPS, enhanced
hippocampal microglial activation and pro-inflammatory cytokine expression was previously
observed in maternal HFD-exposed groups (Bilbo and Tsang, 2010). All of the conditions used
in this project are outlined in Table 1.
Table 1. Number of adult offspring in each challenge condition. Each cell includes an even
number of male and female rats (CORT: mg/kg, LPS: ug/kg). Control offspring were left
undisturbed until sacrifice. The conditions that will be discussed are highlighted.
CORT (s.c.) LPS (i.p.) CORT+ LPS
Dosage Control 5 10 Control 25 50 Control 5, 25 10, 50
CHD 4 12 12 4 12 12 4 12 12
HFD 4 12 12 4 12 12 4 12 12
Total 56 56 56
2.5 Brain tissue preparation
The adult offspring were sacrificed by CO2 inhalation followed by decapitation. Brains were
rapidly dissected, flash-frozen with isopentane, and stored at -80ºC. Using stereotaxic
coordinates, the entire dorsal hippocampus and amygdala was extracted under a Leica CM3050
cryostat. Tissues were homogenized with TRIzol Reagent (Invitrogen, Carlsbad, CA, USA), with

31
1 ml per 50-100 mg of tissue, and RNA, DNA, and proteins were separately extracted. RNA
were first extracted with RNeasy Mini Plus Kits (Qiagen, Valencia, CA, USA), then DNA and
proteins were simultaneously extracted with a manufacturer's protocol for TRIzol isolations
optimized for purity and concentration. Quantification/quality assessments were performed with
a Nanodrop ND-2000C spectrophotometer according to the manufacturers' protocol. The DC
Protein Assay (Bio-Rad, Hercules, CA, USA) was used for the protein quantifications.
2.6 Analyses of transcript abundance by quantitative real-time reverse transcriptase-polymerase
chain reaction (qRT-PCR)
Transcript abundance of genes involved in glucocorticoid signaling, mediating inflammation,
and epigenetic regulation were quantified by StepOne Plus real-time PCR using Fast SYBR
Green PCR master mix (Applied Biosystems, Life Technologies, Carlsbad, CA, USA) and by
relative normalization against the levels of each of five housekeeping genes to compare between
groups (Gapdh, ActB, Ubc, Ywhaz, and rRNA). Table 2 lists the analyzed genes with brief
descriptions. geNorm software selected Gapdh, Ywhaz, and ActB as the three most stably
expressed between the diet and drug conditions. ActB was selected for the analysis due to its
high similarities with results using the geometric mean of the three genes and its common usage
in other studies of HFD. To enable comparisons between runs, transcript abundance of each gene
was quantified by the standard curve method, where the same cDNA mixture of known
concentrations was run with every sample per tissue. All the reactions were performed in
triplicate.

32
Table 2. List of genes analyzed for transcript abundance with qPCR.
Functional Group Gene Abbrev. Description
Glucocorticoid
(GC) Signaling
Glucocorticoid
Receptor
GR Binds GCs to regulate genes involved in stress-induced
HPA-axis activity (Heegde et al., 2015)
Mineralocorticoid
Receptor
MR Binds GCs to regulate genes involved in basal- and
stress-induced HPA-axis activity (Heegde et al., 2015)
Corticotropin-
releasing hormone
CRH Stimulates secretion of ACTH in the pituitary to produce
cortisol/corticosterone (Smith and Vale, 2006)
Pro-inflammatory Cluster of
differentiation
molecule 11b
CD11b Microglial marker (Bilbo and Tsang, 2010).
Nuclear Factor
Kappa Light Chain
Enhancer of
Activated B cells
NFkB Key inflammatory transcription factor (Nelson et al.,
2003).
Interleukin-6 IL-6 Pro-inflammatory cytokine regulated by NFkB
(Libermann and Baltimore, 1990)
Interleukin-
6/Interleukin-10
IL-6/IL-
10
Marker of inflammation and depression (Dhabhar et al.,
2009; Fredericks et al., 2009; Voorhees et al., 2013).
Anti-
inflammatory
Insulin-like growth
factor 1
IGF-1 Neuroprotective; behaves as anti-inflammatory cytokine
in the brain (Mangiola et al., 2015; Bake et al., 2014)
Nuclear factor of
kappa light
polypeptide gene
enhancer in B-cells
inhibitor, alpha
IkBa Inhibits NFkB until, and during activation (Nelson et al.,
2003).
MAP kinase
phosphatase-1
MKP-1 Crucial for resolving inflammation (Vandevyver et al.,
2012); marker for depression (Duric et al. 2010)
Interleukin-10 IL-10 Anti-inflammatory cytokine against, and regulated by,
NFkB (Driessler et al., 2004; Saraiva and O'Garra, 2010).
Epigenetic
regulators
DNA
methyltransferase
1
DNMT1 Maintenance methyltransferase (Jeltsch and Jurkowska,
2014)
DNA
methyltransferase
3a
DNMT3
a
de novo methytransferase (Jeltsch and Jurkowska, 2014)
DNA
methyltransferase
3b
DNMT3
b
de novo methytransferase (Jeltsch and Jurkowska, 2014)
Methyl CpG
binding protein 2
MeCP2 Binds methylated CpG sequences to regulate
transcription (Theisen et al., 2013)
Growth Arrest And
DNA-Damage-
Inducible, Beta
Gadd45
b
Stress and activity-induced DNA demethylation to
enable neurotypical development (Kigar et al., 2015)

33
2.7 Primer specifications
The primers used for qRT-PCR were purchased from Qiagen, or designed using sequence
information from GenBank at the National Center for Biotechnology Information (NCBI;
www.ncbi.nlm.nih.gov), as shown on (Table 3).
Table 3. Primer sequences and catalogue numbers for genes analyzed with qPCR.
Gene Sequence
Forward Primer Reverse Primer
Ubc 5'-CAC CAA GAA GGT CAA ACA GGA A-3' 5'-AAG ACA CCT CCC CAT CAA ACC-3'
Gapdh 5'-ACA TCA AAT GGG GTG ATG CT-3' 5'-GTG GTT CAC ACC CAT CAC AA-3'
ActB 5'-TTT GAG ACC TTC AAC ACC CC-3' 5'-ATA GCT CTT CTC CAG GGA GG-3'
18S rRNA 5'-ATG GTA GTC GCC GTG CCT A-3' 5'-CTG CTG CCT TCC TTG GAT G-3'
Ywhaz 5'-TTG AGC AGA AGA CGG AAG GT-3' 5'-GAA GCA TTG GGG ATC AAG AA-3'
MR 5'-GGC AGC TGC AAA GTC TTC TT-3' 5'-GAC AGT TCT TTC GCC GAA TC-3'
CRH 5'-CTC TCT GGA TCT CAC CTT CCA C-3' 5'-CTA AAT GCA GAA TCG TTT TGG C-3'
CD11b 5'-GAA GCC TTG GCG TGT GAT AG-3' 5'-GAG CAG TTT GTT CCC AAG GG-3'
IGF-1 5'-GCT CTT CAG TTC GTG TGT GG-3' 5'-TGA GTC TTG GGC ATG TCA GT-3'
IkBa 5'-CAG GAT TCT GCA GGT CCA CT-3' 5'-TGG AGC ACT TGG TGA CTT TG-3'
MKP-1 5'-GCT CCA CTC AAG TCT TCT TCC TCC AA-3' 5'-TGG ACT GTT TGC TGC ACA GCT
CAG-3'
IL-10 5'-AGA AGG ACC AGC TGG ACA ACA C-3' 5'-GTC GCA GCT GTA TCC AGA GGG-3'
DNMT1 5'-ACC TAC CAC GCC GAC AT-3' 5'-AGG TCC TCT CCG TAC TCC A-3'
DNMT3a 5'-ACG CCA AAG AAG TGT CTG CT-3' 5'-CTT TGC CCT GCT TTA TGG AG-3'
DNMT3b 5'-GAT GTG ACA CCT AAG AGC AGC AGT AC-3' 5'-CAA ACT CCT TGT CAT CCT GAT ACT
CA-3'
MeCP2 5'-CAA ACA GCG ACG TTC CAT CA-3' 5'-TGT TTA AGC TTT CGC GTC CAA-3'
Gadd45b 5'-GCT GGC CAT AGA CGA AGA AG-3' 5'-GCC TGA TAC CCT GAC GAT GT-3'
GR RT2 quantitative-PCR (qPCR) Primer Assay (PPR52805B, SA biosciences, Qiagen)
NFkB RT2 qPCR Primer Assay (PPR42746A, SA biosciences, Qiagen)
IL-6 RT2 quantitative-PCR (qPCR) Primer Assay (PPR06483B, SA biosciences, Qiagen, Valencia,
CA, USA)

34
2.8 Measurement of DNMT enzyme activity and abundance in the amygdala
To quantify the total enzymatic activity of DNMTs, the EpiQuikTM DNMT Activity/Inhibition
Assay Ultra Kit (Fluorometric) was used (Epigentek, Farmingdale, NY, USA). In triplicates, 5-
10 ug of proteins were added into wells stably coated with an oligonucleotide sequence optimally
designed to be methylated by all active DNMT isoforms in the presence of the substrate, S-
Adenosyl methionine. Anti-5-methylcytosine was added to capture the amount of methylated
DNA, and fluorescence signals generated from a dye-bound secondary antibody were detected to
calculate total enzymatic activity in units of RFU/h/mg with the formula, DNMT Activity =
(Sample RFU - Blank RFU)/(Protein Amount (µg) x hours incubation) x 1000. Using EpiQuikTM
DNMT1 Assay Kit (Epigentek, Farmingdale, NY, USA), a similar principle was used to quantify
the amount of DNMT1 protein colorimetrically using anti-DNMT1 in place of 5mC, with the
formula DNMT1 (ng/mg protein) = (Sample OD - Blank OD)/(Slope x Protein Amount (µg)) x
1000. OD = Optical density.
2.9 Global CpG methylation analysis in the amygdala with Luminometric methylation assay
(LUMA)
LUMA is a restriction enzyme-based method of estimating the global proportion of CpG
methylation using the isoschizomers, HpaII and MspI which both target CCGG sequences, with
the difference being that HpaII cannot cleave the sequence when the internal cytosine is
methylated. Methylation most often occurs on cytosines of CpG dinucleotides (Jones and Takai,
2001), hence it would be an approximate representation of the total amount of methylation
present in a sample of genomic DNA. EcoRI is added to each reaction to control for the
efficiency of each digest, allowing for the cleavage ratios to only capture the methylation

35
difference. A combination of protocols from Karimi et al.(2006)(Original), Pilsner et al.(2010),
and Crudo et al. (2012) were used for this procedure. Briefly, 300 ng of genomic DNA was
added to two duplicate reactions containing 5U each of Hpa/II EcoRI or MspI/EcoRI in Tango
Buffer. Distilled water was included for a total volume of 20 ul. The samples were incubated at
37ºC for 4 h, then heat-inactivated at 80ºC for 20 min. 20 ul of Pyrosequencing annealing buffer
(Qiagen) was added to each reaction, and the samples were transferred onto 96-well
Pyrosequencing plates. The nucleotide dispensation order from Pilsner et al. (2010) was used,
and Pyrosequencing was conducted with PyroMark MD (Qiagen). Peak heights of nucleotide
incorporation were used to calculate % genomic DNA methylation using the formula: 1-
[(HpaII(First G after addition of C)/EcoRI(First T after addition of A))/MspI(G)/EcoRI(T))] x
100, where CG and AT represent the overhangs left behind after each digest.
2.10 Statistical analyses
The weights of pups were analyzed by 2 x 2 day-by-diet repeated measures analysis of variance
(ANOVA), and adults, by 2 x 2 diet by sex ANOVA on SPSS (IBM Corporation, Armonk, NY,
USA). Gene expression, LUMA, and DNMT data were analyzed by 2 x 2 diet by drug ANOVA
on Statview (SAS institute, Cary NC) for each sex. All significant ANOVA tests were followed
by Fisher's PLSD post hoc comparisons. Spearman's Correlations for DNMT activity were
performed on SPSS. IL-6/IL-10 ratios were normalized against non-injected transcript levels in
order to compare between each drug challenge. This was done by subtracting values from the
mean expression levels of non-injected HFD and CHD groups. For the qPCR analyses, all results
used for interpretation are presented as bar graphs.

36
Chapter 3
3 Results
3.1 Perinatal-HFD offspring display differences in body weight throughout development
The Prenatal-HFD exposed offspring displayed lower gestational body weights during PND 1 (p
< 0.05), while maintaining higher body weights after postnatal HFD-exposure by PND 17 (p <
0.01) [Day by diet interaction: F(1,24) = 11.903, p < 0.0001, Figure 4a]. Although the adults had
been fed CHD since PND 21, the perinatal HFD-exposed rats maintained higher body weights
than the CHD group prior to the drug challenges at PND 90 [F(1,165) = 82.455, p < 0.0001], with
males weighing more than females on average [Diet by sex interaction: F(1,165) = 7.587, p < 0.01,
Figure 4b].
Figure 4. Body weights of offspring, as (a) pups from PND 1-21, Bar****: p < 0.0001 Diet by
day interaction; *, **: p < 0.05, p < 0.01 post-hoc comparison within day, and (b) adults prior to
drug challenges, Bar****: p < 0.0001 Main effect of diet; ****: p < 0.0001 Main effect of sex.
a) b)

37
3.2 Perinatal-HFD exposed animals display exacerbated anti-inflammatory transcriptional
responses to CORT challenge in the amygdala
In response to CORT, pro- and anti-inflammatory transcript levels significantly increased for
both diet groups (Table 4; Post-hocs: Supplementary Table 1). Both diet groups for males
exhibited higher relative abundance of IL-6 and IL-6/IL-10 ratios post-CORT (Figure 5a).
Increases for anti-inflammatory molecules, IkBa and MKP-1 were significantly higher for the
HFD males than the CHD males [Diet by drug interaction: IkBa: F(1,23) = 12.020 , p < 0.01, MKP-
1: F(1,23) = 7.977 , p < 0.01; Figure 5b]. This coincided with a maintenance of higher levels of GC
signaling molecules than the CHD males [GR: F(1,23) = 4.909 , p < 0.05, CRH: Diet by drug
interaction: F(1,23) = 3.295 , p < 0.1], which significantly downregulated GR (p < 0.01) while HFD
males slightly increased CRH expression (p < 0.1) (Figure 5c).
The females displayed sparse changes in transcript abundance when responding to CORT (Table
4b). HFD females significantly increase the amount of pro- and anti-inflammatory transcript
levels [IL-6/IL-10: F(1,23) = 5.687 , p < 0.05, IkBa: F(1,23) = 9.065, p < 0.01; Figure 6a,b], while
decreasing in abundance of GR (p < 0.05) (Figure 6c).

38
a) Pro-inflammatory Genes
b) Anti-inflammatory Genes
b) GC-signaling Genes

39
Figure 5. Transcript level data for CORT-challenged males in the amygdala. (a) Pro-
inflammatory transcripts are increased for both diet groups. (b) Perinatal HFD-exposed males
maintain higher transcription of anti-inflammatory genes in response to CORT, and (b)
transcription of GC-signaling genes are sustained and enhanced for Perinatal HFD-exposed
males. Bar**, #: p < 0.01, p < 0.1 post-hoc comparison within diet, **, *: p < 0.01, p < 0.05
post-hoc comparison within drug,
a) Pro-inflammatory Genes b) Anti-inflammatory Genes
c) GC-signaling genes
Figure 6. Transcript level data for CORT-challenged females in the amygdala. (a) Perinatal
HFD-exposed females increase pro- and b) anti-inflammatory transcript abundance in response

40
to CORT, and (b) abundance of GR is reduced for Perinatal HFD-exposed females. Bar*: p <
0.05 post-hoc comparison within diet.
Table 4. Transcriptional response to CORT challenge in the amygdala. 2 x 2 ANOVAs of qPCR
gene expression data separated by (a) males and (b) females. P-values < 0.05 are highlighted.
a) Males
b) Females
Mean relative abundance Diet Drug Diet x Drug
Function Gene HFD CTL CHD CTL HFD CORT CHD CORT F P F P F P
GC signalling GR 1.3 1.246 0.996 1.181 0.305 0.5872 2.414 0.1368 1.003 0.3291
MR 1.155 1.37 1.168 1.236 1.748 0.2027 0.317 0.5802 0.47 0.5019
CRH 0.511 0.542 0.601 0.495 0.147 0.7061 0.047 0.8314 0.479 0.4973
Pro-inflammatory CD11b 0.679 0.811 0.514 0.695 1.812 0.195 1.469 0.2412 0.046 0.8324
NFkB 1.027 0.92 0.891 0.822 1.203 0.2865 2.117 0.162 0.056 0.8153
IL-6 1.255 1.594 1.98 1.594 0.004 0.9493 1.003 0.3306 1.002 0.3308
IL-6/IL-10 0.916 1.053 1.25 1.206 0.207 0.6548 5.687 0.283 0.787 0.3867
Anti-inflammatory IGF-1 0.829 0.913 0.912 0.866 0.028 0.8683 0.027 0.8716 0.342 0.5656
IkBa 0.443 0.447 0.763 0.611 0.839 0.3711 9.065 0.0072 0.932 0.3466
MKP-1 0.574 0.602 0.727 0.608 0.421 0.5248 1.277 0.2732 1.075 0.3135
IL-10 1.568 1.538 1.573 1.311 0.287 0.5988 0.165 0.6893 0.182 0.6745
Epigenetic regulation DNMT1 0.506 0.531 0.694 0.59 0.179 0.6771 1.742 0.2026 0.482 0.496
DNMT3a 0.572 0.647 0.514 0.609 1.251 0.2781 0.403 0.5335 0.018 0.8937
DNMT3b 0.473 0.558 0.274 0.401 1.321 0.2647 3.691 0.0699 0.051 0.823
MeCP2 0.523 0.531 0.524 0.482 0.048 0.8279 0.098 0.7573 0.104 0.7508
Gadd45b 0.397 0.441 0.49 0.464 0.021 0.887 0.849 0.3683 0.322 0.577
Mean relative abundance Diet Drug Diet x Drug
Function Gene HFD CTL CHD CTL HFD CORT CHD CORT F P F P F P
GC signalling GR 1.132 1.035 0.942 0.703 4.909 0.0385 11.796 0.0026 0.875 0.3609
MR 1.095 1.394 1.043 1.021 2.3 0.145 5.459 0.03 3.104 0.0934
CRH 0.411 0.513 0.649 0.34 0.834 0.3721 0.083 0.7767 3.295 0.0845
Pro-inflammatory CD11b 0.498 0.748 0.773 0.501 0.006 0.9401 0.01 0.9197 3.485 0.0767
NFkB 0.825 0.859 0.735 0.677 0.022 0.8829 3.075 0.0948 0.35 0.5608
IL-6 0.683 0.625 1.869 1.547 0.436 0.5165 13.37 0.0016 0.209 0.6523
IL-6/IL-10 0.285 0.245 1.084 1.183 0.038 0.8474 32.65 < 0.0001 0.209 0.6527
Anti-inflammatory IGF-1 0.868 0.946 0.879 0.642 0.805 0.3802 2.716 0.115 3.116 0.0928
IkBa 0.332 0.313 1.097 0.78 15.432 0.0008 207.037 < 0.0001 12.02 0.0024
MKP-1 0.449 0.494 0.997 0.673 4.54 0.0457 31 < 0.0001 7.977 0.0105
IL-10 2.319 2.763 1.906 1.497 0.001 0.9702 3.286 0.0849 0.846 0.3687
Epigenetic regulation DNMT1 0.632 0.731 0.827 0.469 2.781 0.111 0.186 0.6706 8.607 0.0082
DNMT3a 0.871 1.09 0.581 0.454 0.219 0.645 21.908 0.0001 3.063 0.0954
DNMT3b 0.405 0.571 0.45 0.436 0.376 0.5466 0.134 0.7177 0.525 0.4769
MeCP2 0.584 0.599 0.723 0.407 3.786 0.0659 0.118 0.7352 4.586 0.0447
Gadd45b 0.367 0.406 0.589 0.381 1.865 0.1872 2.539 0.1268 3.997 0.0594

41
3.3 Perinatal-HFD exposure incurs similar, but sex-dependent transcriptional responses to
CORT in the hippocampus
In the hippocampus, pro- and anti-inflammatory transcript levels increased in response to CORT
for all groups except the CHD females, where they were all reduced (Table 5; Post-hocs:
Supplementary Table 2). The transcript levels of these genes and GR were initially higher than
the HFD female groups, which had not been observed in the amygdala. Following CORT-
injection, the diet-differences in GR, CD11b, and NFkB levels were thereby abolished (Figure
8a,c). On the other hand, the abundance of anti-inflammatory genes, IkBa and MKP-1 increased
for HFD females to the extent that it exceeded those of CHD females (Figure 8b). A trend of
increased IL-6, and IL-6/IL-10 ratios was also only apparent for HFD-challenged females (p <
0.1, Supplementary Table 2). For both sexes, elevations in anti-inflammatory transcription
associated with either a maintenance of higher GR levels relative to CHD groups, or a lack of
reduction in response to CORT [Males: F(1,23) = 4.655, p < 0.05, Females: Diet by drug
interaction: F(1,23) = 6.457, p < 0.05, Figure 7c, 8c].
a) Pro-inflammatory Genes

42
b) Anti-inflammatory Genes
c) GC-signaling Genes
Figure 7. Transcript level data for CORT-challenged males in the hippocampus. (a) Pro-
inflammatory transcription is increased for both diet groups. (b) Perinatal HFD-exposed males
maintain higher levels of anti-inflammatory genes in response to CORT, and (b) levels of GC-
signaling genes are sustained for Perinatal HFD-exposed males. Bar****, **, *: p < 0.0001, p <
0.01, p < 0.05 post-hoc comparison within diet, *, #: p < 0.05, p < 0.1 post-hoc comparison
within drug.

43
a) Pro-inflammatory Genes
b) Anti-inflammatory Genes c) GC-signaling Genes
Figure 8. Transcript level data for CORT-challenged
females in the hippocampus. (a) Regular diet-exposed
females reduce transcription of pro-inflammatory
genes and (c) GR relative to Perinatal HFD levels in
response to CORT. (b) HFD-CORT females maintain
higher abundance of anti-inflammatory genes than
CHD-CORT. Bar**, *, #: p < 0.01, p < 0.05, p < 0.1
post-hoc comparison within diet, *, #: p < 0.05, p <
0.1 post-hoc comparison within drug.

44
Table 5. Transcriptional response to CORT challenge in the hippocampus. 2 x 2 ANOVAs of
qPCR gene expression data separated by (a) males and (b) females. P-values < 0.05 are
highlighted.
a) Males
b) Females
Mean relative abundance Diet Drug Diet x Drug
Function Gene HFD CTL CHD CTL HFD CORT CHD CORT F P F P F P
GC signalling GR 1.688 1.534 1.62 0.961 4.655 0.0433 2.895 0.1044 1.792 0.1957
MR 1.633 1.49 2.09 1.134 4.837 0.0398 0.041 0.8418 2.653 0.119
Pro-inflammatory CD11b 1.283 1.424 1.559 1.206 0.188 0.6696 0.014 0.9066 1.026 0.3232
NFkB 1.082 0.896 1.286 0.864 4.067 0.0574 0.324 0.5757 0.616 0.4418
IL-6 1.347 0.745 3.01 4.04 0.213 0.6494 28.663 < 0.0001 3.108 0.094
IL-6/IL-10 0.633 0.237 1.291 1.327 0.524 0.478 12.419 0.0023 0.757 0.395
Anti-inflammatory IGF-1 1.199 1.188 2.02 1.486 1.039 0.3209 4.362 0.0505 0.95 0.3419
IkBa 0.504 0.458 2.245 1.128 7.459 0.0129 32.086 < 0.0001 6.333 0.0205
MKP-1 0.607 0.53 2.109 1.112 7.517 0.0126 28.303 < 0.0001 5.515 0.0292
IL-10 3.168 3.054 2.783 3.595 0.329 0.5733 0.016 0.8997 0.579 0.4562
Epigenetic regulation DNMT1 0.588 0.61 1.089 0.576 1.202 0.286 1.086 0.3098 1.423 0.2469
DNMT3a 0.674 0.649 0.799 0.555 1.059 0.3158 0.013 0.9093 0.703 0.4115
MeCP2 0.589 0.674 0.789 0.528 0.456 0.5071 0.043 0.8378 1.752 0.2006
Gadd45b 0.73 0.774 1.295 0.734 1.39 0.2523 1.427 0.2463 1.902 0.1831
Mean relative abundance Diet Drug Diet x Drug
Function Gene HFD CTL CHD CTL HFD CORT CHD CORT F P F P F P
GC signalling GR 1.282 1.818 1.265 1.071 1.416 0.248 7.057 0.0152 6.457 0.0194
MR 1.032 1.245 1.018 0.985 0.751 0.3964 1.746 0.2013 1.411 0.2488
Pro-inflammatory CD11b 0.995 1.695 0.991 0.836 2.253 0.149 5.652 0.0275 5.534 0.029
NFkB 1.068 1.602 1.216 1.052 2.278 0.1468 2.673 0.1177 8.079 0.0101
IL-6 0.83 1.152 1.38 1.104 0.014 0.9067 1.693 0.2079 2.397 0.1373
IL-6/IL-10 0.424 0.806 1.262 0.635 0.23 0.637 1.71 0.2058 3.911 0.0619
Anti-inflammatory IGF-1 1.395 1.633 1.511 1.423 0.213 0.6492 0.082 0.7772 0.995 0.3305
IkBa 0.48 0.634 1.16 0.751 2.136 0.1594 20.98 0.0002 10.486 0.0041
MKP-1 0.439 0.662 0.806 0.514 0.136 0.716 1.374 0.2549 7.651 0.0119
IL-10 2.052 2.049 1.951 1.791 0.034 0.8564 0.164 0.69 0.032 0.8602
Epigenetic regulation DNMT1 1.573 1.709 0.851 0.647 0.014 0.9079 9.355 0.0062 0.341 0.5657
DNMT3a 1.681 1.672 0.685 0.56 0.046 0.8316 11.563 0.0028 0.034 0.8549
MeCP2 1.047 1.34 0.711 0.526 0.236 0.6326 26.153 < 0.0001 4.512 0.0463
Gadd45b 1.297 1.1 0.751 0.553 0.754 0.3955 5.778 0.026 2.12E-06 0.9989

45
3.4 Perinatal HFD-exposed animals display exacerbated anti-inflammatory transcriptional
responses to LPS challenge in the amygdala
Both pro- and anti-inflammatory transcript levels increased for LPS-challenged subjects,
excluding the CHD females (Table 6; Post-hocs: Supplementary Table 3). HFD males
additionally increased in MKP-1 [F(1,23) = 6.450 , p < 0.05, Figure 9a] while displaying more
significant increases in IkBa (p < 0.01) and less significant drops in IL-6/IL-10 ratios (p < 0.05).
HFD females increased IkBa transcript levels while abundance of anti-inflammatory genes were
unchanged for CHD females [F(1,23) = 7.205 , p < 0.05, Figure 10b]. Heightened pro-
inflammatory activity was further evidenced for the CHD females, with higher transcription of
NFkB [F(1,23) = 4.360, p < 0.05; Figure 10a]. Post-hoc analyses revealed this to only be
significant after injection with LPS (p < 0.1) (Figure 10a). Both CHD males and females
reduced MR and GR expression [MR: p < 0.1, GR: F(1,23) = 4.928, p < 0.05; Figure 9b, 10b].
a) Anti-inflammatory Genes b) GC-signaling Genes
Figure 9. Transcript level data for LPS-challenged males in the amygdala. (a) Perinatal HFD-
exposed males increase MKP-1 levels in response to LPS, and (b) levels of MR are sustained for
Perinatal HFD-exposed males. Bar *, #: p < 0.05, p < 0.1 post-hoc comparison within diet.

46
a) Pro-inflammatory Genes b) Anti-inflammatory Genes
b) GC-signaling Genes
Figure 10. Transcript level data for LPS-challenged females in the amygdala. (a) CHD-LPS
females sustain higher abundance of NFkB than HFD-LPS females, while (b) Perinatal HFD-
exposed females increase IkBa in response to LPS, and (c) levels of GR are sustained for
Perinatal HFD-exposed females. Bar *, #: p < 0.05, p < 0.1 post-hoc comparison within diet, *: p
< 0.05 post-hoc comparison within drug.

47
Table 6. Transcriptional response to LPS challenge in the amygdala.. 2 x 2 ANOVAs of qPCR
gene expression data separated by (a) males and (b) females. P-values < 0.05 are highlighted.
a) Males
b) Females
Mean relative abundance Diet Drug Diet x Drug
Function Gene HFD CTL CHD CTL HFD LPS CHD LPS F P F P F P
GC signalling GR 0.83 0.792 0.708 0.698 0.081 0.7795 1.66 0.2123 0.028 0.8695
MR 0.981 1.096 0.97 0.906 0.053 0.8209 0.801 0.3815 0.634 0.4532
Pro-inflammatory CD11b 0.849 1.417 1.094 0.986 1.72 0.2045 0.279 0.6034 3.723 0.068
NFkB 1.029 1.045 1.322 1.535 0.147 0.7055 1.724 0.204 0.11 0.7437
IL-6 0.417 0.495 2.031 1.988 0.001 0.9773 6.491 0.0192 0.01 0.9221
IL-6/IL-10 0.043 0.033 0.912 0.986 0.027 0.8711 22.377 0.0001 0.047 0.8305
Anti-inflammatory IGF-1 1.128 1.107 1.369 1.492 0.156 0.6973 5.851 0.0252 0.303 0.588
IkBa 0.342 0.379 0.678 0.724 0.266 0.6117 17.963 0.0004 0.003 0.9566
MKP-1 0.465 0.495 0.762 0.981 0.652 0.429 6.45 0.0195 0.375 0.5473
IL-10 9.359 15.054 2.322 1.984 0.898 0.3546 12.652 0.002 1.139 0.2986
Mean relative abundance Diet Drug Diet x Drug
Function Gene HFD CTL CHD CTL HFD LPS CHD LPS F P F P F P
GC signalling GR 0.704 0.823 0.565 0.594 0.809 0.3803 4.928 0.0395 0.297 0.5923
MR 0.944 1.055 1.123 1.366 2.201 0.1535 4.254 0.0524 0.305 0.5869
Pro-inflammatory CD11b 0.916 1.035 0.856 0.558 0.463 0.5042 4.163 0.0548 2.512 0.1287
NFkB 1.275 2.402 1.163 3.557 4.36 0.0498 0.383 0.5429 0.565 0.461
IL-6 0.365 0.281 1.573 2.892 0.76 0.3936 7.277 0.0139 0.981 0.3337
IL-6/IL-10 0.122 0.147 1.214 1.082 0.096 0.7602 34.875 < 0.0001 0.208 0.6529
Anti-inflammatory IGF-1 0.953 1.134 0.849 1.158 2.77 0.1116 0.074 0.7888 0.19 0.6673
IkBa 0.369 0.47 0.78 0.906 0.513 0.483 7.205 0.0151 0.006 0.9379
MKP-1 0.813 0.898 0.825 0.986 0.526 0.4778 0.087 0.7713 0.05 0.8258
IL-10 2.928 2.914 1.829 2.495 0.31 0.5836 1.685 0.209 0.337 0.5679

48
3.5 Perinatal-HFD exposed animals display altered pro-inflammatory transcriptional responses
to CORT/LPS challenge in the amygdala
When responding to CORT/LPS, the IL-6/IL-10 ratio increased higher than when injected with
the same dose of LPS alone, suggesting a synergistic effect of CORT. However, post-hoc
analyses suggest it is only significant for CHD groups [Males: F(1,22) = 4.715, p < 0.05, Females:
Diet by drug interaction: F(1,23) = 6.228, p < 0.05; Figure 11]. In this condition, HFD groups
reduce transcript levels of GR, and for females, the levels drop lower than the CHD group
(Table 7; Post-hocs: Supplementary Table 4). Coinciding with the lower levels of GR, MR,
and CRH is lower pro-inflammatory transcription for the HFD females (Figure 13). It is less
clear what is occurring with the HFD males, as higher NFkB levels are maintained alongside a
trend of reduced IL-6/IL-10 ratios (Figure 12a). In response to CORT/LPS, CHD males reduce
MR to levels equivalent to basal levels for HFD males (Figure 12b).
Figure 11. Increases in IL-6/IL-10 ratios between LPS and CORT/LPS. Bar*, #: p < 0.05, p <
0.1 post-hoc comparison within diet; #: p < 0.1 post-hoc comparison within drug. The increases
in IL-6/IL-10 ratios for all groups are significant (p < 0.01).

49
a) Pro-inflammatory Genes
b) GC-signaling Genes
Figure 12. Transcript level data for CORT/LPS-challenged males in the amygdala. (a) HFD-
CORT/LPS males sustain higher NFkB levels, but lower IL-6/IL-10 ratios than CHD-
CORT/LPS males. (b) Regular diet-exposed males reduce levels of MR to Perinatal HFD levels
in response to CORT/LPS. Bar***, **, *: p < 0.001, p < 0.01, p < 0.05 post-hoc comparison
within diet, *, #: p < 0.05, p < 0.1 post-hoc comparison within drug.

50
a) Pro -inflammatory genes b) GC signaling genes
Figure 13. Transcript level data for CORT/LPS-challenged females in the amygdala. (a)
CORT/LPS-induced pro-inflammatory signaling is dampened for Perinatal HFD-exposed
females, and (b) abundance of GC signaling genes are lower than for CHD females. Bar **, *: p
< 0.01, p < 0.05 post-hoc comparison within diet, **, *, #: p < 0.01, p < 0.05, p < 0.1 post-hoc
comparison within drug.

51
Table 7. Transcriptional response to CORT/LPS challenge in the amygdala. 2 x 2 ANOVAs of
qPCR gene expression data separated by (a) males and (b) females. P-values < 0.05 are
highlighted.
a) Males
b) Females
Mean relative abundance Diet Drug Diet x Drug
Function Gene HFD CTL CHD CTL HFD CORT/LPS CHD CORT/LPS F P F P F P
GC signalling GR 1.595 1.628 1.027 0.926 0.091 0.7657 32.485 < 0.0001 0.359 0.5559
MR 1.73 2.297 1.42 1.148 0.882 0.3595 21.517 0.0002 7.104 0.0153
CRH 0.614 0.485 0.603 0.751 0.012 0.9143 2.33 0.1434 2.732 0.1148
Pro-inflammatory CD11b 1.057 1.047 0.891 0.927 0.004 0.9533 0.457 0.5071 0.012 0.9156
NFkB 0.622 0.656 0.409 0.322 0.208 0.6535 22.474 0.0001 1.117 0.3039
IL-6 0.392 0.366 1.461 1.214 0.857 0.3662 42.237 < 0.0001 0.56 0.4635
IL-6/IL-10 0.147 0.185 0.971 1.265 3.059 0.0973 100.509 < 0.0001 1.829 0.193
Anti-inflammatory IGF-1 1.231 1.46 0.782 0.697 0.298 0.5915 21.051 0.0002 1.4 0.2513
IkBa 0.373 0.378 0.694 0.563 1.648 0.2147 26.994 < 0.0001 1.937 0.1801
MKP-1 0.476 0.518 0.525 0.46 0.063 0.8043 0.01 0.9214 1.371 0.2561
IL-10 2.684 2.613 1.545 1.042 0.772 0.3907 17.266 0.0005 0.439 0.5156
Epigenetic regulation DNMT1 1.568 1.486 1.197 1.081 0.584 0.4539 9.007 0.0073 0.017 0.8972
DNMT3a 1.182 1.426 1.158 1.072 0.381 0.5445 2.207 0.1538 1.678 0.2107
DNMT3b 0.399 0.699 0.449 0.379 0.933 0.3463 1.275 0.2728 2.395 0.1382
MeCP2 0.77 0.82 0.873 0.82 0.001 0.9812 0.666 0.4245 0.692 0.4157
Gadd45b 1.295 1.027 1.162 1.386 0.022 0.8828 0.577 0.4562 2.708 0.1154
Mean relative abundance Diet Drug Diet x Drug
Function Gene HFD CTL CHD CTL HFD CORT/LPS CHD CORT/LPS F P F P F P
GC signalling GR 1.826 1.757 0.787 1.193 0.779 0.388 17.633 0.0004 1.549 0.2276
MR 2.203 2.4 1.16 1.493 1.263 0.2745 17.123 0.0005 0.083 0.7764
CRH 0.753 0.535 0.641 1.009 0.447 0.5114 2.588 0.1233 6.758 0.0171
Pro-inflammatoryCD11b 2.583 2.509 1.121 1.431 0.161 0.6921 18.534 0.0003 0.424 0.5222
NFkB 1.703 1.794 0.567 0.836 0.615 0.4421 20.705 0.0002 0.148 0.7042
IL-6 1.736 1.671 1.659 2.989 1.164 0.2934 1.12 0.3025 1.415 0.2481
IL-6/IL-10 0.686 0.639 1.796 4.693 4.359 0.0498 14.315 0.0012 4.651 0.0434
Anti-inflammatoryIGF-1 1.387 1.313 0.852 0.887 0.006 0.9385 3.765 0.0666 0.049 0.8278
IkBa 0.655 0.726 0.777 0.802 0.304 0.5873 1.283 0.2708 0.07 0.7945
MKP-1 0.762 0.757 0.4 0.508 0.225 0.6406 7.94 0.0106 0.27 0.6088
IL-10 2.55 2.639 1.096 0.743 0.18 0.6762 28.941 < 0.0001 0.503 0.4863
Epigenetic regulationDNMT1 1.558 1.572 0.93 1.312 1.111 0.3044 5.577 0.0284 0.957 0.3396
DNMT3a 1.257 1.335 0.788 1.072 0.891 0.3565 3.638 0.0709 0.287 0.5981
DNMT3b 1.065 0.903 0.534 0.9 0.353 0.5591 2.405 0.1366 2.352 0.1408
MeCP2 0.794 0.876 0.859 1.012 1.538 0.2293 1.129 0.3006 0.142 0.7106
Gadd45b 0.978 1.591 1.262 1.485 4.085 0.0568 0.185 0.6716 0.885 0.3582

52
3.6 Diet-dependent transcriptional responses cluster with transcript levels of epigenetic
regulators in the amygdala
Next, we examined the relative abundance of the DNMT isoforms, MeCP2, and Gadd45b in
CORT- and CORT/LPS-challenged individuals, where most of the diet, diet x drug, and sex
effects had been observed (Tables 4, 7; Supplementary Tables 1, 4). Post-hoc analyses revealed
that the directionality of the diet x drug interactions tended to resemble, or ‘cluster’ with those of
GR, MR, and CRH, and the inflammatory genes exhibiting immunosuppressive activity. In
particular, this can be observed for the CORT-injected males, and the CORT/LPS-injected
females in the amygdala (Table 8a,b; Supplementary Table 1a, 4b). For both, the levels of
DNMT1 were commonly clustered.
Table 8. Transcriptional response to CORT and CORT/LPS in the amygdala grouped by post-
hoc comparisons within diet and drug. Changes in transcript levels of epigenetic mediators
cluster with those impacted by Perinatal HFD for (a) CORT-challenged males, and (b)
CORT/LPS-challenged females. # p < 0.1, * p < 0.05, ** p < 0.01, n.s. not significant. Shaded
boxes: Diet x Drug interaction p < 0.05.
a) CORT-injected Males
Function Gene Effect of CORT within diet group Effect of diet within drug group
GC signalling GR CHD CTL > CHD CORT ** HFD CORT > CHD CORT *
MR CHD CTL > CHD CORT * HFD CTL < CHD CTL #
CRH HFD CTL < HFD CORT # HFD CORT > CHD CORT *
Anti-inflammatory IGF-1 CHD CTL > CHD CORT # HFD CORT > CHD CORT #
IkBa CTL < CORT, HFD + CHD ** HFD CORT > CHD CORT **
MKP-1 HFD CTL < HFD CORT ** HFD CORT > CHD CORT *
Epigenetic regulation DNMT1 CHD CTL > CHD CORT * HFD CORT > CHD CORT **
MeCP2 CHD CTL > CHD CORT # HFD CORT > CHD CORT *
Gadd45b HFD CTL < HFD CORT # HFD CORT > CHD CORT #

53
b) CORT/LPS-injected females
3.7 DNMT activity reduces in response to CORT and CORT/LPS for Perinatal HFD-exposed
males, but not CORT/LPS females in the amygdala
Our transcript analyses of the DNMTs provoked us to examine what may be occurring with the
enzymes on the molecular level. We observed that in response to CORT, HFD males showed
robust decreases in DNMT activity (p < 0.05), while it slightly increased for CHD males (p <
0.1) (Figure 14a). For the CORT/LPS condition, HFD males once again decreased DNMT
activity (p < 0.05), while it remained unchanged for CHD (Figure 14b). Females did not show
changes in DNMT activity in response to CORT/LPS (Figure14c). Spearman's Correlations
revealed that DNMT activity correlates with DNMT1 transcript levels, and not DNMT3a or
DNMT3b in both CORT and CORT/LPS-injected males. DNMT1 strongly correlates negatively
to activity for CORT (rho = -0.578, p < 0.01), while a positive trend is apparent in response to
CORT/LPS (rho = 0.364, p < 0.1) (Table 9). In both instances, DNMT1 also strongly correlates
to DNMT3a transcript levels (CORT: rho = 0.487, p < 0.05, CORT+LPS: rho = 0.786, p < 0.01).
There were non-significant increases in protein levels in CORT-injected HFD and CHD males,
Function Gene Effect of CORT/LPS within diet group Effect of diet within drug group
GC signalling GR CTL > CORT/LPS, HFD **, CHD * HFD CORT/LPS < CHD CORT/LPS **
MR CTL > CORT/LPS, CHD **, HFD * HFD CORT/LPS < CHD CORT/LPS **
CRH CHD CTL < CHD CORT/LPS ** HFD CORT/LPS < CHD CORT/LPS **
Pro-inflammatory CD11b CTL > CORT/LPS, HFD **, CHD * HFD CORT/LPS < CHD CORT/LPS *
NFkB CTL > CORT/LPS, HFD **, CHD * HFD CORT/LPS < CHD CORT/LPS #
IL-6/IL-10 CTL < CORT/LPS, HFD + CHD ** HFD CORT/LPS < CHD CORT/LPS #
Epigenetic regulation DNMT1 HFD CTL > HFD CORT/LPS * HFD CORT/LPS < CHD CORT/LPS *
DNMT3a HFD CTL > HFD CORT/LPS # HFD CORT/LPS < CHD CORT/LPS *
DNMT3b HFD CTL > HFD CORT/LPS * n.s.
Gadd45b n.s. HFD CTL < CHD CTL #

54
which did not correlate with either transcript levels or DNMT activity (mRNA: rho = 0.015, p =
0.946, DNMT activity: rho = 0.109, p = 0.629) (Figure 14d).
0
500000
1000000
1500000
2000000
2500000
3000000
3500000
4000000
4500000
Non-injected CORT+LPS
DN
MT
Act
ivit
y (
RF
U/h
/mg
)
HFD-Males
CHD-Males
0
200
400
600
800
1000
1200
DN
MT
1 (
ng
/mg
Pro
tein
)
HFD
CHD
0
500000
1000000
1500000
2000000
2500000
3000000
3500000
Non-injected CORT
DN
MT
Act
ivit
y (
RF
U/h
/mg
)
HFD-Males
CHD-Males
0
1000000
2000000
3000000
4000000
5000000
6000000
7000000
Non-injected CORT+LPS
DN
MT
Act
ivit
y (
RF
U/h
/mg
)
HFD-Females
CHD-Females
**
____*____
a) CORT-injected males
b) CORT/LPS-injected males
c) CORT/LPS-injected females
d) DNMT1 protein levels
Figure 14. DNMT Activity and Protein
Assays. a) Total DNMT activity for
CORT-challenged males. **: p < 0.01
post-hoc diet comparisons within drug.
Post-hoc drug comparisons within diet
(not shown): CHD p < 0.1, HFD p <
0.05. b) Total DNMT activity for
CORT/LPS-challenged males. Bar *: p
< 0.05 post-hoc drug comparisons
within diet (HFD). c) Total DNMT
activity for CORT/LPS-challenged
females (n.s.). d) DNMT1 protein levels
for CORT-challenged males (n.s.).
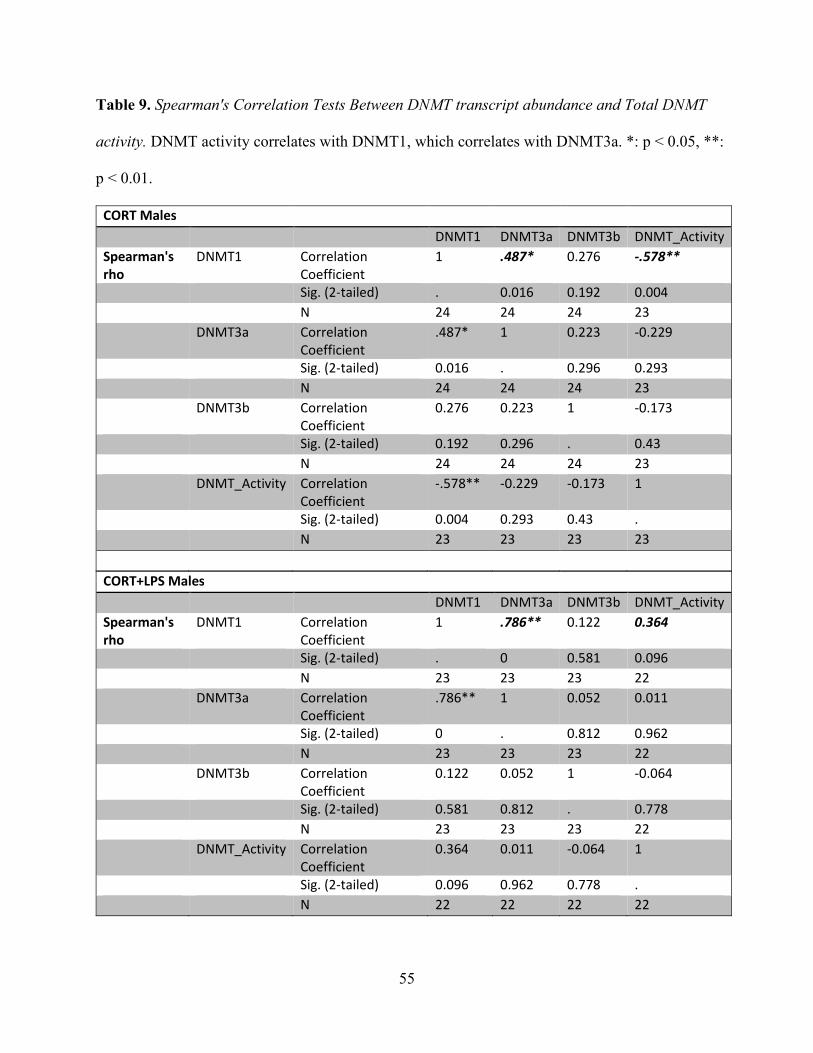
55
Table 9. Spearman's Correlation Tests Between DNMT transcript abundance and Total DNMT
activity. DNMT activity correlates with DNMT1, which correlates with DNMT3a. *: p < 0.05, **:
p < 0.01.
CORT Males
DNMT1 DNMT3a DNMT3b DNMT_Activity
Spearman's
rho
DNMT1 Correlation
Coefficient
1 .487* 0.276 -.578**
Sig. (2-tailed) . 0.016 0.192 0.004
N 24 24 24 23
DNMT3a Correlation
Coefficient
.487* 1 0.223 -0.229
Sig. (2-tailed) 0.016 . 0.296 0.293
N 24 24 24 23
DNMT3b Correlation
Coefficient
0.276 0.223 1 -0.173
Sig. (2-tailed) 0.192 0.296 . 0.43
N 24 24 24 23
DNMT_Activity Correlation
Coefficient
-.578** -0.229 -0.173 1
Sig. (2-tailed) 0.004 0.293 0.43 .
N 23 23 23 23
CORT+LPS Males
DNMT1 DNMT3a DNMT3b DNMT_Activity
Spearman's
rho
DNMT1 Correlation
Coefficient
1 .786** 0.122 0.364
Sig. (2-tailed) . 0 0.581 0.096
N 23 23 23 22
DNMT3a Correlation
Coefficient
.786** 1 0.052 0.011
Sig. (2-tailed) 0 . 0.812 0.962
N 23 23 23 22
DNMT3b Correlation
Coefficient
0.122 0.052 1 -0.064
Sig. (2-tailed) 0.581 0.812 . 0.778
N 23 23 23 22
DNMT_Activity Correlation
Coefficient
0.364 0.011 -0.064 1
Sig. (2-tailed) 0.096 0.962 0.778 .
N 22 22 22 22

56
3.8 DNMT activity correlates with global CpG methylation in CORT-injected male groups in the
amygdala
Given that strong diet-dependent differences in DNMT activity were evident for the CORT-
challenged males in the amygdala, we examined whether this would be reflect genome-wide
changes in methylation using the LUMA technique. It was observed that CHD males sustained
significantly lower methylation levels than HFD males after responding to CORT [F(1,19) =
10.455, p < 0.01, Figure 15a). Correlating the DNMT activity of CORT-challenged groups
revealed significances with highly linear R2 values (HFD: rho = 0.829 , p < 0.05, R2 = 0.9635,
CHD: rho = 1.000, p < 0.01, R2 = 0.9902; Figure 15b). Therefore, DNMTs at least in part
mediates transcriptional responses to CORT that are apparent at an inter-individual level.
Figure 15. Global CpG methylation measured by LUMA in the amygdala of CORT-injected
males. (a) HFD males maintain higher % methylation than CHD in response to CORT, **: p
<0.01 post-hoc comparison within drug. (b) DNMT activity correlates with methylation levels,
HFD: rho = 0.829 , p < 0.05, R2 = 0.9635, CHD: rho = 1.000, p < 0.01, R2 = 0.9902.
a) % Global CpG methylation b) Global methylation vs. DNMT activity

57
Chapter 4
4 Discussion
The goal of this study was to examine the effects of perinatal HFD-exposure on the
transcriptional response to acute stressors in the brain during adulthood. Overall, we found
enhancements in anti-inflammatory signaling in response to CORT and/or LPS in both the
amygdala and hippocampus that appeared to be dependent on GR transcript levels. The diet-
dependence was the most apparent in the amygdala of CORT-challenged HFD males and
CORT/LPS-challenged HFD females, as we had previously found with non-stressed offspring
(Sasaki et al., 2013). Preliminary findings suggest impairments in mediators of DNA methylation
to be related to these diet-differences, since the DNMT activity of CORT-challenged HFD males
was significantly reduced, and global CpG methylation failed to decrease in response to CORT;
although the involvement of other means of epigenetic regulation, such as miRNAs or histone
modifications, cannot be discounted. Together, the findings suggest that early exposure to
maternal overnutrition may place offspring at particular risk for developing infections, and
behavioral/cognitive differences during stressful events encountered during adulthood,
depending on the nature of the stressor. We will start by discussing results from the CORT-
challenge.
4.1 Interpretation of results from CORT challenge
4.1.1 Diet x Sex differences in the Amygdala
Sorrells et al. (2009) suggest that GR is activated to suppress the immune response that initiates
in response to stress, which can be in the form of moderate, chronic doses of CORT, or acute

58
high doses of CORT (Dhabhar, 2000). However, CORT is also known to enhance the immune
response depending on the dose, duration, and type of tissue (Sorrells et al., 2009; Dhabhar,
2009; Bowers et al., 2008). For example, basal levels of CORT are necessary to induce immune
responses during a stressful event (Sorrells et al., 2009). This is comparable to how severe
injuries initially induce a systemic inflammatory response that is followed by an anti-
inflammatory response (Gouel-Chéron et al., 2012), where sepsis arises from simultaneous and
profound elevations of both responses (Hotchkiss et al., 2013; Boomer et al., 2011). GR is well-
established to elicit its immunosuppressive effects by initiating the transcription of anti-
inflammatory molecules including IkBa and MKP-1, and through preventing the transcription
factor NFkB from binding to its target gene binding-sites (Rhen and Cidlowski, 2005). This
includes the pro- and anti-inflammatory cytokines, IL-6 and IL-10 (Libermann and Baltimore,
1990; Saraiva and O'Garra, 2010).
An increased IL-6/IL-10 ratio was observed 3 hours post-CORT, supporting the literature
suggesting that CORT initially induces an immune response to stress. Furthermore, the
expression of GR, MKP-1, and IkBa were tightly correlated, in that perinatal HFD males
sustained higher expressions of each gene, while CHD males maintained relatively lower levels
of each. Analogous to hyperactive immune responses to injury, the simultaneous elevation of
pro- and anti-inflammatory transcripts in the brain may indicate an increased risk for mild-to-
high grades of septic encephalopathy, which is known to lead to long-term cognitive and
behavioral changes such as anxiety-like behavior, depression, and memory and learning deficits
(Calsavara et al., 2013; Ziaja, 2013).
Reduced negative feedback of the HPA-axis from the HFD males was also implicated by
the higher and increased transcription of CRH relative to CHD. This could be caused by the

59
elevations of IL-10 and GR in the amygdala relative to CHD, which are both known to activate
CRH in the hypothalamus and/or amygdala (Kolber et al., 2008; Smith et al., 1999). In the
amygdala, the activity of CRH has long been suggested to enhance anxiety (McGill et al., 2006;
Samaco et al., 2012), reinforcing the possibility of higher anxiety of HFD males responding to
CORT.
Comparing the IL-6/IL-10 ratio between sexes reveals that males sustain much greater
immunosuppressive activity in the absence of stress, and that they increase inflammation to
levels similar to females after being challenged with CORT [F(1,45) = 11.177, p < 0.01, Figure
16]. This supports common findings that females have basally active immune systems that are
more efficient at eliminating pathogens (Scotland et al., 2011; Rohleder et al., 2001). Hence,
during stress, females appear to have a higher threshold required to activate GR, as the levels of
IL-6 to IL-10 are already sufficient to respond to CORT, and does not need to further increase.
This would explain the lack of changes in transcript levels observed.
Figure 16. Comparison of IL-6/IL-10 ratios between CORT-challenged males and females in the
amygdala. Males sustain lower inflammatory activity, then reach levels equivalent to females

60
post-CORT. Bar **, *: p < 0.01, p < 0.05 post-hoc comparison within diet, ****: post-hoc
comparison within drug and diet.
On the other hand, HFD females have a significant increase in IL-6/IL-10 that co-occurs
with significant increases in IkBa, and decreased GR in response to CORT. It has been indicated
that the NFkB pathway can self-regulate by increasing the expression of IkBa independently of
GR (Sorrells et al., 2009). Therefore, the reduced transcription of GR may be inappropriately
causing elevations in IL-6/IL-10, while NFkB induces IkBa as a compensatory mechanism.
Under the assumption that transcript levels in the brain are representative of protein
levels in the plasma, the heightened IL-6/IL-10 ratio in responding to stress may be indicative of
depressive behavior according to a number of studies (Dhabhar et al., 2009; Fredericks et al.,
2009; Voorhees et al., 2013). Given that the periphery is known to send immune cells to the
blood-brain-barrier, cytokine receptors lining the vagus nerve and as recently discovered,
through lymphatic vessels connected to the brain (Louveau et al., 2015), to communicate and
propagate its inflammatory state, this could be a possibility (Maier and Watkins, 2012; Kronfol,
2003). Since MKP-1 is also a marker for depression, and it is elevated in HFD males, they may
also display more depressive behavior (Duric et al., 2010). Therefore, behavioral aberrations
potentially exist to a higher extent for both male and female HFD groups when responding to
stress. Behavioral studies will be needed to confirm this.

61
4.1.2 Females' distinct response in the Hippocampus
Here, we report very similar transcriptional responses between the amygdala and
hippocampus when responding to CORT, especially for males. On the other hand, CHD females
did not display any changes in transcript abundance in the amygdala, while transcription of many
genes reduced in response to CORT in the hippocampus. This appeared to relate to the CHD
group having higher initial transcript levels of these genes than the HFD group. While direct
comparisons of stress-invoked inflammatory changes has been sparse, the brain region-
specificity could be supported by a study that measured the activity and plasticity of subregions
of the hippocampus and amygdala in CHD rats following stressful exposures to a Morris Water
Maze test. It was discovered that long-term potentiation was differentially affected between the
two brain regions (Kavushansky et al., 2006). Since neuronal activity is regulated by cytokines,
the study may support the existence of distinct immune responses between the two regions
(Albensi and Mattson, 2000; Datson et al., 2013). Our results suggested that non-stressed HFD
females possess lower inflammatory activity in the hippocampus, but that they become more
active than CHD females in both limbic brain areas during stress with the higher levels of IkBa
and MKP-1, and exclusive enrichments of IL-6/IL-10 – again accompanied with lack of reduced
abundance of GR. However, considering that the levels become equivalent to those of CHD
females, it is arguable that GR is in fact meant to be functioning in a pro-inflammatory manner,
since CD11b and NFkB displayed reductions in transcript levels in a very similar manner to GR.
In either case, the transcript abundance of GR appears to coincide with differential responses
between the diet groups. The consequence this may have on performance in stressful learning,
memory, and fear-conditioning tasks such as the water maze may be interesting to examine.
Given that pro-inflammatory cytokines is induced and crucial for normal learning, and that

62
memory is impaired by excessively high levels (Williamson et al., 2011), HFD exposure would
likely have an effect in this cohort.
Taken together, perinatal HFD-exposed males in this study demonstrate exacerbated anti-
inflammatory responses, while HFD females show both higher pro- and anti-inflammatory
responses to CORT, which both co-occur with differences in transcription of GR. This could
suggest an increased susceptibility to infections and anxiety behavior for males following
physiological stress, and the risk for developing mild-to-severe grades of inflammatory-related
illnesses such as septic encephalopathy, which is characterized by anxiety, depression, and
cognitive impairments for both sexes.
4.2 Results from LPS challenge in the amygdala
Similar to the HFD females' response to CORT, both pro- and anti-inflammatory transcription
simultaneously increased for both HFD groups responding to LPS while the CHD groups
primarily increased in pro-inflammatory transcription. This again suggests an anti-inflammatory
bias on the part of the perinatal HFD-exposed animals, which occurs with the characteristic lack
of reductions in GR, as well as MR for males. The similarity to the CORT-challenged group is
likely due to the gradual secretion of CORT that naturally occurs after exposure to LPS for the
purpose of ameliorating the immune response - supporting the significance of GR in causing the
diet differences (Quan et al., 2000). Our finding contradicts two studies: One, that suggests MR
to have a pro-inflammatory effect under low-moderate CORT exposure (Changtong et al., 2012),
and another finding from Bilbo and Tsang (2010) where the protein expression of pro-
inflammatory cytokine, IL-1β were elevated in the hippocampus, and microglial activation was
enhanced both basally, and post-LPS challenge. In this study, we did not find significant effects

63
of diet in the hippocampus, nor did we find many in either the amygdala or hippocampus with
the dose used in that study (not shown). Going by CD11b levels, basal microglia concentrations
may have instead been reduced in the HFD-exposed males. Therefore, it's possible that the action
of MR may only be pro-inflammatory depending on the initial state of surrounding microglia.
Given the context-dependency of GR and MR (Sorrells et al., 2009), this could be the case, and
warrants further investigation. Put together, this would support the general idea that the
inflammatory, hence behavioral effects of perinatal HFD would depend on how it affects the
HPA-axis, as well the responsiveness of microglia separately, both of which could vary with
numerous factors such as sex, species strain, handling, or other variations in cohorts. Once more,
the coinciding elevation of pro- and anti-inflammatory transcript abundance may suggest
increased health risks with perinatal HFD-exposure, including grades of septic encephalopathy,
with the accompanying affect and cognitive impairments.
4.3 Results from CORT/LPS challenge in the amygdala
Although CORT is classically known to be immunosuppressive, there have been numerous
studies indicating that GR activation by acute or chronic stress can increase LPS-induced
expression of pro-inflammatory cytokines in limbic brain areas, excluding the hypothalamus
(Sorrells et al., 2009; Frank et al., 2009; Munhoz et al., 2006; de Pablos et al., 2006). This has
been shown to occur at least in part by GR-dependent proliferation and increased activation of
microglia, the immune cells of the brain (Nair and Bonneau, 2006; Frank et al., 2006), and by
lack of inductions of MKP-1 and/or IkBa (Munhoz et al., 2010). The change in function of GR is
speculated to occur because pro-inflammatory responses can be protective in neurons (e.g. when
facilitating hippocampal synaptic plasticity; Albensi and Mattson, 2000), while being damaging
to microglia (Sorrells et al., 2009). Since the pro-inflammatory effects usually occur when CORT

64
is given hours prior to LPS, and not simultaneously, we predicted that previous, perinatal HFD
exposure to CORT would be sufficient to provoke a similar response.
Compared to a high-dose LPS treatment alone (50 ug/kg), we indeed observed elevated
IL-6/IL-10 ratios with the addition of CORT for the CHD groups, with very high levels for the
CHD females. However, pro-inflammatory transcript levels were reduced for HFD females
relative to the CHD group, while it is less clear what is occurring for the HFD males, as IL-6/IL-
10 ratios are slightly higher for CHD males, but NFkB levels are higher for HFD males. In this
case, the CHD females maintain higher abundance of GR and MR than the HFD females -
further suggesting the involvement of GR in directing both pro- and anti-inflammatory signaling
depending on the stressor. The observation of GR decreasing, rather than being sustained,
supports past studies with LPS given after CORT - GR protein levels were shown to be reduced,
even though they were found to be necessary for increasing inflammation (Sorrells et al., 2009).
More support of the dependency on GR is the higher abundance of CD11b, a marker for
microglia, in the CORT/LPS-challenged CHD females relative to the HFD females. Further, the
increased CRH levels for CHD groups supports the enhanced LPS response as being a result of
lower negative feedback on the HPA-axis. Although the results suggest HFD females have an
elevated risk of developing infections against bacteria, their lower IL-6/IL-10 ratio and MKP-1
transcripts may suggest a decreased risk of sickness-induced depression – pointing to the delicate
balance that seems to exist between adaptive versus ‘maladaptive’ responses to stress, in that that
some levels of anxiety, for instance, might be necessary for learning, as previously implicated by
the maternal HFD study by Bilbo and Tsang (2010). Put together, perinatal HFD, combined with
CORT, impairs the pro-inflammatory, neuroprotective response to LPS in females.

65
4.4 Comparison of results with Sasaki et al. (2013) Maternal HFD Study
Here, we report that the majority of differences under baseline (non-injected) conditions occur in
the hippocampus of females, while none were apparent in the amygdala. The very opposite was
discovered in our laboratory’s previous study (Sasaki et al., 2013). Also, results between the
HFD and CHD males were similar in the hippocampus, but slightly different in the amygdala. In
this study, only MR and CD11b were different under baseline (HFD < CHD), whereas some
differences in GR, pro-, and anti-inflammatory transcript levels had been shown before. While
the same diet paradigms, primer sets, and storage/extraction procedures were used, we could not
account for any differences in handling, time of sacrifice, or exposures to cues from challenged
rodents within the room, which may have all contributed to these differing results. In summary,
Figure 17 re-illustrates the CORT/LPS pathway with respect to each gene analyzed in this study
with coloured arrows to indicate the effects of perinatal HFD-exposure we observed.

66
Figure 17. Simplified diagram of the effect of Perinatal HFD on the general CORT/LPS
pathway. Overview: An Inflammatory-challenge such as LPS activates NFkB, which
translocates to the nucleus to induce the transcription of pro-inflammatory cytokines such as IL-6
to initiate the immune response (Libermann and Baltimore, 1990). Over time (dashed arrows),
CORT is secreted to activate GR and repress NFkB activity by blocking its binding to consensus
sequences, and through inducing the expression of anti-inflammatory molecules such as IkBa
and MKP-1 (Rhen and Cidlowski, 2005). NFkB also negatively inhibits itself by progressively
increasing the expression of anti-inflammatory molecules (Sorrells et al., 2009). In the brain,
prior CORT exposure has been shown to enhance immune responses to LPS (Frank et al., 2006).

67
The green arrows indicate where we predominantly observed enhancements in activity as a result
of perinatal HFD-exposure (immunosuppression by CORT), while the orange arrow indicates
where it is reduced (pro-inflammation by CORT). GR is bolded to indicate its central role in the
diet-differences.
4.5 Maternal HFD transmission of CORT as possible mechanism for differential GC
sensitivity
In order to explain these effects, it would be reasonable to question whether perinatal exposure to
HFD may be priming offspring to respond to CORT later in life by directly increasing fetal
exposure to maternal circulating levels of CORT. Indeed, maternal HFD has been reported to
increase plasma CORT and decrease 11β-HSD-2 expression and activity in rodents, which is an
enzyme that catalyzes the inactivation of maternal GCs before they are transferred to the fetus
(Bellisario et al., 2015). Moreover, these conditions are known to lead to reduced gestational
body weight (Bellisario et al., 2015; Lindsay et al., 1996), which independently results in much
of the downstream mental health outcomes in humans (Grissom and Reyes, 2013; Rivera et al.,
2015). The HFD-exposed offspring showed reduced body weight at birth, suggesting that a
heightened exposure to CORT may have altered the way the offspring respond to CORT and
CORT/LPS during adulthood - with receptors of males being able to bind a higher concentration
of CORT, while females reduce transcript levels more intensely with the addition of LPS.
Potential evidence for this is the reduction of basal transcript levels of MR that we
observed in the amygdala of males (Table 6), which should theoretically increase the occupancy
of CORT to GR. This may have permanently conditioned GR to bind higher concentrations of
CORT before downregulating itself during a stress response. Alternatively, the early exposures

68
to GCs could have changed the expression of corticosteroid-binding globulin (CBG), which
escorts CORT through the blood and keeps it in an inactive state (Dhillo et al., 2002). If CBG
levels are higher in HFD-exposed animals, then the downregulation of GR may occur at later
time-points due to decreased contact with CORT - thereby prolonging HPA-axis activity.
However, this is less likely, as we saw higher pro- and anti-inflammatory activity, which
suggests that CORT is activating GR more frequently than with CHD animals. Nonetheless, we
cannot discount the possibility that the microglia of HFD-exposed animals had become more
sensitive to GR, potentially through impaired MeCP2 functionality, as we will discuss below.
Overall, direct perinatal overexposure to CORT offers one possible mechanism for the diet-
dependent transcriptional responses to CORT and CORT/LPS observed in this study.
4.6 Role of Epigenetic Regulators: MeCP2 as potential hub for manifestation of diet- and
sex- differences
In numerous, but not all instances, the transcript changes of epigenetic regulators resemble, or
‘cluster’ with that of GR and its downstream target genes. This can especially be observed for
CORT-challenged males, and CORT/LPS-challenged females in the amygdala. In both cases,
DNMT1 levels were commonly clustered with those of the genes - very similar to a study
between anxious humans which showed concurrent increases in gene expression of all DNMT
isoforms and IL-6 in peripheral blood mononuclear cells, with self-reports of anxiety (Murphy et
al., 2014). This observation provoked us to examine the DNMT activity of these cohorts, and it
was found that HFD males displayed greatly reduced total DNMT activity that tightly negatively
correlated with DNMT1 transcript levels and global CpG methylation levels. However, DNMT1
protein levels were not different across diet and treatment groups, and did not correlate to either
activity or transcript levels. This more closely mirrors what has been found in previous studies of

69
DNMT activity in the amygdala and medial preoptic area of the hypothalamus, where levels of
protein did not change in the presence of differing DNMT activity, suggesting alternate means of
changing activity, such as phosphorylation and/or reduced binding affinity to interacting partners
(McCarthy et al., 2010; Nugent et al., 2015). Reduced interactions with MeCP2 due to changes
in its phosphorylation state would be a strong candidate for explaining the lowered activity, as
this was observed to occur for maternal HFD-exposed groups that showed impaired DNMT
activity in the prefrontal cortex in another study (McKee et al. 2014). Hence, the changes in
transcription of DNMT1 may merely be capturing the effects on GR without affecting actual
expression or function, as GR has been found to target the DNMTs downstream when
responding to CORT (Urb et al., 2015). Co-immunoprecipitation (Co-IP) studies will be needed
to clarify the effect of reduced DNMT activity on interactions with MeCP2 and GR.
On the other hand, there were no differences in DNMT activity across diet and drug for
the CORT/LPS-injected females. We speculated that perhaps, then, demethylase activity may be
responsible for the diet-differences observed in responding to CORT/LPS. This was a possibility,
considering how both DNMTs and the demethylase enzyme, Tet (specifically, Tet1) require
interactions with MeCP2 for its activity (Kimura and Shiota, 2003; Cartron et al., 2013), and
MeCP2 itself represses genes in the microglia induced by GCs and TNF-α, another pro-
inflammatory stimulus (Cronk et al., 2015). There are also findings that Gadd45b modulates the
expression of pro-inflammatory cytokines such as IL-6 in the amygdala (Kigar et al., 2015), and
that adult females, but not males show decreases in Gadd45b expression in response to early
caregiver maltreatment (Blaze and Roth, 2013) – which we have also observed in the amygdala
in response to perinatal HFD. Hence, future studies on measurements of demethylase activity
and Co-IP will be required to confirm this hypothesis. It is particularly apparent that more focus

70
needs to be placed on MeCP2 with studies involving effects of perinatal stress on future stress
challenges.
4.7 Explaining the effects of DNMT activity on Global CpG methylation levels
Since the DNMT activity kits used specifically measured the ability of DNMTs to methylate a
DNA substrate, it was initially surprising to observe the decreased activity occurring with no
changes in global methylation levels for the CORT-challenged HFD males, whereas the levels
significantly reduced for the CHD males. To begin with, reductions in methylation can only be
occurring through active demethylase activity, or passively, through replication events where
DNMT1 does not copy the methylation marks onto newly synthesized DNA. Since DNMT
activity was not reduced for the CHD males, the latter does not appear to apply, and the results
may reinforce the existence of impairments in both methylase and demethylase activity through
perinatal HFD-exposure, where GR-dependent effects on inflammatory pathways depend on the
type of stressor and sex of the stressed individual. In which case, DNMT activity seems essential
for responding to CORT in males, important enough to incur interindividual differences in global
CpG methylation levels, but not for CORT/LPS-challenged females. In order to clarify that the
absence or presence of DNA methylation is leading to the diet-dependent transcriptional
responses to each challenge, future bisulfite pyrosequencing studies will have to be conducted
for the genes studied, particularly GR, which uses epigenetic modifications to down-regulate
itself after activation with CORT (Ramamoorthy and Cidlowski, 2013). To attain a greater
picture of our findings, microarray studies should also be performed to examine potential
discrepancies between transcript responses, and actual gene expression changes, since mRNA
stability can be compromised by miRNA, another epigenetic mechanism.

71
4.8 Conclusion
To summarize, we report that rats exposed to perinatal HFD respond to the acute stress of
CORT, LPS, and CORT/LPS with predominantly enhanced immunosuppressive activity seeming
to relate to GR transcript abundance, and altered abundance of genes associated with anxiety and
depression in a sex and brain region-specific manner. As GR is known to interact with epigenetic
mediators to regulate the transcription of target genes when responding to stress, including its
own downregulation, impaired DNA methylase and demethylase activity through reduced
associations with MeCP2 may at least in part explain the differential responses. Future
experiments including Co-IP, bisulfite pyrosequencing, and more protein activity and
quantification assays will be required to confirm this hypothesis.
References
Afari, N., & Buchwald, D. (2003). Chronic fatigue syndrome: A review. American Journal of
Psychiatry, 160(2), 221–236. doi:10.1176/appi.ajp.160.2.221
Albensi, B. C., & Mattson, M. P. (2000). Evidence for the involvement of TNF and NF-kappaB
in hippocampal synaptic plasticity. Synapse (New York, N.Y.), 35(2), 151–159.
doi:10.1002/(SICI)1098-2396(200002)35:2<151::AID-SYN8>3.0.CO;2-P
Anacker, C., Zunszain, P. a, Carvalho, L. a, & Pariante, C. M. (2012). Europe PMC Funders
Group The glucocorticoid receptor : Pivot of depression and of antidepressant treatment ?,
36(3), 415–425. doi:10.1016/j.psyneuen.2010.03.007.The

72
Bake, S., Selvamani, A., Cherry, J., & Sohrabji, F. (2014). Blood brain barrier and
neuroinflammation are critical targets of IGF-1-mediated neuroprotection in stroke for
middle-aged female rats. PLoS ONE, 9(3). doi:10.1371/journal.pone.0091427
Barker, E. D., Kirkham, N., Ng, J., & Jensen, S. K. G. (2013). Prenatal maternal depression
symptoms and nutrition, and child cognitive function. The British Journal of Psychiatry :
The Journal of Mental Science, 203(6), 417–21. doi:10.1192/bjp.bp.113.129486
Bartha, J. L., Romero-Carmona, R., & Comino-Delgado, R. (2003). Inflammatory cytokines in
intrauterine growth retardation. Acta Obstetricia et Gynecologica Scandinavica, 82(12),
1099–1102. doi:10.1046/j.1600-0412.2003.00259.x
Bellisario, V., Panetta, P., Balsevich, G., Baumann, V., Noble, J., Raggi, C., … Cirulli, F.
(2015). Maternal high-fat diet acts as a stressor increasing maternal glucocorticoids’
signaling to the fetus and disrupting maternal behavior and brain activation in C57BL/6J
mice. Psychoneuroendocrinology, 60, 138–150. doi:10.1016/j.psyneuen.2015.06.012
Benros, M. E., Nielsen, P. R., Nordentoft, M., Eaton, W. W., Dalton, S. O., & Mortensen, P. B.
(2011). Autoimmune diseases and severe infections as risk factors for schizophrenia: A
30-year population-based register study. American Journal of Psychiatry, 168(12), 1303–
1310. doi:10.1176/appi.ajp.2011.11030516
Bilbo, S. D., & Schwarz, J. M. (2009). Early-life programming of later-life brain and behavior: a
critical role for the immune system. Frontiers in Behavioral Neuroscience, 3(August), 1–
14. doi:10.3389/neuro.08.014.2009

73
Bilbo, S. D., & Tsang, V. (2010). Enduring consequences of maternal obesity for brain
inflammation and behavior of offspring. The FASEB Journal : Official Publication of the
Federation of American Societies for Experimental Biology, 24(6), 2104–2115.
doi:10.1096/fj.09-144014
Bilbo, S. D., Yirmiya, R., Amat, J., Paul, E. D., Watkins, L. R., & Maier, S. F. (2008). Bacterial
infection early in life protects against stressor-induced depressive-like symptoms in adult
rats. Psychoneuroendocrinology, 33(3), 261–9. doi:10.1016/j.psyneuen.2007.11.008
Binder, E. B. (2009). The role of FKBP5, a co-chaperone of the glucocorticoid receptor in the
pathogenesis and therapy of affective and anxiety disorders. Psychoneuroendocrinology,
34, S186–S195. doi:10.1016/j.psyneuen.2009.05.021
Blaze, J., & Roth, T. L. (2013). Exposure to caregiver maltreatment alters expression levels of
epigenetic regulators in the medial prefrontal cortex. International Journal of
Developmental Neuroscience, 31(8). doi:10.1016/j.ijdevneu.2013.10.001
Bolton, J. L., & Bilbo, S. D. (2014). Developmental programming of brain and behavior by
perinatal diet: focus on inflammatory mechanisms. Dialogues in Clinical Neuroscience,
16(3), 307–20. Retrieved from
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=4214174&tool=pmcentrez&r
endertype=abstract
Boomer, J. S. (2011). Immunosuppression in Patients Who Die of Sepsis and Multiple Organ
Failure. JAMA: The Journal of the American Medical Association, 306(23), 2594.
doi:10.1001/jama.2011.1829

74
Bourre, J. M., Gozlan-Devillierre, N., Morand, O., & Baumann, N. (1979). Importance of
exogenous saturated fatty acids during brain development and myelination in mice.
Annales de Biologie Animale Biochimie Biophysique, 19(1B), 173–180.
doi:10.1051/rnd:19790205
Bowers, S. L., Bilbo, S. D., Dhabhar, F.S., & Nelson, R.J. (2008) Stressor-specific alterations in
corticosterone and immune responses in mice. Brain, Behavior, and Immunity, 22, 105–
113. doi:10.1016/j.bbi.2007.07.012
Calsavara, A. C., Rodrigues, D. H., Miranda, A. S., Costa, P. a., Lima, C. X., Vilela, M. C., …
Teixeira, A. L. (2013). Late anxiety-like behavior and neuroinflammation in mice
subjected to sublethal polymicrobial sepsis. Neurotoxicity Research, 24(2), 103–108.
doi:10.1007/s12640-012-9364-1
Can,O. D., Ulupinar, E.,Ozkay, U. D., Yegin, B., andOzturk, Y. (2012). The effect of simvastatin
treatment on behavioral parameters, cognitive performance, and hippocampal
morphology in rats fed a standard or a high-fat diet. Behav. Pharmacol. 23, 582–592. doi:
10.1097/FBP.0b013e328356c3f2
Canani, R. B., Costanzo, M. Di, Leone, L., Bedogni, G., Brambilla, P., Cianfarani, S., …
Agostoni, C. (2011). Epigenetic mechanisms elicited by nutrition in early life Nutrition
Research Reviews Nutrition Research Reviews, 198–205.
doi:10.1017/S0954422411000102
Cartron, P.-F., Nadaradjane, A., Lepape, F., Lalier, L., Gardie, B., & Vallette, F. M. (2013).
Identification of TET1 Partners That Control Its DNA-Demethylating Function. Genes &
Cancer, 4(5-6), 235–41. doi:10.1177/1947601913489020

75
Chantong, B., Kratschmar, D. V, Nashev, L. G., Balazs, Z., & Odermatt, A. (2012).
Mineralocorticoid and glucocorticoid receptors differentially regulate NF-kappaB activity
and pro-inflammatory cytokine production in murine BV-2 microglial cells. Journal of
Neuroinflammation, 9(1), 260. doi:10.1186/1742-2094-9-260
Chiba, S., Numakawa, T., Ninomiya, M., Richards, M. C., Wakabayashi, C., & Kunugi, H.
(2012). Chronic restraint stress causes anxiety- and depression-like behaviors,
downregulates glucocorticoid receptor expression, and attenuates glutamate release
induced by brain-derived neurotrophic factor in the prefrontal cortex. Progress in Neuro-
Psychopharmacology and Biological Psychiatry, 39(1), 112–119.
doi:10.1016/j.pnpbp.2012.05.018
Chmielarz, P., Kuśmierczyk, J., Parlato, R., Schütz, G., Nalepa, I., & Kreiner, G. (2013).
Inactivation of glucocorticoid receptor in noradrenergic system influences anxiety- and
depressive-like behavior in mice. PloS One, 8(8), e72632.
doi:10.1371/journal.pone.0072632
Colman, I., Jones, P. B., Kuh, D., Weeks, M., Naicker, K., Richards, M., & Croudace, T. J.
(2014). Early development, stress and depression across the life course: pathways to
depression in a national British birth cohort. Psychological Medicine, 44, 2845–2854.
doi:10.1017/S0033291714000385
Cronk, J. C., Litvak, V., Kipnis, J., Derecki, C., Ji, E., Xu, Y., … Baker, W. (2015). Methyl-CpG
Binding Protein 2 Regulates Microglia and Macrophage Gene Expression in Response to
Article Methyl-CpG Binding Protein 2 Regulates Microglia and Macrophage Gene
Expression in Response to Inflammatory Stimuli, 679–691.

76
Crudo, A., Petropoulos, S., Moisiadis, V. G., Iqbal, M., Kostaki, A., Machnes, Z., … Matthews,
S. G. (2012). Prenatal synthetic glucocorticoid treatment changes DNA methylation states
in male organ systems: Multigenerational effects. Endocrinology, 153(7), 3269–3283.
doi:10.1210/en.2011-2160
Datson, N. A., Van Den Oever, J. M. E., Korobko, O. B., Magarinos, A. M., De Kloet, E. R., &
McEwen, B. S. (2013). Previous history of chronic stress changes the transcriptional
response to glucocorticoid challenge in the dentate gyrus region of the male rat
hippocampus. Endocrinology, 154(C), 3261–3272. doi:10.1210/en.2012-2233
de Pablos, R.M., Villaran, R.F., Arguelles, S., Herrera, A.J., Venero, J.L., Ayala, A., Cano, J., &
Machado, A. (2006). Stress increases vulnerability to inflammation in the rat prefrontal
cortex. Journal of Neuroscience, 26(21), 5709–5719. doi:10.1523/JNEUROSCI.0802-
06.2006
de Souza, C. T., Araujo, E.P., Bordin, S., Ashimine, R., Zollner, R. L., Boschero, A. C., Saad, M.
J., & Velloso, L.A. (2005) Consumption of a fat-rich diet activates a proinflammatory
response and induces insulin resistance in the hypothalamus. Endocrinology, 146, 4192–
4199. doi:10.1210/en.2004-1520
Dhabhar, F. S. (2000). Acute Stress Enhances While Chronic Stress Suppresses Skin Immunity:
The Role of Stress Hormones and Leukocyte Trafficking. Annals of the New York
Academy of Sciences, 917, 876–893. doi: 10.1111/j.1749-6632.2000.tb05454.x
Dhabhar, F. S. (2009). Enhancing versus suppressive effects of stress on immune function:
Implications for immunoprotection and immunopathology. Neuroimmunomodulation, 16,
300–317. doi:10.1159/000216188

77
Dhabhar, F. S., Burke, H. M., Epel, E. S., Mellon, S. H., Rosser, R., Reus, V. I., & Wolkowitz,
O. M. (2009). Low serum IL-10 concentrations and loss of regulatory association
between IL-6 and IL-10 in adults with major depression. Journal of Psychiatric
Research, 43(11), 962–969. doi:10.1016/j.jpsychires.2009.05.010
Dhillo, W. S., Kong, W. M., Le Roux, C. W., Alaghband-Zadeh, J., Jones, J., Carter, G., …
O’Shea, D. (2002). Cortisol-binding globulin is important in the interpretation of
dynamic tests of the hypothalamic-pituitary-adrenal axis. European Journal of
Endocrinology, 146, 231–235. doi:10.1530/eje.0.1460231
Djordjevic, J., Djordjevic, a., Adzic, M., Niciforovic, a., & Radojcic, M. B. (2010). Chronic
stress differentially affects antioxidant enzymes and modifies the acute stress response in
liver of wistar rats. Physiological Research, 59(11), 729–736.
Doherty, a S., Mann, M. R., Tremblay, K. D., Bartolomei, M. S., & Schultz, R. M. (2000).
Differential effects of culture on imprinted H19 expression in the preimplantation mouse
embryo. Biology of Reproduction, 62(6), 1526–1535. doi:10.1095/biolreprod62.6.1526
Driessler, F., Venstrom, K., Sabat, R., Asadullah, K., & Schottelius, a J. (2004). Molecular
mechanisms of interleukin-10-mediated inhibition of NF-kappaB activity: a role for p50.
Clinical and Experimental Immunology, 135(1), 64–73. doi:10.1046/j.1365-
2249.2004.02342.x
Duan, S. Z., Zhang, W. C., & Zheng, X. J. (2013). An Update on Actions of Mineralocorticoid
Receptor in the Cardiovascular System : From Clinical Studies to Basic Research, 6(2),
91–94. doi:10.7156/najms.2013.0602091

78
Dunn, G.A., & Bale, T.L. (2009) Maternal high-fat diet promotes body length increases and
insulin insensitivity in second-generation mice. Endocrinology 150(11), 4999–5009. doi:
10.1210/en.2009-0500
Durairaj, R. V., & Koilmani, E. R. (2014). Environmental enrichment modulates glucocorticoid
receptor expression and reduces anxiety in Indian field male mouse Mus booduga through
up-regulation of microRNA-124a. General and Comparative Endocrinology, 199, 26–32.
doi:10.1016/j.ygcen.2014.01.005
Duric, V., Banasr, M., Licznerski, P., Schmidt, H. D., Stockmeier, C. a, Simen, A. a, … Duman,
R. S. (2010). A negative regulator of MAP kinase causes depressive behavior. Nature
Medicine, 16(11), 1328–1332. doi:10.1038/nm.2219
El-Haschimi, K., Pierroz, D. D., Hileman, S. M., Bjorbaek, C., Flier, J. S. (2000). Two defects
contribute to hypothalamic leptin resistance in mice with diet-induced obesity. Journal of
Clinical Investigation, 105, 1827–1832. doi:10.1172/JCI9842
Fagiolini, M., Jensen, C. L., & Champagne, F. a. (2009). Epigenetic influences on brain
development and plasticity. Current Opinion in Neurobiology, 19(2), 207–12.
doi:10.1016/j.conb.2009.05.009
Fagerås, M., Jenmalm, M. C., Garofalo, P., Böttcher, M. F., Garofalo, R. P., & Björkstén,
B. (1999). Cytokines in breast milk from allergic and nonallergic mothers. Pediatric
Research, 47(1), 157–162. doi:10.1203/00006450-200005000-00006

79
Fatemi, M., Hermann, A., Gowher, H., & Jeltsch, A. (2002). Dnmt3a and Dnmt1 functionally
cooperate during de novo methylation of DNA. European Journal of Biochemistry,
269(20), 4981–4984. doi:10.1046/j.1432-1033.2002.03198.x
Feldker, D. E. M., Morsink, M. C., Veenema, A. H., Datson, N. A., Proutski, V., Lathouwers, D.,
… Vreugdenhil, E. (2006). The effect of chronic exposure to highly aggressive mice on
hippocampal gene expression of non-aggressive subordinates. Brain Research, 1089(1),
10–20. doi:10.1016/j.brainres.2006.02.110
Flegal, K. M., Carroll, M. D., Ogden, C. L., & Curtin, L. R. (2010). Prevalence and Trends in
Obesity Among US Adults, 1999-2008. JAMA, 303(3), 235–41. doi:
10.1001/jama.2009.2014.
Frank, M. G., Baratta, M. V., Sprunger, D. B., Watkins, L. R., & Maier, S. F. (2006). Microglia
serve as a neuroimmune substrate for stress-induced potentiation of CNS pro-
inflammatory cytokine responses. Brain, Behavior, and Immunity, 21(1), 47–59.
doi:10.1016/j.bbi.2006.03.005
Frank, M.G., Miguel, Z.D., Watkins, L.R., & Maier, S.F. (2009). Prior exposure to
glucocorticoids sensitizes the neuroinflammatory and peripheral inflammatory responses
to E. coli lipopolysaccharide. Brain, Behavior, and Immunity, 24(1), 19–30. doi:
10.1016/j.bbi.2009.07.008.
Fredericks, C. a., Drabant, E. M., Edge, M. D., Tillie, J. M., Hallmayer, J., Ramel, W., …
Dhabhar, F. S. (2010). Healthy young women with serotonin transporter SS
polymorphism show a pro-inflammatory bias under resting and stress conditions. Brain,
Behavior, and Immunity, 24(3), 350–357. doi:10.1016/j.bbi.2009.10.014

80
Freeman, D. J. (2010). Effects of maternal obesity on fetal growth and body composition:
implications for programming and future health. Seminars in Fetal and Neonatal
Medicine, 15(2), 113–118. doi:10.1016/j.siny.2009.09.001
Giriko, C. A., Andreoli, C. A., Mennitti, L. V., Hosoume, L. F., Souto Tdos, S., Silva, A. V., et
al. (2013). Delayed physical and neurobehavioral development and increased aggressive
and depression-like behaviors in the rat offspring of dams fed a high-fat diet.
International Journal of Developmental Neuroscience, 31, 731–739. doi:
10.1016/j.ijdevneu.2013.09.001
Godfrey, K. M., Gluckman, P. D., & Hanson, M. A. (2010). Developmental origins of metabolic
disease: life course and intergenerational perspectives. Trends in endocrinology and
metabolism, 21(4), 199-205. doi: 10.1016/j.tem.2009.12.008.
Gouel-Chéron, A., Allaouchiche, B., Guignant, C., Davin, F., Floccard, B., & Monneret, G.
(2012). Early interleukin-6 and slope of monocyte human leukocyte antigen-DR: A
powerful association to predict the development of sepsis after major trauma. PLoS ONE,
7(3). doi:10.1371/journal.pone.0033095
Grissom, N. M., George, R., & Reyes, T. M. (2015). The hypothalamic transcriptional response
to stress is severely impaired in offspring exposed to adverse nutrition during gestation.
Neuroscience, (July). doi:10.1016/j.neuroscience.2015.07.022
Grissom, N. M., & Reyes, T. M. (2013). Gestational overgrowth and undergrowth affect
neurodevelopment: Similarities and differences from behavior to epigenetics.
International Journal of Developmental Neuroscience, 31(6), 406–414.
doi:10.1016/j.ijdevneu.2012.11.006

81
Heim, C., Newport, D. J., Heit, S., Graham, Y. P., Wilcox, M., Bonsall, R., … Nemeroff, C. B.
(2000). Pituitary-adrenal and autonomic responses to stress in women after sexual and
physical abuse in childhood. JAMA : The Journal of the American Medical Association,
284(5), 592–597. doi:joc92111 [pii]
Herrero, A. I., Sandi, C., & Venero, C. (2006). Individual differences in anxiety trait are related
to spatial learning abilities and hippocampal expression of mineralocorticoid receptors.
Neurobiology of Learning and Memory, 86(2), 150–9. doi:10.1016/j.nlm.2006.02.001
Hotchkiss, R. S., Monneret, G., & Payen, D. (2013). Sepsis-induced immunosuppression: from
cellular dysfunctions to immunotherapy. Nature Reviews. Immunology, 13(12), 862–74.
doi:10.1038/nri3552
Hsieh, C.-L. (2005). The de novo methylation activity of Dnmt3a is distinctly different than that
of Dnmt1. BMC Biochemistry, 6, 6. doi:10.1186/1471-2091-6-6
Hyman, S. E. (2009). How adversity gets under the skin. Nature Neuroscience, 12(3), 241–243.
doi:10.1038/nn0309-241
Jeltsch, A., & Jurkowska, R. Z. (2014). New concepts in DNA methylation. Trends in
Biochemical Sciences, 39(7), 310–318. doi:10.1016/j.tibs.2014.05.002
Jones, P. A, & Takai, D. (2001). The role of DNA methylation in mammalian epigenetics.
Science (New York, N.Y.), 293(5532), 1068–1070. doi:10.1126/science.1063852
Jones, P. A. (2012). Functions of DNA methylation: islands, start sites, gene bodies and beyond.
Nature Reviews. Genetics, 13(7), 484–92. doi:10.1038/nrg3230

82
Juruena, M. F., Pariante, C. M., Papadopoulos, A. S., Poon, L., Lightman, S., & Cleare, A. J.
(2013). The role of mineralocorticoid receptor function in treatment-resistant depression.
Journal of Psychopharmacology (Oxford, England), 27(12), 1169–79.
doi:10.1177/0269881113499205
Kanatsou, S., Kuil, L. E., Arp, M., Oitzl, M. S., Harris, A. P., Seckl, J. R., … Joels, M. (2015).
Overexpression of mineralocorticoid receptors does not affect memory and anxiety-like
behavior in female mice Article type : Received on : Accepted on : Frontiers website
link : Citation : Overexpression of mineralocorticoid receptors does not affect memo.
Frontiers in Behavioral Neuroscience, 9(July), 1–9. doi:10.3389/fnbeh.2015.00182
Kang, S. S., Kurti, A., Fair, D. a, & Fryer, J. D. (2014). Dietary intervention rescues maternal
obesity induced behavior deficits and neuroinflammation in offspring. Journal of
Neuroinflammation, 11(1), 156. doi:10.1186/s12974-014-0156-9
Karimi, M., Johansson, S., & Ekström, T. J. (2006). Using LUMA: A luminometric-based assay
for global DNA-methylation. Epigenetics, 1(1), 45–48. doi:10.4161/epi.1.1.2587
Kavushansky, A., Vouimba, R. M., Cohen, H., & Richter-Levin, G. (2006). Activity and
plasticity in the CA1, the dentate gyrus, and the amygdala following controllable vs.
uncontrollable water stress. Hippocampus, 16(1), 35–42. doi:10.1002/hipo.20130
Kigar, S. L., Chang, L., & Auger, A. P. (2015). Gadd45b is an epigenetic regulator of juvenile
social behavior and alters local pro-inflammatory cytokine production in the rodent
amygdala. Brain, Behavior, and Immunity, 46, 60–9. doi:10.1016/j.bbi.2015.02.018

83
Kimura, H., & Shiota, K. (2003). Methyl-CpG-binding protein, MeCP2, is a target molecule for
maintenance DNA methyltransferase, Dnmt1. Journal of Biological Chemistry, 278(7),
4806–4812. doi:10.1074/jbc.M209923200
Klok, M. D., Giltay, E. J., Van der Does, a J. W., Geleijnse, J. M., Antypa, N., Penninx, B. W. J.
H., … DeRijk, R. H. (2011). A common and functional mineralocorticoid receptor
haplotype enhances optimism and protects against depression in females. Translational
Psychiatry, 1(12), e62. doi:10.1038/tp.2011.59
Kohli, R. M., & Zhang, Y. (2013). TET enzymes, TDG and the dynamics of DNA
demethylation. Nature, 502(7472), 472–9. doi:10.1038/nature12750
Kolber, B. J., Roberts, M. S., Howell, M. P., Wozniak, D. F., Sands, M. S., & Muglia, L. J.
(2008). Central amygdala glucocorticoid receptor action promotes fear-associated CRH
activation and conditioning. Proceedings of the National Academy of Sciences of the
United States of America, 105(33), 12004–12009. doi:10.1073/pnas.0803216105
Koldzic-Zivanovic, N., Tu, H., Juelich, T. L., Rady, P. L., Tyring, S. K., Hudnall, S. D., …
Hughes, T. K. (2006). Regulation of adrenal glucocorticoid synthesis by interleukin-10:
A preponderance of IL-10 receptor in the adrenal zona fasciculata. Brain, Behavior, and
Immunity, 20(5), 460–468. doi:10.1016/j.bbi.2005.09.003
Kolodkin, M. H., & Auger, A. P. (2011). Sex difference in the expression of DNA
methyltransferase 3a (DNMT3a) in the rat amygdala during development. Journal of
Neuroendocrinology, 23(7), 577–583. doi:10.1111/j.1365-2826.2011.02147.x

84
Korte, S. M. (2001). Corticosteroids in relation to fear, anxiety and psychopathology.
Neuroscience and Biobehavioral Reviews, 25(2), 117–142. doi:10.1016/S0149-
7634(01)00002-1
Korte, S. M., Korte-Bouws, G. a, Koob, G. F., De Kloet, E. R., Bohus, B., Neurosciences, C., …
Laboratories, S. (1996). Mineralocorticoid and glucocorticoid receptor antagonists in
animal models of anxiety. Pharmacology, Biochemistry, and Behavior, 54(1), 261–7.
doi:0091-3057(95)02172-8 [pii]
Kossintseva, I., Wong, S., Johnstone, E., Guilbert, L., Olson, D. M., & Mitchell, B. F. (2006).
Proinflammatory cytokines inhibit human placental 11beta-hydroxysteroid
dehydrogenase type 2 activity through Ca2+ and cAMP pathways. American Journal of
Physiology. Endocrinology and Metabolism, 290(2), E282–8.
doi:10.1152/ajpendo.00328.2005
Krakowiak, P., Walker, C. K., Bremer, a. a., Baker, a. S., Ozonoff, S., Hansen, R. L., & Hertz-
Picciotto, I. (2012). Maternal Metabolic Conditions and Risk for Autism and Other
Neurodevelopmental Disorders. Pediatrics, 129(5), e1121–e1128.
doi:10.1542/peds.2011-2583
Kronfol, Z. [Ed]. (2003). Cytokines and mental health. Cytokines and Mental Health. Kluwer
Academic Publishers. Retrieved from:
http://ovidsp.ovid.com/ovidweb.cgi?T=JS&PAGE=reference&D=psyc4&NEWS=N&AN
=2003-00899-000.
Ladd, C. O., Huot, R. L., Thrivikraman, K. V, Nemeroff, C. B., & Plotsky, P. M. (2004). Long-
term adaptations in glucocorticoid receptor and mineralocorticoid receptor mRNA and

85
negative feedback on the hypothalamo-pituitary-adrenal axis following neonatal maternal
separation. Biological Psychiatry, 55(4), 367–75. doi:10.1016/j.biopsych.2003.10.007
Lai, M., Horsburgh, K., Bae, S.-E., Carter, R. N., Stenvers, D. J., Fowler, J. H., … Macleod, M.
R. (2007). Forebrain mineralocorticoid receptor overexpression enhances memory,
reduces anxiety and attenuates neuronal loss in cerebral ischaemia. The European Journal
of Neuroscience, 25(6), 1832–42. doi:10.1111/j.1460-9568.2007.05427.x
Levitt, N. S., Lindsay, R. S., Holmes, M. C., & Seckl, J. R. (1996). Dexamethasone in the last
week of pregnancy attenuates hippocampal glucocorticoid receptor gene expression and
elevates blood pressure in the adult offspring in the rat. Neuroendocrinology, 64(6), 412-
8. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/8990073
Li, M., Han, F., & Shi, Y. (2011). Expression of locus coeruleus mineralocorticoid receptor and
glucocorticoid receptor in rats under single-prolonged stress. Neurological Sciences,
32(4), 625–631. doi:10.1007/s10072-011-0597-1
Libermann, T. a, & Baltimore, D. (1990). Activation of interleukin-6 gene expression through
the NF-kappa B transcription factor. Molecular and Cellular Biology, 10(5), 2327–2334.
doi:10.1128/MCB.10.5.2327.Updated
Lillycrop, K. a, Phillips, E. S., Torrens, C., Hanson, M. a, Jackson, A. a, & Burdge, G. C. (2008).
Feeding pregnant rats a protein-restricted diet persistently alters the methylation of
specific cytosines in the hepatic PPAR alpha promoter of the offspring. The British
Journal of Nutrition, 100(2), 278–282. doi:10.1017/S0007114507894438

86
Lillycrop, K. a, Slater-Jefferies, J. L., Hanson, M. a, Godfrey, K. M., Jackson, A. a, & Burdge,
G. C. (2007). Induction of altered epigenetic regulation of the hepatic glucocorticoid
receptor in the offspring of rats fed a protein-restricted diet during pregnancy suggests
that reduced DNA methyltransferase-1 expression is involved in impaired DNA
methylation and . The British Journal of Nutrition, 97(6), 1064–73.
doi:10.1017/S000711450769196X
Louveau, A., Smirnov, I., Keyes, T. J., Eccles, J. D., Rouhani, S. J., Peske, J. D., … Kipnis, J.
(2015). Structural and functional features of central nervous system lymphatic vessels.
Nature. doi:10.1038/nature14432
MacKenzie, E. M., Odontiadis, J., Le Mellédo, J. M., Prior, T. I., & Baker, G. B. (2007). The
relevance of neuroactive steroids in schizophrenia, depression, and anxiety disorders.
Cellular and Molecular Neurobiology, 27(5), 541–574. doi:10.1007/s10571-006-9086-0
Maier, S. F., & Watkins, L. R. (2012). Consequences of the Inflamed Brain. Retrieved from The
Dana Foundation website:
http://www.dana.org/Publications/ReportOnProgress/Consequences_of_the_Inflamed_Br
ain/
Makino, S., Hashimoto, K., & Gold, P. W. (2002). Multiple feedback mechanisms activating
corticotropin-releasing hormone system in the brain during stress. Pharmacology
Biochemistry and Behavior, 73(1), 147–158. doi:10.1016/S0091-3057(02)00791-8
Mangiola, A., Vigo, V., Anile, C., Bonis, P. De, Marziali, G., & Lofrese, G. (2015). Role and
Importance of IGF-1 in Traumatic Brain Injuries, 2015.

87
Markham, J. a., & Koenig, J. I. (2011). Prenatal stress: Role in psychotic and depressive
diseases. Psychopharmacology, 214(1), 89–106. doi:10.1007/s00213-010-2035-0
Mccarthy, M. M., Auger, A. P., Bale, T. L., Vries, G. J. De, Gregory, a, Forger, N. G., …
Wilson, M. E. (2010). The Epigenetics of Sex Differences in the Brain. Journal of
Neuroscience, 29(41), 12815–12823. doi:10.1523/JNEUROSCI.3331-09.2009.
McGill, B. E., Bundle, S. F., Yaylaoglu, M. B., Carson, J. P., Thaller, C., & Zoghbi, H. Y.
(2006). Enhanced anxiety and stress-induced corticosterone release are associated with
increased Crh expression in a mouse model of Rett syndrome. Proceedings of the
National Academy of Sciences of the United States of America, 103(48), 18267–18272.
doi:10.1073/pnas.0608702103
McKee, S., Grissom, N. M., Bowman, N., & Reyes, T. M. (2014, November). Perinatal high fat
diet leads to DNMT1 deficits in the offspring prefrontal cortex: mRNA overexpression,
reduced activity and cytoplasmic sequestration. Poster presented at the Neuroscience
2014 Conference of Society for Neuroscience, Washington, DC.
McGowan, P. O., Sasaki, A., D’Alessio, A. C., Dymov, S, Labonté, B, & Szyf, M. E. A. (2010).
Epigenetic regulation of the glucocorticoid receptor in human brain associates with
childhood abuse. Nature Neuroscience, 12(3), 342–348. doi:10.1038/nn.2270.Epigenetic
Milanski, M., Degasperi, G., Coope, a., Morari, J., Denis, R., Cintra, D. E., … Velloso, L. a.
(2009). Saturated Fatty Acids Produce an Inflammatory Response Predominantly through
the Activation of TLR4 Signaling in Hypothalamus: Implications for the Pathogenesis of
Obesity. Journal of Neuroscience, 29(2), 359–370. doi:10.1523/JNEUROSCI.2760-
08.2009

88
Mikaelsson, M. A., Constância, M., Dent, C. L., Wilkinson, L. S., & Humby, T. (2013).
Placental programming of anxiety in adulthood revealed by Igf2-null models. Nature
Communications, 4(May), 2311. doi:10.1038/ncomms3311
Mitra, R., Ferguson, D., & Sapolsky, R. M. (2009). Mineralocorticoid Receptor Overexpression
in Basolateral Amygdala Reduces Corticosterone Secretion and Anxiety. Biological
Psychiatry, 66(7), 686–690. doi:10.1016/j.biopsych.2009.04.016
Mitra, R., & Sapolsky, R. M. (2008). Acute corticosterone treatment is sufficient to induce
anxiety and amygdaloid dendritic hypertrophy. Proceedings of the National Academy of
Sciences of the United States of America, 105(14), 5573–5578.
doi:10.1073/pnas.0705615105
Mueller, B. R., & Bale, T. L. (2008). Sex-specific programming of offspring emotionality after
stress early in pregnancy. The Journal of Neuroscience : The Official Journal of the
Society for Neuroscience, 28(36), 9055–9065. doi:10.1523/JNEUROSCI.1424-08.2008
Munhoz, C. D., Lepsch, L. B., Kawamoto, E. M., Malta, M. B., Lima Lde, S., Avellar, M. C.,
Sapolsky, R. M., & Scavone, C. (2006). Chronic unpredictable stress exacerbates
lipopolysaccharide-induced activation of nuclear factor- kappaB in the frontal cortex and
hippocampus via glucocorticoid secretion. Journal of Neuroscience, 26(14), 3813–3820.
doi: 10.1523/JNEUROSCI.4398-05.2006
Munhoz, C. D., Sorrells S. F., Caso J. R., Scavone C., & Sapolsky, R.M. (2010). Glucocorticoids
exacerbate lipopolysaccharide-induced signaling in the frontal cortex and hippocampus in
a dose-dependent manner. Journal of Neuroscience, 30(41), 13690–13698. doi:
10.1523/JNEUROSCI.0303-09.2010

89
Murphy, T. M., Donovan, A. O., Mullins, N., Farrelly, C. O., McCann, A., Malone, K., …
Malone, K. (2014). Anxiety is associated with higher levels of global DNA methylation
and altered expression of epigenetic and interleukin-6 genes. Psychiatric Genetics,
(August 2015), 1–8. doi:10.1097/YPG.0000000000000055
Naef, L., Gratton, A., & Walker, C.-D. (2013). Exposure to high fat during early development
impairs adaptations in dopamine and neuroendocrine responses to repeated stress. Stress
(Amsterdam, Netherlands), 3890(5), 1–9. doi:10.3109/10253890.2013.805321
Nair, A., & Bonneau, R.H. (2006). Stress-induced elevation of glucocorticoids increases
microglia proliferation through NMDA receptor activation. Journal of
Neuroimmunology, 171, 72–85. doi:10.1016/j.jneuroim.2005.09.012
Najjar, S., Pearlman, D. M., Alper, K., Najjar, A., & Devinsky, O. (2013). Neuroinflammation
and psychiatric illness. Journal of Neuroinflammation, 10, 1–24. doi:10.1186/1742-2094-
10-43
Nawa, H., & Takei, N. (2006). Recent progress in animal modeling of immune inflammatory
processes in schizophrenia: Implication of specific cytokines. Neuroscience Research.
doi:10.1016/j.neures.2006.06.002
Nelson, G., Wilde, G. J. C., Spiller, D. G., Kennedy, S. M., Ray, D. W., Sullivan, E., … White,
M. R. H. (2003). NF-kappaB signaling is inhibited by glucocorticoid receptor and STAT6
via distinct mechanisms. Journal of Cell Science, 116(Pt 12), 2495–503.
doi:10.1242/jcs.00461

90
Niehrs, C., & Schäfer, A. (2012). Active DNA demethylation by Gadd45 and DNA repair.
Trends in Cell Biology, 22(4), 220–227. doi:10.1016/j.tcb.2012.01.002
Nugent, B. M., Wright, C. L., Shetty, A. C., Hodes, G. E., Lenz, K. M., Mahurkar, A., …
Mccarthy, M. M. (2015). Brain feminization requires active repression of masculinization
via DNA methylation. Nature Neuroscience, 18(August 2014). doi:10.1038/nn.3988
O'Connor, T. G. (2002). Maternal antenatal anxiety and children’s behavioural/emotional
problems at 4 years: Report from the Avon Longitudinal Study of Parents and Children.
The British Journal of Psychiatry, 180(6), 502–508. doi:10.1192/bjp.180.6.502
O’Donovan, A., Hughes, B. M., Slavich, G. M., Lynch, L., Cronin, M.-T., O’Farrelly, C., &
Malone, K. M. (2010). Clinical anxiety, cortisol and interleukin-6: Evidence for
specificity in emotion–biology relationships. Brain, Behavior, and Immunity, 24(7),
1074–1077. doi:10.1016/j.bbi.2010.03.003
Oberlander, T. F., Weinberg, J., Papsdorf, M., Grunau, R., Misri, S., & Devlin, A. M. (2008).
Prenatal exposure to maternal depression, neonatal methylation of human glucocorticoid
receptor gene (NR3C1) and infant cortisol stress responses. Epigenetics, 3(2), 97–106.
doi:10.4161/epi.3.2.6034
Ong, Z. Y., Muhlhausler, B. S. (2011). Maternal ‘‘junk-food’’ feeding of rat dams alters food
choices and development of the mesolimbic reward pathway in the offspring. The FASEB
Journal : Official Publication of the Federation of American Societies for Experimental
Biology, 25(7), 2167–2179. doi: 10.1096/fj.10-178392

91
Peleg-Raibstein, D., Luca, E., & Wolfrum, C. (2012). Maternal high-fat diet in mice programs
emotional behavior in adulthood. Behavioural Brain Research, 233(2), 398–404.
doi:10.1016/j.bbr.2012.05.027
Pham, T. D., MacLennan, N. K., Chiu, C. T., Laksana, G. S., Hsu, J. L., & Lane, R. H. (2003).
Uteroplacental insufficiency increases apoptosis and alters p53 gene methylation in the
full-term IUGR rat kidney. American Journal of Physiology. Regulatory, Integrative and
Comparative Physiology, 285(5), R962–R970. doi:10.1152/ajpregu.00201.2003
Pilsner, J. R., Lazarus, A. L., Nam, D. H., Letcher, R. J., Sonne, C., Dietz, R., & Basu, N. (2010).
Mercury-associated DNA hypomethylation in polar bear brains via the LUminometric
Methylation Assay: A sensitive method to study epigenetics in wildlife. Molecular
Ecology, 19(2), 307–314. doi:10.1111/j.1365-294X.2009.04452.x
Postal, M., Costallat, L. T. L., & Appenzeller, S. (2011). Neuropsychiatric manifestations in
systemic lupus erythematosus: epidemiology, pathophysiology and management. CNS
Drugs, 25(9), 721–36. doi:10.2165/11591670-000000000-00000
Purcell, R. H., Sun, B., Pass, L. L., Power, M. L., Moran, T. H., & Tamashiro, K. L. (2011)
Maternal stress and high-fat diet effect on maternal behavior, milk composition, and pup
ingestive behavior. Physiology & Behavior, 104(3), 474–479. doi:
10.1016/j.physbeh.2011.05.012
Qin, C., Greenwood-Van Meerveld, B., & Foreman, R. D. (2003). Spinal neuronal responses to
urinary bladder stimulation in rats with corticosterone or aldosterone onto the amygdala.
Journal of Neurophysiology, 90(4), 2180–2189. doi:10.1152/jn.00298.2003

92
Quan, N., He, L., Lai, W., Shen, T., & Herkenham, M. (2000). Induction of IkBa mRNA
Expression in the Brain by Glucocorticoids : A Negative Feedback Mechanism for
Immune-to-Brain Signaling, 20(17), 6473–6477.
Ramamoorthy, S., & Cidlowski, J. a. (2013). Ligand-Induced Repression of the Glucocorticoid
Receptor Gene Is Mediated by an NCoR1 Repression Complex Formed by Long-Range
Chromatin Interactions with Intragenic Glucocorticoid Response Elements. Molecular
and Cellular Biology, 33(9), 1711–1722. doi:10.1128/MCB.01151-12
Ratnayake, U., Quinn, T., Walker, D. W., & Dickinson, H. (2013). Cytokines and the
neurodevelopmental basis of mental illness. Frontiers in Neuroscience, 7(7 OCT), 180.
doi:10.3389/fnins.2013.00180
Reul, J. M. H. M., Gesing, A., Droste, S., Stec, I. S. M., Weber, A., Bachmann, C., … Linthorst,
A. C. E. (2000). The brain mineralocorticoid receptor: Greedy for ligand, mysterious in
function. European Journal of Pharmacology, 405(1-3), 235–249. doi:10.1016/S0014-
2999(00)00677-4
Rhen, T., & Cidlowski, J. a. (2005). Antiinflammatory action of glucocorticoids--new
mechanisms for old drugs. The New England Journal of Medicine, 353(16), 1711–1723.
doi:10.1056/NEJMra050541
Rivera, H. M., Christiansen, K. J., & Sullivan, E. L. (2015). The role of maternal obesity in the
risk of neuropsychiatric disorders. Frontiers in Neuroscience, 9(June), 1–16.
doi:10.3389/fnins.2015.00194

93
Robson, P. J., Blannin, A. K., Walsh, N. P., Castell, L. M., & Gleeson, M. (1999). Effects of
exercise intensity, duration and recovery on in vitro neutrophil function in male athletes.
International Journal of Sports Medicine, 20(2), 128–35. doi:10.1055/s-2007-971106
Rodriguez, A. (2010). Maternal pre-pregnancy obesity and risk for inattention and negative
emotionality in children. Journal of Child Psychology and Psychiatry, and Allied
Disciplines, 51(2), 134–143. doi: 10.1111/j.1469-7610.2009.02133.x
Rohleder, N., Schommer, N. C., Hellhammer, D. H., Engel, R., & Kirschbaum, C. (2001). Sex
differences in glucocorticoid sensitivity of proinflammatory cytokine production after
psychosocial stress. Psychosomatic Medicine, 63(6), 966–72. Retrieved from
http://www.ncbi.nlm.nih.gov/pubmed/11719636
Rozeboom, A. M., Akil, H., & Seasholtz, A. F. (2007). Mineralocorticoid receptor
overexpression in forebrain decreases anxiety-like behavior and alters the stress response
in mice. Proceedings of the National Academy of Sciences of the United States of
America, 104(11), 4688–4693. doi:10.1073/pnas.0606067104
Samaco, R. C., Mandel-Brehm, C., McGraw, C. M., Shaw, C. a, McGill, B. E., & Zoghbi, H. Y.
(2012). Crh and Oprm1 mediate anxiety-related behavior and social approach in a mouse
model of MECP2 duplication syndrome. Nature Genetics, 44(2), 206–211.
doi:10.1038/ng.1066
Saraiva, M., & O’Garra, A. (2010). The regulation of IL-10 production by immune cells. Nature
Reviews. Immunology, 10(3), 170–181. doi:10.1038/nri2711

94
Sarrazin, N., Di Blasi, F., Roullot-Lacarrière, V., Rougé-Pont, F., Le Roux, A., Costet, P., …
Piazza, P. V. (2009). Transcriptional effects of glucocorticoid receptors in the dentate
gyrus increase anxiety-related behaviors. PloS One, 4(11), e7704.
doi:10.1371/journal.pone.0007704
Sasaki, A., de Vega, W. C., St-Cyr, S., Pan, P., & McGowan, P. O. (2013). Perinatal high fat diet
alters glucocorticoid signaling and anxiety behavior in adulthood. Neuroscience, 240, 1–
12. doi:10.1016/j.neuroscience.2013.02.044
Sasaki, A., Erb, S., & McGowan, P. O. (2015). The effect of maternal overnutrition on reward
and anxiety in offspring. Manuscript submitted for publication.
Schaefer, C. a, Brown, a S., Wyatt, R. J., Kline, J., Begg, M. D., Bresnahan, M. a, & Susser, E.
S. (2000). Maternal prepregnant body mass and risk of schizophrenia in adult offspring.
Schizophrenia Bulletin, 26(2), 275–86. Retrieved from
http://www.ncbi.nlm.nih.gov/pubmed/10885630
Schmitt, A., Malchow, B., Hasan, A., & Falkai, P. (2014). The impact of environmental factors
in severe psychiatric disorders. Frontiers in Neuroscience, 8(February), 1–10.
doi:10.3389/fnins.2014.00019
Schwarz, J. M., Hutchinson, M. R., & Bilbo, S. D. (2011). Early-Life Experience Decreases
Drug-Induced Reinstatement of Morphine CPP in Adulthood via Microglial-Specific
Epigenetic Programming of Anti-Inflammatory IL-10 Expression. Journal of
Neuroscience, 31(49), 17835–17847. doi:10.1523/JNEUROSCI.3297-11.2011

95
Scotland, R. S., Stables, M. J., Madalli, S., Watson, P., & Gilroy, D. W. (2011). Sex differences
in resident immune cell phenotype underlie more efficient acute inflammatory responses
in female mice. Blood, 118(22), 5918–27. doi:10.1182/blood-2011-03-340281
Shapero, B. G., Black, S. K., Liu, R. T., Klugman, J., Bender, R. E., Abramson, L. Y., & Alloy,
L. B. (2014). Stressful life events and depression symptoms: the effect of childhood
emotional abuse on stress reactivity. Journal of Clinical Psychology, 70(3), 209–23.
doi:10.1002/jclp.22011
Sharma, D., Bhave, S., Gregg, E., & Uht, R. (2013). Dexamethasone induces a putative repressor
complex and chromatin modifications in the CRH promoter. Molecular Endocrinology
(Baltimore, Md.), 27(November), 1142–52. doi:10.1210/me.2013-1079
Simmons, R. (2008). Perinatal programming of obesity. Seminars in Perinatology, 32(5), 371–4.
doi:10.1053/j.semperi.2008.08.004
Smith, E. M., Cadet, P., Stefano, G. B., Opp, M. R., & Hughes, T. K. Jr. (1999). IL-10 as a
mediator in the HPA axis and brain. Journal of Neuroimmunology, 100, 140-8.
Smith, S. M., & Vale, W. W. (2006). The role of the hypothalamic-pituitary-adrenal axis in
neuroendocrine responses to stress. Dialogues in Clinical Neuroscience, 8(4), 383–395.
doi:10.1038/nrendo.2011.222
Smythe, J. W., Murphy, D., Timothy, C., & Costall, B. (1997). Hippocampal mineralocorticoid,
but not glucocorticoid, receptors modulate anxiety-like behavior in rats. Pharmacology,
Biochemistry, and Behavior, 56(3), 507–513.

96
Sorrells, S. F., Caso, J. R., Munhoz, C. D., & Sapolsky, R. M. (2009). The stressed CNS: when
glucocorticoids aggravate inflammation. Neuron, 64(1), 33–9.
doi:10.1016/j.neuron.2009.09.032
Stein-Behrens, B., Mattson, M. P., Chang, I., Yeh, M., & Sapolsky, R.. (1994) Stress exacerbates
neuron loss and cytoskeletal pathology in the hippocampus. Journal of Neuroscience, 14,
5373–5380.
Strohle, A., & Holsboer, F. (2003). Stress responsive neurohormones in depression and anxiety.
Pharmacopsychiatry, 36 Suppl 3, S207–14. doi:10.1055/s-2003-45132
Sullivan, E. L., Grayson, B., Takahashi, D., Robertson, N., Maier, A., Bethea, C. L., Smith,
M.S., Coleman, K., & Grove, K. L. (2010) Chronic consumption of a high-fat diet during
pregnancy causes perturbations in the serotonergic system and increased anxiety-like
behavior in nonhuman primate offspring. Journal of Neuroscience, 30(10), 3826–30. doi:
10.1523/JNEUROSCI.5560-09.2010
Sullivan, E. L., Riper, K. M., Lockard, R., & Valleau, J. C. (2015). Maternal high-fat diet
programming of the neuroendocrine system and behavior. Hormones and Behavior.
doi:10.1016/j.yhbeh.2015.04.008
Suzuki, M. M., & Bird, A. (2008). DNA methylation landscapes: provocative insights from
epigenomics. Nature Reviews. Genetics, 9(6), 465–76. doi:10.1038/nrg2341
Svennerholm, L., & Vanier, M. T. (1978). Lipid and fatty acid composition of human cerebral
myelin during development. Advances in experimental medicine and biology, 100, 27–41.
Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/696475

97
Tamashiro, K. L., Terrillion, C. E., Hyun, J., Koenig, J. I., & Moran, T. H. (2009). Prenatal stress
or high-fat diet increases susceptibility to diet- induced obesity in rat offspring. Diabetes,
58(5), 1116–1125. doi: 10.2337/db08-1129
Teicher, M. H., & Samson, J. A. (2013). Childhood Maltreatment and Psychopathology: A Case
for Ecophenotypic Variants as Clinically and Neurobiologically Distinct Subtypes.
American Journal of Psychiatry, 170(10), 1114. doi:10.1176/appi.ajp.2013.12070957
Ter Heegde, F., De Rijk, R. H., & Vinkers, C. H. (2014). The brain mineralocorticoid receptor
and stress resilience. Psychoneuroendocrinology, 52, 92–110.
doi:10.1016/j.psyneuen.2014.10.022
Theisen, J. W. M., Gucwa, J. S., Yusufzai, T., Khuong, M. T., & Kadonaga, J. T. (2013).
Biochemical analysis of histone deacetylase-independent transcriptional repression by
MeCP2. The Journal of Biological Chemistry, 288(10), 7096–104.
doi:10.1074/jbc.M112.438697
Tronche, F., Kellendonk, C., Kretz, O., Gass, P., Anlag, K., Orban, P. C., … Schütz, G. (1999).
Disruption of the glucocorticoid receptor gene in the nervous system results in reduced
anxiety. Nature Genetics, 23(1), 99–103. doi:10.1038/12703
Tu, H., Rady, P. L., Juelich, T., Tyring, S. K., Koldzic-Zivanovic, N., Smith, E. M., & Hughes,
T. K. (2007). Interleukin-10 regulated gene expression in cells of hypothalamic-pituitary-
adrenal axis origin. Cellular and Molecular Neurobiology, 27(2), 161–170.
doi:10.1007/s10571-006-9137-6

98
Urb, M., Anier, K., Matsalu, T., Aonurm-Helm, A., & Timmusk, T. (2015). Corticosterone
induces DNA methyltransferases expression in rat cortical neurons. SpringerPlus,
4(Suppl 1), P50. doi:10.1186/2193-1801-4-S1-P50
Van den Bergh, B. R. H., Mennes, M., Stevens, V., Van Der Meere, J., Börger, N., Stiers, P., …
Lagae, L. (2006). ADHD deficit as measured in adolescent boys with a continuous
performance task is related to antenatal maternal anxiety. Pediatric Research, 59(1), 78–
82. doi:10.1203/01.pdr.0000191143.75673.52
Van Lieshout, R. J., Robinson, M., & Boyle, M. H. (2013). Maternal pre-pregnancy body mass
index and internalizing and externalizing problems in offspring. Canadian Journal of
Psychiatry. Revue Canadienne de Psychiatrie, 58(3), 151–9. Retrieved from
http://www.ncbi.nlm.nih.gov/pubmed/23461886
Van Winkel, R., Stefanis, N. C., & Myin-Germeys, I. (2008). Psychosocial stress and psychosis.
A review of the neurobiological mechanisms and the evidence for gene-stress interaction.
Schizophrenia Bulletin, 34(6), 1095–105. doi:10.1093/schbul/sbn101
Vandevyver, S., Dejager, L., Van Bogaert, T., Kleyman, A., Liu, Y., Tuckermann, J., & Libert,
C. (2012). Glucocorticoid receptor dimerization induces MKP1 to protect against TNF-
induced inflammation. Journal of Clinical Investigation, 122(6), 2130–2140.
doi:10.1172/JCI60006
Varese, F., Smeets, F., Drukker, M., Lieverse, R., Lataster, T., Viechtbauer, W., … Bentall, R. P.
(2012). Childhood Adversities Increase the Risk of Psychosis: A Meta-analysis of
Patient-Control, Prospective- and Cross-sectional Cohort Studies. Schizophrenia Bulletin,
38(4), 661–671. doi:10.1093/schbul/sbs050

99
Voorhees, J. L., Tarr, A. J., Wohleb, E. S., Godbout, J. P., Mo, X., Sheridan, J. F., … Marsh, C.
B. (2013). Prolonged Restraint Stress Increases IL-6, Reduces IL-10, and Causes
Persistent Depressive-Like Behavior That Is Reversed by Recombinant IL-10. PLoS
ONE, 8(3). doi:10.1371/journal.pone.0058488
Wang, X., Athayde, N., & Trudinger, B. (2003). A proinflammatory cytokine response is present
in the fetal placental vasculature in placental insufficiency. American Journal of
Obstetrics and Gynecology, 189(5), 1445–1451. doi:10.1067/S0002-9378(03)00652-5
Weaver, I. C. G., Cervoni, N., Champagne, F. a, D’Alessio, A. C., Sharma, S., Seckl, J. R., …
Meaney, M. J. (2004). Epigenetic programming by maternal behavior. Nature
Neuroscience, 7(8), 847–854. doi:10.1038/nn1276
Weaver, I. C. G., D’Alessio, A. C., Brown, S. E., Hellstrom, I. C., Dymov, S., Sharma, S., …
Meaney, M. J. (2007). The transcription factor nerve growth factor-inducible protein a
mediates epigenetic programming: altering epigenetic marks by immediate-early genes.
The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 27(7),
1756–1768. doi:10.1523/JNEUROSCI.4164-06.2007
Weinstock, M. (2005). The potential influence of maternal stress hormones on development and
mental health of the offspring. Brain, Behavior, and Immunity, 19(4), 296–308.
doi:10.1016/j.bbi.2004.09.006
Weinstock, M. (2008). The long-term behavioural consequences of prenatal stress. Neuroscience
and Biobehavioral Reviews, 32(6), 1073–86. doi:10.1016/j.neubiorev.2008.03.002

100
Welberg, L. a M., Seckl, J. R., & Holmes, M. C. (2000). Inhibition of 11β-hydroxysteroid
dehydrogenase, the foetoplacental barrier to maternal glucocorticoids, permanently
programs amygdala GR mRNA expression and anxiety-like behaviour in the offspring.
European Journal of Neuroscience, 12(3), 1047–1054. doi:10.1046/j.1460-
9568.2000.00958.x
Welberg, L. a M., Seckl, J. R., & Holmes, M. C. (2001). Prenatal glucocorticoid programming of
brain corticosteroid receptors and corticotrophin-releasing hormone: possible
implications for behaviour. Neuroscience, 104(1), 71–79. doi:10.1016/S0306-
4522(01)00065-3
White, C. L., Pistell, P. J., Purpera, M. N., Gupta, S., Fernandez-Kim, S.-O., Hise, T. L., …
Bruce-Keller, A. J. (2009). Effects of high fat diet on Morris maze performance,
oxidative stress, and inflammation in rats: contributions of maternal diet. Neurobiology of
Disease, 35(1), 3–13. doi:10.1016/j.nbd.2009.04.002
Williamson, L. L., Sholar, P. W., Mistry, R. S., Smith, S. H., & Bilbo, S. D. (2011). Microglia
and Memory: Modulation by Early-life Infection. The Journal of Neuroscience : The
Official Journal of the Society for Neuroscience, 31(43), 15511–15521.
doi:10.1523/JNEUROSCI.3688-11.2011
Zaretsky, M. V, Alexander, J. M., Byrd, W., & Bawdon, R. E. (2004). Transfer of inflammatory
cytokines across the placenta. Obstetrics and Gynecology, 103(3), 546–550.
doi:10.1097/01.AOG.0000114980.40445.83
Ziaja, M. (2013). Septic Encephalopathy. Current Neurology and Neuroscience Reports, 13(10),
383. doi:10.1007/s11910-013-0383-y

101
Appendix
Supplementary Table 1. Transcriptional response to CORT challenge in the amygdala (post-
hocs). Fisher's PLSD post-hocs of qPCR gene expression data separated by (a) males and (b)
females. CORT-induced anti-inflammatory signaling is exacerbated in Perinatal HFD-exposed
groups. # p < 0.1, * p < 0.05, ** p < 0.01. Shaded boxes: Diet x Drug interaction p < 0.05.
a) Males
b) Females
Function Gene Effect of CORT within diet group Effect of diet within drug group
GC signalling GR CHD CTL > CHD CORT ** HFD CORT > CHD CORT *
MR CHD CTL > CHD CORT * HFD CTL < CHD CTL #
CRH HFD CTL < HFD CORT # HFD CORT > CHD CORT *
Pro-inflammatory CD11b HFD CTL < CHD CTL #
NFkB CTL > CORT, HFD + CHD #
IL-6 CTL < CORT, HFD + CHD **
IL-6/IL-10 CTL < CORT, HFD + CHD **
Anti-inflammatory IGF-1 CHD CTL > CHD CORT # HFD CORT > CHD CORT #
IkBa CTL < CORT, HFD + CHD ** HFD CORT > CHD CORT **
MKP-1 HFD CTL < HFD CORT ** HFD CORT > CHD CORT *
IL-10 CHD CTL > CHD CORT #
Epigenetic regulation DNMT1 CHD CTL > CHD CORT * HFD CORT > CHD CORT **
DNMT3a CTL > CORT, HFD + CHD **
DNMT3b
MeCP2 CHD CTL > CHD CORT # HFD CORT > CHD CORT *
Gadd45b HFD CTL < HFD CORT # HFD CORT > CHD CORT #
Function Gene Effect of CORT within diet group Effect of diet within drug group
GC signalling GR HFD CTL > HFD CORT *
MR
CRH
Pro-inflammatory CD11b
NFkB
IL-6
IL-6/IL-10 HFD CTL < HFD CORT *
Anti-inflammatory IGF-1
IkBa HFD CTL < HFD CORT *
MKP-1
IL-10
Epigenetic regulation DNMT1
DNMT3a
DNMT3b
MeCP2
Gadd45b

102
Supplementary Table 2. Transcriptional response to CORT challenge in the hippocampus
(post-hocs). Fisher's PLSD post-hocs of qPCR gene expression data separated by (a) males and
(b) females. CORT-induced anti-inflammatory signaling is exacerbated in Perinatal HFD-
exposed groups. # p < 0.1, * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001. Shaded
boxes: Diet x Drug interaction p < 0.05.
a) Males
b) Females
Function Gene Effect of CORT within diet group Effect of diet within drug group
GC signalling GR CHD CTL > CHD CORT * HFD CORT > CHD CORT *
MR CHD CTL > CHD CORT * HFD CORT > CHD CORT #
Pro-inflammatory CD11b
NFkB
IL-6 CTL < CORT, CHD ***, HFD * HFD CTL > CHD CTL #
IL-6/IL-10 CHD CTL < CHD CORT **
Anti-inflammatory IGF-1
IkBa CTL < CORT, CHD ****, HFD ** HFD CORT > CHD CORT *
MKP-1 CTL < CORT, CHD ****, HFD ** HFD CORT > CHD CORT *
IL-10
Epigenetic regulation DNMT1
DNMT3a
MeCP2
Gadd45b
Function Gene Effect of CORT within diet group Effect of diet within drug group
GC signalling GR CHD CTL > CHD CORT ** HFD CTL < CHD CTL *
MR
Pro-inflammatory CD11b CHD CTL > CHD CORT ** HFD CTL < CHD CTL *
NFkB CHD CTL > CHD CORT ** HFD CTL < CHD CTL *
IL-6 HFD CTL < HFD CORT #
IL-6/IL-10 HFD CTL < HFD CORT #
Anti-inflammatory IGF-1
IkBa HFD CTL < HFD CORT ** HFD CTL < CHD CTL, HFD CORT > CHD CORT *
MKP-1 CHD CTL > CHD CORT *, HFD CTL < HFD CORT # HFD CTL < CHD CTL, HFD CORT > CHD CORT #
IL-10
Epigenetic regulation DNMT1 CHD CTL > CHD CORT **
DNMT3a CHD CTL > CHD CORT, CHD **, HFD #
MeCP2 CHD CTL > CHD CORT, CHD ***, HFD * HFD CORT > CHD CORT #
Gadd45b CHD CTL > CHD CORT *

103
Supplementary Table 3. Transcriptional response to LPS challenge in the amygdala (post-
hocs). Fisher's PLSD post-hocs of qPCR gene expression data separated by (a) males and (b)
females. LPS-induced anti-inflammatory signaling is exacerbated in perinatal HFD-exposed
groups. # p < 0.1, * p < 0.05, ** p < 0.01, *** p < 0.001.
a) Males
b) Females
Function Gene Effect of LPS within diet group Effect of diet within drug group
GC signalling GR
MR CHD CTL > CHD LPS #
Pro-inflammatory CD11b HFD CTL < CHD CTL *
NFkB
IL-6 CTL < LPS, HFD + CHD *
IL-6/IL-10 CTL < LPS, CHD **, HFD *
Anti-inflammatory IGF-1 CHD CTL < CHD LPS *
IkBa CTL < LPS, HFD **, CHD *
MKP-1 HFD CTL < HFD LPS *
IL-10 CTL > LPS, HFD ***, CHD *
Function Gene Effect of LPS within diet group Effect of diet within drug group
GC signalling GR CHD CTL > CHD LPS #
MR CTL < LPS, HFD + CHD #
Pro-inflammatory CD11b CHD CTL > CHD LPS *
NFkB HFD LPS < CHD LPS *
IL-6 CTL < LPS, HFD **, CHD #
IL-6/IL-10 CTL < LPS, HFD + CHD **
Anti-inflammatory IGF-1
IkBa HFD CTL < HFD LPS *
MKP-1
IL-10

104
Supplementary Table 4. Transcriptional response to CORT/LPS challenge in the amygdala
(post-hocs). Fisher's PLSD post-hocs of qPCR gene expression data separated by (a) males and
(b) females. CORT/LPS-induced pro-inflammatory response is altered in Perinatal HFD-exposed
groups. # p < 0.1, * p < 0.05, ** p < 0.01, *** p < 0.001. Shaded boxes: Diet x Drug interaction
p < 0.05.
a) Males
b) Females
Function Gene Effect of CORT/LPS within diet group Effect of diet within drug group
GC signalling GR CTL > CORT/LPS, HFD *, CHD ***
MR CHD CTL > CHD CORT/LPS ** HFD CTL < CHD CTL *
CRH CHD CTL < CHD CORT/LPS **
Pro-inflammatory CD11b
NFkB CTL > CORT/LPS, CHD ***, HFD * HFD CORT/LPS > CHD CORT/LPS *
IL-6 CTL < CORT/LPS, HFD ***, CHD **
IL-6/IL-10 CTL < CORT/LPS ** HFD CORT/LPS < CHD CORT/LPS #
Anti-inflammatory IGF-1 CTL > CORT/LPS, CHD **, HFD *
IkBa CTL < CORT/LPS, HFD + CHD **
MKP-1
IL-10 CTL > CORT/LPS, CHD **, HFD *
Epigenetic regulation DNMT1 CHD CTL > CHD CORT/LPS *
DNMT3a CHD CTL > CHD CORT/LPS #
DNMT3b
MeCP2
Gadd45b CHD CTL < CHD CORT/LPS **
Function Gene Effect of CORT/LPS within diet group Effect of diet within drug group
GC signalling GR CTL > CORT/LPS, HFD **, CHD * HFD CORT/LPS < CHD CORT/LPS **
MR CTL > CORT/LPS, CHD **, HFD * HFD CORT/LPS < CHD CORT/LPS **
CRH CHD CTL < CHD CORT/LPS ** HFD CORT/LPS < CHD CORT/LPS **
Pro-inflammatory CD11b CTL > CORT/LPS, HFD **, CHD * HFD CORT/LPS < CHD CORT/LPS *
NFkB CTL > CORT/LPS, HFD **, CHD * HFD CORT/LPS < CHD CORT/LPS #
IL-6
IL-6/IL-10 CTL < CORT/LPS, HFD + CHD ** HFD CORT/LPS < CHD CORT/LPS #
Anti-inflammatory IGF-1 CHD CTL > CHD CORT/LPS #
IkBa
MKP-1 HFD CTL > HFD CORT/LPS *
IL-10 CTL > CORT/LPS, HFD + CHD **
Epigenetic regulation DNMT1 HFD CTL > HFD CORT/LPS * HFD CORT/LPS < CHD CORT/LPS *
DNMT3a HFD CTL > HFD CORT/LPS # HFD CORT/LPS < CHD CORT/LPS *
DNMT3b HFD CTL > HFD CORT/LPS *
MeCP2
Gadd45b HFD CTL < CHD CTL #