liver-specffic and shared cell membrane antigens
TRANSCRIPT

Immunology 1978 35 289
Liver-specffic and shared cell membrane antigens
STUDIES BY LIGHT- AND ELECTRON MICROSCOPY
URSULA J. BEHREN S & F. PARONETTO Immunopathology Laboratory, VeteransAdministration Hospital, Bronx, N. Y. 10468
Received 13 October 1977; acceptedfor publication 21 December 1977
Summary. Liver-specific and shared saline-insolublecell surface antigens were localized by immuno-fluorescence as well as by light- and electron micro-scopic immunoenzyme techniques. Antisera againstpurified mouse liver cell membranes were surfacemembrane but not organ-specific. Variable quanti-ties of shared antigens were present in endoderm-and mesoderm-derived organs but not in ectodermalnerve tissue. Species crossreactivity was observedfor the rat. Repeated absorption produced liver-specific antisera that reacted with antigenic sitesdistributed along the entire hepatocyte and sinu-soidal cell surfaces. For the precise localization aswell as the detection of low concentrations of bothliver-specific and nonspecific antigens, the ultra-structural visualization of reactive sites provedessential.
INTRODUCTION
The functional diversity of different cell types isreflected in differences in chemical composition,charge density, and the distribution of distinctmetabolic sites at the cell surface. The dynamics and
Correspondence: Dr Ursula J. Behrens, ImmunopathologyLaboratory, Veterans Administration Hospital, Bronx,N.Y. 10468, U.S.A.0019-2805/78/0800-0289$02.00© 1978 Blackwell Scientific Publications
complex architecture of cell membranes have beenamply reviewed (Winzler, 1970; Hendler, 1971;Kraemer, 1971; Siekevitz, 1972). Immunologicalstudies indicate that antigenic cell surface de-terminants of unrelated cells exhibit a high degree ofoverlap; however, certain cell membrane antigensare organ- or tissue-specific (Moscona, 1968;Hanna & Nelken, 1969; Sell, Mori, Rack, Gurner &Coombs, 1969; Goldschneider & Moscona, 1971,1972).The location of cell- and organ-specific as well as
shared antigens is of importance in the elucidationof immunological disorders in neoplasia and auto-immune reactions.
In chronic hepatitis, the alteration of cell mem-branes has been implicated to play a role in theensuing tissue damaging immunological reaction(Meyer zum Buschenfelde & Hopf, 1974). Little isknown, however, of the precise location of liver-specific surface antigens. In this study, the distri-bution of saline-insoluble specific and shared surfaceantigens of mouse liver cells was investigated byindirect immunofluorescence and immunoenzymelight- and electron microscopy.
MATERIALS AND METHODS
Isolation ofplasma membranesFemale AKR mice (National Cancer Institute, Silver
289

Ursula J. Behrens & F. Paronetto
Spring, Maryland) were bled from axillary vesselsunder ether anaesthesia. Livers were removed andwashed free of residual blood with chilled 0-85%NaCl. The tissues were minced in two volumes ofhypotonic buffer (1 mm NaHCO3: 0 5 mm CaCl2,pH 7 5) and then homogenized in a Dounce homo-genizer (pestle A) by 6-8 gentle downstrokes. Cellmembranes were isolated under ice-cold conditionsby a method similar to that described by Ray (1970).The homogenate was diluted 100-fold the initialtissue volume with hypotonic buffer and left stand-ing for 10 min with occasional stirring. Cell nucleiwere removed by brief centrifugation (30 sec at1300 g. The supernatant was centrifuged at 3400 gfor 45 min. The resulting pellet was resuspended in100 volumes of hypotonic buffer and centrifuged at3400 g for 15 min. The last step was repeated. Thepellet was resuspended in a small volume of hypo-tonic buffer and diluted with 60% sucrose (in 5 mmTris: 5 mm CaCl2, pH 7 4) to a final concentrationof 48% sucrose. The cell membrane fraction (CM)was isolated by flotation through a sucrose stepgradient (Behrens & Hollander, 1976). To removeresidual contaminants from the CM layer, found atthe 41 %/37% sucrose interface, the flotation stepwas repeated. The CM layer was washed and centri-fuged twice in 150 ml of saline. Portions of thisfraction were assayed for marker enzymes, and fixedin osmic acid (1% in 0-1 M sodium cacodylate,pH 7 3) for electron microscopic examination.
Chemical assaysProtein was determined by the method of Lowry,Rosebrough, Farr & Randall (1951). Succinicdehydrogenase activity was assayed by the method ofReid (1960) and glucose-6-phosphatase, Mg++-dependent ATPase, and 5'-nucleotidase activitieswere determined as described by Ray (1970).Inorganic phosphate released by the phosphataseswas measured by the method of Chen, Toribara &Warner (1956).
Immunization ofrabbitsOne mg of mouse liver CM protein, suspended in0 5 ml saline and emulsified in an equal volume ofcomplete Freund's adjuvant (GIBCO, Grand Island,New York) was injected into two albino rabbits(Camm Research, Wayne, New Jersey) at four (twointramuscular, two subcutaneous) sites. Seven daysafter the third weekly injection, the animals werebled from the ear or by cardiac puncture. A third
rabbit received six weekly injections of 3 mg of liverCM protein. All animals were bled before im-munization to obtain normal control sera.
ImmunofluorescenceAcetone-fixed cryostat sections from organs ob-tained from AKR mice were reacted for 30 min withappropriate dilutions (in phosphate-buffered saline(PBS)) of antisera, followed by goat-anti-rabbit Ig(IgG, IgA, IgM), fluorescein conjugated (BehringDiagnostics, Somerville, New Jersey) (30 min).Between steps the slides were washed three times(for 5 min each) with PBS and finally mounted inelvanol (Rodrigues & Deinhardt, 1960). To excludenonspecific immunofluorescence, normal rabbitserum was substituted for antiserum.
Absorption ofantiseraAntisera were diluted either 1: 5, 1: 10, or 1: 20 inPBS and mixed with 100-300 mg/ml serum (un-diluted) or lyophilized homogenates of kidney,intestine, spleen, heart, lung, and thymus. Absorp-tions were carried out (2 or 3 times) at 370 for30 min and overnight at 4°. The resulting liver-specific antiserum was used as negative control forall tissues other than liver. Antiserum absorbed withliver homogenate was used as negative control forliver tissue.
Light microscopic immunoenzyme techniqueAcetone-fixed cryostat sections of AKR mousetissues were washed with Tris-HCI, pH 7-6, and pro-cessed by the unlabelled antibody enzyme method(Sternberger, 1973). All antisera were diluted withTris-HCI, containing 0 1% gelatin. Briefly, the slideswere treated in sequence with normal sheep serum(diluted 1: 30 with saline), appropriate dilutions(unabsorbed antiserum 1: 60-1: 1280; liver specificantiserum 1: 10-1: 40) of anti cell membraneserum, sheep anti rabbit serum (Antibodies Inc.,Davis, California) (diluted 1: 10), peroxidase:antiperoxidase complex (PAP) (Accurate Chemical& Scientific Corp., Hicksville, New York) (diluted1: 10) for 10 min each. Between steps, the slides wererinsed with Tris-HCl, pH 7 -6, from a pipette andleft standing in Tris buffer for 5 min. After reactionwith PAP, the slides were treated with 3,3'-diamino-benzidine (DAB) (Polysciences Inc., Warrington,Pennsylvania) as described by Graham & Karnovsky(1966), and then exposed to 1% 0S04 (in 0-1 M
290

Liver cell membrane antigens
sodium cacodylate, pH 7 4) for 30 min, washedwith Tris buffer and mounted in elvanol.
Electron microscopic immunoenzyme techniqueMouse tissues, not larger than 0 5 mm in diameter,were fixed in 3 % glutaraldehyde in Millonig's bufferfor 45 min at room temperature. The tissues were
then rinsed and left overnight at 4° in buffer. Thefixed tissue was sliced into smaller pieces with a
sharp razor blade and processed in small test tubesby the procedure described for light microscopy.To facilitate penetration, the time of incubation withantisera was increased to 24 h, allowing 8-10 h forwashing. To further improve penetration, portionsof each tissue were incubated with 0 2% tritonX-100 (Pickel, Joh & Reis, 1976), for 30 min atroom temperature following glutaraldehyde fixa-tion. Two controls were included: (1) to account fornonspecific reactions with PAP, endogenous tissueperoxidase, and excessive osmiophilia, sheep antirabbit serum was omitted; (2) to detect nonspecificantigen-antibody reactions, anti-liver CM sera were
substituted by normal rabbit sera. During the reac-
tion with antisera, the tubes were placed on a rockingplatform. The tissues were postfixed in 1% OS04in Millonig's buffer, pH 7 4, for 30 min at room
temperature, dehydrated in ethanol and embeddedin Epon 812 (Luft, 1961). Thin sections were cuteither on an MT-2 Porter Blum or an LKB ultra-microtome and examined in the electron microscopebefore and after staining with uranyl acetate andlead citrate (Venable & Coggeshall, 1965). Electronmicrographs were taken with a Phillips 300 electronmicroscope at 80 kV.
RESULTS
Evaluation of isolated cell membranesThe average yield of AKR mouse liver CM protein(8 determinations) as compared to the total proteinof initial homogenates was 2-1%±0-17 (S.E.). 5'-nucleotidase and Mg++-dependent ATPase, knownmarker enzymes of liver cell membranes (Ray, 1970),showed significantly increased activities in CM frac-tions as compared to homogenates (Table 1).Activities of glucose-6-phosphatase (a marker of theendoplasmic reticulum) and succinic dehydrogenase(a mitochondrial marker) were negligible in CM
Figure 1. Isolated mouse liver CM fraction. No mitochondriaor elements of rough endoplasmic reticulum are visible.Junctional complexes, found only between cell membranes(arrows) are seen. ( x 21,000)
Table 1. Activities of selected marker enzymes in homogenates and CM fractions of AKR mouseliver
Specific activity*
% Total activityrecovered in CM
Enzymes Homogenate CM fraction fraction
5'-nucleotidase 1-74 ± 0.1 It (6)1 21-08 ±2-8 (6) 23 55 1-82Mg++-ATPase 7-42 ± 1-25 (4) 41-93 ± 355 (4) 7-04 0-77Glucose-6-phosphatase 5-16 ± 1-16 (5) 1-21 ±0-24 (5) 0 54 0-21Succinic dehydrogenase 1-60 ±0 33 (7) 0-32 ±0-08 (7) 0 44 ±010
* All enzyme activities are expressed in micro moles of product produced per mg protein perhour.
t Mean ± S.E.$ Numbers in parentheses = number of determinations.
291

Ursula J. Behrens & F. Paronetto
preparations (less than 1% of total activity ofhomogenates). Electron microscopic examination ofdifferent CM fractions gave no evidence of con-tamination with mitochondria or elements of therough endoplasmic reticulum, but showed smoothmembrane vesicles and junctional complexes foundonly between cell membranes (Fig. 1).
Antisera against liver CMAll rabbits injected with purified CM material pro-duced CM-specific but not organ-specific antibodies.Titres of antisera, designated as the highest dilutionat which fluorescence was clearly visible, ranged from1: 80-1: 2560. Extensive absorption resulted inrelatively low liver-specific titres, ranging from1:5 to 1: 80.
Immunoenzyme light microscopyIndirect immunofluorescence and the light micro-scopic immunoenzyme technique gave identicalstaining patterns, and only results of the lattermethod will be reported here. Before absorption,antisera produced cell surface staining in theendoderm-derived liver, lung, and intestine. Thesame reaction was observed in the mesoderm-derived organs heart, skeletal muscle, spleen,thymus, kidney, and adrenal cortex. Ectodermalnerve tissues, such as brain and adrenal medulla,showed negative results.Examples of antigenic sites shared with liver cell
surfaces are demonstrated in Figs 2-4. The intestine(Fig. 2) shows a heavy reaction along the mucosaand weaker CM staining in cell layers belowthe luminal surface. In skeletal muscle (Fig. 3), thesarcolemma is strongly DAB-positive, and in thespleen (Fig. 4), plasma membranes are clearlyoutlined.
After absorption, antisera reacted only with liversections. As compared to the honeycomb patternproduced by nonspecific anti-CM serum (Fig. 5),liver-specific antiserum gave a much weaker reaction(Fig. 6), producing DAB-positive deposits thatappear to line sinusoidal walls and bile canaliculi.The type of cell and the precise location of thismaterial cannot be discerned by light microscopy.
Sections of rat organs were tested for speciescrossreactivity by immunofluorescence only. Stain-ing patterns similar to those described in mice wereobserved in this species.
Immunoenzyme electron microscopyProlonged incubation with antisera facilitatedadequate penetration in kidney, heart, skeletalmuscle, intestine, and lung, whereas in liver, spleenand thymus, only peripheral cells showed reactionproduct. Exposure to triton X-100 after glutaralde-hyde fixation resulted in adequate antiserumpenetration of the latter tissues to demonstrateantigenic sites. Alterations in location of reactiveantigens or effects on ultrastructure due to treat-ment with the detergent, with the exception of someseparation of cells, were not observed.
Liver-specific antisera produced intense stainingof granular material along the surface of sinusoidalendothelium and a weaker reaction on the irregularhepatocyte CM within the space of Disse (Fig. 7).In bile canaliculi and along the entire length ofadjacent surface membranes, DAB-positive materialwas seen as a thin continuous line (Fig. 8). By lightmicroscopy liver-specific staining of the hepatocyteCM appeared to be limited to secretory sites, pre-sumably due to low concentrations of antigens onthe less active CM surfaces. Kupffer cells (Fig. 9)and lipid-storing cells (Fig. 10), found in hepaticsinusoids, showed granules of dense reaction pro-duct on their surfaces.
In liver control sections treated with antiserumthat had been absorbed with liver homogenate, noantigen-antibody complexes could be visualized.Comparative results obtained with anti-CM serumbefore and after absorption are shown in Figs 11and 12, respectively.As observed by light microscopy, unabsorbed
anti-CM sera gave a CM-specific reaction in allorgans tested, with the exception of nerve tissue.Organs of mesodermal origin, such as spleen,thymus, heart, and kidney, showed distinct cellsurface staining. In skeletal muscle, the sarcolemmashowed a relatively heavy coating (Fig. 13), whereasno reaction was observed when liver-specific anti-serum was used (Fig. 14). Similar staining patternswere produced in endoderm-derived cell systems. Inthe lung, dense material covered the pneumocyteCM (Fig. 15), and in the intestine, the large surfacearea of the luminal brush border was coated withreaction product. Absorbed (liver-specific) antiserumgave negative results in both lung (Fig. 17) andintestine (Fig. 18). Similar observations were madein the kidney, where shared antigens were presenton the CM of glomerular podocytes, vascularendothelium, and tubular brush border. Antigenic
292

Liver cell membrane antigens
'6 :A..s' 8 "." -hFigures 2-5. Acetone-fixed mouse tissue sections treated with unabsorbed anti-mouse liver CM serum. Light micrographs.(x 500)Figure 2. Intestinal mucosa coated with dense reaction product. A weakly positive reaction is seen on cell surfaces below thelumen. Figure 3. Skeletal muscle. Only the sarcolemma is DAB-positive. Figure 4. Spleen. Cell surfaces show dense rims ofreaction product. Figure 5. Liver showing a honeycomb staining pattern.
Figure 6. Liver reacted with liver-specific anti-CM serum. Staining appears to be present in sinusoids and bile canaliculi. ( x 500).
293

Ursula J. Behrens & F. Paronetto
~ ~~--. .. 3 i|a
acttn lea ctrt wit h ecpiono igue 7 an ntcutrtie)
suracwihi th spc of Dis ()i cvrd ihthnyiprsdsaie atra. (x 2800. Fgr8.Bi l caaiuu (B)
o~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~Ht,
S~~b ~
andadacn heaoct CM so cotnossraetiig.(280). Fiue9 uie el()wti iuod(i
P ' ~O
showsfakadlaB-poitivte deoit sth xetonits firrgulresurae7 an 18,000). Figunrse1.Lpd-() soigcl ihnsiuod(ihwn
the same reaction pattern as the Kupifer cell. ( x 18,000).
294

Liver cell membrane antigens
13 A
Figure 11. Liver reacted with unabsorbed anti-CM serum. CM and surface of bile canaliculus (B) are covered with dense reactionproduct. ( x 28,000).Figure 12. Liver reacted with anti-CM serum absorbed with liver. Cell surfaces within space of Disse (D) and of endothelium(E) show no reaction product. ( x 18,000).Figure 13. Skeletal muscle reacted with unabsorbed anti-CM serum. Sarcolemma (S) shows a heavy coat of DAB-positivematerial. ( x 12,000).Figure 14. Skeletal muscle reacted with liver-specific anti-CM serum. Sarcolemma (S) is free of reaction product. ( x 12,000).
295
Ursula J. Behrens & F. Paronetto
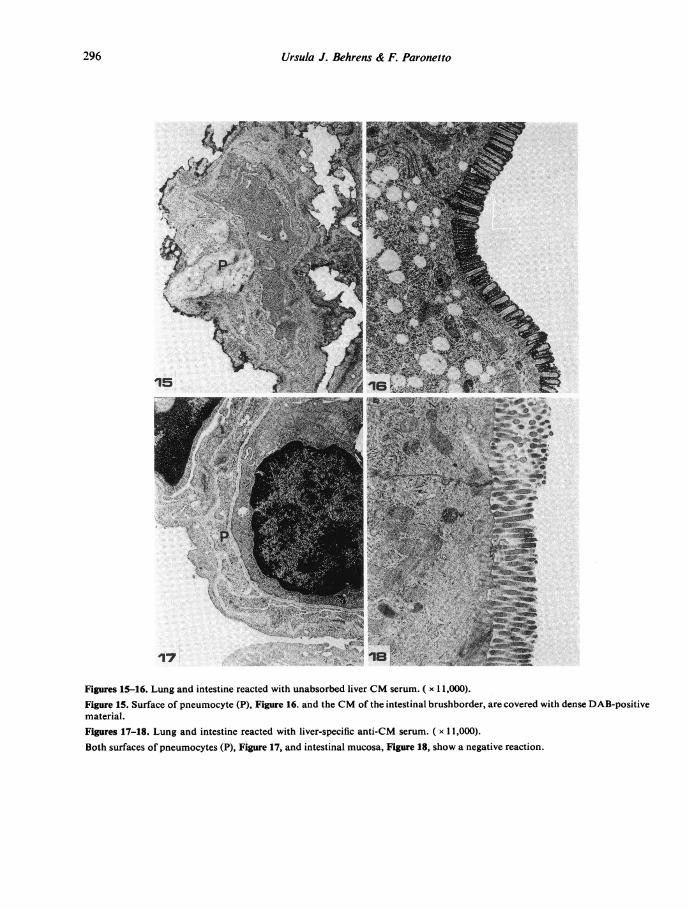
Figures 15-16. Lung and intestine reacted with unabsorbed liver CM serum. ( x 11,000).Figure 15. Surface of pneumocyte (P), Figure 16. and the CM of the intestinal brushborder, are covered with dense DAB-positivematerial.Figures 17-18. Lung and intestine reacted with liver-specific anti-CM serum. ( x 11,000).Both surfaces of pneumocytes (P), Figure 17, and intestinal mucosa, Figure 18, show a negative reaction.
296

Liver cell membrane antigens
Table 2. Quantitative differences of common surface antigens in different organs
Reciprocal titres* of anti-CM serum for:
Liver Intestine Kidney Spleen Thymus Heart Skel. muscle
2560 1280 640 1280 2560 640 640
* Titres are designated as the highest dilution at which CM fluorescence was clearly visible.
sites within the tubular lumen could not be resolvedby light microscopy.
Quantitative differences in shared surface antigensAnti-liver CM serum of the highest titre was usedfor the determination of comparative titres of anti-bodies against shared antigens. Differences in anti-serum dilutions at which fluorescence was detectablein different organs (Table 2) indicate that antigenscommon to functionally different cells vary inquantity.
DISCUSSION
Antisera against isolated mouse liver cell membranesproduced cell surface-specific staining by immuno-fluorescence and the unlabelled antibody enzymetechnique, indicating that the liver CM fractionused for immunization was of high purity. This wasfurther confirmed by marker enzyme activities andmorphological observations.Unabsorbed antiserum against mouse liver CM
lacked organ and species specificity. Variablequantities of shared surface antigens were found incell systems of endodermal and mesodermal origin.Ectoderm-derived nerve cells, mainly concernedwith the transmission of electrical impulses, however,did not react with anti-liver-CM serum. Speciescrossreactivity was observed by immunofluorescencein the rat.Repeated absorption produced organ-specific
anti-CM serum, although of comparatively lowtitre. Our results indicate that liver-specific saline-insoluble surface antigens represent only a smallpercentage of the total number of antigens. Similarfindings were reported by other authors (Sell et al.,1969) for human CM antigens in different organs,including liver.
Results obtained by both indirect immunofluores-
cence and the light microscopic unlabelled antibodyenzyme technique indicate that liver-specific CMantigens are concentrated in sinusoids and bilecanaliculi. Similar observations made by immuno-fluorescence only, in mouse, rat, rabbit, and humanliver, have been reported in the literature (Sulit-zeanu & Slavin, 1968; Sheffield & Emmelot, 1972;Hopf, Meyer zum Buschenfelde & Freudenberg,1974; Kramkova & Beloshapkina, 1974). However,different methods for the isolation of liver CM wereused, and it is difficult to determine, whether one isdealing with identical surface antigens. No com-parative immunoenzyme light and electron micro-scopic studies have been reported.The ultrastructure of liver, treated with organ-
specific antiserum, revealed that all liver cell types,including hepatocytes, Kupffer cells, lipid-storingcells, and sinusoidal endothelium, have liver-specific antigens on their surfaces. Since antiserawere produced against a CM preparation of a hetero-geneous cell population, these results are not sur-prising. Presumably there are more than oneantigen involved that are not necessarily common toall liver cell types. These antigenic sites are distri-buted along the entire surface of hepatocytes andsinusoidal cells and appear to be most concentratedon the endothelial surface and in bile canaliculi. Forthe immunoenzyme technique used in this study, theamount of reaction product is not only a function ofantigen concentration but also of antiserum dilution(Bigbee, Kosek & Eng, 1977) and does not neces-sarily correlate with antigen density. However,since fluorescent staining which corresponds directlyto antigen concentration, was intense in sinusoidsand bile canaliculi, it is conceivable that these highlyspecialized structures are coated with a large numberof specific antigens. By light microscopy, low con-centrations of CM antigens located on nonvilloussurfaces of hepatocytes as well as the precise locationof antigenic sites in sinusoids could not be resolved.The nature of liver-specific antigens demonstrated
297

298 Ursula J. Behrens & F. Paronetto
here, and the significance of specific and sharedantigens in autoimmune hepatitis remains to bedetermined.
ACKNOWLEDGMENTS
This study was supported by Veterans Administra-tion Research Funds.The preparation of tissue sections and assistance
with immunofluorescence and immunoenzymetechniques by Mr Joseph Fiorillo is gratefullyacknowledged.The authors also wish to thank Mr John
Bodnaruk, Department of Chemical Engineering,City College, C.U.N.Y., for his co-operation in theuse of the electron microscope and darkroomfacilities.
REFERENCES
BEHRENS U.J. & HOLLANDER V.P. (1976) Cell membranesialoglycopeptides of corticoid-sensitive and -resistantlymphosarcoma P1798. Cancer Res. 36, 172.
BIGBEE J.W., KOSEK J.C. & ENG L.F. (1977) Effects ofprimary antiserum dilution on staining of 'antigen-rich'tissues with the peroxidase antiperoxidase technique.J. Histochem. Cytochem. 25, 443.
CHEN P.S., JR, TORIBARA T.Y. & WARNER H. (1956) Micro-determination of phosphorus. Anal. Chem. 28, 1756.
EMMELOT P. & BENEDETTI E.L. (1967) On the possible involve-ment of the plasma membrane in the carcinogenicprocess. In: Carcinogenesis: A broad critique, M. D.Anderson Hospital and Tumor Inst. Symp., Universityof Texas. Williams and Wilkins, Baltimore, Md.
GOLDSCHNEIDER I. & MOSCONA S.A. (1971) Tissue-specificantigenic determinants on embryonic cell surfacesdemonstrated by antisera prepared against suspensionsof liver cells. Anat. Rec. 169, 478.
GOLDSCHNEIDER I. & MOSCONA A.A. (1972) Tissue-specificcell surface antigens in embryonic cells. J. cell Biol. 53,435.
GRAHAM R.C. & KARNOVSKY J.J. (1966) The early stages ofabsorption of injected horseradish peroxidase in theproximal tubules of mouse kidney; ultrastructural cyto-chemistry by a new technique. J. Histochem. Cytochem.14, 291.
HANNA N. & NELKEN D. (1969) Thrombocyte-specificantigens. Detection of organ-specific antigens by hetero-specific antithrombocyte serum. Immunology, 16, 601.
HENDLER R.W. (1971) Biological membrane ultrastructure.Physiol. Rev. 51, 66.
HOPF U., MEYER ZUM BUSCHENFELDE K.H. & FREUDENBERGJ. (1974) Liver-specific antigens of different species. II.Localization of a membrane antigen at cell surfaces ofisolated hepatocytes. Clin. exp. Immunol. 16, 117.
KRAMKOVA N.I. & BELOSHAPKINA T.D. (1974) Antigen ofmouse bile capillaries and cuticle of intestinal mucosa.Nature (Lond.), 251, 627.
KRAEMER P.M. (1971) Complex carbohydrates of animalcells: Biochemistry and physiology of the cell periphery.In: Biomembranes, 1, 67 (ed. by Manson). Plenum Press.
LOWRY O.H., ROSEBROUGH N.J., FARR A.L. & RANDALLR.J. (1951) Protein measurement with the Folin phenolreagent. J. biol. Chem. 193, 265.
LUFT J.H. (1961) Improvements in epoxy embeddingmethods. J. biophys. biochem. Cytol. 9, 409.
MEYER ZUM BOSCHENFELDE K.H. & HOPF U. (1974) Studieson the pathogenesis of experimental chronic hepatitis.Brit. J. exp. Path. 55, 498.
MOSCONA A.A. (1968) Cell aggregation: properties of specificcell ligands and their role in the formation of multi-cellular systems. Develop. Biol. 18, 250.
PICKEL V.M., JOH T.H. & REIs D.J. (1976) Monaminesynthesizing enzymes in central dopaminergic, noradre-nergic and serotonergic neurons. Immunocytochemicallocalization by light and electron microscopy. J. Histo-chem. Cytochem. 24, 792.
RAY T.K. (1970) A modified method for the isolation of theplasma membranes from rat liver. Biochim. biophys. Acta,196, 1.
REID E. (1960) Assay of marker enzymes. In: SubcellularComponents-Preparation and Fractionation (ed. by G. D.Birnie). University Park Press, Baltimore.
RODRIGuEs L. & DEINHARDT F. (1960) Preparation of asemipermanent mounting medium for fluorescent anti-body studies. Virology, 12, 316.
SELL K.W., MORI W., RACK J.H., GURNER B.W. & COoMBsR.R.A. (1969) Organ-specific membrane antigens.Attempts to produce specific antisera for mixed anti-globulin tests on disaggregated cells. Brit. J. exp. Path.50, 413.
SHEFFIELD J.B. & EMMELOT P. (1972) Studies on plasmamembranes. XVI. Tissue-specific antigens in the livercell membrane. Exp. Cell Res. 71, 97.
SIEKEVITZ P. (1972) Biological membranes: The dynamics oftheir organization. Ann. Rev. Biochem. 240, 117.
STERNBERGER L.A. (1973) Enzyme immunocytochemistry. In:Electronmicroscopy of enzymes-principles and methods(ed. by M. S. Hayat). Van Nostrand Reinhold Company,New York.
SULITZEANU D. & SLAVIN M. (1968) Partial purification ofan antigen lining the rat bile canaliculi. Clin. exp. Im-munol. 3, 1005.
VENABLE J.H. & COGGESHALL R.A. (1965) A simplified leadcitrate stain for use in electron microscopy. J. cell Biol.25, 407.
WINZLER R.J. (1970) Carbohydrates in cell surfaces. Int. Rev.Cytol. 29, 77.