main patterns and individual differences in baboon skull split-lines and theories of causes of...
TRANSCRIPT

Main Patterns and Individual Differences in Baboon Skull Split-lines and Theories of Causes of Split-line Orientation in Bone '
N. C. TAPPEN Department of Anthropology, University of Wisconsin, Milwaukee, Wisconsin 53201
ABSTRACT Split-line pattcrns are reported in skulls of five adult male baboons. While variations in pattern occur in all parts of the skull. these variations are rela- tively minor in the following regions: supraorbital, lateral orbital, medial orbital, nasal bones, zygomatico-alveolar crest, nasal opening, alveolar process of maxilla and mandible. Wide differences in pattern occur in these regions: infraorhital, zygomatic bone, body of maxilla, and frontal bone posterior to the supraorbital area. The major variability in split-line orientation indicates that oversimplified interpretations of the patterns in terms of ( 1) conformity to gross structure, or (2) direction of bone growth. are untenable. The variation3 do not contradict a functional interpretation in which mechanical forces and skull form interact to different degrees in different individuals. however. Skulls of a variety of primates are useful for functional analysis, because they have similar structural plans, but the differences are well outside the normal range OC variation for a single species.
A reccnt report (Tappen, '67) outlined major features of split-line patterns in single skulls from local populations of ba- boons ( P a p i o ) , mangabeys (Cercocebus), and guenons (Cercopithecus) and pointed out some differences among these genera which appeared to be related to peculiari- ties in the structure of the skulls of each species. Space limitations placed upon the original report did not allow presentation of details of the patterns and variations within species. Because individual differ- ences were most striking in the baboons, and because these differences give evidence relevant to general theories of the causes of split-line oricntations in bone, more ex- tensive descriptions of the split-line or- ganization in all baboon sltulls processed are presented here.
Benninghoff ('25 introduced the tech- nique of making repeated needle punctures into decalcified compact bone. In the great majority of areas these punctures produce cracks rather than round holes. Moreover, in many areas the great majority of the fissures are oriented in the same general direction, producing patterns of lines along the surface. These patterns are made ap- parent by making many punctures and in-
AM. 3. PHYS. ANTRROP, 33' 61-72
serting India ink into the splits. Cortical bone may thus be shown to have a "grain" analogous to that of wood. Benninghoff ('25) believed that the orientation patterns were a response to mechanical forces, such as tension or compression, acting upon the bone. Comparative studies (e.g. Tap- pen. '53, '54) tend to support this assump- tion. Microscopic studies (Tappen, '67 j show a close correspondence of split-line orientation with that of the axes of Haver- sian systems, but split-lines are also highly oriented in circumferential lamellar bone where no Haversian systems are present. In this case i t is probable that split-lines correspond to the direction of orientation of the bulk of the collagen fibers which make up 95% of the bone matrix.
Evans ('65), in a communication at- tacking the theory that split-line patterns are a function of mechanical forces acting upon bone and defending alternative expla- nations. cites a study by Dempster ('65) indicating that split-line orientation pat- terns in the human skull are closely related to its gross structural features. This essen- tially constitutes yet another hypothesis ~~
1 Supported by National Institutes of Health grant HD 02033.
61

62 N. C. TAPPEN
concerning the factors responsible for split- line orientation. According to Dempster, “cylindrical surfaces typically show a lengthwise grain transverse to the curva- ture; trenches show a crosswise grain, basin-like fossae often have a circular or spiral grain. The margins of foramina and thin laminae with unsupported borders characteristically show a grain parallel to the edge. Spines and sharp processes have elongate grain patterns.” The “grain” was revealed by split-line preparations. The im- plication is that the shape of the bony structures in the human skull determines the orientation of the split-line patterns, and presumably this would also be true for any bone in which the patterns may be demonstratfd. This explanation €or split- line orientation has not previously been of- fered, to my knowledge, and it deserves close examination. Information on baboon skull split-lines provides a partial com- parative test.
MATERIALS AND METHODS
Split-line preparations were made on skulls of five adult male baboons from the region of the confluence of the Kibwezi and Athi rivers, Kenya, according to methods described by Tappen (’54). The skulls are from the uniquely valuable collection cap- tured and autopsied in the field by McGill et al. (’60) as part of a study on primate atherosclerosis. Brains had been removed at autopsy, and the cranial vault had been discarded, but the skulls are otherwise com- plete, with fully erupted dentition. The skulls were not otherwise selected for any particular characteristics so far as I am aware; the entire collection was on loan to Dr. Clyde Snow. and the specimens used were returned at my request for any five adult male skulls.
The skulls were decalcified in dilute ni- tric acid after the teeth had been protected from demineralization by application of layers of melted dental impression wax. After washing, splits were produced by punctures from a sharpened teasing needle perpendicular to the plane of the bone sur- face. The needle may penetrate through the outer compact bone, which varies greatly in thickness in different regions. making cracks 1-3 mm in length. Where the direction of orientation of the split is
obvious the needle is left in position and used to lengthen the split by pulling in the direction of least resistance. This pro- cedure makes i t possible to reveal with pre- cision patterns which converge, diverge or curve. Attempts to pull the needle in other directions meet immediate heavy resist- ance. Occasionally resistance is met in the direction which previously offered least re- sistance. Where this does not signal the encounter with another system OP split- lines, i t probably is caused by the buildup of osseous lamellae around a secondary transverse iVolkmann’s) canal in the bone. In such areas separate punctures are made at 1-2 mm intervals until the orientation is re-established or disappears.
India ink is inserted into the splits as they are held open with the teasing needle, using a hypodermic needle or a fine-pointed pen, and excess ink is blotted and wiped away with a wet rag. Inked split-lines are prepared at 2-3 mm intervals to reveal the general patterns in any bony area. Splits placed closer together become difficult to distinguish, especially in bone with enough porosity to allow excess diffusion of the ink. Where lengthened split-lines in a par- ticular area are approximately parallel, punctures between them are nearly always parallel to them. This procedure is followed as a test of the validity of the pattern. The additional splits close after the needle is removed and can seldom be seen unless ink is inserted. The split-lines are thus sampling the general structural orientation of the compact bone at different localities.
Split-line preparations were made by Carolyn Pons and Donald Duvall under the supervision of the author. Major patterns were confirmed by making additional needle punctures in the area without mark- ing the resultant splits with ink. In some critical areas a t the junctures of major sys- terns of split-lines the preparations were completed bp the author.
OBSERVATIONS
Split-line preparations arc illustrated in figures 1-3. Areas showing closest similari- ties in all specimens are described first, with those regions which contain notable variations from specimen to specimen fol- lowing. The skulls are identified numeri- cally from the original tags. with the ex-

BABOON SKULL SPLIT-LINES 6 3
ception of one specimen whose tag had been lost, referred to subsequently as “NO Number.”
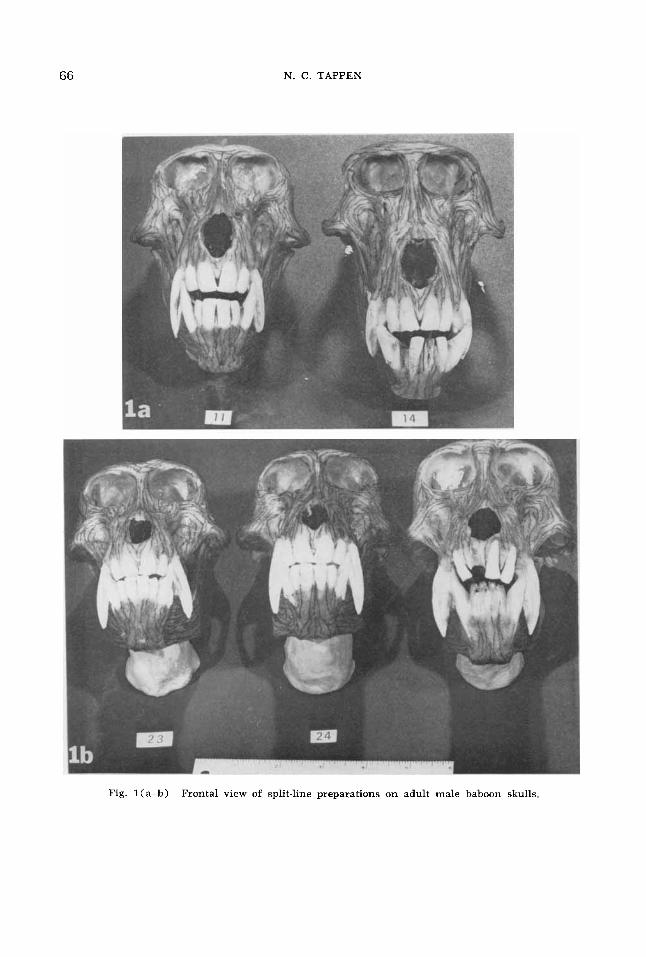
A . Areas with similar patterns 1. Supraorbital. The patterns are all
oriented parallel to the superior orbital margin. Above the region between the or- bits there is a tendency for the split-lines to dip toward the nasal bones, most strongly illustrated in number 24.
The split-lines con- tinue parallel to the orbital margins beyond the zygomatico-frontal suture. Variations below this point will be described in Sec- tion B-1 ( lnfraorbi tal) .
3 . Medial orbital. Split-lines nearest the orbital border tend to curve away from the lines paralleling the superior border of the orbit, continuing to conform closely to the outlines of the orbital margin as they course parallel to those in the superior por- tions of the nasal bones. Near the lower margins of the medial border of the orbits the lines continue mainly parallel to the margins of the nasal bones, except for “No Number.” Further mention of this area will be undertaken with the description of the infraorbital region, where major varia- tions occur, since there are patterns of transition and continuity between the two.
4. Nasal bones. The patterns course parallel to the axis of the bones from the naso-frontal suture to the border of the nasal opening
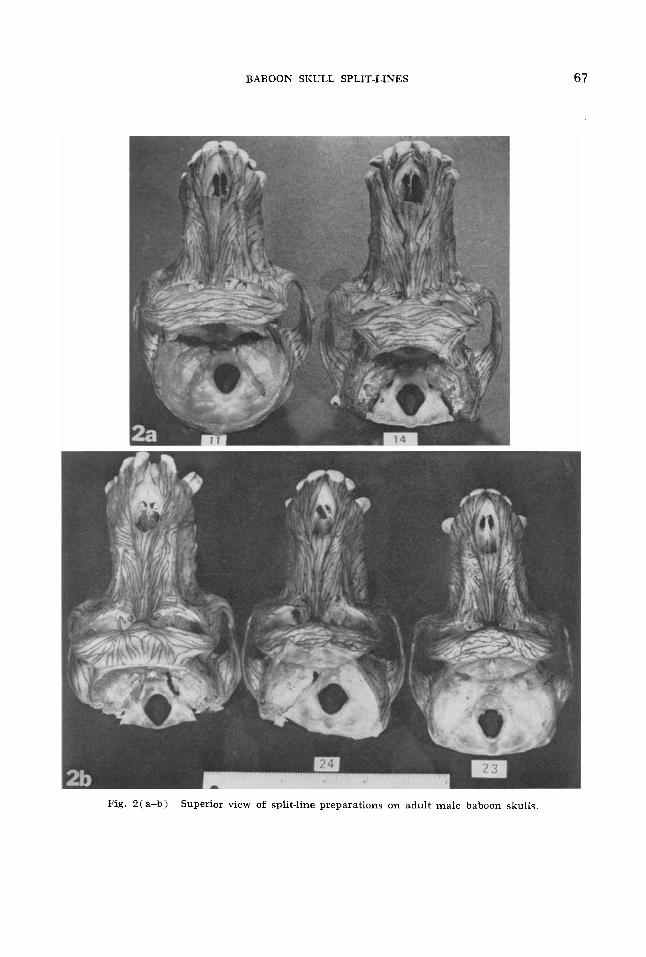
5. Zyyomatico-alveolar crest and maxil- E R ~ Y ridge. The zygomatico-alveolar crest is especially pronounced in the baboon skulls, continuous with the zygomatic arch and projecting to an area in the maxilla ex- ternal to the roots of the first and second molar teeth. Split-lines in all specimens show a strong tendency to run parallel to the lateral border of this crest on both the facial and temporal surfaces. Viewed an- teriorly, the lines closest to the border of the crest continue into the zygomatic arch, or, in a few cases, continue with the other ascending lines in the crest and, along with others lateral to them in the body of the zygomatic bone (see Section B-21, recurve above the fossa medial to the zygomatico- alveolar crest to become continuous with lines conforming to the axis of a ridge in the maxilla which becomes apparent an-
2. Lateral orbital.
terior to the orbit and continues to the re- gion of the apex of the root of the canine tooth. This structure, here termed the maxillary ridge, may be observed in all views, but is clearest in figure 2. The bone in the fossa surrounded by these recurving split-lines is extremely thin, and the split- line patterns do not show any consistent trend i n the different specimens, nor on opposite sides of the same specimen.
6. Area of nasal cavity opening. Split- lines in the nasal bones continue parallel to its axis all the way to the superior border of the nasal opening. Only in “No Number” is there a slight lateral deviation partially conforming to thc curvature of the border of the aperture. Split-lines in the premax- illa course approximately parallel to the lateral border, except in number 11, where the lines deviate laterally from above down- ward. As the lower margin of the nasal opening curves toward the midline, the split-lines tend not to conform to this curva- ture, but to continue to the margins of the alveoli of the incisor teeth. However. the split-lines closest to the border of the open- ing conform to its outlines until they reach the triangular area bounded by the inferior border of the opening and the central in- cisor teeth. where they turn and proceed toward the most inferior part of the alveo- lar area between the central incisors.
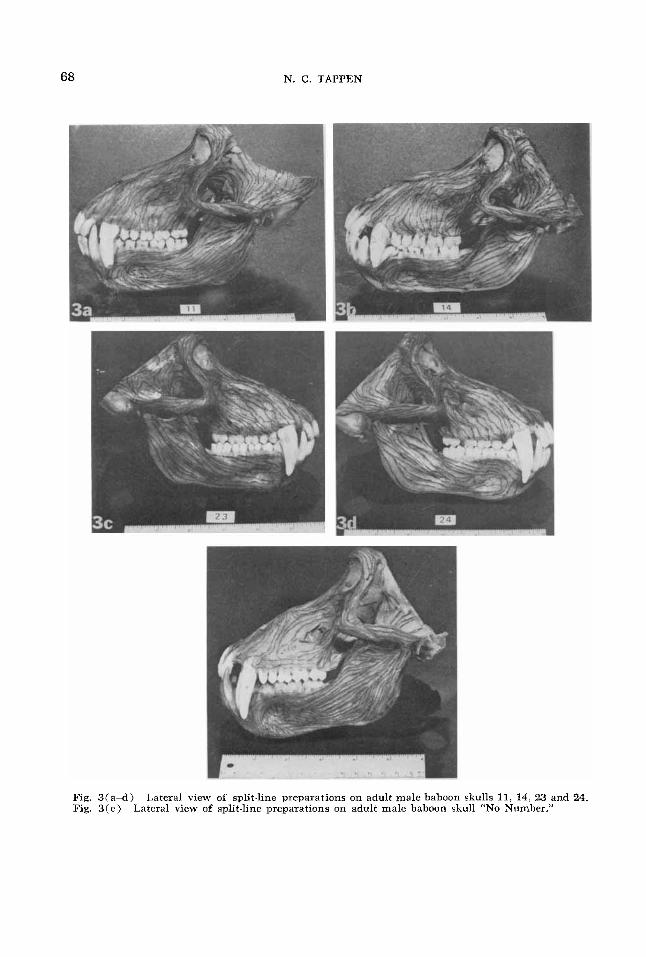
7. Alueolar process of maxilla. The general orientation of the split-lines in this area tends to conform to the axes of the roots of the teeth, as already mentioned for the incisors in the preceding paragraph. This is true in all cases for the bone as- sociated with the roots of the canines. More variation occurs in the areas of the pre- molars and molars, as illustrated in figure 3. The general tendency is for the split- lines to descend toward the alveolar mar- gins at various degrees of obliquity, as seen in numbers 11, 14. 23 and 24, with some tendency for the split-lines near alveolar borders to become approximatelv parallel to these margins in limited areas. This tendency is much exaggerated in “No Num- ber,” so that the split-lines are parallel to the alveolar margin as far as the apices of the molar and premolar roots. In none of the specimens are the patterns on the op- posite sides in the same individuals precise duplicates of the ones shown, although

64 N. C. TAPPEN
none show radical departures from those illustrated in figure 3 .
8. Temporal fossa. This area is incom- plete in all specimens because of the re- moval of brains al autopsy. The remaining portion is closely equivalent in all speci- mens and shows a substantial uniformity of pattern despite the prevalence of very thin cortical bone in the lateral walls of the orbits. Split-lines continuing from the zygomatic arch fan out in the fossa, the ones on and closest to the supramastoid crest and its continuation, the temporal line. running parallel to these structures, while the split-lines farther removed gradu- ally begin to ascend the temporal squama. The most anterior of these ascending lines recurve forward in the lateral orbital wall to become continuous with split-lines along the lateral orbital margins, zygomatic area and zygomatico-alveolar crest. The prepara- tions in “No Number” were not completed at the time of photographing figure 3, but subsequent processing showed no essential differences from other specimens.
9. Mandible. Viewed anteriorly, all mandibles have the same basic pattern of split-lines arranged parallel to the axes of the roots of the incisor and canine teeth. In lateral view there is also considerable uniformity. Split-lines in the ascending ramus are directed toward the condylar and coronoid processes, with a somewhat vari- able area near the mandibular notch show- ing patterns parallel to this border and connecting the two processes. Split-lines at the tip of the coronoid process recurve backward to parallel the two borders; in some individuals the split-lines paralleling the anterior and mandibular notch margins meet virtually at a point. The zygomatic arch obscures this pattern in figure 3. The split-lines near the posterior border of the ramus are approximately parallel to this border as they descend from the condylar area, curving at the gonial area to continue parallel to the inferior border of the body of the mandible. Split-lines descending from the condyle tend to be directed away from the angle more and more as they are further removed from it in distance, form- ing a much smaller arc as they turn ap- proximately parallel to the axis of the body. The most anterior of these split-lines be- come confluent with others in the ramus
which parallel the anterior border as they descend from the coronoid process. These lines course parallel to the oblique line. which becomes prominent immediately posterior to the third molar tooth, and con- tinue more or less parallel to the alveolar border to the sectorial lower first premolar. The alveolar margin here dips sharply in conforming to the peculiar shape of this tooth, and the split-lines also dip to parallel the margin. These lines are continuous with others which have turned upward after coursing parallel to the inferior mar- gin of the mandible, so that a loop of split- lines tends to form around the deep fossa which lies below the roots of the second premolar and first molar. This fossa is most apparent in the photographs of numbers 11, 14 and “No Number,” but is present in all specimens. Cortical bone tends to be thin within the fossa itself, and split-line orientation is variable.
Comment . While the split-line patterns described thus far are in regions which show relative uniformity, i t should be ob- served that some variability is found among the different individuals, and even on left and right sides in the same individ- ual. Where this variabilitv has seemed rather noteworthy it has been pointed out. The regions to be described next show vari- ations which are frequently quite striking and appear to have considerable impor. tance for theories of bone organization.
B. Areas with differing patterns 1. Infraorbital. The strong differences
in pattern along the inferior border of the orbit are clearly shown in figures 1 and 2. In the frontosphenoidal process of the zygomatic bone the split-lines nearest the orbital margin conform closely to its con- tours in their orientation, while the lines closer to the temporal border continue downward toward the zygomatic arch. Number 24 provides a notable exception, since the split-lines tend toward the hori- zontal throughout much of this region. The most lateral split-lines in this case are par- allel to the temporal border, and a thin area immediately adjacent to the orbital border courses parallel to this margin on the right side. However, the intermediate split-lines radiate more and more as the body of the zygomatic bone is approached, so that they

BABOON SKULL SPLIT-LINES 65
gradualIy assume the inferolateral orien- tation of the split-lines in this region seen in the other specimens. On the left side these split-lines proceed to the orbital mar- gin, so no lines are parallel to the orbital border on this side until the vicinity of the zygoniatico-maxillary juncture is ap- proached.
The differences are even more striking along the mcdial portion of the infraorbital region. In numbers 14, 24 and “No Num- ber” the split-lines in the immediate vicin- ity of the orbital border are approximately parallel to it, although this area is quite thin in number 24. In numbers 11 and 23, however, split-lines are at right angles to this pattern, entering the orbit as continn- ations of split-lines in the muzzle which are approximately parallel to the orienta- tion of the axis of the nasal bones. This system of lines is also present in the other specimens, but is replaced near the orbit by the split-lines parallel to the orbital mar- gin. In “No Xumber” a small ridge contin- ues from the muzzle, and the split-lines in it interrupt the continuity of much of the rather wide area of these split-lines paral- lel to the orbit, especially on the right side, but do not reach the orbital border.
The medial border of the lower orbital niargin had been cut away on the right side in number 24, apparently while the animal was being skinned. The split-line pattern in this small region is consequent- ly unknown.
2. Zyyomatic region and zygornatic arch. Below the lateral orbital region the split- lines near the temporal border of the zygomatic bone continue from the fronto- sphenoidal process approximately parallel to the border in all specimens. continuing to the base of the zygomatic arch. In this region the orientations of the split-lines vary markedly. In number 14 and “No Number,” and in number 23 on the left side, the descending split-lines turn at the temporal border to continue approximately parallel to the axis of the zygomatic arch. The split-lines along the masseteric border are also approximately parallel to the arch in these specimens, but are continuous with lines parallel to the border of the zy- gomatico-alveolar crest. In number 24 and the right side of number 23, the lines from above continue to the masseteric border at
the base of the zygomatic arch, cutting off the system of lines ascending along the border of the zygomatico-alveolar crest and only gradually turning in the zygomatic arch in conformity with its axis. In num- ber 11 the pattern is intermediate, tending toward that of number 24, but there is a thin zone at the extreme masseteric border in which the split-lines are continuous with those ascending near the margin of the zygoimatico-alveolar crest. In all speci- mens the split-lines of this region show a continuity with those of the maxillary crest, recurving above the fossa separating the two regions. except for number 11 on the right side. Here the pattern of lines ascending vertically to enter the orbit reaches its most lateral extent, apparently interfering with a tendency for the split- lines in the zygomatic and maxillary crest areas to meet. On the left side of the same specimen the area of recurvature is small but definitely present.
The split-lines in the zygomatic arch tend to conform in direction to its axis in the more posterior regions, especially in the zygomatic process of the temporal bone (fig. 3 ) . They also conform to the changes in contour of the arch as it narrows pos- teriorly, and the split-lines then continue into the supra-mastoid crest.
3. Body of the maxilla. Split-lines coursing irom the region below the orbits and €rum the frontal process of the maxilla tend to converge with those coursing par- allel to the axis of the maxillary ridge (de- scribed in Section A-5) in the area lateral to the naso-premaxillary suture above the nasal opening. It is here that the ridge be- comes most pronounced (it is much more exaggerated in other baboons, notably man- drills), but in numbers 11, 23 and “NO Kumber” the continuing split-lines, in- stead of conforming in direction to its axis, turn laterally as they continue toward the alveolar region. There is also a tendency for this to occur in numbers 14 and 24, but the laterally deviating split-lines do not cross all the way over the ridge.
4. Frontal bone posterior to supraorbital area. Much of this region had been cut away at autopsy, so generalizations must be tentative. In all specimens except “No Number,” the split-lines are mainly trans- verse, continuing the main pattern of the
66 N. C . TAPPEN
Fig. l(a-b) Frontal view of split-line preparations on adult male baboon skulls.
BABOON SKULL SPLIT-LINES
Fig. 2(a-b) Superior view of split-line preparations on adult male baboon skulls.
67
68 N. C. TAPPEN
Fig. 3(a-d) Fig. 3 ( e )
Lateral view of split-line preparations on adult male baboon skulls 11, 14. 23 and 24. Lateral view of split-line preparations on adult male baboon skull “No Number.”

BABOON SKULL SPLIT-LINES 69
supraorbital split-lines. Tn “No Number” the main trend is antero-posterior, with transitional zones from the supraorbital split-lines. In “No Number“ the main trend is antero-posterior, with transitional zones from the supraorbital orientation indicated in some areas.
DISCUSSION
Major variations in split-line patterns in the baboon skulls show clearly that the shape of skull structures cannot be a pri- mary determinant of orientation of split- lines. The striking individual differences in Orientation of the lines in the infra- urbital, zygomatic, maxillary ridge and frontal bone regions are all associated with structures which are otherwise directly comparable and do not exhibit very notice- able morphological variation. The same may be said for some of the less striking variations observed in other parts of the baboon sltulls.
Dr. Dempster unfortunately died before he was able to publish more than the ab- stract (Dempster, ’65), parts of which have been quoted in the introduction to this article. It is not clear from the ab- stract what he believed to be the causes of split-line orientation, although there is a definite implication that split-line pat- terns are a response to the shape of the structures in which they appear. However, he also stated that “bone is weaker in ten- sion than in compression and several times weaker across the grain than along the grain. In terms of these physical proper- ties, each of the above grain patterns rep- resents a mechanically strong arrangement of- bone.” There would thus be functional importance in the orientation patterns of bone even if’ the mechanical forces acting on the bone did not contribute to its split- line orientation, if Dempster’s mechanical analysis is correct,
Another possible explanation for split- line orientation proposed by Evans and Goff (’57) was that i t represents the direction of growth of bone. Some evidence against this hypothesis as a general explanation has been presented elsewhere (Tappen, ‘641, and the great differences in split-line orientation in areas such as the baboon infraorbital region makes this seem even more improbable. It seems very unlikely
that longitudinal bone growth produces the lower orbital border pattern in numbers 11 and 23 while transverse growth is respon- sible for the development of this region in number 14 and “No Number.” The relative uniformity of growth process in this part of the face in man has been determined by Bjork (’68).
Contrasting patterns of split-lines in the lower orbital border in different species (Tappen, ’53) show the lines conforming to the orientation of this margin in man and entering the orbit at right angles to it in a chimpanzee. The present study shows that the contrast can occur in the same age and sex group of a single species. The dif- ferences between the human and chimpan- zee patterns of split-lines in the orbital and zygomatic areas were indicated to be closely related to differences in the distance between the orbits and the area of origin of the masseter muscle. and possibly to dif- ferences in the relative amount of force ex- erted by the muscle. In the baboon skulls striking differences in pattern can be found where gross morphological differences are much smaller. This indicates that general- izations about the “grain” of cortical bone need to be based on split-line preparations from more than a single species. The value of the sliulls of other primates, similar in basic structures but beyond the range of normal human variability, has been empha- sized in previous publications on compara- tive split-line analysis (e g., Tappen, ’53, ’54, ’67 1.
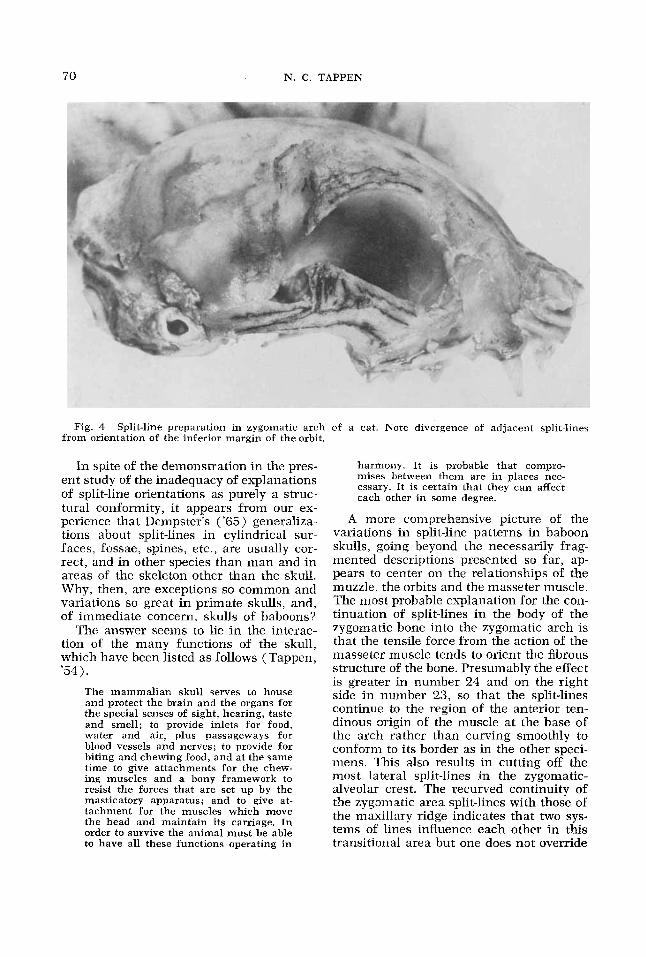
One additional illustration of the rela- tionship of bone structure to split-line ori- entation may be of value. In early work de- veloping the split line technique, I made a series of separated split-lines along the in- ferior orbital border of a cat, illustrated in figure 4. I had unconsciously anticipated Dempster, assuming that the splits would be oriented along the margin, and made a series of punctures accordingly. The en- larged photograph of this preparation shows that each of the splits was actually directed obliquely rather than being deter- mined by the alignment of the border of the orbit. A more accurate picture of the split-line situation would have resulted from the present method of continuation of the splits in the direction of least resist- ance.
70 N. C. TAPPEN
Fig. 4 Split-line preparation in zygomatic arch of a cat. Note divergence of adjacent split-lines from orientation of the inferior margin of the orbit.
In spite of the demonstration in the pres- ent study of the inadequacy of explanations of split-line orientations as purely a struc- tural conformity, it appears from our ex- perience that Dempster's ('65) generaliza- tions about split-lines in cylindrical sur- faces, fossae, spines, etc., are usually cor- rect, and in other species than man and in areas of the skeleton other than the skull. Why, then, are exceptions so common and variations so great in primate skulls, and, of immediate concern, skulls of' baboons?
The answer seems to lie in the interac- tion of the many functions of the skull, which have been listed as follows (Tappen, '54).
The mammalian skull serves to house and protect the brain and the organs for the special senses of sight, hearing, taste and smell; to provide inlets for food, water and air, plus passageways for blood vessels and nerves; to provide f o r biting and chewing food, and at the same time to give attachments for the chew- ing muscles and a bony framework to resist the forces that are set up by the masticatory apparatus; and to give at- tachment for the muscles which move the head and maintain its carriage. In order to survive the animal must be able to have all these functions operating in
harmony. It is probable that compro- mises between them are in places nec- cssary. It is certain that they can affect each other in some degree.
A inore comprehensive picture of the variations in split-line patterns in baboon skulls, going beyond the necessarily frag- mented descriptions presented so far, ap- pears to center 011 the relationships of the muzzle, the orbits and the masseter muscle. The most probable explanation for the con- tinuation of split-lines in the body of the zygomatic bone into the zygomatic arch is that the tensile force from the action of the masseter muscle tends to orient the fibrous structure of the bone. Presumably the effect is greater in number 24 and on the right side in number 23, so that the split-lines continue to the region of the anterior ten- dinous origin of the muscle at the base of the arch rather than curving smoothly to conform to its border as in the other speci- mens. This also results in cutting off the most lateral split-lines in the zygomatic- alveolar crest. The recurved continuity of the zygomatic area split-lines with those of the maxillary ridge indicates that two sys- tems of lines influence each other in this transitional area but one does not override

BABOON SKULL SPLIT-LINES 71
the other. In number 24 this area of transi- tion is so extensive that i t appears to over- flow into the fronto-sphenoidal process of the zygomatic bone, largely cutting off the continuity of the split-lines otherwise con- forming to the outlines of the orbital mar- gins.
The extent to which the action of the inasseter muscle may affect the split-line patterns encircling the lower orbital mar- gins, a5 in number 14 and “No Number,” or meeting the orbital margin perpendicu- larly, a5 in numbers 11 and 23, is uncer- tain. The perpendicular lines are a contia- uation of a general pattern associated with the muzzle. I t seems plausible that this is an area in which the action of mechanical forces and the resistance and histological responses offered by the bone are in rela- tively delicate balance. allowing consider- able variability in split-line pattern from individual to individual. This variabilty could result from small differences in pro- portions of the skull, size of the dentition, strength of the chewing musculature, and even from different feeding or chewing habits.
Partial support for this interpretation is provided by the zygomatic and circumorbi- tal regions of a series of infant, juvenile and adult gorillas (Tappen, ’57, ’ 6 3 ) . Here the relative size of the split-line patterns primarily associated with the origin of the masseter muscle increases greatly with age, at the expense of systems of lines parallel- ing the orbit in the infraorbital and lateral orbital regions. The structural organization of this area differs considerably from the baboons, of course, but the evidence of age changes in pattern in one species does indi- cate a functional basis for individual differ- ences. In any case, this internretation is flexible enough to encompass the variabil- ity in split-line patterns, while explana-
tions of the patterns as conformity to struc- tural features alone, or simply delineations of the direction of bone growth, are too rigid to be plausible primary factors in the orientation of split-lines on the basis of the actual findings of comparative investiga- tions.
LITERATURE CITED Benninghoff, A. 1925 Spaltlinien am Knochen,
eine Methode zur Ermilllung der Architektur platter Knochen. Verhandl. Anat. Gesellsch., 34: 189-206.
Bjerk, A. 1968 The use of metallic implants in the study of facial growth i n children: Method and application. Am. J. Phys. Anthrop., 29: 243- 254.
Dempster, W. T. 1965 The grain of cortical bone i n relation to structural features of the adult skull. Anat. Rec.? 151: 342-343.
Evans, F. G . 1965 A commentary on the sig- nificance of stresscoat and split-line patterns 0x1 bone. Am. J. Phys. Anthrop., 23: 189-195.
Evans, F. G., and C. W. Goff 1957 A compara- tive study of the primate femur by means of stresscoat and the split-line technique. Am. J. Phys. Anthrop., 15: 59-90.
McGill, H. C., J. P. Strong, R. L. Holman and N. T. Werthessen 1960 Arterial lesions in the Kenya baboon. Circ. Res., 8: 670-679.
Tappen, N. C . 1953 A functional analysis of the facial skeleton with split-line technique. Am. J. Phys, Anthrop., 11 : 503-532.
A comparative functional analysis of primate skulls by the split-line technique. Hum. Biol., 26: 220-238.
.. __- 1957 A comparison of split-line patterns in the skulls of a juvenile and an adult male gorilla, Am. J. Phys. Anthrop., 15: 49-58.
1963 Split-line patterns in a series or skulls of infant, juvenile and adult gorillas. Am. J. Phys. Anthrop., 21: 135-151.
An examination of alternative ex- planations of split-line orientation jn compact bone. Am. J . Phys. Anthrop., 22: 423-441.
____ 1967 Some relationships between split- line patterns and underlying structure in pri- mate skeletons. In: Neue Ergebnisse der Prima- tologie Progress in Primatology. D. R. Starck, R. Schneider and H.-J. Kuhn, eds. Gustav Fisch- er Verlag, Stuttgart, pp. 80-89.
3 954
1964