markers and methods to verify mesenchymal stem cell ... · markers and methods to verify...
TRANSCRIPT

RnDSy-lu-2945
Scott Schachtele, Ph.D., Christine Clouser, Ph.D., and Joy Aho, Ph.D.
Markers and Methods to Verify Mesenchymal Stem Cell Identity, Potency, and Quality
WHITE PAPER
ABSTRACTMesenchymal stem cells (MSCs) are multipotent cells that are functionally defined by their capacity to self-renew and their ability to differentiate into multiple cell types including adipocytes, chondrocytes, and osteocytes. Translation of MSC-based therapies has been confounded by MSC population heterogeneity as well as non-standardized methods for their definition and characterization. This white paper begins by defining and discussing the evolution of MSC nomenclature. It then provides a detailed description of MSC markers and how they may vary by tissue source and species. Ultimately, this review discusses how to standardize MSC characterization by selecting markers for isolation, characterization, and validation.
INTRODUCTIONMesenchymal stem cells (MSCs) are multipotent cells that adhere to plastic, have a fibroblast-like morphology, express a specific set of surface antigens, and differentiate into adipocytes, chondrocytes, and osteocytes.1 Clinically, MSCs are of interest for their ability to modulate the immune system as well as their potential to regenerate tissues. However, the translation of MSC-based therapies has been hindered by the heterogeneity of the isolated cells as well as the lack of standardized methods for their definition and characterization. At the forefront of these issues is the lack of a MSC-specific marker that can be used to definitively isolate and characterize MSCs. The absence of a MSC-specific marker limits the purity of MSCs isolated by methods such as positive and negative selection. As a result, MSCs are typically isolated by plastic adherence, which generates a heterogeneous population of cells that differ in their growth kinetics and differentiation potential. This heterogeneity can increase experimental variability, decrease the efficiency of differentiation, and contribute to conflicting data in the literature. In an attempt to increase consistency in the definition and verification of MSCs, the International Society for Cellular Therapy (ISCT) proposed minimal criteria for the definition of human MSCs based on plastic adherence, multipotency, and the expression of a panel of surface antigens (Box 1).1 Since proposing the minimal criteria in 2006, several advances in MSC characterization and marker identification have been made. These advances have led to improved methods for isolating, verifying, and assessing the quality of MSCs. This review summarizes the most recent MSC markers, definitions, and the latest methods used to characterize MSCs.
KEY TOPICS• Defining Mesenchymal Stem Cells
• Established and New MSC Markers
• Methods for MSC Isolation
• MSC Marker Variation by Species
• Methods for MSC Identification and Characterization
• Media Definitions for MSC Expansion
Box 1. Minimal Experimental Criteria for MSC as proposed by the International Society for Cellular Therapy
• Adherence to plastic.
• ≥95% of the MSC population must express CD73/5′-Nucleotidase, CD90/Thy1, and CD105/Endoglin as measured by flow cytometry.
• ≤2% of the MSC population must express CD34, CD45, CD11b/Integrin alpha M or CD14, CD79 alpha or CD19, and HLA Class II.
• Multipotent differentiation potential as demonstrated by staining of in vitro differentiated cells.

Learn more | rndsystems.com/msc
DEFINING MESENCHYMAL STEM CELLSThe in vivo identity and characteristics as well as the therapeutic potential of MSCs have been debated since their discovery by Alexander Friedenstein in 1968. As a result, several names have been proposed to reflect the changing understanding of MSC properties (Figure 1). However, newly proposed nomenclature for MSCs is not universally adopted by the field, resulting in the use of several different terms to describe the same cell type (Figure 1). While “mesenchymal stem cell” is commonly used to describe multipotent cells that express specific surface antigens and adhere to plastic, some scientists argue that the definition is vague, encompasses cells isolated from virtually every tissue, and does not necessarily predict the stem cell properties of the cells in an in vivo setting.2
Because MSCs isolated from bone marrow have been shown to self-renew and regenerate skeletal tissue in vivo, some scientists suggest that these cells are more accurately defined by the term “skeletal stem cell.”3,4,5 Alternatively, the term “medicinal signaling cell” has been proposed as a replacement for “mesenchymal stem cell” as a way to indicate that the primary therapeutic benefit of MSCs may be attributed to secretion of bioactive molecules.6
In this review, the term “mesenchymal stromal cells” is used to describe a heterogeneous population of cells that are adherent to plastic, have a fibroblast-like morphology, and express a specific set of marker proteins. Within this heterogeneous population are a subset of cells referred to as “mesenchymal stem cells,” which are multipotent, self-renewing cells that have the ability to differentiate into adipocytes, chondrocytes, and osteoblasts when cultured in vitro. In addition to the conceptual definitions of MSCs, the ISCT offers an experimental definition of MSCs (Box 1) to facilitate identification of human MSCs.
The controversy over the definition and nomenclature of MSCs centers in part on the minimal criteria proposed by the ISCT. According to the minimal criteria, MSCs are not restricted to bone marrow but can be isolated from a wide range of tissues.
The definition also suggests that MSCs isolated from different tissues are phenotypically and functionally equivalent, which would imply that the cells share the same regenerative capabilities in vivo. Thus, MSCs isolated from bone marrow and MSCs isolated from placenta would be expected to have the same ability to regenerate skeletal tissue in vivo. Some scientists argue that this assumption is unfounded and that the defining features of MSCs, as proposed by the ISCT, are little more than characteristics shared by connective tissues and
perivascular cells.2 Therefore, MSCs isolated from adipose tissues may not demonstrate the same regenerative properties in vivo as MSCs isolated from bone marrow. In fact, it has been proposed that MSCs are perivascular cells that are lineage-restricted, tissue-specific progenitors and that the local environment and perivascular position could account for the expression of common antigens.2 If this hypothesis is correct, MSCs isolated from bone marrow would have the ability to regenerate skeletal tissue in vivo, while adipose-derived MSCs would be restricted to the regeneration of adipose tissue in vivo. These issues emphasize the controversies surrounding the nomenclature of MSCs and such controversies will persist in the field until experiments are performed to assess the in vivo properties of MSCs isolated from different tissues sources.
The defining features of MSCs, as proposed by the ISCT. are little more than characteristics shared by connective tissues and perivascular cells.
Figure 1. The History and Timeline of Mesenchymal Stem Cell Nomenclature. MSCs were originally identified by Alexander Friedenstein as colony forming unit fibroblasts (CFU-F) and osteogenic stem cells.7,8 However, since this time, a number of terms have been used to describe mesenchymal stem cells (MSCs) including “stromal stem cells,” which was proposed in 1988 by Maureen Owen, Ph.D. and colleagues to indicate that the cells reside in the stromal rather than the hematopoietic compartment. Rather than highlight the cells’ compartmental origin, Arnold Caplan, Ph.D. proposed the term “mesenchymal stem cells” in 1991 to emphasize the self-renewal property and differentiation potential of the cells.9 However, this nomenclature was challenged by James Dennis, Ph.D. and colleagues who suggested that the cells may be progenitors rather than stem cells. As a result, the term “mesenchymal progenitor cells” was proposed.”10 In 2000, Paolo Bianco, M.D. and Pamela Gehron Robey, Ph.D. coined “skeletal stem cell” to specify that the cells give rise to components of the skeletal system, while just two years later, the term “multipotent adult progenitor cells” was proposed by Yuehua Jiang, M.D. and colleagues to describe the multipotent nature and potential progenitor status of the cells.3,11 Since no direct evidence demonstrated the ability of MSCs to self-renew and differentiate in vivo, in 2006, the International Society for Cellular Therapy proposed the term “multipotent mesenchymal stromal cells.”1 More recently, in 2010, Arnold Caplan, Ph.D. suggested that the acronym MSC stand for “medicinal signaling cells” to reflect that the primary therapeutic benefit of MSCs may be attributed to the secretion of bioactive molecules rather than tissue regeneration.6
History and Timeline of Mesenchymal Stem Cell Nomenclature
CFU-F andOsteogenic Stem Cells
(Friedenstein)
Stromal Stem Cells(Maureen Owen and colleagues)
Mesenchymal Progenitor Cells(James Dennis and colleagues)
Multipotent Adult Progenitor Cellsand Mesodermal Progenitor Cells
(Yuehua Jiang and colleagues)
Medicinal Signaling Cells(Arnold Caplan)
Mesenchymal Stem Cells(Arnold Caplan)
Skeletal Stem Cells(Paolo Bianco and Pamela Gehron Robey)
Multipotent MesenchymalStromal Cells
(ISCT)
1991
1999
2000
2002
2006
1988 2010
1970s
© R&D Systems, Inc.
For the purpose of this review, the term “Mesenchymal Stem Cells” (MSCs) refers to multipotent, self-renewing cells that express a specific set of surface antigens and can differentiate into adipocytes, chondrocytes, and osteocytes when cultured in vitro.

MARKERS FOR MESENCHYMAL STEM CELL ISOLATION AND VERIFICATIONIn 1970, Friedenstein and colleagues described a method for isolating fibroblast-like colonies from rodent bone marrow.7 These clonogenic cells were isolated based on their relatively high adhesion to plastic compared to other cells in the bone marrow compartment. The method used by Friedenstein is still widely used today for isolation of MSCs from unpurified bone marrow or Ficoll™-purified bone marrow mononuclear cells.
While isolation by plastic adherence is efficient, it does not yield a homogeneous population of cells. The isolated cells demonstrate different growth kinetics and differentiation capacities and include a small population of contaminating hematopoietic cells. Cell homogeneity can be increased over several weeks by culturing under conditions that preferentially expand MSCs and select against expansion of hematopoietic cells.12 However, the remaining MSCs can still demonstrate a range of proliferation rates and differen tiation potentials. This heterogeneity may contribute to experimental variability, contradictory data, and poor pre-clinical outcomes.
Established and New Markers for MSC Identification and VerificationBefore the ISCT published the minimal criteria for defining MSCs in 2006 (Box 1), variability in the methods used to isolate, expand, and characterize MSCs made it difficult to compare data from different labs. The ISCT criteria offered a quality control step for researchers to verify MSC identity, which decreased experimental variability and increased the transparency of published data.
The ISCT proposed positive and negative markers that enabled researchers to distinguish MSCs from other cells in the bone marrow compartment. The negative markers were selected to include surface antigens that are expressed by hematopoietic cells, while the positive markers were selected to include surface antigens that are absent from most hematopoietic cells (Table 1). However, ISCT acknowledges that the criteria must be met with some flexibility, particularly as they relate to expression of the negative marker, HLA Class II. Specifically, HLA Class II can be expressed by MSCs under certain conditions, such as cytokine stimulation.1 Therefore, cells that meet all other criteria, but are also positive for HLA Class II, can be designated as MSCs if an adjective is used to indicate that the cells were stimulated. According to the ISCT, CD34 is a negative marker of MSCs. However, some reports suggest that the CD34 negative status is an artifact of cell culture conditions.13,14,15 In fact, several groups have shown that MSCs isolated from adipose tissue express CD34 at the time of isolation but lose expression while in culture.16,17,18 Expression of CD34 by MSCs is also supported by the fact that the STRO-1 antibody (Clone STRO-1), which is commonly used to identify MSCs, was developed using CD34+ bone marrow as the immunogen.19 The debate over the use of CD34 as a negative marker raises the possibility that markers may vary depending on the MSC tissue source.
Learn more | rndsystems.com/msc
Table 1: Negative and Positive MSC Markers as Proposed by the International Society for Cell Therapy (ISCT)
Negative HSC Marker Used to Exclude Peer-reviewed References
CD3420,21 Primitive hematopoietic cells and endothelial cells22 23, 24
CD4520,21 Leukocytes25 26, 27
CD11b and CD1420,21 Monocytes and macrophages28,29 27, 30
CD79 alpha and CD19 alpha31 B cells32 33, 34
HLA Class II31 Antigen presenting cells and lymphocytes1,35* 36, 37
Positive MSC Markers Biological Role Peer-reviewed References
CD73/5′-Nucleotidase21 Catalyzes production of extracellular adenosine from AMP38 39, 40
CD90/Thy120 Wound repair, cell-cell and cell-matrix interactions41; see Figure 2
27, 42
CD105/Endoglin21 Vascular homeostasis; modulates TGF-beta functions via interaction with TGF-beta RI and TGF-beta RII43
24, 36, 44
*HLA Class II may be expressed on stimulated MSCs.
Figure 2. Detection of CD90/Thy1 in Human Mesenchymal Stem Cells by Flow Cytometry. Human mesenchymal stem cells were stained with Sheep Anti-Human/Porcine/Canine CD90/Thy1 Antigen Affinity-purified Polyclonal Antibody (Catalog # AF2067, filled histogram) or isotype control antibody (Catalog # 5-001-A, open histogram), followed by PE-conjugated Anti-Sheep IgG Secondary Antibody (Catalog # F0126).
101 102 103 104
20
40
100
Rela
tive
Cell
Num
ber
1000
80
60
CD90/Thy1

THE LATEST MARKERS USED TO VERIFY MSC IDENTITYSince the ISCT-proposed markers were published in 2006, several additional markers have become widely accepted as MSC markers. While no markers have been identified that are exclusively expressed by MSCs, the growing panel of MSC-associated markers enables researchers to simultaneously verify expression of several MSC-associated surface antigens, which increases the confidence in the identification and verification of isolated MSCs.
STRO-1The STRO-1 monoclonal antibody was the first antibody that was successfully used to enrich Colony-forming Unit-Fibroblasts (CFU-Fs) from human bone marrow.19,45 While STRO-1 is a widely used marker of MSCs, little is known about the identity of its cognate antigen. In addition to MSCs, STRO-1 recognizes nucleated erythroid cells.46
CD271/NGF RCD271, also known as low affinity nerve growth factor receptor (NGF R) or p75NTR, is highly expressed by a fraction of clonogenic human bone marrow MSCs and is currently one of the most common and selective markers for enriching MSCs from bone marrow.47,48 Since CD271/NGF R is expressed at low levels by hematopoietic cells, separation of CD271/NGF R+bright versus CD271/NGFdim cells is typically used to distinguish MSCs from hematopoietic cells. Figure 3 demonstrates CD271/NGF R staining in human MSCs.
CD200CD200 was discovered as part of a screen to identify surface antigens expressed by MSCs but not hematopoietic cells.49 CD200+ cells are clonogenic and differentiate into adipocytes, chondrocytes, and osteocytes. However, a subsequent study showed that the expression of CD200 varied widely among twenty different human MSC lines, suggesting that the use of CD200 as a marker of MSCs may depend on the cell line or tissue source.50
Ganglioside GD2Ganglioside GD2 is a glycosphingolipid that has been reported as a marker that is expressed by MSCs but not hematopoietic cells. GD2 was expressed by 95% of CD45–CD105+CD73+ cells isolated from bone marrow. However, further studies are warranted to examine if GD2 expression is donor-dependent as Sivasubramaniyan et al. were unable to detect GD2 in primary bone marrow cells.45
Frizzled-9Frizzled-9, also known as CD348, was identified as part of a screen to detect MSC-specific markers. In bone marrow-derived MSCs, Frizzled-9 is expressed in CD271bright cells,51 whereas in placenta-derived MSCs, Frizzled-9 is expressed by highly clonogenic CD271/NGFdim cells.52 The difference in cell expression is likely due to the fact that MSC expression of CD271/NGF R appears to be dependent on the tissue source used for MSC isolation.
Figure 3. CD271/NGF R in Human Mesenchymal Stem Cells. NGF R/TNFRSF16 was detected in immersion fixed human mesenchymal stem cells using Mouse Anti-Human/Canine NGF R/TNFRSF16 Monoclonal Antibody (Catalog # MAB367) at 10 µg/mL for 3 hours at room temperature. Cells were stained using the NorthernLights™ 557-conjugated Anti-Goat IgG Secondary Antibody (red; Catalog # NL001) and counterstained with DAPI (blue). Specific staining was localized to cell surfaces.
Figure 4. Detection of Alkaline Phosphatase in Rat Stem Cells by Flow Cytometry. Rat Mesenchymal Stem Cells were stained with Goat Anti-Mouse Alkaline Phosphatase/ALPL Antigen Affinity-purified Polyclonal Antibody (Catalog # AF2910, filled histogram) or isotype control antibody (Catalog # AB-108-C, open histogram), followed by Phycoerythrin-conjugated Anti-Goat IgG Secondary Antibody (Catalog # F0107).
10
40
80
Rela
tive
Cell
Num
ber
1000
30
20
70
60
50
101 102 103 104
Alkaline Phosphatase/ALPL
Learn more | rndsystems.com/msc

Tissue Non-specific Alkaline Phosphatase (TNAP)TNAP, also referred to as Alkaline Phosphatase/ALPL, is identical to the Mesenchymal Stem Cell Antigen-1 (MSCA-1) and has been used as a marker for the prospective isolation of endometrium-derived MSCs. However, TNAP+ cells show limited multipotency and expression levels do not correlate with colony forming activity.53,54 Therefore, TNAP may be of limited use for MSC verification. Figure 4 shows expression of Alkaline Phosphatase in rat MSCs using flow cytometry.
USE OF MARKERS FOR MESENCHYMAL STEM CELL ISOLATIONThe use of markers to verify MSC identity serves as an important quality control step that can save significant time and reduce experimental variability. In an attempt to further decrease experimental variability, several labs have tried to increase the purity of isolated MSCs through positive and negative selection. Antibodies against CD45 or other hematopoietic markers can be used to negatively select MSCs, while antibodies against markers such as CD271/NGF R can be used to positively select MSCs. The success of positive and negative selection varies depending on the specificity of the marker, the starting cell population, and the isolation strategy. Although negative and positive selection of MSCs has gained acceptance in the field, there are some concerns over using such an isolation strategy.12 First, positive selection can lead to cross-linking of surface antigens, which may activate signaling pathways and alter cell phenotype. Second, any type of selection increases the risk of isolating a distinct cell population, which may preferentially differentiate towards a specific lineage. Despite these limitations, cells isolated by positive selection have been reported to have increased clonogenicity compared to cells isolated based on plastic adhesion.16,19,48,55
Negative Selection of MSCsNegative selection of cells is often preferred over positive selection since the isolated cells remain untouched by antibodies, beads, and magnets. In contrast, cells isolated by positive selection are labeled with antibodies and/or beads which may interfere with downstream applications and introduce experimental variability, either by preventing antibody binding or by stimulating signaling pathways. MSCs can be negatively selected by using a cocktail of antibodies to proteins expressed by hematopoietic cells (Figure 5). For human bone marrow stromal cells, common targets of negative selection include CD2, CD3, CD11b/Integrin alpha M, CD14, CD15/Lewis X, CD16/Fc gamma RIII, CD19, CD38, CD56/NCAM-1, CD66b/CEACAM-8, CD123/IL-3 R alpha, and CD235a/Glycophorin while CD45 and TER-119 are common targets for negative selection of murine MSCs. To improve the purity of negatively selected human MSCs, Modder et al. followed negative selection of MSCs (using antibodies to CD2, CD3, CD11b/Integrin alpha M, CD14, CD15/Lewis X, CD16/ Fc gamma RIII, CD19, CD56/NCAM-1, CD123/IL-3 R alpha, and CD235a/Glycophorin A) with fluorescence activated cell sorting (FACS) to isolate lin–/CD34–/CD31/PECAM-1- cells.56 This second selection step removed endothelial cells and precursors, yielding a cell population that was 97% negative for the hematopoietic marker CD45.56
Figure 5. Enrichment of MSCs from C57/BL6 Mouse Compact Bone Using the MagCellect™ Mesenchymal Stem Cell Isolation Kit (Catalog # MAGM212B). Cells before (A) and after (B) mesenchymal stem cell enrichment were stained with Anti-Mouse PDGF Rα Alexa Fluor® 488-conjugated Antibody (R&D Systems, Catalog # FAB1062G) and an Anti-Mouse CD45-APC Monoclonal Antibody (R&D Systems, Catalog # FAB114A). (C) Morphology of isolated and unstained MSCs demonstrating a typical mesenchymal stem/stromal cell shape.
PDG
F Rα
100
101
100
102
103
104
101 102 103 104
CD45
PDG
F Rα
100
101
100
102
103
104
101 102 103 104
CD45
Before MSC Enrichment After MSC Enrichment
Learn more | rndsystems.com/msc
A. B. C.

Positive Selection of MSCs
CD271/NGF RMSCs express high levels of CD271/NGF R while hematopoietic progenitor cells of the erythroid lineage express low levels of CD271/NGF R (CD271/NGFdim). Isolation of MSCs by CD271/NGF R positive selection is currently the gold standard to which new putative markers are compared. MSCs isolated by CD271/NGF R positive selection show lower hematopoietic cell contamination and higher clonogenicity compared to MSCs isolated by plastic adherence.16,48 Because CD271/NGF R is expressed by other cells in the bone marrow, selection by CD271/NGF R alone does not yield 100% pure MSCs.
CD105/EndoglinMSCs isolated by CD105/Endoglin positive selection demonstrate tri-lineage differentiation and higher colony-forming units-fibroblasts (CFU-F) compared to unsorted, plastic adherent cells.55 CD105/Endoglin is expressed by MSCs as well as other cells within the bone marrow. Therefore, selection based on CD105/Endoglin does not yield 100% pure MSCs. For example, flow cytometric analysis of freshly isolated human bone marrow cells enriched by CD105/Endoglin positive selection demonstrated a purity of 79%.55
STRO-1The STRO-1 antibody recognizes MSCs as well as nucleated erythroid cells. Therefore, the isolated cells do not approach 100% purity. To improve purity, FACS can be used to sort cells by expression of STRO-1 and Glycophorin A, a marker of erythroid cells. Isolation of STRO-1+/Glycophorin A– cells has been shown to yield a population with 100-fold enrichment of CFU-F compared to unfractionated bone marrow cells.19
Ganglioside GD2Ganglisoside GD2 is negatively expressed by hematopoietic cells and positively expressed by MSCs, making it a promising marker for MSC isolation. Sorting of bone marrow cells based on GD2 expression yielded cells with the ability to differentiate into adipocytes, chondrocytes, and osteocytes.57 However, Sivasubramaniyan et al. were only able to detect GD2 in cultured MSCs and did not detect expression in primary MSCs.45
SUSD2Sushi domain containing 2 (SUSD2) is a type 1 transmembrane protein that was recently identified as a marker that can be used for prospective isolation of MSCs.45 Antibodies directed against SUSD2 recognize CD271/NGF Rbright, but not CD271/NGFdim cells, suggesting that SUDS2 can differentiate between MSCs and hematopoietic cells. Sivasubramaniyan et al. showed that SUSD2+ but not SUSD2– cells, gave rise to CFU-F and were able to differentiate into adipocytes, chondrocytes, and osteoblasts.45 If these data are corroborated by other laboratories, GD2 may be revealed as one of the most specific MSC markers.
Positive Selection Using a Combination of Mesenchymal Stem Cell MarkersThe purity of MSCs can be increased by using more than one MSC marker for positive selection. As an example, a recent publication demonstrated that selection of mesenchymal stem cells using CD271/NGF R, CD90/Thy1, and CD106/VCAM-1 resulted in a highly clonogenic population of cells.58 Specifically, the addition of CD106/VCAM-1 as a marker for positive selection led to isolation of cells with five times greater clonogenic potential compared to the cells isolated with CD271/NGF R and CD90/Thy1 alone.
Learn more | rndsystems.com/msc

MARKERS AND ASSAYS TO EVALUATE MSC AGING, SENESCENCE AND POTENCYMSCs have a finite replicative capacity in culture. From early passage numbers, MSCs begin to demonstrate senescence-associated changes such as reduced proliferative capacity, changes in morphology, shortening of telomeres, and an accumulation of the cyclin-dependent kinase inhibitor p16INK4a/CDKN2A.59 Within approximately 5–12 passages, MSCs demonstrate decreased clonogenicity and differentiation potential.59,60,61,62 In parallel, several publications indicate that the expression of specific MSC markers, including CD106/VCAM-1,63,64,65 CD146/MCAM,63 and STRO-1,19 decreases with increasing passage number.
The decrease in differentiation potential of MSCs in culture emphasizes the need to verify MSC identity and multipotency both after isolation and after the cells have been passaged in culture. The parallel reduction in differentiation potential and MSC marker expression has led to the suggestion that CD106/VCAM-1, CD146/MCAM, and STRO-1 may be used as indicators of MSC age or potency.66 Additional research is needed to determine whether the decrease in MSC marker expression correlates with a reduction in differentiation potential and whether such a correlation is dependent on species or tissue source. If the reduction of specific MSC markers can be correlated with reduced differentiation potential or increasing age, then the markers could be used to simultaneously verify the identity and quality of MSCs. While more research is needed to examine the potential correlation between marker expression and MSC multipotency, several non-MSC markers and assays are commonly used to assess cell quality and replicative capacity.
USE OF NON-MSC MARKERS AND ASSAYS TO EVALUATE CELL QUALITYMSC aging or senescence can be assessed by examining the senescence-associated beta-galactosidase (SA-beta-Gal) activity, p16INK4a/CDKN2A levels, and cell clonogenicity by CFU-F.68,69 While telomere shortening is a hallmark of cell senescence, the length of telomeres varies from donor to donor, making telomere length a potentially unreliable measurement of MSC senescence.70
SA-beta-GalHistochemical detection of senescence-associated lysosomal beta-galactosidase (SA-beta-Gal) expression by senescent cells is the one of the most common assays used to assess senescence of MSCs and several other cell types.59,71 Although the assay was first published in 1995, it continues to be a standard assay for evaluating senescence. Cells are typically fixed and incubated with a SA-beta-Gal staining solution containing X-gal which generates a blue precipitate when cleaved by beta-galactosidase.
Alpha-L-FucosidaseRecently Hildebrand and colleagues identified alpha-L-Fucosidase (alpha-Fuc) as a novel marker of senescence that is upregulated in response to replicative, DNA damage-, and oncogene-induced senescence.72 Furthermore, as a marker of senescence, alpha-Fuc was equivalent or more reliable than SA-beta-Gal at the transcriptional and enzymatic level. If additional publications support this claim, then alpha-Fuc activity may become the preferred method for assessing senescence.
p16INK4a/CDKN2Ap16INK4a/CDKN2A is a cylcin-dependent kinase inhibitor that is highly expressed in senescent cells.73 Under physiological, non-senescent conditions, p16INK4a/CDKN2A is repressed by BMI1, a polycomb protein that maintains MSC self-renewal.74 P16INK4a expression in senescent MSCs positively correlates with activity of senescence associated beta-Galactosidase (SA-beta-Gal) and negatively correlates with expression of the proliferation marker MKI67/Ki-67.
Colony-Forming Unit-Fibroblast (CFU-F) AssayThe colony-forming unit-fibroblast assay can be used to estimate the proliferative and clonogenic potential of MSCs in culture. Bone marrow stromal cells are plated on plastic culture dishes at a low density and cultured under conditions that allow individual CFU-F to adhere and proliferate. By plating the cells at a low density, single colony clusters arise from the clonal expansion of a single CFU-F. The number of colonies present indicates the number of CFU-F present in the original sample. However, the colonies are only clonal when plated at a low density since at higher densities adherent cells with limited growth potential can be observed.75
Markers Reported to be Independent of Passage Number/Time in Culture
CD73/5′-Nucleotidase, CD90/Thy1, CD105/Endoglin21
Markers that are Reported to Decrease with Increasing Passage Number/Time in Culture
CD106/VCAM-1, STRO-12263,64,65
Marker Reported to be Associated with Differentiation Potential
CD106/VCAM-167
Table 2: The Effect of Time in Culture on MSC Marker Expression.
Learn more | rndsystems.com/msc
MSC MARKER VARIATION BY SPECIESMSCs isolated from animals such as mice, pigs, sheep, goats, horses, and dogs are used in preclinical trials and in the veterinarian clinic for the treatment of conditions such as osteoarthritis in dogs and musculoskeletal conditions in horses.76,77 Despite the widespread use of MSCs from non-humans, there are no established minimal criteria for the identification of MSCs in non-humans. While all MSCs display plastic adherence and tri-lineage differentiation, not all express the same panel of surface antigens that has been described for human MSCs. Most non-human MSCs express CD29/Integrin alpha 9 beta 1 and CD44. However, the expression of CD73/5′-Nucleotidase, CD90/Thy1, and CD105/Endoglin varies depending on species and strain.
For some animals, the limited availability of species-specific antibodies has made it difficult to establish positive and negative expression of MSC markers. In the absence of species-specific antibodies, anti-human antibodies are often used to examine the marker expression profile of non-human MSCs. However, if the antibodies are not examined for cross-reactivity, a negative finding may indicate that the marker is not expressed or that the antibody does not recognize the antigen. Results by Catherine De Shawer and colleagues emphasize the importance of verifying cross-reactivity.78 Their studies demonstrated that many of the anti-human and anti-mouse antibodies used to assess the phenotype and identity of equine MSCs did not-cross react with the equine antigens. For example, none of the anti-human CD34 antibodies recognized equine CD34, yet equine MSCs are reported to be negative for CD34 expression.78 These studies emphasize the need to include proper controls when using anti-human antibodies to evaluate the expression of non-human MSC markers. Table 3 includes results from individual publications that examined surface antigen expression of non-human MSCs.
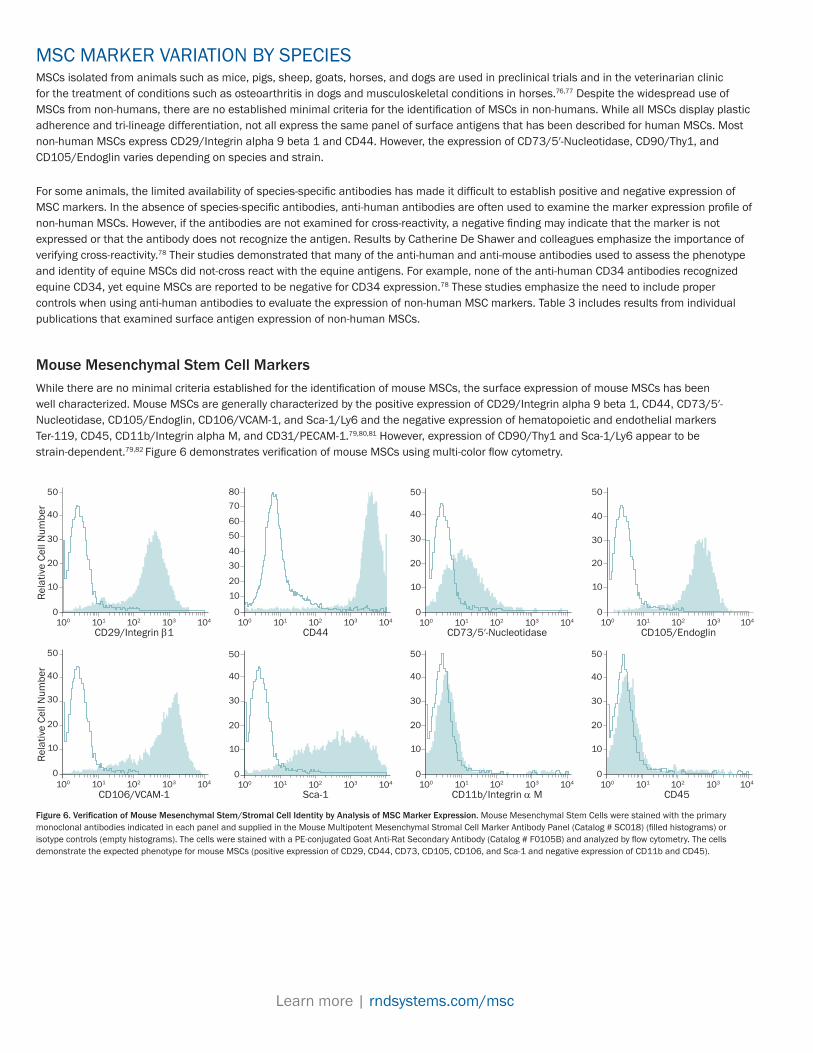
Mouse Mesenchymal Stem Cell MarkersWhile there are no minimal criteria established for the identification of mouse MSCs, the surface expression of mouse MSCs has been well characterized. Mouse MSCs are generally characterized by the positive expression of CD29/Integrin alpha 9 beta 1, CD44, CD73/5′-Nucleotidase, CD105/Endoglin, CD106/VCAM-1, and Sca-1/Ly6 and the negative expression of hematopoietic and endothelial markers Ter-119, CD45, CD11b/Integrin alpha M, and CD31/PECAM-1.79,80,81 However, expression of CD90/Thy1 and Sca-1/Ly6 appear to be strain-dependent.79,82 Figure 6 demonstrates verification of mouse MSCs using multi-color flow cytometry.
10
30
50
Rela
tive
Cell
Num
ber
1000
20
40
101 102 103 104
CD29/Integrin β1
10
30
80
1000
20
5060
70
40
101 102 103 104
CD44
10
30
50
1000
20
40
101 102 103 104
CD73/5′-Nucleotidase
10
30
50
1000
20
40
101 102 103 104
CD105/Endoglin
10
30
50
Rela
tive
Cell
Num
ber
1000
20
40
101 102 103 104
CD106/VCAM-1
10
30
50
1000
20
40
101 102 103 104
Sca-1
10
30
50
1000
20
40
101 102 103 104
CD11b/Integrin α M
10
30
50
1000
20
40
101 102 103 104
CD45
Figure 6. Verification of Mouse Mesenchymal Stem/Stromal Cell Identity by Analysis of MSC Marker Expression. Mouse Mesenchymal Stem Cells were stained with the primary monoclonal antibodies indicated in each panel and supplied in the Mouse Multipotent Mesenchymal Stromal Cell Marker Antibody Panel (Catalog # SC018) (filled histograms) or isotype controls (empty histograms). The cells were stained with a PE-conjugated Goat Anti-Rat Secondary Antibody (Catalog # F0105B) and analyzed by flow cytometry. The cells demonstrate the expected phenotype for mouse MSCs (positive expression of CD29, CD44, CD73, CD105, CD106, and Sca-1 and negative expression of CD11b and CD45).
Learn more | rndsystems.com/msc
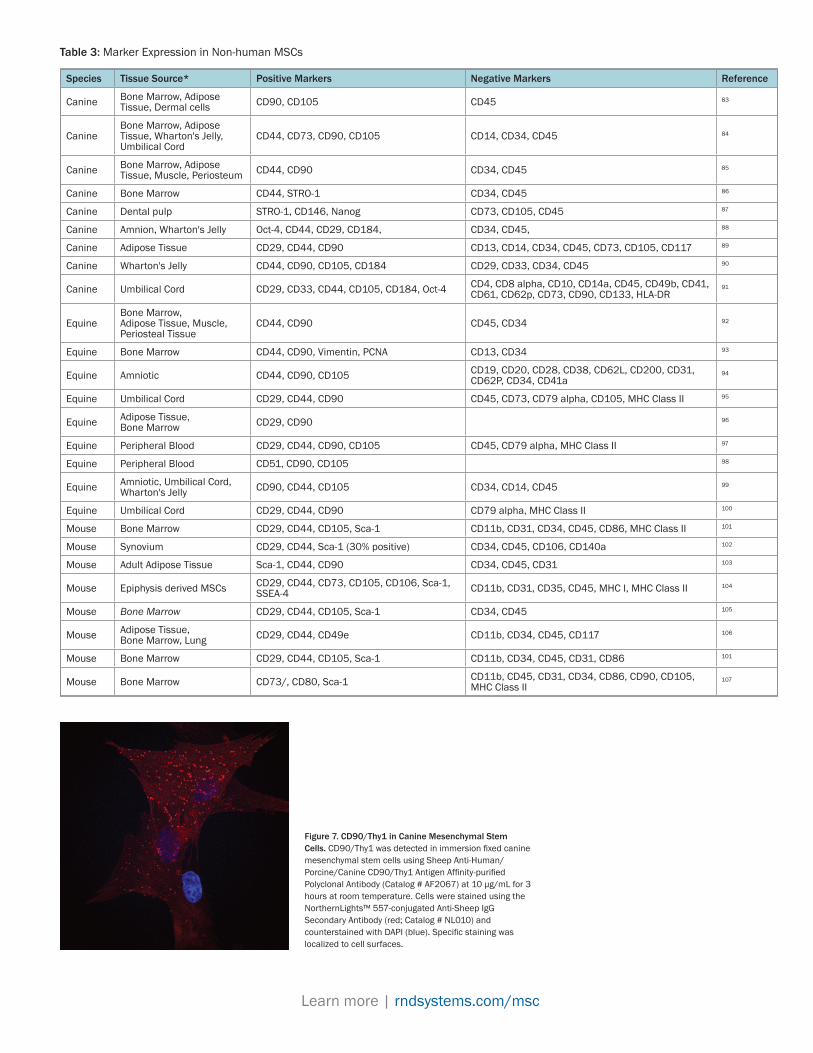
Species Tissue Source* Positive Markers Negative Markers Reference
Canine Bone Marrow, Adipose Tissue, Dermal cells CD90, CD105 CD45 83
Canine Bone Marrow, Adipose Tissue, Wharton's Jelly, Umbilical Cord
CD44, CD73, CD90, CD105 CD14, CD34, CD45 84
Canine Bone Marrow, Adipose Tissue, Muscle, Periosteum CD44, CD90 CD34, CD45 85
Canine Bone Marrow CD44, STRO-1 CD34, CD45 86
Canine Dental pulp STRO-1, CD146, Nanog CD73, CD105, CD45 87
Canine Amnion, Wharton's Jelly Oct-4, CD44, CD29, CD184, CD34, CD45, 88
Canine Adipose Tissue CD29, CD44, CD90 CD13, CD14, CD34, CD45, CD73, CD105, CD117 89
Canine Wharton's Jelly CD44, CD90, CD105, CD184 CD29, CD33, CD34, CD45 90
Canine Umbilical Cord CD29, CD33, CD44, CD105, CD184, Oct-4 CD4, CD8 alpha, CD10, CD14a, CD45, CD49b, CD41, CD61, CD62p, CD73, CD90, CD133, HLA-DR
91
Equine Bone Marrow, Adipose Tissue, Muscle, Periosteal Tissue
CD44, CD90 CD45, CD34 92
Equine Bone Marrow CD44, CD90, Vimentin, PCNA CD13, CD34 93
Equine Amniotic CD44, CD90, CD105 CD19, CD20, CD28, CD38, CD62L, CD200, CD31, CD62P, CD34, CD41a
94
Equine Umbilical Cord CD29, CD44, CD90 CD45, CD73, CD79 alpha, CD105, MHC Class II 95
Equine Adipose Tissue, Bone Marrow CD29, CD90 96
Equine Peripheral Blood CD29, CD44, CD90, CD105 CD45, CD79 alpha, MHC Class II 97
Equine Peripheral Blood CD51, CD90, CD105 98
Equine Amniotic, Umbilical Cord, Wharton's Jelly CD90, CD44, CD105 CD34, CD14, CD45 99
Equine Umbilical Cord CD29, CD44, CD90 CD79 alpha, MHC Class II 100
Mouse Bone Marrow CD29, CD44, CD105, Sca-1 CD11b, CD31, CD34, CD45, CD86, MHC Class II 101
Mouse Synovium CD29, CD44, Sca-1 (30% positive) CD34, CD45, CD106, CD140a 102
Mouse Adult Adipose Tissue Sca-1, CD44, CD90 CD34, CD45, CD31 103
Mouse Epiphysis derived MSCs CD29, CD44, CD73, CD105, CD106, Sca-1, SSEA-4 CD11b, CD31, CD35, CD45, MHC I, MHC Class II 104
Mouse Bone Marrow CD29, CD44, CD105, Sca-1 CD34, CD45 105
Mouse Adipose Tissue, Bone Marrow, Lung CD29, CD44, CD49e CD11b, CD34, CD45, CD117 106
Mouse Bone Marrow CD29, CD44, CD105, Sca-1 CD11b, CD34, CD45, CD31, CD86 101
Mouse Bone Marrow CD73/, CD80, Sca-1 CD11b, CD45, CD31, CD34, CD86, CD90, CD105, MHC Class II
107
Table 3: Marker Expression in Non-human MSCs
Figure 7. CD90/Thy1 in Canine Mesenchymal Stem Cells. CD90/Thy1 was detected in immersion fixed canine mesenchymal stem cells using Sheep Anti-Human/Porcine/Canine CD90/Thy1 Antigen Affinity-purified Polyclonal Antibody (Catalog # AF2067) at 10 µg/mL for 3 hours at room temperature. Cells were stained using the NorthernLights™ 557-conjugated Anti-Sheep IgG Secondary Antibody (red; Catalog # NL010) and counterstained with DAPI (blue). Specific staining was localized to cell surfaces.
Learn more | rndsystems.com/msc

METHODS FOR MSC IDENTIFICATION, VERIFICATION, AND CHARACTERIZATIONMSCs are identified by phenotypic assays, which assess marker expression and functional assays, which assess the differen tiation potential of the cell population. Table 4 includes the most common methods for verifying MSC identity, as well as the advantages and disadvantages of each method. For analysis of marker expression, flow cytometry and immunocytochemistry are efficient methods that reveal the marker profile of individual cells, whereas protein arrays and immunoblotting demonstrate the average marker expression by a population of cells.
Table 4: Methods for MSC Verification and Characterization
Method Type of Verification Advantages Disadvantages
Flow Cytometry Phenotypic
• Reveals the phenotype of individual cells• Examines several markers simultaneously• Can be used to isolate specific populations
(when equipped with a cell sorter)• High sensitivity• High speed analysis• Can use gating to remove dead cells from analysis
• Requires expensive equipment• Requires skilled operator• Potential cross-reactivity when using several
antibodies• Potential autofluorescence• Cell sorting by FACS can be time consuming
and expensive
Immunofluorescence/ Immunocytochemistry Phenotypic
• Reveals localization of marker proteins• Can assess multiple markers simultaneously• More efficient than Western blot analysis• Can use live or fixed cells
• Requires specialized equipment• Potential for cross-reactivity when using
multiple antibodies• Potential autofluorescence• Photobleaching• More time consuming than flow cytometry
Western Blot Phenotypic• High sensitivity• High specificity• Does not require specialized equipment
• Does not reveal heterogeneity of cell population• Low throughput
Protein Array Phenotypic
• High sensitivity• High specificity• Analyze several markers simultaneously• More efficient than individual Western blots• Cost effective compared to individual
Western blots• Does not require specialized equipment• Ideal for screening cell populations
• Does not reveal heterogeneity of cell population• Low throughput
Real-time RT-PCR Phenotypic
• Detects early changes in marker expression induced by differentiation
• Analyze several markers simultaneously• High sensitivity and specificity
• Yields the average marker expression of a population
• Does not reveal heterogeneity
Induction of Differentiation Functional
• Does not rely on MSC markers• Not dependent on tissue or species type
• Time consuming• Cannot recover MSCs
Learn more | rndsystems.com/msc

Functional Verification of Mesenchymal Stem CellsMesenchymal stem cells are defined by their ability to differentiate into adipocytes, chondrocytes, and osteocytes in culture. MSCs can be efficiently differentiated using a combination of media and supplements to induce adipogenesis, chondrogenesis, and osteogenesis. Additionally, small molecules can be used to enhance differentiation. Successful differentiation can be verified by staining the differentiated cells for expression of lineage markers and/or by assessing the functionality of the cells. Confirmation of tri-lineage differentiation provides excellent evidence for the verification of MSC identity. Furthermore, functional analysis is a well-accepted method of verifi cation that is not dependent on the tissue or species type. Figure 8 shows an example of functional verification of human MSCs.
Phenotypic Verification of MSCs by Multi-Color Flow CytometryFlow cytometry is an efficient method that can be used to examine the phenotype of cells at the single cell level. Multi-color flow cytometry allows users to examine multiple MSC markers simultaneously, which can reveal heterogeneity in a population that might not be obvious when examining multiple samples stained for individual markers. Specifically, multi-color flow cytometry can reveal the percentage of cells that express several MSC markers, whereas single-color flow cytometry can only reveal the percentage of cells that express each MSC marker individually. Additionally, many flow cytometers are equipped with a cell sorting function, which can be used to enrich for MSCs. Figure 9 demonstrates verification of human MSCs by multi-color flow cytometry.
Figure 9. Human Bone-Marrow Derived Cells Fulfill the ISCT's Definition of Human MSCs Based on Marker Expression. Human bone marrow-derived mesenchymal stem cells were stained using the antibodies and reagents provided in the Human Mesenchymal Stem Cell Marker Verification Multi-Color Flow Cytometry Kit (Catalog # FMC020). The data shows positive expression of MSC-associated surface antigens CD73, CD90, and CD105. In contrast, minimal expression of antigens recognized by the Negative Marker Cocktail was detected.
5′-N
ucle
otid
ase/
CD73
100
101
100
102
103
104
101 102 103 104
Negative Marker Cocktail-PE Negative Marker Cocktail-PE Negative Marker Cocktail-PE
CD90
/Thy
1
100
101
100
102
103
104
101 102 103 104
Endo
glin
/CD
105
100
101
100
102
103
104
101 102 103 104
Figure 8. Verification of Multipotency using the Human Mesenchymal Stem Cell Functional Identification Kit. Human mesenchymal stem cells were cultured in StemXVivo™ Mesenchymal Stem Cell Expansion Media (Catalog # CCM004) and differentiation was induced as indicated using the media supplements included in the Human Mesenchymal Stem Cell Functional Identification Kit (Catalog # SC006). The kit also contains a Goat Anti-Mouse FABP-4 Antigen Affinity-purified Polyclonal Antibody (adipocytes), a Goat Anti-Human Aggrecan Antigen Affinity-purified Polyclonal Antibody (chondrocytes), and a Mouse Anti-Human Osteocalcin Monoclonal Antibody (osteocytes) for the confirmation of differentiation status. The cells were stained using the NorthernLights™ 557-conjugated Donkey Anti-Goat (Catalog # NL001; red) or Anti-Mouse (Catalog # NL007; red) IgG Secondary Antibodies, The nuclei were counterstained with DAPI (blue).
© R&D Systems, Inc.
Human Mesenchymal Stem Cells
Chondrogenic Differentiation 14–21 days
Adipogenic Differentiation 7–21 days
Osteogenic Differentiation 14–21 days
Learn more | rndsystems.com/msc

SUMMARYMesenchymal stem cells have been isolated from a wide range of species and tissues using several techniques. MSCs are isolated as a heterogeneous population of cells that differ in growth kinetics and differentiation potentials and may be contaminated with terminally or partially differentiated cells. Prior to 2006, the heterogeneity of the cell population combined with the different techniques used to isolate, culture, and define MSCs led to experimental variability, inefficient differentiation, and contradictory data. The publication of the Minimal Criteria to Define Human MSCs in 2006 offered standards by which MSCs could be functionally and phenotypically defined.
Since the 2006 criteria, several studies suggest that the minimal criteria should be revised to account for newly identified markers as well as for species and tissue-specific variations in MSC phenotype. While the functional definition of MSCs is unlikely to change, the phenotypic definition appears to vary depending on the species and tissue source. Tissue-specific and species-specific MSC markers will likely emerge as source-specific MSCs are systematically characterized and as a wider variety of species-specific antibodies become available.
As MSCs become better characterized for their in vitro properties and their in vivo abilities, it is likely that the definition and nomenclature will continue to evolve. What markers do you find most useful for MSC isolation and characterization?
MEDIA FOR MSC EXPANSIONMedia formulations used for growing MSCs can vary by application as well as by research stage (ie, basic or translational). For many researchers, deciphering which variant of MSC media best suits their needs can be a daunting prospect. Specific formulations of MSC media now exist to address these various applications, including serum-containing, serum-free, xeno-free, and GMP-grade media.108,109,110 Table 5 discusses MSC media nomenclature and provides a quick reference for understanding the various media options and when to use them in your research.
Media Type Description
Serum-containing
Basic MSC expansion media is serum-containing, meaning it includes serum or plasma. Serum contains a variety of nutrients critical for cell survival and growth including growth factors, hormones, macromolecules, and adhesion factors. Serum-containing MSC media most commonly contains fetal bovine serum with fetal calf serum also being used by researchers. In general, MSCs tend to grow more robustly in serum-containing media but the lack of control over the unknown variables within serum can confound experimental results and be problematic when moving into clinical trials. While serum-containing MSC media is most commonly being used in research laboratories, it is also being used as an ancillary reagent in cell production for stem cell therapy.
Serum-freeSerum-free MSC expansion media does not contain serum or plasma. In comparison to serum-containing media, serum-free media provides more batch-to-batch consistency for the user and gives them more control over culture conditions. Serum-free MSC expansion media can also be used as an ancillary reagent for production of cells for cell therapy.
Xeno-free Xeno-free MSC expansion media does not contain any animal-derived components. Xeno-free medium may potentially contain human-derived components, such as human serum, holo-transferrin, or insulin. It is often used by researchers working to translate a therapy from research to clinic.
GMP-grade GMP-grade media is manufactured so that all products, materials, equipment, and processes used in production are traceable and controlled. Although universally recognized, there are regional differences in the documentation required by regulatory agencies regarding GMP-grade media.
Chemically-definedAll of the components in the media are known and it does not contain undefined components such as serum, tissue extracts, platelet lysate, etc. Chemically-defined media contains products and materials with known chemical structure such as, small molecules, salts, fatty acids, etc. It can also include recombinant proteins that are not made in animals.
Table 5: Description of Media used for MSC Expansion
Figure 10. Improved Expression of Mesenchymal Stem Cell (MSC) Markers in StemXVivo® Xeno-Free Human MSC Expansion Media. Human bone marrow-derived MSCs were expanded for 4 passages in StemXVivo® Xeno-Free Human MSC Expansion Media (Catalog # CCM021) or a competitor’s xeno-free MSC expansion media. The cells were harvested and stained for the expression of positive and negative MSC markers. Positive MSC markers were detected with APC-conjugated Mouse Anti-Human Endoglin/CD105 Monoclonal Antibody (Catalog # FAB10971A), PE-conjugated Mouse Anti-Human 5′-Nucleotidase/CD73 Monoclonal Antibody (Catalog # FAB5795P), and APC-conjugated Mouse Anti-Human CD90/Thy1 Monoclonal Antibody (Catalog # FAB2067A). Expression of the negative MSC marker, CD45 was detected with PE-conjugated Mouse Anti-Human CD45 Monoclonal Antibody (Catalog # FAB1430P). Quadrants were set based on isotype controls.
5′-N
ucle
otid
ase/
CD73
-PE
100100
101
102
103
104
101 102 103 104
Endoglin/CD105-APC
CD45
-PE
100100
101
102
103
104
101 102 103 104
CD90/Thy1-APC
5′-N
ucle
otid
ase/
CD73
-PE
100100
101
102
103
104
101 102 103 104
Endoglin/CD105-APC
CD45
-PE
100100
101
102
103
104
101 102 103 104
CD90/Thy1-APC
89%
69%
95%
71%
StemXVivo® Xeno-free Human MSC Media Competitor Xeno-free MSC Media
Learn more | rndsystems.com/msc

1. Dominici, M. et al. (2006) Cytotherapy 8:315.
2. Bianco, P. et al. (2013) Nat. Med. 19:35.
3. Bianco, P. & P. Gehron Robey (2000) J. Clin. Invest. 105:1663.
4. Méndez-Ferrer, S. et al. (2010) Nature 466:829.
5. Sacchetti, B. et al. (2007) Cell 131:324.
6. Caplan, A.I. (2010) Tissue Engin. Part A 16:2415.
7. Friedenstein, A.J. et al. (1970) Cell Tissue Kinet. 3:393.
8. Friedenstein, A.J. et al. (1968) Transplantation 6:230.
9. Caplan, A.I. (1991) J. Orthop. Res. 9:641.
10. Dennis, J.E. et al. (1999) J. Bone Miner. Res. 14:700.
11. Jiang, Y. et al. (2002) Nature 418:41.
12. Mosna, F. et al. (2010) Stem Cells Dev. 19:1449.
13. Lin, C.S. et al. (2012) Cytotherapy 14:1159.
14. Zimmerlin, L. et al. (2013) Cytotherapy 83:134.
15. Kaiser, S. et al. (2007) Cytotherapy 9:439.
16. Quirici, N. et al. (2010) Stem Cells Dev. 19:915.
17. Yoshimura, K. et al. (2006) J. Cell. Physiol. 208:64.
18. Pachón-Peña, G. et al. (2011) J. Cell. Physiol. 226:843.
19. Simmons, P.J. & B. Torok-Storb (1991) Blood 78:55.
20. Colter, D.C. et al. (2000) Proc. Natl. Acad. Sci. USA. 97:3213.
21. Pittenger, M.F. et al. (1999) Science 284:143.
22. Krause, D.S. et al. (1996) Blood 87:1.
23. Numasawa, Y. et al. (2011) Stem Cells 29:1405.
24. Vaculik, C. et al. (2012) J. Invest. Dermatol. 132:563.
25. Hermiston, M.L. et al. (2003) Annu. Rev. Immunol. 21:107.
26. Harvanová, D. et al. (2011) Folia Biol (Praha) 57:119.
27. Pilz, G.A. et al. (2011) Cytometry A 79:635.
28. Mazzone, A. &G. Ricevuti (1995) Haematologica 80:161.
29. Wagner, C. et al. (2001) Eur. J. Immunol. 31:1173.
30. Nareika, A. et al. (2008) J. Endocrinol. 196:45.
31. Lubis, A.M.T. et al. (2011) Acta Med. Indones-Indones J. Intern. Med. 43:178.
32. Tabera, S. et al. (2008) Haematologica 93:1301.
33. Gómez-Piña, V. et al. (2012) J. Leukoc. Biol. 91:933.
34. Notley, C.A. et al. (2011) J. Immunol. 186:4967.
35. Thorsby, E. (2009) Tissue Antigens 74:101.
36. Tran, N.T. et al. (2012) Stem Cells Dev. 21:1165.
37. Albert, R. et al. (2012) PLoS One 7:e47187.
38. Resta, R. et al. (1998) Immunol. Rev. 161:95.
39. Yu, W. et al. (2011) PLoS One 6:e18704.
40. Sowa, N.A. et al. (2010) J. Neurosci. 30:2235.
41. Rege, T.A. &J.S. Hagood (2006) FASEB J. 20:1045.
42. De Bruyn, C. et al. (2011) Stem Cells Dev. 20:547.
43. Duff, S.E. et al. (2003) FASEB J. 17:984.
44. Riekstina, U. et al. (2008) Cytotechnology 58:153.
45. Sivasubramaniyan, K. et al. (2013) Stem Cells Dev 22:1944.
46. Lin, G. et al. (2011) Stem Cells Dev. 20:1747.
47. Jones, E.A. et al. (2002) Arthritis Rheum. 46:3349.
48. Quirici, N. et al. (2002) Exp. Hematol. 30:783.
49. Delorme, B. et al. (2008) Blood 111:2631.
50. Pietilä, M. et al. (2012) PLoS One 7:e31671.
51. Bühring, H.J. et al. (2007) Ann. N.Y. Acad. Sci. 1106:262.
52. Battula, V.L. et al. (2008) Differentiation 76:326.
53. Alexander, D. et al. (2010) Cell Physiol. Biochem. 26:1073.
54. Kim, Y.H. et al. (2012) Stem Cells Dev. 21:2958.
55. Aslan, H. et al. (2006) Stem Cells 27:1728.
56. Mödder, U.I. et al. (2012) Bone 50:804.
57. Martinez, C. et al. (2007) Blood 109:4245.
58. Mabuchi, Y. et al. (2013) Stem Cell Rep. 1:152.
59. Wagner, W. et al. (2008) PLoS One 3:e2213.
60. Bonab, M.M. et al. (2006) BMC Cell Biol. 7:14.
61. Izadpanah, R. et al. (2008) Cancer Res. 68:4229.
62. Madeira, A. et al. (2012) PLoS One 7:e43523.
63. Abramov, N. et al. (2011) Stem Cells Dev. 20:53.
64. Jung, E.M. et al. (2011) Biochem. Biophys. Res. Commun. 404:463.
65. Ooi, Y.Y. et al. (2013) World J. Stem Cells 5:34.
66. Boxall, S.A. &E. Jones (2012) Stem Cells Int. 2012:975871.
67. Liu, F. et al. (2008) J. Bone Miner.Metab. 26:312.
68. Vidal, M.A. et al. (2012) Stem Cells Dev. 21:273.
69. Zhang, D.-Y. et al. (2011) PLoS One 6:e21397.
70. Mareschi, K. et al. (2006) J. Cell. Biochem. 97:744.
71. Dimri, G.P. et al. (1995) Proc. Natl. Acad. Sci. USA 92:9363.
72. Hildebrand, D.G. et al. (2013) Cell Cycle 12:1922.
73. Shibata, K.R. et al. (2007) Stem Cells 25:2371.
74. Zhang, H.W. et al. (2010) J. Bone Miner. Res. 25:640.
75. Bianco, P. et al. (2008) Cell Stem Cell 2:313.
76. Cyranoski, D. (2013) Nature 496:148.
77. De Schauwer, C. et al. (2013) Vet. Q. in press.
78. De Schauwer, C. et al. (2012) Cytometry A 81:312.
79. Baddoo, M. (2003) J. Cell. Biochem. 89:1235.
80. Peister, A. et al. (2004) Blood 103:1662.
81. Tropel, P. (2004) Exp. Cell. Res. 295:395.
82. Phinney, D.G. et al. (1999) J. Cell. Biochem. 72:570.
83. Ock, S.A. et al. (2012) Cell Transplant. in press.
84. Kang, B.-J. et al. (2012) J. Vet. Sci. 13:299.
85. Kisiel, A.H. et al. (2012) Am. J. Vet. Res. 73:1305.
86. Hodgkiss-Geere, H.M. et al. (2012) Vet J. 194:361.
87. Dissanayaka, W.L. et al. (2011) J. Endod. 37:1074.
88. Filioli Uranio, M. et al. (2011) Mol. Reprod. Dev. 78:361.
89. N.M, V. et al. (2010) Cell Transplant. 19:279.
90. Lee, K.S. et al. (2013) Res. Vet. Sci. 94:144.
91. Seo, M.-S. et al. (2009) J. Vet. Sci. 10:181.
92. Radtke, C.L. et al. (2013) Am. J. Vet. Res. 74:790.
93. Maia, L. et al. (2013) Microsc. Res. Tech. 76:618.
94. Seo, M.-S. et al. (2013) J. Vet. Sci. 14:151.
95. De Schauwer, C. et al. (2013) Equine Vet. J. 45:518.
96. Ranera, B. et al. (2012) BMC Vet. Res. 8:142.
97. Spaas, J.H. et al. (2013) Vet. J. 195:107.
98. Dhar, M. et al. (2012) Equine Vet. J. 44:600.
99. Iacono, E. et al. (2012) Reproduction 143:455.
100. De Schauwer, C. et al. (2011) Tissue Eng. Part C 17:1061.
101. Zhu, H. et al. (2010) Nat. Protoc. 5:550.
102. Futami, I. et al. (2012) PLoS One 7:e45517.
103. Sági, B. et al. (2012) Stem Cells Dev. 21:814.
104. Cheng, C.-C. et al. (2012) PLoS One 7:e36085.
105. Shen, J. et al. (2011) Sci. Rep. 1:67.
106. Nora, C.C. et al. (2012) Stem Cells Dev. 21:1761.
107. Schurgers, E. et al. (2010) Arthritis Res. Ther. 12:R31.
108. Solomon, J. et al. (2016) Cryotherapy. 18:1.
109. Usta, S.N. et al. (2014) Ann. of Trans. Med. 2:97.
110. Medicino M. et al. (2014) Cell Stem Cell. 2:141. 12:R31.
REFERENCES
WP_MSC_13763
Global [email protected] bio-techne.com/find-us/distributors TEL +1 612 379 2956North America TEL 800 343 7475 Europe | Middle East | Africa TEL +44 (0)1235 529449China [email protected] TEL +86 (21) 52380373 bio-techne.com